3.1. Soil Properties
The means and standard deviations (s.d.) of the measured physical attributes are presented here to provide estimates of soil properties at the sampled locations within paddocks (
Table 4). Soils on the study site showed a degree of variability in physical attributes among paddocks assigned to the two stocking methods. Percentages of different particle fractions of soils in paddocks assigned to continuous stocking were less consistent than in sub-paddocks assigned to rotational stocking, which was likely due to the greater number of soils encountered within these larger units [
25]. Paddocks receiving continuous stocking had greater percentages of sand and lower amounts of silt than in rotational-stocked sub-paddocks, while percentages of clay in soils were similar. Moist bulk densities of soils within paddocks receiving rotational stocking were lower than in paddocks receiving continuous stocking, most likely due to the higher percentage of sand in soils of continuously stocked paddocks [
34]. Amounts of soil organic matter showed distinct differences among stocking methods. Paddocks managed by continuous stocking had lower concentrations than were recorded in paddocks managed under rotational stocking. However, organic matter in soils was more variable in paddocks receiving rotational stocking (c.v. = 29% vs. 24%).
A review of these properties (
Table 4) indicates the two stocking methods may have affected some of the physical properties of soil, as has been noted in responses to both longer- and shorter-term applications of stocking methods [
13,
35,
36]. However, the amount of natural variability (e.g., standard deviation) present in the measured attributes, which is also a component of the landscapes enclosed within paddock boundaries [
1], indicated the means were not consistent within different paddocks assigned to the same stocking method. The location of paddocks within the landscape of the study area likely affected these properties. The continuous stocked paddocks had predominantly western exposures, while rotationally stocked sub-paddocks had largely easterly exposures. Such differences in exposure can have large effects on catena-based soil development within landscapes [
1,
2,
34]. This difference in predominant exposure likely contributed to the higher amounts of sand, corresponding greater bulk densities, and lower amounts of silt, noted in the continuously stocked paddocks.
Stocking methods have some capacity to result in changes in soil organic matter and bulk density in the US Southern Great Plains (SGP) over time. Research in south-central Oklahoma, United States [
35], noted increasing bulk densities of soils of perennial grassland in response to 10 years of different animal densities applied by rotational stocking, compared to no grazing. However, the higher bulk density of soils in the continuously stocked paddocks of the current study may be related to the greater amounts of sand that were recorded [
34]. Concentrations of soil organic matter were lower in the continuously stocked paddocks, though amounts were variable within paddocks managed under both stocking methods. However, the distribution of soil properties and the native grasslands they support varies within landscapes without grazing [
2]. For example, variable patterns were recorded in the spatial distribution of bulk density and organic matter of soils in 1.6 ha paddocks of southern tallgrass prairie within 2 km of this study site, after 25 years of three sustained stocking methods [
15]. Distribution patterns noted within those paddocks were definable both within and across multiple paddocks, often within 20 m of spatial scale, which defined the catenae-scale organization of the landscape. Such patterns indicate soils in the area of the current study, which were defined as members of different families and subgroups of Mollisols, likely had similar non-uniform distributions, and variable, fine-scale patterns of spatial distribution.
3.2. Mineral N
Main effects related to stocking method (0.15 <
p < 0.76) and stocking interactions with paddock locations (0.21 <
p < 0.34) were not significant for NO
3−, NH
4+, or total N. This lack of effect by stocking method in general, and their effects on N fluxes at paddock locations, was unexpected given the number of years of applied treatments and broad range of sampled locations (
n = 16). These locations included a series with a low likelihood of animal visitation, such as the middle area of paddocks under continuous stocking under the low animal densities that were applied. In comparison, studies in introduced perennial grasslands reported significantly greater amounts of NO
3− and total mineral N with increase in length of time cattle grazed rotationally stocked sub-paddocks, but found no differences between effects of longer rotational or continuous stocking [
10,
37]. Studies on the effects of stocking methods on soil properties of native prairie in northcentral Texas, United States, also reported no differences in NO
3−-N concentrations between continuous and rotationally stocked native prairie [
13].
Stocking methods interacted with soil depth for NO
3− (F
1, 194 = 3.4;
p = 0.07) and total N (F
1, 194 = 3.4;
p = 0.07). The stocking method × with time of year interaction on NH
4+ flux was also significant (F
1, 194 = 16.6;
p < 0.01). Main effects related to paddock location were also significant (0.01 <
p < 0.08) for NO
3−, NH
4+ and total N. The highest amount of total mineral and NO
3−N within the stocking method × soil depth interactions (
Table 5) were noted for the upper 7.5 cm of the profile under rotational stocking, with the second-highest amounts recorded for the upper 7.5 cm under continuous stocking. The lowest amounts of NO
3− flux were recorded in the upper soil depth under rotational stocking. Within the stocking method × time of year interaction on NH
4+, the highest and second-highest amounts of flux were recorded, respectively, in response to rotational and continuous stocking during March. The lowest amounts of flux were noted for responses to rotational stocking during August. The greater total and NO
3-N fluxes recorded in the upper 7.5 cm of soil was similar to results noted in studies on other warm-season grasslands [
10]. The higher amounts noted during March were likely related to cooler soil temperatures that occur during winter and early-spring in the region, which result in lower amounts of mineralization of NH
4+ to NO
3− [
3]. Research in paddocks of other warm-season grasses in Florida, United States, noted no differences in concentrations of NH
4+ among different forms of rotational and continuous stocking [
10].
