ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress


 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
- 1st = 25 days after sowing (Crown root Initiation)
- 2nd = 55 days after sowing (Tillering stage)
- 3rd = 80 days after sowing (Heading stage)
- 4th = 110 days after sowing (Milky stage/soft dough)
3. Results
3.1. Plant Height, Root Length and Spike Length
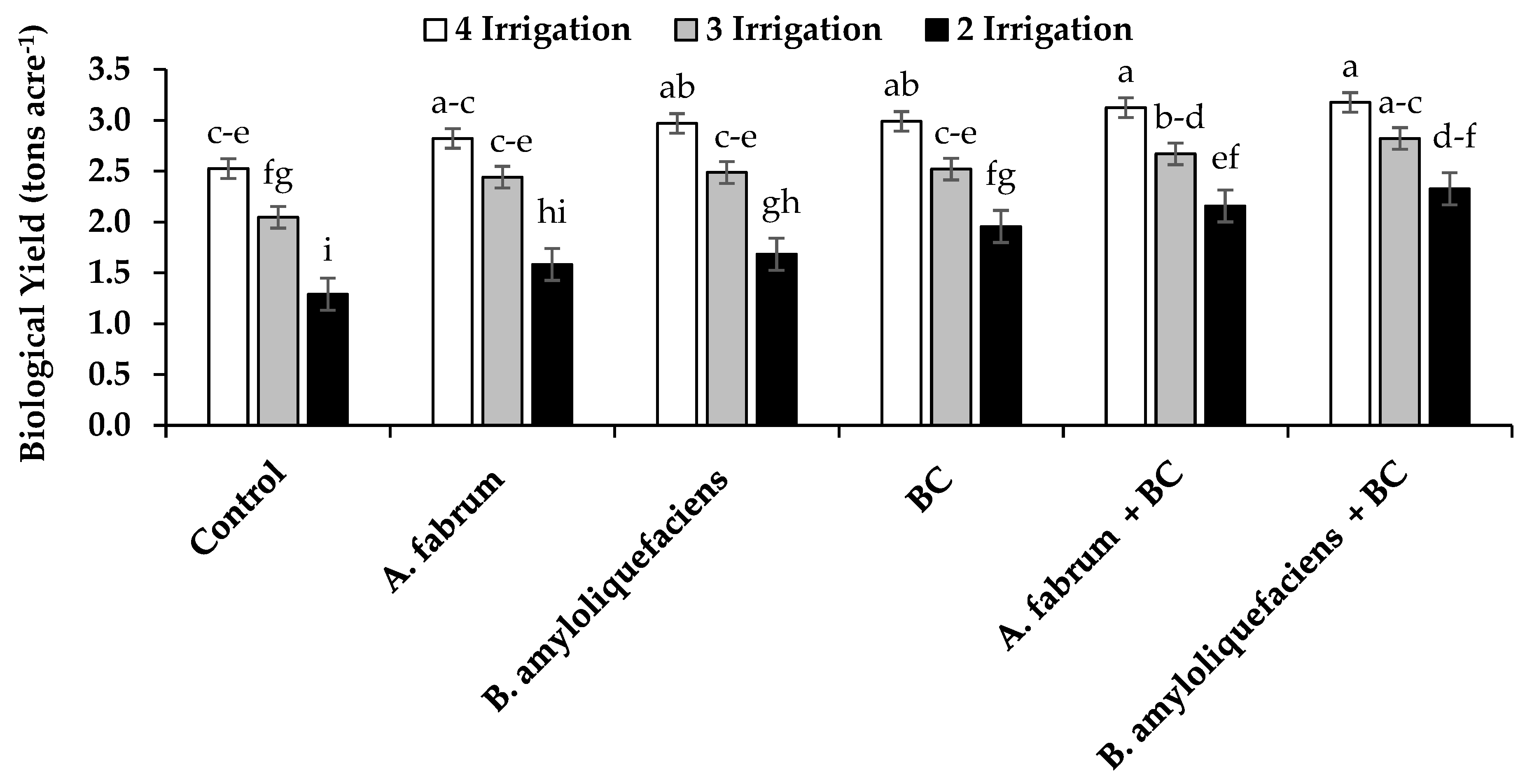
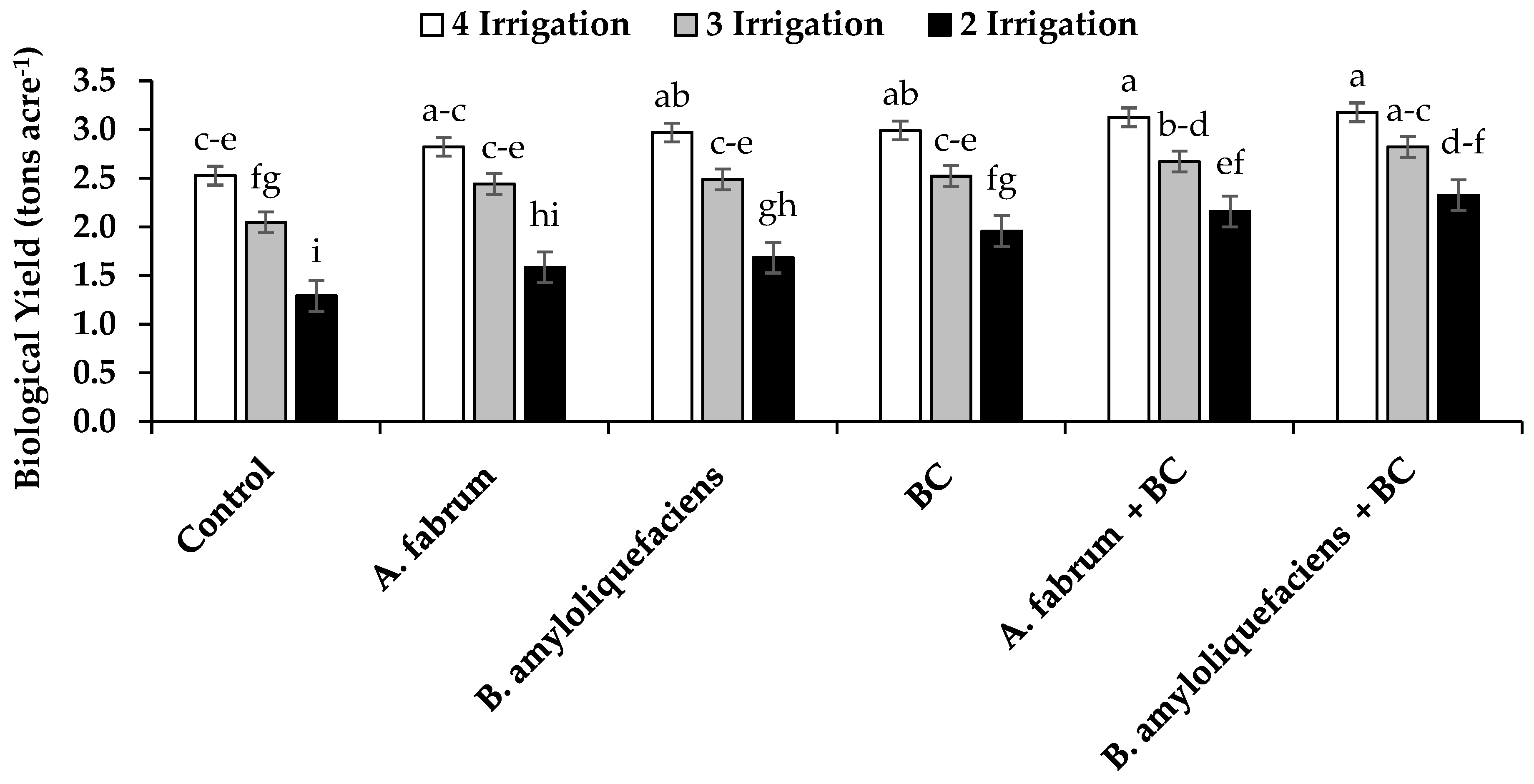
3.2. Grain, Straw and Biological Yield
3.3. Spikelets Spike−1, Grains Spik−1 and 1000 Grain Weight
3.4. N, P and K Concentration in Grains
3.5. N, P and K Concentration in Shoot
3.6. Gas Exchange Attributes
3.7. Chlorophyll Content
3.8. Electrolyte Leakage
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Bos, C.; Juillet, B.; Fouillet, H.; Turlan, L.; Daré, S.; Luengo, C.; N’tounda, R.; Benamouzig, R.; Gausserès, N.; Tomé, D.; et al. Postprandial metabolic utilization of wheat protein in humans. Am. J. Clin. Nutr. 2005, 81, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef]
- Ahmad, Z.; Waraich, E.A.; Akhtar, S.; Anjum, S.; Ahmad, T.; Mahboob, W.; Hafeez, O.B.A.; Tapera, T.; Labuschagne, M.; Rizwan, M. Physiological responses of wheat to drought stress and its mitigation approaches. Acta Physiol. Plant. 2018, 40, 80. [Google Scholar] [CrossRef]
- Kilic, H.; Yağbasanlar, T. The effect of drought stress on grain yield, yield components and some quality traits of durum wheat (Triticum turgidum ssp. Durum) cultivars. Not. Bot. Horti Agrobot. Clujnapoca 2010, 38, 164–170. [Google Scholar]
- Waraich, E.A.; Ahmad, R.; Ashraf, M. Role of mineral nutrition in alleviation of drought stress in plants. Aust. J. Crop Sci. 2011, 5, 764–777. [Google Scholar]
- Griffin, M.T.; Montz, B.E.; Arrigo, J.S. Evaluating climate change induced water stress: A case study of the lower cape fear basin, nc. Appl. Geogr. 2013, 40, 115–128. [Google Scholar] [CrossRef]
- Munir, T.; Perkins, M.; Kaing, E.; Strack, M. Carbon dioxide flux and net primary production of a boreal treed bog: Responses to warming and water-table-lowering simulations of climate change. Biogeosciences 2015, 12, 1091–1111. [Google Scholar] [CrossRef]
- Bechtold, M.; De Lannoy, G.; Koster, R.; Reichle, R.; Mahanama, S.; Bleuten, W.; Bourgault, M.; Brümmer, C.; Burdun, I.; Desai, A.; et al. Peat-clsm: A specific treatment of peatland hydrology in the nasa catchment land surface model. J. Adv. Model. Earth Syst. 2019. [Google Scholar] [CrossRef]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with acc deaminase producing rhizobacteria isolated from acidic soil of northeast india. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef]
- Zhang, S.; Kang, H.; Yang, W. Climate change-induced water stress suppresses the regeneration of the critically endangered forest tree nyssa yunnanensis. PLoS ONE 2017, 12, e0182012. [Google Scholar] [CrossRef]
- Strack, M.; Munir, T.M.; Khadka, B. Shrub abundance contributes to shifts in dissolved organic carbon concentration and chemistry in a continental bog exposed to drainage and warming. Ecohydrology 2019. [Google Scholar] [CrossRef]
- Preston, K.L.; Rotenberry, J.T.; Redak, R.A.; Allen, M.F. Habitat shifts of endangered species under altered climate conditions: Importance of biotic interactions. Glob. Chang. Biol. 2008, 14, 2501–2515. [Google Scholar] [CrossRef]
- Allen, M.R.; Barros, V.R.; Broome, J.; Cramer, W.; Christ, R.; Church, J.A.; Clarke, L.; Dahe, Q.; Dasgupta, P.; Dubash, N.K. IPCC Fifth Assessment Synthesis Report-Climate Change 2014 Synthesis Report. 2014. Available online: https://ar5-syr.ipcc.ch/ (accessed on 28 June 2019).