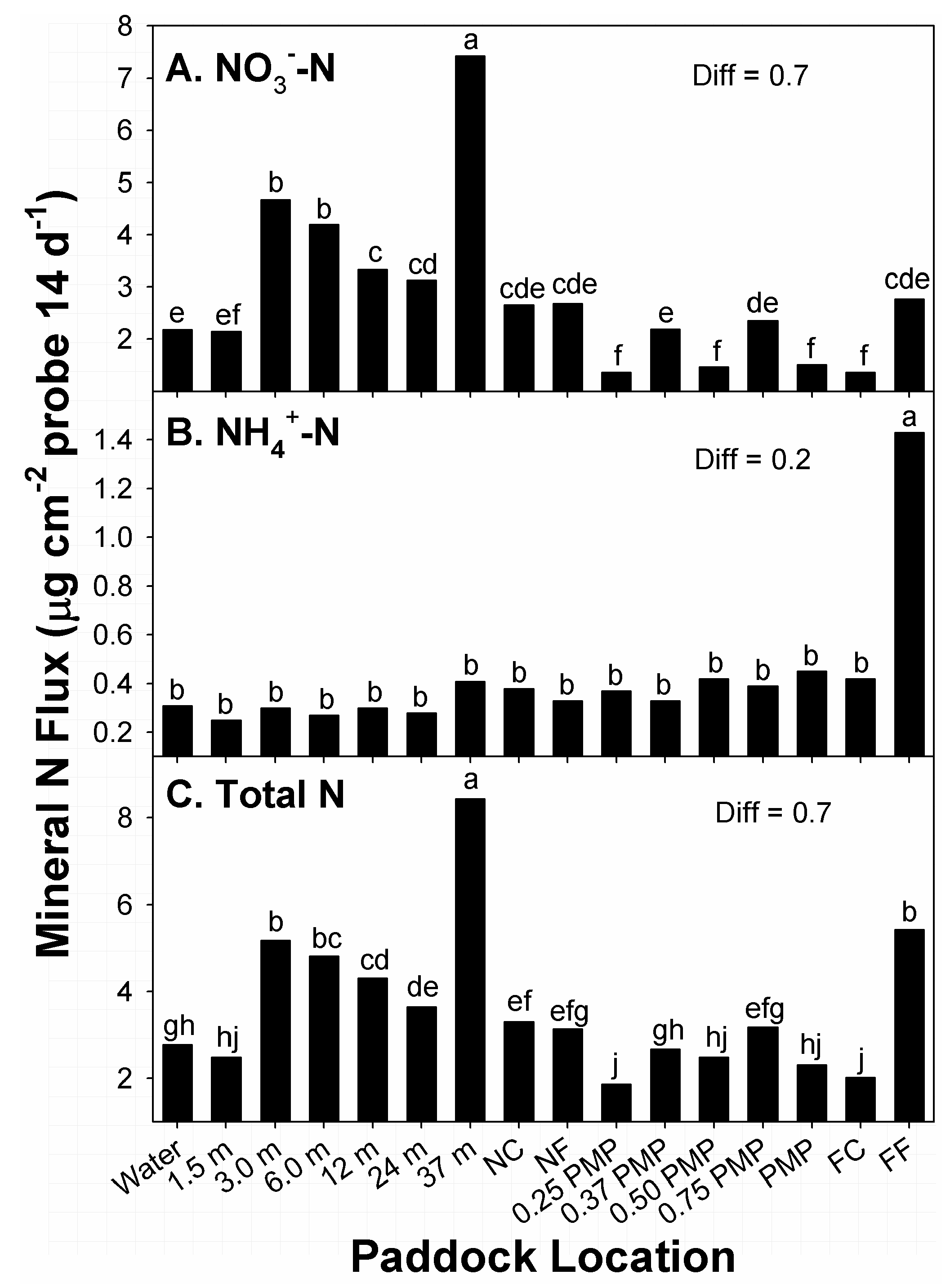
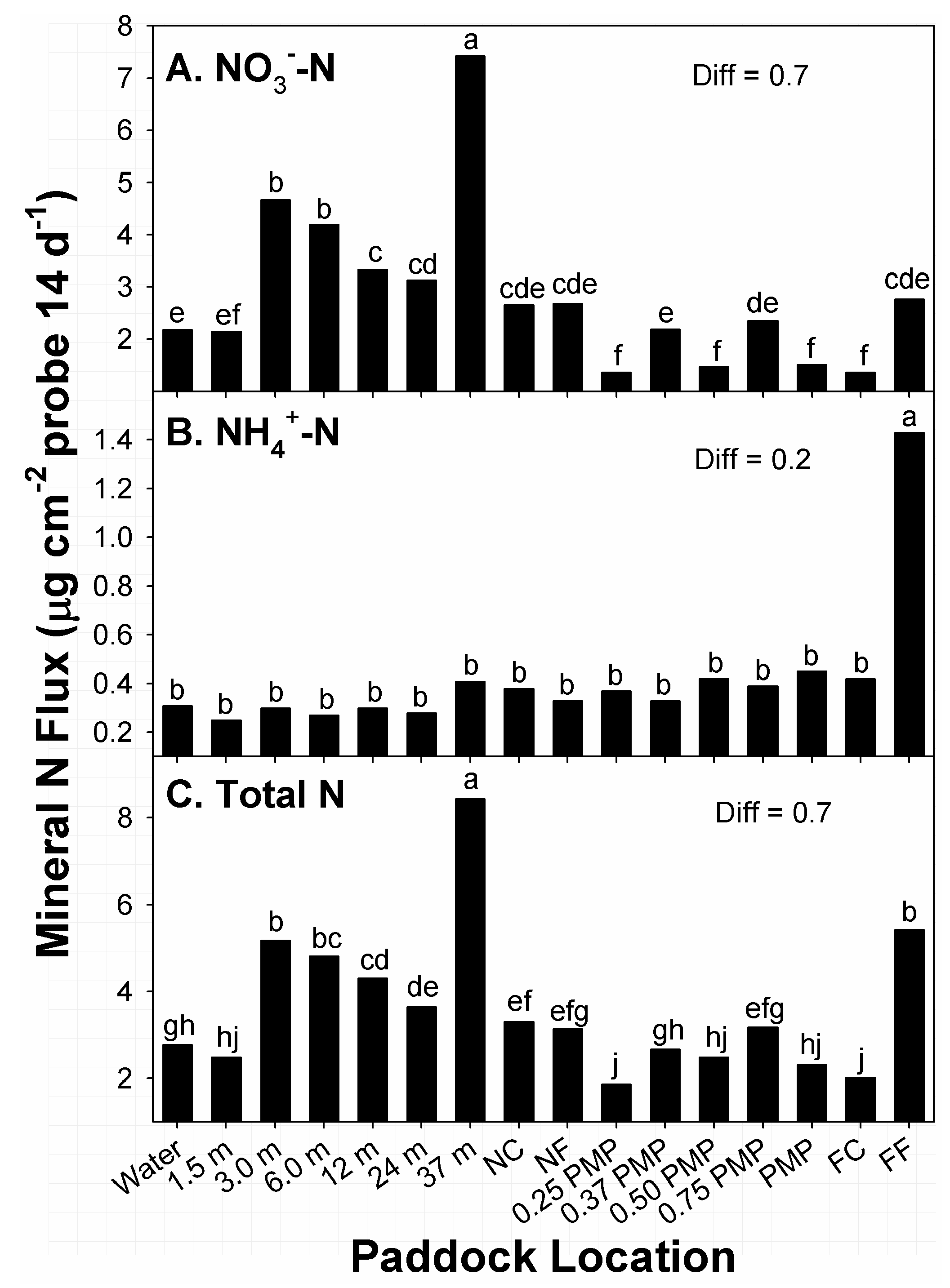
The greatest amounts of flux in NO
3− among paddock locations occurred at locations 37 m from water sources, with the second-greatest amounts noted from 3 to 6 m from water (
Figure 2A). Alternatively, the lowest amounts of flux were recorded at locations near 0.25 PMP, 0.5 PMP, PMP, and FC. Locations with the second-lowest amounts of flux in NO
3− were noted at water sources and 1.5 m from water. Amounts of flux at remaining locations belonged to groups with intermediate amounts. Overall, there was a general decline in amounts of NO
3− flux from 3 m from water sources to the far corner of paddocks (FC), with the notable exception of 37 m.