- Mishra, V.; Cherkauer, K.A. Retrospective droughts in the crop growing season: Implications to corn and soybean yield in the midwestern united states. Agric. Meteorol. 2010, 150, 1030–1045. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.K.; Liu, S.; Rudd, J.C.; Xue, Q.; Payton, P.; Finlayson, S.A.; Mahan, J.; Akhunova, A.; Holalu, S.V.; Lu, N. Physiology and transcriptomics of water-deficit stress responses in wheat cultivars tam 111 and tam 112. J. Plant Physiol. 2014, 171, 1289–1298. [Google Scholar] [CrossRef] [PubMed]
- Munir, T.M.; Khadka, B.; Xu, B.; Strack, M. Mineral nitrogen and phosphorus pools affected by water table lowering and warming in a boreal forested peatland. Ecohydrology 2017, 10, e1893. [Google Scholar] [CrossRef]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zafar-Ul-Hye, M.; Shahjahan, A.; Danish, S.; Abid, M.; Qayyum, M.F. Mitigation of cadmium toxicity induced stress in wheat by acc-deaminase containing pgpr isolated from cadmium polluted wheat rhizosphere. Pak. J. Bot. 2018, 50, 1727–1734. [Google Scholar]
- Sharp, R.E.; LeNoble, M.E. Aba, ethylene and the control of shoot and root growth under water stress. J. Exp. Bot. 2002, 53, 33–37. [Google Scholar] [CrossRef]
- Danish, S.; Zafar-ul-Hye, M.; Hussain, M.; Shaaban, M.; Núñez-Delgado, A.; Hussain, S.; Qayyum, M.F. Rhizobacteria with ACC-deaminase activity improve nutrient uptake, chlorophyll contents and early seedling growth of wheat under PEG-induced osmotic stress. Intl. J. Agric. Biol. 2019, 21, 1212–1220. [Google Scholar]
- Danish, S.; Zafar-ul-Hye, M. Co-application of acc-deaminase producing pgpr and timber-waste biochar improves pigments formation, growth and yield of wheat under drought stress. Sci. Rep. 2019, 9, 5999. [Google Scholar] [CrossRef] [PubMed]
- Stromberger, M.E.; Abduelafez, I.; Byrne, P.; Elamari, A.A.; Manter, D.K.; Weir, T. Genotype-specific enrichment of 1-aminocyclopropane-1-carboxylic acid deaminase-positive bacteria in winter wheat rhizospheres. Soil Sci. Soc. Am. J. 2017, 81, 796–805. [Google Scholar] [CrossRef]
- Salem, G.; Stromberger, M.E.; Byrne, P.F.; Manter, D.K.; El-Feki, W.; Weir, T.L. Genotype-specific response of winter wheat (Triticum aestivum L.) to irrigation and inoculation with acc deaminase bacteria. Rhizosphere 2018, 8, 1–7. [Google Scholar] [CrossRef]
- Kumputa, S.; Vityakon, P.; Saenjan, P.; Lawongsa, P. Carbonaceous greenhouse gases and microbial abundance in paddy soil under combined biochar and rice straw amendment. Agronomy 2019, 9, 228. [Google Scholar] [CrossRef]
- Basak, B.B.; Biswas, D.R. Co-inoculation of potassium solubilizing and nitrogen fixing bacteria on solubilization of waste mica and their effect on growth promotion and nutrient acquisition by a forage crop. Biol. Fertil. Soils 2010, 46, 641–648. [Google Scholar] [CrossRef]
- Mwajita, M.R.; Murage, H.; Tani, A.; Kahangi, E.M. Evaluation of rhizosphere, rhizoplane and phyllosphere bacteria and fungi isolated from rice in kenya for plant growth promoters. SpringerPlus 2013, 2, 606. [Google Scholar] [CrossRef]
- Belimov, A.A.; Safronova, V.I.; Sergeyeva, T.A.; Egorova, T.N.; Matveyeva, V.A.; Tsyganov, V.E.; Borisov, A.Y.; Tikhonovich, I.A.; Kluge, C.; Preisfeld, A. Characterization of plant growth promoting rhizobacteria isolated from polluted soils and containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2001, 47, 642–652. [Google Scholar] [CrossRef]
- Mayak, S.; Tirosh, T.; Glick, B.R. Plant growth-promoting bacteria confer resistance in tomato plants to salt stress. Plant Physiol. Biochem. 2004, 42, 565–572. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Munir, A.; Asghar, H.N.; Shaharoona, B.; Arshad, M. Effectiveness of rhizobacteria containing acc deaminase for growth promotion of peas (Pisum sativum) under drought conditions. J. Microbiol. Biotechnol. 2008, 18, 958–963. [Google Scholar]
- Gundale, M.J.; DeLuca, T.H. Charcoal effects on soil solution chemistry and growth of koeleria macrantha in the ponderosa pine/douglas-fir ecosystem. Biol. Fertil. Soils 2006, 43, 303–311. [Google Scholar] [CrossRef]
- Hartmann, M.; Fliessbach, A.; Oberholzer, H.R.; Widmer, F. Ranking the magnitude of crop and farming system effects on soil microbial biomass and genetic structure of bacterial communities. FEMS Microbiol. Ecol. 2006, 57, 378–388. [Google Scholar] [CrossRef] [PubMed]
- De Jesus Duarte, S.; Glaser, B.; Pellegrino Cerri, C.E. Effect of biochar particle size on physical, hydrological and chemical properties of loamy and sandy tropical soils. Agronomy 2019, 9, 165. [Google Scholar] [CrossRef]
- Wacal, C.; Ogata, N.; Basalirwa, D.; Handa, T.; Sasagawa, D.; Acidri, R.; Ishigaki, T.; Kato, M.; Masunaga, T.; Yamamoto, S.; et al. Growth, seed yield, mineral nutrients and soil properties of sesame (Sesamum indicum L.) as influenced by biochar addition on upland field converted from paddy. Agronomy 2019, 9, 55. [Google Scholar] [CrossRef]
- De la Rosa, J.M.; Rosado, M.; Paneque, M.; Miller, A.Z.; Knicker, H. Effects of aging under field conditions on biochar structure and composition: Implications for biochar stability in soils. Sci. Total Environ. 2018, 613, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Paneque, M.; De la Rosa, J.M.; Kern, J.; Reza, M.; Knicker, H. Hydrothermal carbonization and pyrolysis of sewage sludges: What happen to carbon and nitrogen? J. Anal. Appl. Pyrolysis 2017, 128, 314–323. [Google Scholar] [CrossRef]
- Lehmann, J. Bio-energy in the black. Front. Ecol. Environ. 2007, 5, 381–387. [Google Scholar] [CrossRef]
- Singh, B.; Singh, B.P.; Cowie, A.L. Characterisation and evaluation of biochars for their application as a soil amendment. Aust. J. Soil Res. 2010, 48, 516–525. [Google Scholar] [CrossRef]
- Hagemann, N.; Subdiaga, E.; Orsetti, S.; De la Rosa, J.M.; Knicker, H.; Schmidt, H.-P.; Kappler, A.; Behrens, S. Effect of biochar amendment on compost organic matter composition following aerobic composting of manure. Sci. Total Environ. 2018, 613, 20–29. [Google Scholar] [CrossRef]
- Sarfraz, M.; Mehdi, S.; Abid, M.; Akram, M. External and internal phosphorus requirement of wheat in bhalike soil series of pakistan. Pak. J. Bot. 2008, 40, 2031–2040. [Google Scholar]
- Ahmad, I.; Bibi, F.; Ullah, H.; Munir, T.M. Mango fruit yield and critical quality parameters respond to foliar and soil applications of zinc and boron. Plants 2018, 7, 97. [Google Scholar] [CrossRef]
- Dworkin, M.; Foster, J.W. Experiments with some microorganisms which utilize ethane and hydrogen. J. Bacteriol. 1958, 75, 592–603. [Google Scholar] [PubMed]
- El-Tarabily, K.A. Promotion of tomato (Lycopersicon esculentum mill.) plant growth by rhizosphere competent 1-aminocyclopropane-1-carboxylic acid deaminase-producing streptomycete actinomycetes. Plant Soil 2008, 308, 161–174. [Google Scholar] [CrossRef]
- Glickmann, E.; Dessaux, Y. A critical examination of the specificity of the salkowski reagent for indolic compounds produced by phytopathogenic bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [PubMed]
- Vazquez, P.; Holguin, G.; Puente, M.E.; Lopez-Cortes, A.; Bashan, Y. Phosphate-solubilizing microorganisms associated with the rhizosphere of mangroves in a semiarid coastal lagoon. Biol. Fertil. Soils 2000, 30, 460–468. [Google Scholar] [CrossRef]
- Sheng, X.F.; He, L.Y. Solubilization of potassium-bearing minerals by a wild-type strain of bacillus edaphicus and its mutants and increased potassium uptake by wheat. Can. J. Microbiol. 2006, 52, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.; Pratt, P. Methods of Analysis for Soils, Plants and Waters; University of California, Division of Agricultural Sciences: Oakland, CA, USA, 1961; pp. 169–176. [Google Scholar]