The distribution of amounts of NH
4+ flux recorded at paddock locations differed from the distribution noted for NO
3− (
Figure 2B). The greatest amounts of flux in NH
4+ among paddock locations occurred along fence lines furthest from water sources (FF). Amounts of flux (0.3 to 0.5 µg NH
4+ cm
−2) at all remaining sampled locations were ≤31% of this level, and belonged to the same means group. The distribution of NH
4+ within paddocks was unusual, given the 3- to 7-fold greater amounts at FF, across stocking methods, compared to the other locations. Studies in paddocks of tame warm-season grasses noted greater amounts of NH4+ in zones of paddocks that were closest and furthest from water, relative to intermediate locations [
37]. The driver for high amounts of NH
4+ flux at FF in the current study was not clear, as these locations were not consistent in terms of landscape positions, or distances from water. The FF locations were situated at both tread and toe positions of the landscapes of the two continuously stocked and four rotationally stocked units that were sampled. However, the amounts of NH
4+ flux recorded in differences among locations were small, covering a range of 1.1 µg cm
−2 of surface areas of probes.
The distribution of amounts of flux in total mineral N recorded at paddock locations were somewhat similar to the distribution pattern noted for NO
3− (
Figure 2C). This similarity was related to the high amounts of NO
3− relative to the total amount; 65% to 88% of total mineral N at 14 of 16 locations was NO
3−. The greatest amounts of flux in total N among paddock locations occurred at locations 37 m from water sources, with the second-greatest amounts noted from 3.0 from water, and at sampled locations along the fence furthest from water sources. Alternatively, the lowest amounts of flux were recorded at locations near 0.25 PMP and FC. Locations with the second-lowest amounts of flux in total N were noted within 1.5 m of water sources, 0.37 PMP, 0.5 PMP, and PMP. Amounts of flux at remaining locations belonged to different groups with intermediate amounts. As with NO
3−, there was a general decline in amounts of flux in total N from 3.0 m from water sources to the far corner of paddocks, with the notable exception of 37 m.
Other studies have noted hotspots of mineral N in soils at specific locations within paddocks, especially in closer proximity to water, corners, and along fences, due to uneven distribution of animal use of paddock areas [
38]. For example, research [
8] reported that cattle grazing shortgrass rangeland in northeast Colorado, United States, spent ~27% of the time on paddocks at locations near water sources and corners, which represented 2.5% of the total area of paddocks, resulting in shifts in N distributions. Studies undertaken to define redistribution of N in tamegrass paddocks in Florida, United States, noted greater amounts of mineral N in zones closer to water and shade, relative to the remainder of paddock areas [
10,
37]. These distribution patterns were similar across different forms of rotational and continuous stocking applied to bermudagrass (
Cynodon dactylon) and bahiagrass (
Paspalum notatum) paddocks [
10,
37]. In comparison, the distribution of flux in plant available NO
3− and total mineral N in the current study showed variability among sampled locations, but higher amounts occurred at locations other than in immediate proximity to water. Further, fluxes in plant available N at other (supposed) high-traffic locations (i.e., NC and FC) were among the lower groups of mean responses that were recorded. Other studies on native rangeland noted positive correlations between heterogeneity in paddock use by cattle and size of paddocks [
11]. Given the variable distributions of mineral N in the current study, the large sizes of paddocks and sub-paddocks may have limited uniform use of paddock (and sub-paddock) areas by cattle.
3.3. Phosphorus and Magnesium
There were significant soil depth (F
1, 194 = 86.8;
p < 0.01) and paddock location (F
15, 194 = 5.3;
p < 0.01) main effects on flux of available P. The interaction between stocking method and time of year was also significant (F
1, 194 = 24.8;
p < 0.01) for flux in available P. Among effects of soil depth, greater amounts of P flux were noted within the upper 7.5 cm depth than the 7.5–15 cm increment (0.75 versus 0.28 µg P cm
−2; Diff = 0.45 µg). Greater amounts of flux in plant-available P within the uppermost segment of the soil profile was not unexpected, as such responses have been reported elsewhere for a range of different grasslands [
3]. Larger concentrations in P were recorded in the upper 15 cm of soil than in deeper increments of bermudagrass paddocks in Florida, United States, managed under continuous and rotational stocking methods, with similar responses across stocking methods [
10].
Within the stocking method × time of year interaction (
Table 6), the largest fluxes in plant-available P occurred in response to rotational stocking during March, while the second-largest occurred under continuous stocking during August. The lowest flux in available P within the interaction was recorded for responses to rotational stocking during August. Differences among mean amounts of plant-available P during the growing season are driven by amounts present in soil solution at the start of growing seasons, plus amounts that become soluble during growing seasons [
3]. Responses to continuous stocking indicated a degree of consistency between these factors and uptake by plants (
Table 6), while declines in P flux under rotational stocking in summer may indicate depletion of available pools. However, the concentration of plant-available P in soils is generally low [
3], so the effects of stocking methods during different times of the growing season are unclear. It is difficult to assess what such low fluxes in plant-available P represent, as it is only a portion of the entire P pool. Other research on P distributions within paddocks of grassland under rotational and continuous stocking noted differences in amounts related to lengths of applied grazing [
10,
37].
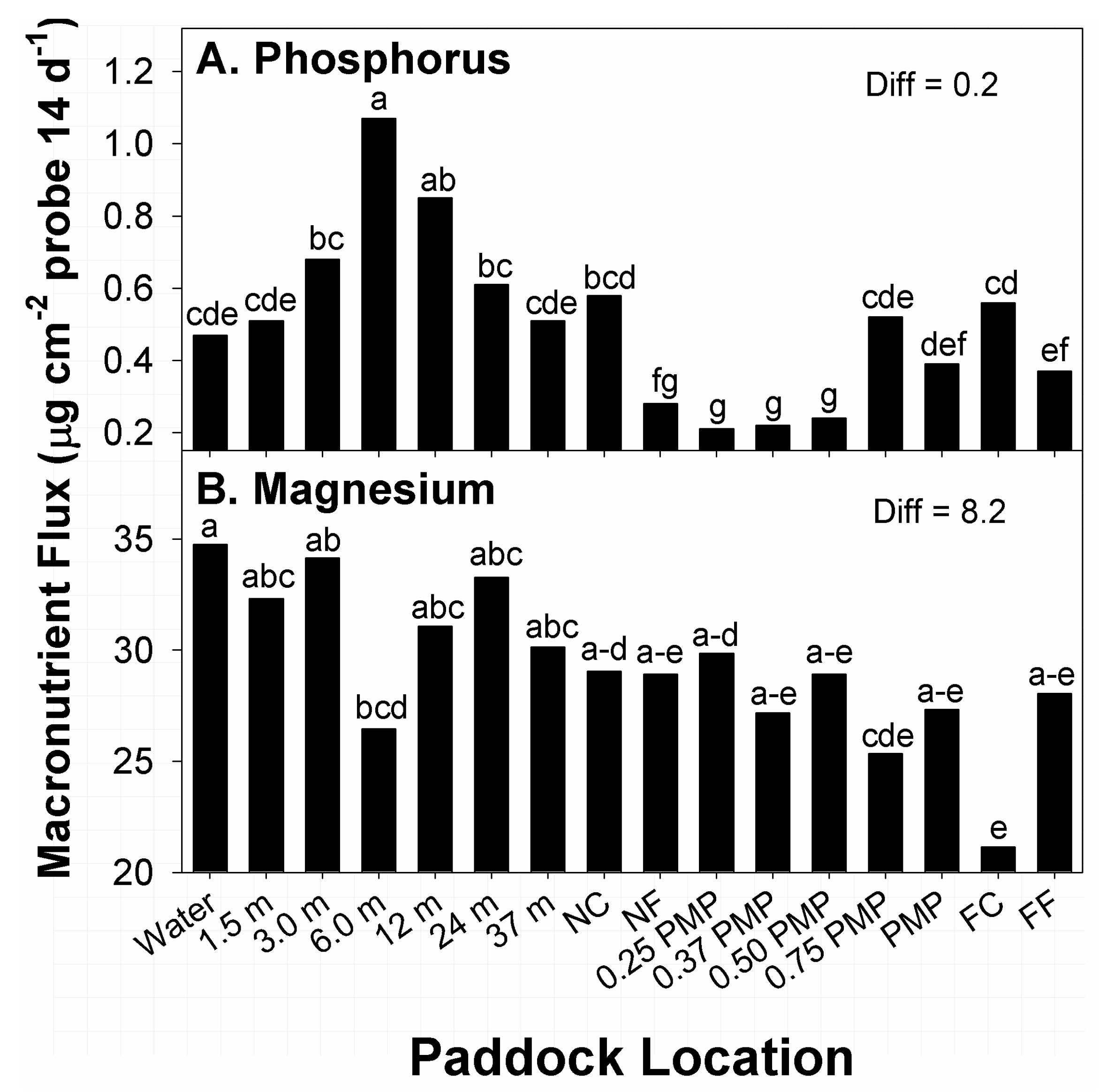
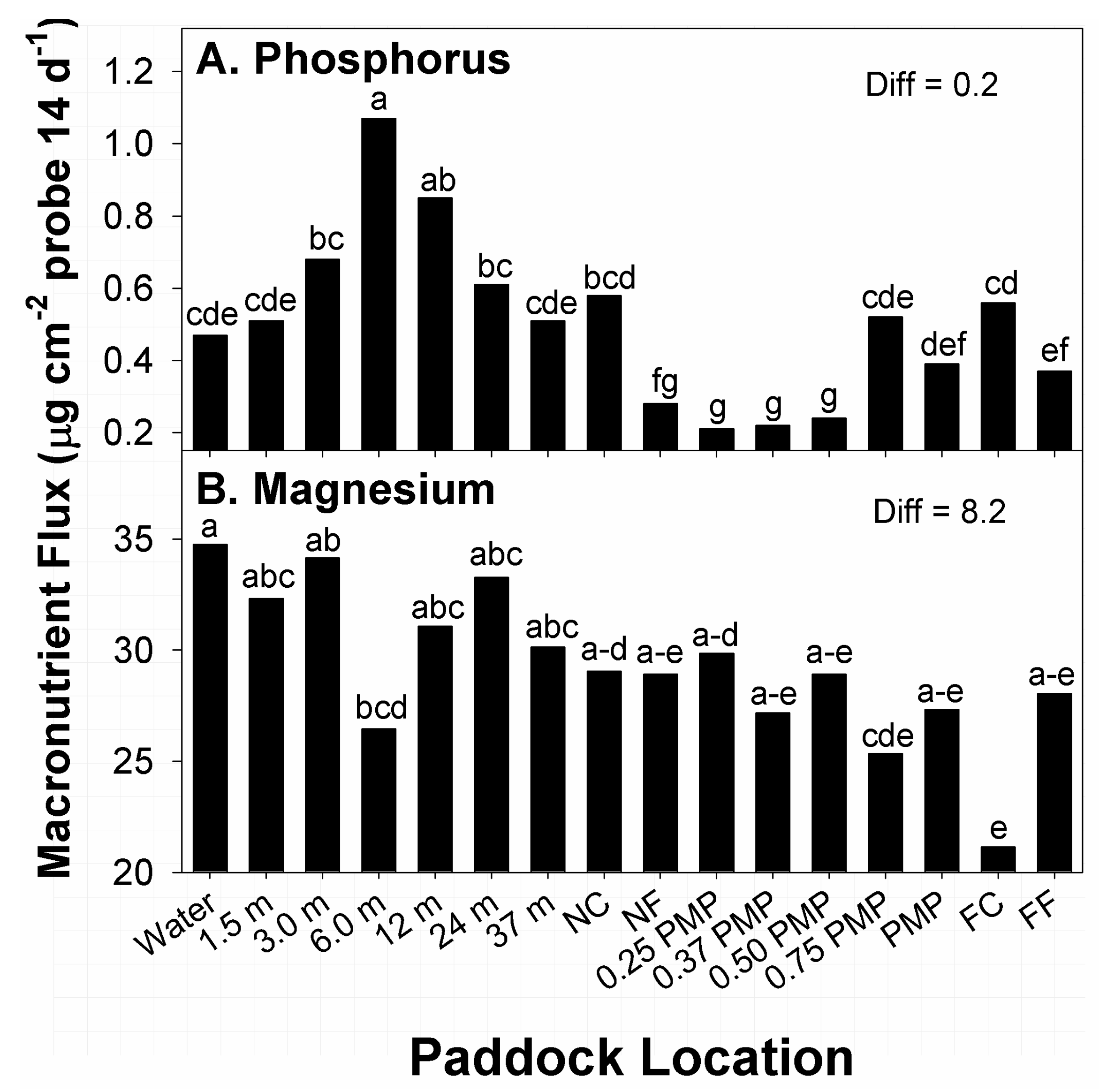
Among effects related to paddock location, the greatest amounts of flux in P were recorded at 6 m from water, while the second-greatest amounts occurred at 12 m from water (
Figure 3A). In contrast, locations with the lowest amounts of P flux were noted at 0.25 PMP through 0.5 PMP. Locations with the second-lowest amounts of P flux occurred at NF and FF. Flux in P at all other locations belonged to different means groups, with intermediate amounts recorded. The potential of the two applied stocking methods to affect distribution of plant-available P within different areas of paddocks (sub-paddocks) was not entirely clear. While there were definite areas with high and low P flux, these zones did not translate to locations normally considered high-traffic areas, such as water sources, corners, or fences [
37,
38]. Cattle could have loafed roughly 6 to 12 m from water in all sampled paddocks and sub-paddocks, but that would have also resulted in higher flux in other neighboring locations.