- Tandon, H.; Cescas, M.; Tyner, E. An acid-free vanadate-molybdate reagent for the determination of total phosphorus in soils 1. Soil Sci. Soc. Am. J. 1968, 32, 48–51. [Google Scholar] [CrossRef]
- Jones, J.B., Jr.; Wolf, B.; Mills, H.A. Plant Analysis Handbook. A Practical Sampling, Preparation, Analysis, and Interpretation Guid; Micro-Macro Publishing, Inc.: Athens, GA, USA, 1991. [Google Scholar]
- Ronsse, F.; Van Hecke, S.; Dickinson, D.; Prins, W. Production and characterization of slow pyrolysis biochar: Influence of feedstock type and pyrolysis conditions. Gcb Bioenergy 2013, 5, 104–115. [Google Scholar] [CrossRef]
- Gee, G.; Bauder, J. Particle-size analysis. Agronomy 1986, 4, 255–293. [Google Scholar]
- Walkley, A. An examination of methods for determining organic carbon and nitrogen in soils. J. Agric. Sci. 1935, 25, 598–609. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus. In Method of Soil Analysis, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 403–430. [Google Scholar]
- Ahmad, I.; Akhtar, M.J.; Zahir, Z.A.; Naveed, M.; Mitter, B.; Sessitsch, A. Cadmium-tolerant bacteria induce metal stress tolerance in cereals. Environ. Sci. Pollut. R. 2014, 21, 11054–11065. [Google Scholar] [CrossRef]
- Ahmad, M.T.; Asghar, N.; Saleem, M.; Khan, M.Y.; Zahir, Z.A. Synergistic effect of rhizobia and biochar on growth and physiology of maize. Agron. J. 2015, 107, 2327–2334. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Zahir, Z.A.; Naveed, M.; Arshad, M. Rhizobacteria containing acc-deaminase confer salt tolerance in maize grown on salt-affected fields. Can. J. Microbiol. 2009, 55, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Shaharoona, B.; Arshad, M.; Zahir, Z.A. Effect of plant growth promoting rhizobacteria containing acc-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (vigna radiata L.). Lett. Appl. Microbiol. 2006, 42, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.; Rehman, T.; Yates, C. Logit models for identifying the factors that influence the uptake of new ‘no-tillage’technologies by farmers in the rice–wheat and the cotton–wheat farming systems of pakistan’s punjab. Agric. Syst. 2003, 75, 79–95. [Google Scholar] [CrossRef]
- Kumar, K.; Prihar, S.; Gajri, P. Determination of root distribution of wheat by auger sampling. Plant Soil 1993, 149, 245–253. [Google Scholar] [CrossRef]
- Newman, E. A method of estimating the total length of root in a sample. J. Appl. Ecol. 1966, 3, 139–145. [Google Scholar] [CrossRef]
- Munir, T.; Khadka, B.; Xu, B.; Strack, M. Partitioning forest-floor respiration into source based emissions in a boreal forested bog: Responses to experimental drought. Forests 2017, 8, 75. [Google Scholar] [CrossRef]
- Tekalign, T.; Hammes, P. Growth and productivity of potato as influenced by cultivar and reproductive growth: I. Stomatal conductance, rate of transpiration, net photosynthesis, and dry matter production and allocation. Sci. Hortic. 2005, 105, 13–27. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts:Polyphenoloxidase in beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Nacl-induced senescence in leaves of rice (oryza sativa L.) cultivars differing in salinity resistance. Ann. Bot. 1996, 78, 389–398. [Google Scholar] [CrossRef]
- Steel, R.G.D.; Torrie, J.H. Principles and Procedures of Statistics, a Biometrical Approach; McGraw-Hill Kogakusha, Ltd.: New York, NY, USA, 1980. [Google Scholar]
- Horel, Á.; Tóth, E.; Gelybó, G.; Dencső, M.; Farkas, C. Biochar amendment affects soil water and CO2 regime during capsicum annuum plant growth. Agronomy 2019, 9, 58. [Google Scholar] [CrossRef]