The primary sources of plant-available P in soils during growing seasons is derived from recycling of P in soil organic matter via microbes, and through inputs and decomposition of livestock feces, which are slow processes [
3,
10]. Small amounts are also derived by weathering of parent materials of soils. In this instance, a likely contributing factor for the lower amounts of flux in available P at the more centrally-located positions in paddocks could be a lack of use of these areas by cattle [
10], or a lack of deposition of excreta in these areas when frequented by cattle. Cattle grazing larger areas of rangeland can travel up to 1.6 km day
−1 while searching for forage, though areas frequented in such travel may not be grazed or used for loafing [
38,
39]. Other research on P distributions within paddocks of grasslands under rotational and continuous stocking noted higher amounts of inorganic P closer to water or shade compared to paddock centers and more distant locations (78–130 [
10], and 8–17 [
40] ppm), or different times of growing seasons (11–27 ppm [
9]), but no effects of stocking method on distributions, as was recorded in the current study.
Among effects on plant available Mg, main effects related to paddock location (F
15, 195 = 1.8;
p = 0.03) and stocking method × time of year interactions (F
1, 194 = 12.2;
p < 0.01) were significant. All other main and interaction effects were not (0.11 <
p < 0.60). Within stocking method × time of year interactions (
Table 6), the largest fluxes in plant-available Mg occurred in response to rotational stocking during March, while the second-largest occurred under continuous stocking during August. The lowest flux in Mg occurred in response to rotational stocking during August.
Differences in mean responses in this interaction varied compared to results of other studies. Studies on both native prairie and tame pasture have reported higher amounts of Mg in soils under rotational than continuous stocking [
3,
13], as was noted in the current study, in addition to changes in distribution patterns related to stocking methods [
37]. Research has also recorded declines in Mg concentrations in soils through leaching under grazing [
3,
39], as in the decline in flux recorded under rotational stocking between March and August. In contrast, the increase in Mg over the growing season under continuous stocking may be related to inputs via feces, which is the primary input source, due to yearlong residency on paddocks [
3,
37]. Soil type can also have some effects on flux in available Mg, with lower amounts recorded in sandier soils. However, the primary sources of plant-available Mg are found in animal excreta, and primarily (>75% of total) in feces [
3], so the longer grazing time afforded by continuous stocking provided a greater opportunity for Mg enrichment. The current study noted higher amounts under rotational stocking in March, but continuous stocked paddocks, dominated by sandier soils, generated similar high fluxes in August.
The greatest amounts of flux in available Mg among paddock locations, across stocking methods, occurred at water sources, with the second-greatest amounts noted at 3 m from water (
Figure 3B). Alternatively, the lowest amounts of flux were recorded at FC locations. Locations with the second-lowest amounts of flux in Mg were noted at 0.75 PMP. Amounts of Mg flux at remaining locations belonged to groups with intermediate amounts, with some mean fluxes belonging to 5 means groups. Therefore, there was a degree of similarity in amounts of Mg flux at different locations along transects. A total of 13 means belonged to the group containing the largest response, while 7 responses belonged to the means group with the lowest amounts of flux. Overall, there was a general decline in Mg flux from water sources to FC, but the degree of variability present in flux among paddock locations makes identifying the presence of hot spots difficult. Research on paddocks of introduced perennial grasses managed by different stocking methods reported greater amounts of Mg in larger zones of paddocks closer to water or shade [
10,
37]. A similar effect was noted in the current study in the lower means of Mg flux with increasing distance from water, though the test for differences noted a degree of similarity between locations near water through FF, which were distances of 423 and 976 m from water.
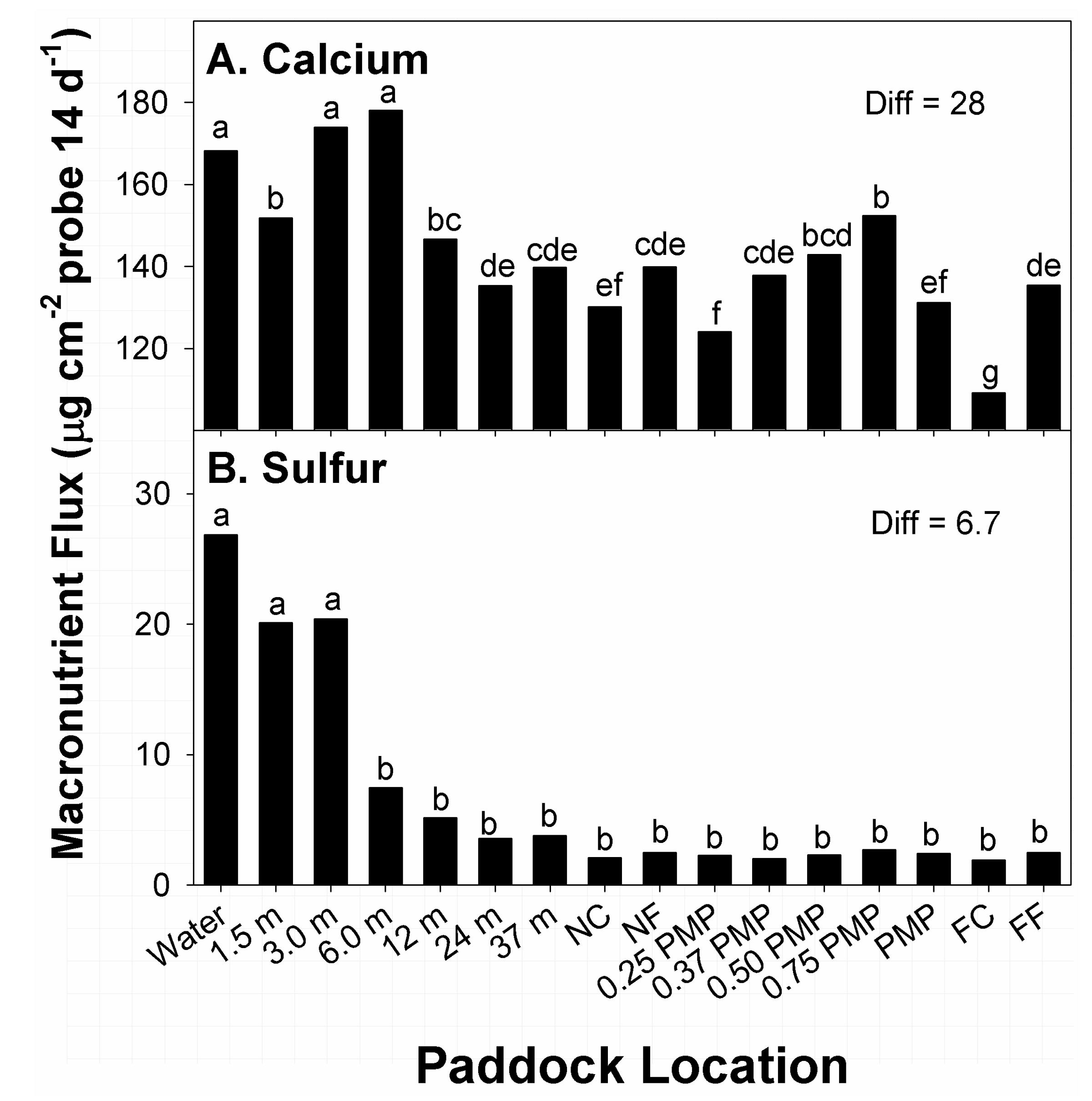
3.4. Calcium and Sulfur
Main effects of time of growing season (F
1, 194 = 155.7;
p < 0.01) and paddock location (F
15, 194 = 2.3;
p < 0.01) on flux in available Ca were significant; all other main and interaction effects were not (0.11 <
p < 0.90). The lack of effects related to stocking methods was unexpected. Earlier research on native prairie in north-central Texas, United States, noted greater amounts of Ca in surface soils of rotational stocked sub-paddocks than in larger, continuously stocked paddocks [
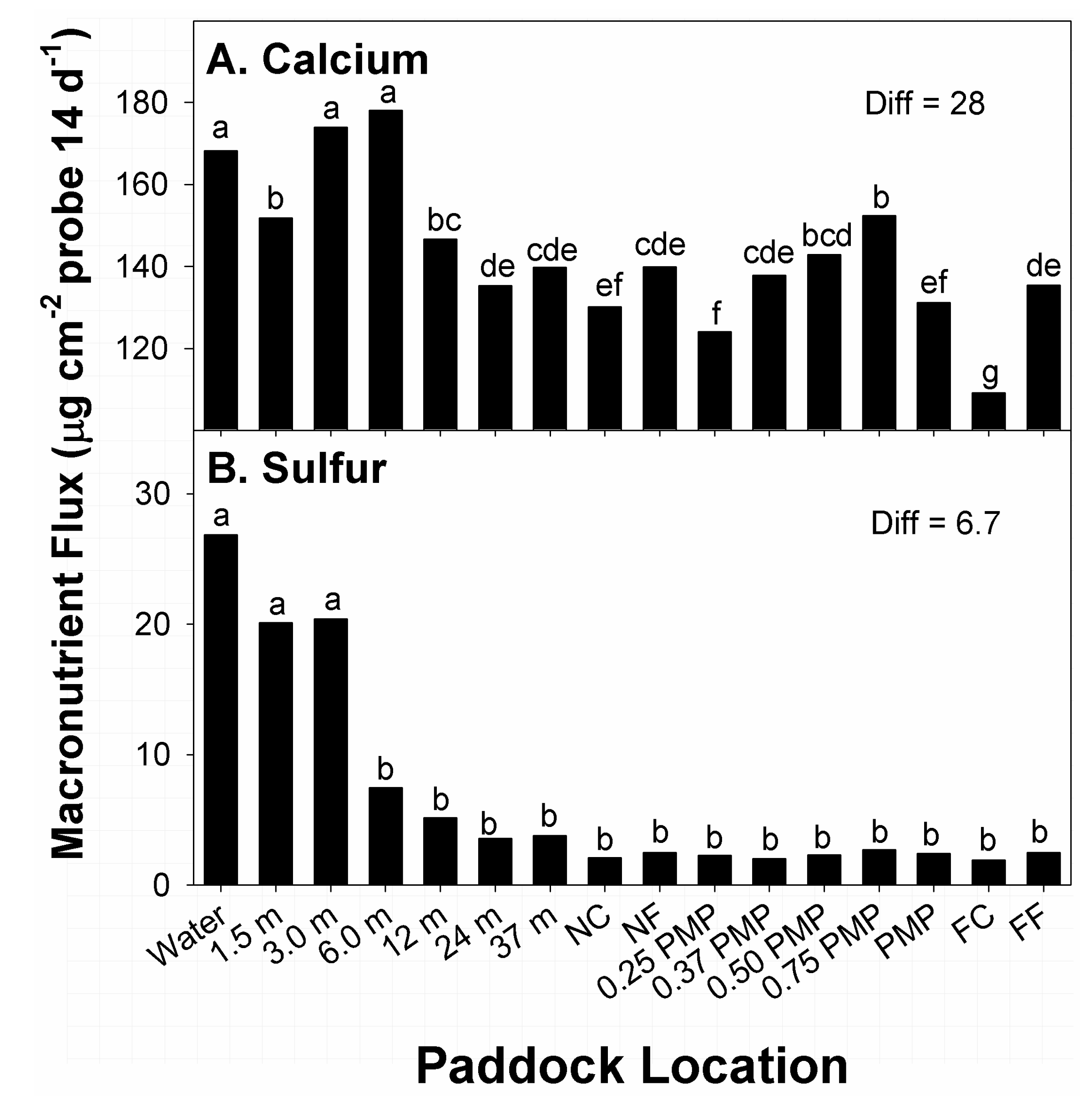
13]. Among times of growing season, higher flux in available Ca was noted at time of sampling during August, compared to March (181 vs. 116 µg Ca cm
−2 probe; Diff = 5.4 µg). Among effects related to paddock location, the greatest amounts of flux in Ca were recorded at water sources 3.0 and 6.0 m from water (
Figure 4A). Locations with the second greatest amounts of Ca flux were noted at 1.5 m from water, and 0.75 PMP. Locations with the lowest and second-lowest amounts of Ca flux were noted at far corners from water sources, and 0.25 PMP, respectively. Flux in Ca at all other locations belonged to different means groups with intermediate amounts recorded.
Calcium in excreta from animals is primarily found in feces, with <3% of animal inputs to soil found in urine. Fecal sources of Ca are also more significant drivers of Ca flux in grassland soils than amounts derived from decomposition of plant residues, or breakdown of parent materials of soils [
3]. However, reports of Ca inputs to grassland soils via cattle excreta varies widely. Further, Ca movement from dung into available pools in soils is not well defined, particularly in native rangelands of the United States. A study on soil responses of native prairie ecosystems in northcentral Texas, United States, reported higher concentrations of Ca in soils of paddocks managed by rotational than continuous stocking, with both concentrations lower than was recorded under no grazing [
13]. While the current study did show greater amounts of Ca flux in close proximity to water across stocking methods, other locations that would be considered high-traffic zones for cattle (i.e., corners and fences) tended to show lower flux, despite the application of stocking methods for 6 years.