- Sánchez-Monedero, M.A.; Cayuela, M.L.; Sánchez-García, M.; Vandecasteele, B.; D’Hose, T.; López, G.; Martínez-Gaitán, C.; Kuikman, P.J.; Sinicco, T.; Mondini, C. Agronomic evaluation of biochar, compost and biochar-blended compost across different cropping systems: Perspective from the european project fertiplus. Agronomy 2019, 9, 225. [Google Scholar] [CrossRef]
- Amonette, J.E.; Joseph, S. Characteristics of biochar: Microchemical properties. In Biochar for Environmental Management; Routledge: London, UK, 2012; pp. 65–84. [Google Scholar]
- Glick, B.R.; Liu, C.; Ghosh, S.; Dumbroff, E.B. Early development of canola seedlings in the presence of the plant growth-promoting rhizobacterium pseudomonas putida gr12-2. Soil Biol. Biochem. 1997, 29, 1233–1239. [Google Scholar] [CrossRef]
- Xie, H.; Pasternak, J.J.; Glick, B.R. Isolation and characterization of mutants of the plant growth-promoting rhizobacterium pseudomonas putida gr12-2 that overproduce indoleacetic acid. Curr. Microbiol. 1996, 32, 67–71. [Google Scholar] [CrossRef]
- Mohite, B. Isolation and characterization of indole acetic acid (iaa) producing bacteria from rhizospheric soil and its effect on plant growth. J. Soil Sci. Plant Nutr. 2013, 13, 638–649. [Google Scholar] [CrossRef]
- Pérez-Fernández, M.; Alexander, V. Enhanced plant performance in cicer arietinum l. Due to the addition of a combination of plant growth-promoting bacteria. Agriculture 2017, 7, 40. [Google Scholar] [CrossRef]
- Ma, Y.; Látr, A.; Rocha, I.; Freitas, H.; Vosátka, M.; Oliveira, R.S. Delivery of inoculum of rhizophagus irregularis via seed coating in combination with pseudomonas libanensis for cowpea production. Agronomy 2019, 9, 33. [Google Scholar] [CrossRef]
- De la Rosa, J.M.; Paneque, M.; Hilber, I.; Blum, F.; Knicker, H.E.; Bucheli, T.D. Assessment of polycyclic aromatic hydrocarbons in biochar and biochar-amended agricultural soil from southern spain. J. Soils Sediments 2016, 16, 557–565. [Google Scholar] [CrossRef]
- Chan, K.Y.; Zwieten, L.V.; Meszaros, I.; Downie, A.; Joseph, S. Using poultry litter biochars as soil amendments. Aust. J. Soil Res. 2008, 46, 437–444. [Google Scholar] [CrossRef]
- Singh, A.; Singh, A.P.; Singh, S.K.; Rai, S.; Kumar, D. Impact of addition of biochar along with pgpr on rice yield, availability of nutrients and their uptake in alluvial soil. J. Pure Appl. Microbiol. 2016, 10, 2181–2188. [Google Scholar]
- Shabala, S. Regulation of potassium transport in leaves: From molecular to tissue level. Ann. Bot. 2003, 95, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, S.; Davies, W.J. Aba-based chemical signalling: The co-ordination of responses to stress in plants. Plant Cell Environ. 2002, 25, 195–210. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.M.; Lima, I.; Xing, B.; Gaskin, J.W.; Steiner, C.; Das, K.C.; Ahmedna, M.; Rehrah, D.; Watts, D.W.; Busscher, W.J.; et al. Characterization of designer biochar produced at different temperatures and their effects on a loamy sand. Ann. Environ. Sci. 2009, 3, 195–2006. [Google Scholar]
- Lehmann, J.; Rillig, M.C.; Thies, J.; Masiello, C.A.; Hockaday, W.C.; Crowley, D. Biochar effects on soil biota—A review. Soil Biol. Biochem. 2011, 43, 1812–1836. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Adrees, M.; Zia-ur-Rehman, M.; Qayyum, M.F.; Ok, Y.S.; Murtaza, G. Effect of biochar on alleviation of cadmium toxicity in wheat (Triticum aestivum L.) grown on cd-contaminated saline soil. Environ. Sci. Pollut. R. 2018, 25, 25668–25680. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Soil | Unit | Value | Biochar | Unit | Value |
|---|---|---|---|---|---|
| Sand | % | 55 | pH | - | 7.26 |
| Silt | % | 25 | ECe | dS m−1 | 1.22 |
| Clay | % | 30 | Volatile Matter | % | 8.96 |
| Texture | Sandy Clay Loam | Ash Content | % | 28.9 | |