Differences in amounts of flux of plant-available Ca during different seasons have been previously reported. Dickinson and Craig [
40] reported Ca in soils increased with amount of precipitation received. However, the current study showed lower amounts of Ca flux during spring, under wetter conditions in conjunction with precipitation events, compared to flux recorded in summer. Precipitation received during individual calendar years in the US SGP, including the study site, is highly variable in amount and timing of precipitation [
41]. Such variability in received precipitation would result in some level of variability of plant-available Ca and other macronutrients. Larger precipitation events than were encountered during this study may be required. As such, Ca movement from dung into soil without such events likely occurs at rates that are similar to amounts derived from organic matter in dung [
42,
43].
The main effects of time of growing season (F
1, 194 = 5.4;
p = 0.02) and paddock location (F
15, 194 = 2.0;
p = 0.02) on flux in available S were significant; all other main and interaction effects were not (0.34 <
p < 0.98). Among times of growing season, higher flux in available S was recorded at time of sampling during August than March (4.8 vs. 3.7 µg S cm
−2 probe; Diff = 0.7 µg). Among effects of paddock location, the greatest amounts of flux in available S were recorded within 3.0 m of water sources (
Figure 4B). Thereafter, amounts of S flux at 6.0 m from water to FC belonged to the same means group and displayed low and consistent amounts of flux. Flux of available S at these locations ranged from 6 to 28% of amounts recorded closer to water sources.
The majority (>90%) of plant-available S within the uppermost sections of soil profiles in grazed temperate grasslands are present in labile organic forms derived from animal excreta and plant residues [
44]. Inputs of plant-available S from grazing animals is related to relationships between retention and throughput of consumed forage in animals. Amounts of S in urine and feces varies with type of grassland, location within landscape where deposited, and form of management. Research in Australia and New Zealand reported 50 to 70% of S excreted by cattle was labile forms of sulfate in urine [
44,
45]. Alternatively, S concentration in dung tends to be small (~0.3% of dung dry matter) and generally in organic forms that mineralize at slow rates, similar to soil organic matter [
3,
44].
Cattle retain roughly 25% of S within biomass of consumed forage, with the remainder excreted, primarily in urine [
3]. The current study showed redistribution of S within grazed paddocks was limited, and largely occurred in proximity (≤3.0 m) to water, compared to other locations. Further, other high traffic areas (corners and fences) showed low amounts of flux compared to water sources. In comparison, research in Florida, United States, [
10] reported no significant effects of stocking method or paddock locations related to S distribution within grazed bermudagrass paddocks.
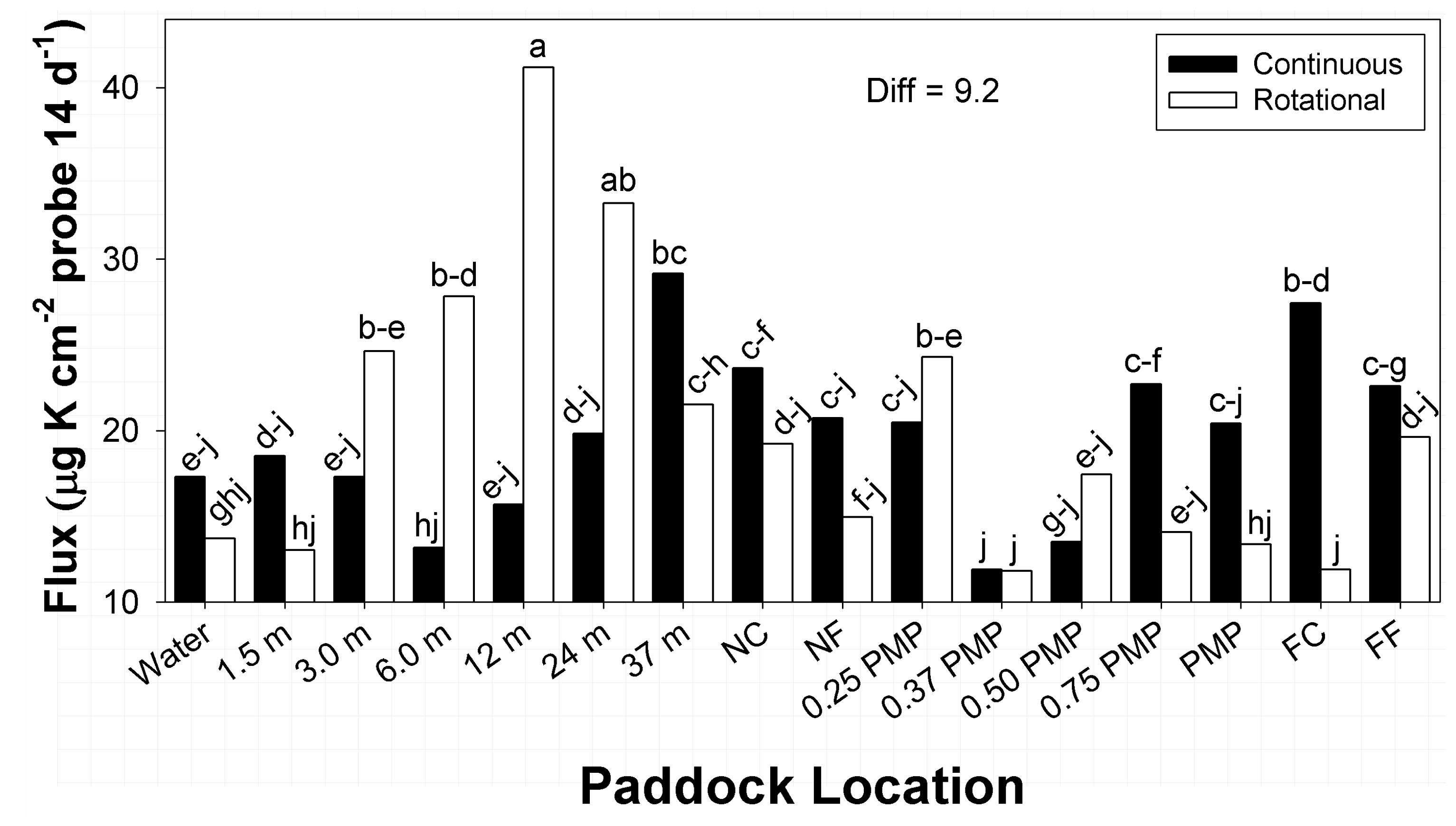
3.5. Potassium
There were significant differences in main effects related to time of year (
F1, 194 = 37.4;
p < 0.01) and soil depth (F
1, 194 = 66.9;
p < 0.01) on plant-available K, as was the stocking method × paddock location interaction (F
15, 194 = 2.3;
p = 0.01). All other main and interaction effects were not significant (0.14 <
p < 0.96). Greater amounts of flux were noted during March than were recorded in August (24.4 vs. 14.7 μg K cm
−2 probe; Diff = 7.4 µg). Among soil depths, greater amounts of flux were noted within the upper 7.5 cm depth of profile than the 7.5 to 15 cm increment (26.1 vs. 13.7 μg K cm
−2 probe; Diff = 7.4 µg). Main effects of time of year and soil depth were similar to results reported in the broader literature, with greater amounts of K flux noted during periods with greater amounts soil water and in sections of soil profiles near the surface [
3,
15]. Sampling during March of the current study occurred after a series of precipitation events that allowed moisture in the upper sections of soil profile to approximate field capacity. Such conditions are important in movement of K from plant residues and excreta into soil solution [
3]; drought conditions prior to sampling in August would limit such movement, and reduce amounts of plant-available K in soils.
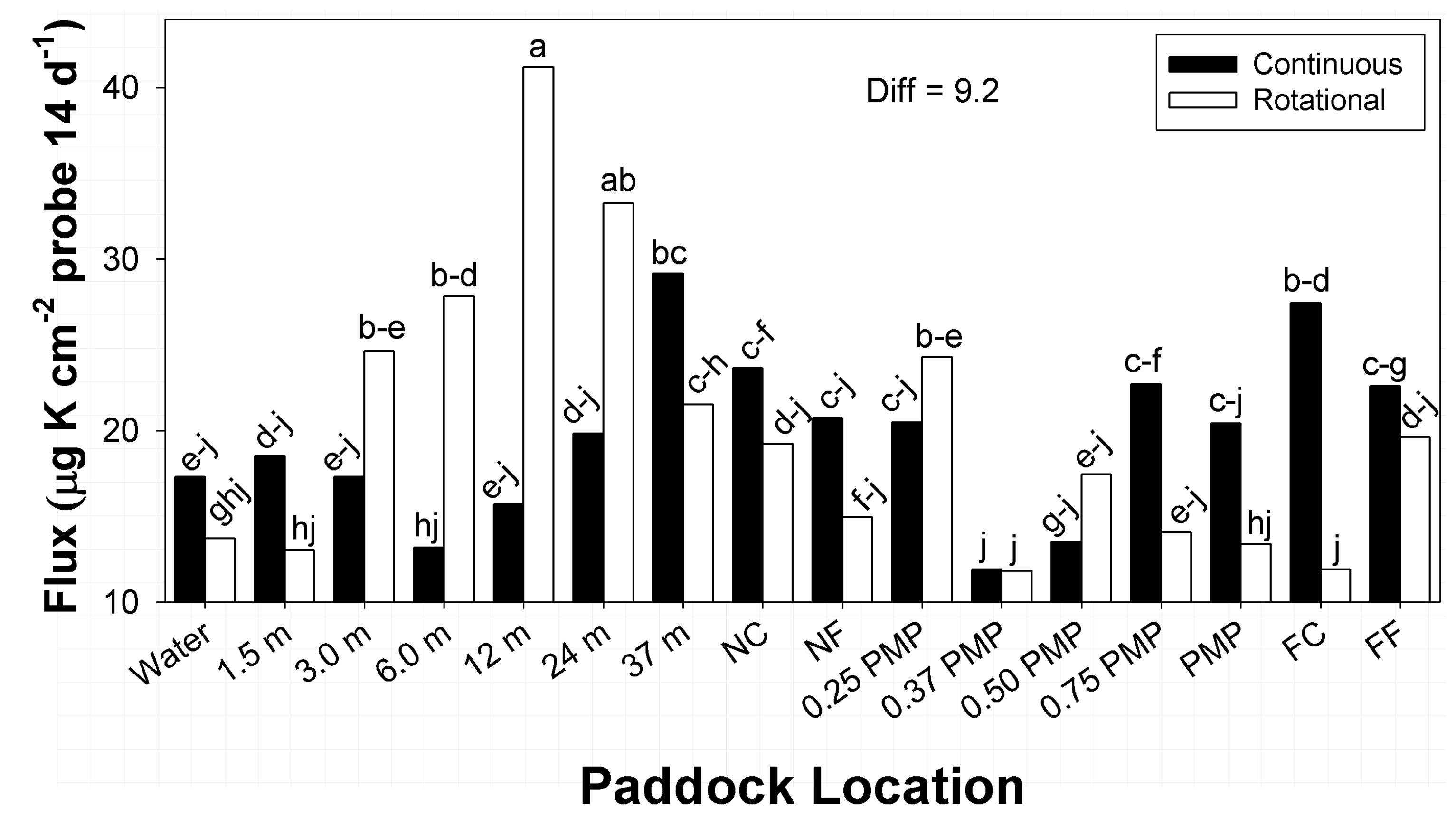
Within the stocking method × paddock location interaction in flux of available K, the greatest and second greatest amounts occurred, respectively, under rotational stocking at 12 and 24 m from water sources (
Figure 5). Amounts at 37 m from water in continuous stocked paddocks had some similarity to amounts at 24 m locations in rotational stocked paddocks. In contrast, the lowest flux in available K were recorded at 0.37 PMP under both stocking methods, and FC in rotational stocked paddocks. All remaining means within the interaction belonged to ranges of means groups, with many values belonging to 7 means groups within the interaction. Amounts of flux in plant available K under both stocking methods displayed undulations among low and high amounts with increasing distance from water, though at different locations.
The interaction between stocking method and paddock location on K flux (
Figure 5) was the only occurrence of stocking method affecting the distribution of a macronutrient within paddock space. However, this impact is not clear and displayed a degree of variability, as was recorded for most of the macronutrients. Potassium is highly labile and can be easily leached from excreta and grassland soils, particularly with occurrences of high rainfall [
3], which are uncommon in the US SGP [
41]. Cattle excreta (particularly urine) is a primary source of plant-available K, and amounts in urine patches tend to exceed uptake requirements of plants [
3,
35]. Therefore, amounts of K flux could be related to some combination of conservation of K in soils by drought and local distribution of urine patches, though many of these locations did not translate to areas normally noted for high animal use [
37,
38]. Effects related to stocking method and paddock location on K flux showed both similarities and differences from results of other studies. Experiments testing distributions of macronutrients within paddocks of bahiagrass reported no difference among rotational and continuous stocking for soil K, but higher concentrations of K in zones near water and shade [
37]. In comparison, experiments on stocking methods applied to bermudagrass paddocks also reported no difference in K concentrations in soils among stocking methods [
10]. Similarly, K concentrations in soils of native prairie in northcentral Texas, United States, reported no differences among continuous and rotational stocking [
13]. In contrast, amounts of flux in available K in the current study showed differences among stocking methods that occurred as undulations across paddocks and sub-paddocks.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}