| pHs | - | 8.52 | Fixed Carbon | % | 62.1 |
| ECe | dS m−1 | 3.69 | Total N | % | 0.21 |
| Organic Matter | % | 0.45 | Total P | % | 0.62 |
| Total N | % | 0.02 | Total K | % | 1.61 |
| Extractable P | mg kg−1 | 5.26 | Total Na | % | 0.19 |
| Extractable K | mg kg−1 | 170 | |||
| Treatments | Plant Height (cm) | Root Length (cm) | Spike Length (cm) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | ||||||||||||
| IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | |||||||
| 4I | 3I | 2I | 4I | 3I | 2I | 4I | 3I | 2I | ||||
| Control | 59.0 d–f | 48.1 g,h | 33.0 i | 46.7 D | 8.82 d–f | 5.65 h,i | 4.46 i | 6.31 D | 5.75 | 4.86 | 4.34 | 4.98 B |
| A. fabrum | 65.5 c–f | 58.3 e,f | 40.1 h,i | 54.6 C | 10.1 b–d | 7.08 f–h | 6.22 g–i | 7.79 C | 6.68 | 6.22 | 4.87 | 5.92 A |
| B. amyloliquefaciens | 66.2 b–e | 60.0 d–f | 39.6 h,i | 55.3 B,C | 10.3 b–d | 7.46 f–h | 6.36 g,h | 8.03 C | 6.69 | 6.27 | 4.78 | 5.91 A |
| BC | 71.2 a–c | 60.9 d–f | 45.6 h | 59.2 B | 11.5 a–c | 9.67 c–e | 7.81 e–g | 9.67 B | 6.67 | 6.46 | 4.96 | 6.03 A |
| A. fabrum + BC | 76.8 a | 68.3 a–d | 56.5 f,g | 67.2 A | 12.8 a | 11.2 a–c | 8.46 d–f | 10.8 A | 7.13 | 6.33 | 5.10 | 6.19 A |
| B. amyloliquefaciens + BC | 75.5 a,b | 68.3 a–d | 59.7 d–f | 67.8 A | 13.1 a | 11.9 a,b | 8.49 d–f | 11.2 A | 7.12 | 6.55 | 5.05 | 6.24 A |
| ME (I) | 69.0 A | 60.7 B | 45.8 C | 11.1 A | 8.83 B | 6.97 C | 6.67 A | 6.1 B | 4.85 C | |||
| Treatments | Spikelets spike−1 | Grains Spike−1 | 1000 Grains Weight (g) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | ||||||||||||
| IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | |||||||
| 4I | 3I | 2I | 4I | 3I | 2I | 4I | 3I | 2I | ||||
| Control | 15.3 | 13.0 | 10.7 | 13.0 D | 37.7 | 27.7 | 22.0 | 29.1 C | 35.3 b–d | 28.2 f,g | 20.2 h | 27.9 C |
| A. fabrum | 16.0 | 13.7 | 13.0 | 14.2 C,D | 38.3 | 33.7 | 26.7 | 32.9 B,C | 34.2 c–f | 30.7 c–g | 25.5 g,h | 30.2 B,C |
| B. amyloliquefaciens | 16.0 | 13.3 | 12.7 | 14.0 C | 39.3 | 35.0 | 29.7 | 34.7 B | 35.2 b–e | 31.0 c–g | 26.8 g | 31.0 B |
| BC | 16.3 | 14.3 | 13.0 | 14.6 B,C | 47.3 | 42.0 | 32.7 | 40.7 A | 36.3 a–c | 31.1 c–g | 29.4 d–f | 32.3 B |
| A. fabrum + BC | 17.0 | 15.3 | 14.3 | 15.6 A,B | 45.3 | 40.7 | 39.0 | 41.7 A | 42.5 a | 34.7 b–e | 29.4 d–f | 35.5 A |
| B. amyloliquefaciens + BC | 17.3 | 15.7 | 15.3 | 16.1 A | 49.0 | 43.0 | 39.3 | 43.8 A | 40.8 a,b | 36.5 a–c | 28.9 e–f | 35.4 A |
| ME (I) | 16.3 A | 14.2 B | 13.2 C | 42.8 A | 37.0 B | 31.6 C | 37.4 A | 32.0 B | 26.7 C | |||
| Treatments | Grains Nitrogen (%) | Grains Phosphorus (%) | Grains Potassium (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | ||||||||||||
| IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | |||||||
| 4I | 3I | 2I | 4I | 3I | 2I | 4I | 3I | 2I | ||||
| Control | 2.58 a–e | 1.99 h–j | 1.54 k | 2.04 D | 0.66 d–g | 0.43 i | 0.25 j | 0.44 D | 0.50 b–e | 0.45 e | 0.31 f | 0.42 D |
| A. fabrum | 2.78 a,b | 2.21 f–i | 1.79 j,k | 2.26 C | 0.71 c–f | 0.60 f–h | 0.45 i | 0.59 C | 0.55 a–c | 0.47 d,e | 0.43 e | 0.48 C |
| B. amyloliquefaciens | 2.73 a–c | 2.33 e–h | 1.86 i–k | 2.31 C | 0.75 a–e | 0.61 e–h | 0.48 h,i | 0.61 B,C | 0.56 a–c | 0.48 c–e | 0.46 e | 0.50 C |
| BC | 2.74 a–c | 2.55 b–f | 2.13 g–j | 2.47 B | 0.78 a–d | 0.68 c–g | 0.54 g–i | 0.67 B | 0.58 a | 0.50 b–e | 0.46 e | 0.51 B,C |
| A. fabrum + BC | 2.87 a,b | 2.68 a–d | 2.33 d–h | 2.63 A,B | 0.87 a,b | 0.73 b–f | 0.62 e–h | 0.74 A | 0.60 a | 0.54 a–d | 0.49 c–e | 0.55 A,B |
| B. amyloliquefaciens + BC | 2.92 a | 2.76 a–c | 2.42 c–g | 2.70 A | 0.87 a | 0.82 a–c | 0.66 d–g | 0.78 A | 0.61 a | 0.57 a,b | 0.50 b–e | 0.56 A |
| ME (I) | 2.77 A | 2.42 B | 2.01 C | 0.77 A | 0.65 B | 0.50 C | 0.57 A | 0.50 B | 0.44 C | |||
| Treatments | Shoot Nitrogen (%) | Shoot Phosphorus (%) | Shoot Potassium (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | |||||||||
| IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | ||||
| 4I | 3I | 4I | 3I | 4I | 3I | ||||
| Control | 1.88 a | 1.33 c | 1.60 B | 0.42 | 0.26 | 0.34 C | 1.85 | 1.56 | 1.71 D |
| A. fabrum | 1.85 a | 1.58 b,c | 1.71 A,B | 0.46 | 0.36 | 0.41 B | 2.20 | 1.97 | 2.09 C |
| B. amyloliquefaciens | 1.85 a | 1.57 b,c | 1.71 A,B | 0.45 | 0.38 | 0.42 B | 2.20 | 2.01 | 2.11 C |
| BC | 1.88 a | 1.59 b | 1.73 A,B | 0.48 | 0.42 | 0.45 A,B | 2.56 | 2.17 | 2.37 B |
| BC + A. fabrum | 1.93 a | 1.75 a,b | 1.84 A | 0.51 | 0.43 | 0.47 A | 2.69 | 2.47 | 2.58 A |
| BC + B. amyloliquefaciens | 1.94 a | 1.75 a,b | 1.85 A | 0.53 | 0.45 | 0.49 A | 2.68 | 2.44 | 2.56 A |
| ME (I) | 1.89 A | 1.59 B | 0.47 A | 0.39 B | 2.37 A | 2.10 B | |||
| Treatments | Photosynthetic Rate (µmol (CO2) m−2 s−1) | Transpiration Rate (mmol (H2O) m−2 s−1) | Stomatal Conductance (µmol (CO2) m−2 s−1) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | |||||||||
| IE (T × I) | ME | IE (T × I) | ME (T) | IE (T × I) | ME (T) | ||||
| 4I | 3I | (T) | 4I | 3I | 4I | 3I | |||
| Control | 14.5 | 9.07 | 11.8 C | 4.35 | 2.97 | 3.66 D | 150.7 | 105.3 | 128.0 C |
| A. fabrum | 16.1 | 10.7 | 13.4 C | 4.90 | 4.23 | 4.56 C | 148.3 | 125.7 | 137.0 C |
| B. amyloliquefaciens | 15.9 | 10.1 | 13.0 B,C | 5.41 | 4.17 | 4.79 B,C | 166.7 | 127.7 | 147.2 B,C |
| BC | 17.4 | 13.5 | 15.5 A,B | 6.27 | 4.64 | 5.46 B | 181.3 | 145.0 | 163.2 A,B |
| A. fabrum + BC | 18.6 | 15.6 | 17.1 A | 7.35 | 5.85 | 6.60 A | 193.0 | 156.3 | 174.7 A |
| B. amyloliquefaciens + BC | 19.1 | 15.8 | 17.5 A | 7.86 | 5.43 | 6.64 A | 193.3 | 172.3 | 182.8 A |
| ME (I) | 16.9 A | 12.5 B | 6.02 A | 4.55 B | 172.2 A | 138.7 B | |||
| Treatments | Chlorophyll a (mg g−1) | Chlorophyll b (mg g−1) | Total Chlorophyll (mg g−1) | Electrolyte Leakage (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. of Irrigations (I) | ||||||||||||
| IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | IE (T × I) | ME (T) | |||||
| 4I | 3I | 4I | 3I | 4I | 3I | 4I | 3I | |||||
| Control | 0.87 | 0.68 | 0.77 C | 0.42 | 0.34 | 0.38 C | 1.29 | 1.02 | 1.15 C | 41.0 | 59.3 | 50.2 A |
| A. fabrum | 0.91 | 0.78 | 0.85 B,C | 0.47 | 0.39 | 0.43 B,C | 1.38 | 1.17 | 1.28 B,C | 40.3 | 55.3 | 47.8 A,B |
| B. amyloliquefaciens | 0.90 | 0.78 | 0.84 B,C | 0.48 | 0.38 | 0.43 B,C | 1.37 | 1.16 | 1.27 B,C | 41.3 | 54.0 | 47.7 A,B |
| BC | 0.99 | 0.85 | 0.92 B | 0.48 | 0.42 | 0.45 B | 1.47 | 1.27 | 1.37 B | 41.0 | 47.0 | 44.0 A,B |
| A. fabrum + BC | 1.16 | 0.98 | 1.07 A | 0.53 | 0.45 | 0.49 A,B | 1.68 | 1.44 | 1.56 A | 39.0 | 41.0 | 40.0 B |
| B. amyloliquefaciens + BC | 1.10 | 1.06 | 1.08 A | 0.59 | 0.49 | 0.54 A | 1.69 | 1.55 | 1.62 A | 37.0 | 42.3 | 39.7 B |
| ME (I) | 0.99 A | 0.86 B | 0.49 A | 0.41 B | 1.48 A | 1.27 B | 39.9 B | 49.8 A | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zafar-ul-Hye, M.; Danish, S.; Abbas, M.; Ahmad, M.; Munir, T.M. ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress. Agronomy 2019, 9, 343. https://doi.org/10.3390/agronomy9070343
Zafar-ul-Hye M, Danish S, Abbas M, Ahmad M, Munir TM. ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress. Agronomy. 2019; 9(7):343. https://doi.org/10.3390/agronomy9070343
Chicago/Turabian StyleZafar-ul-Hye, Muhammad, Subhan Danish, Mazhar Abbas, Maqshoof Ahmad, and Tariq Muhammad Munir. 2019. "ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress" Agronomy 9, no. 7: 343. https://doi.org/10.3390/agronomy9070343
APA StyleZafar-ul-Hye, M., Danish, S., Abbas, M., Ahmad, M., & Munir, T. M. (2019). ACC Deaminase Producing PGPR Bacillus amyloliquefaciens and Agrobacterium fabrum along with Biochar Improve Wheat Productivity under Drought Stress. Agronomy, 9(7), 343. https://doi.org/10.3390/agronomy9070343