Effect of a Late Spring Application of Hydrogen Cyanamide on High-Chill Peaches


Abstract
1. Introduction
2. Materials and Methods
2.1. Peach Materials and Spray Treatment
2.2. Data Collection.
2.3. Statistical Analysis
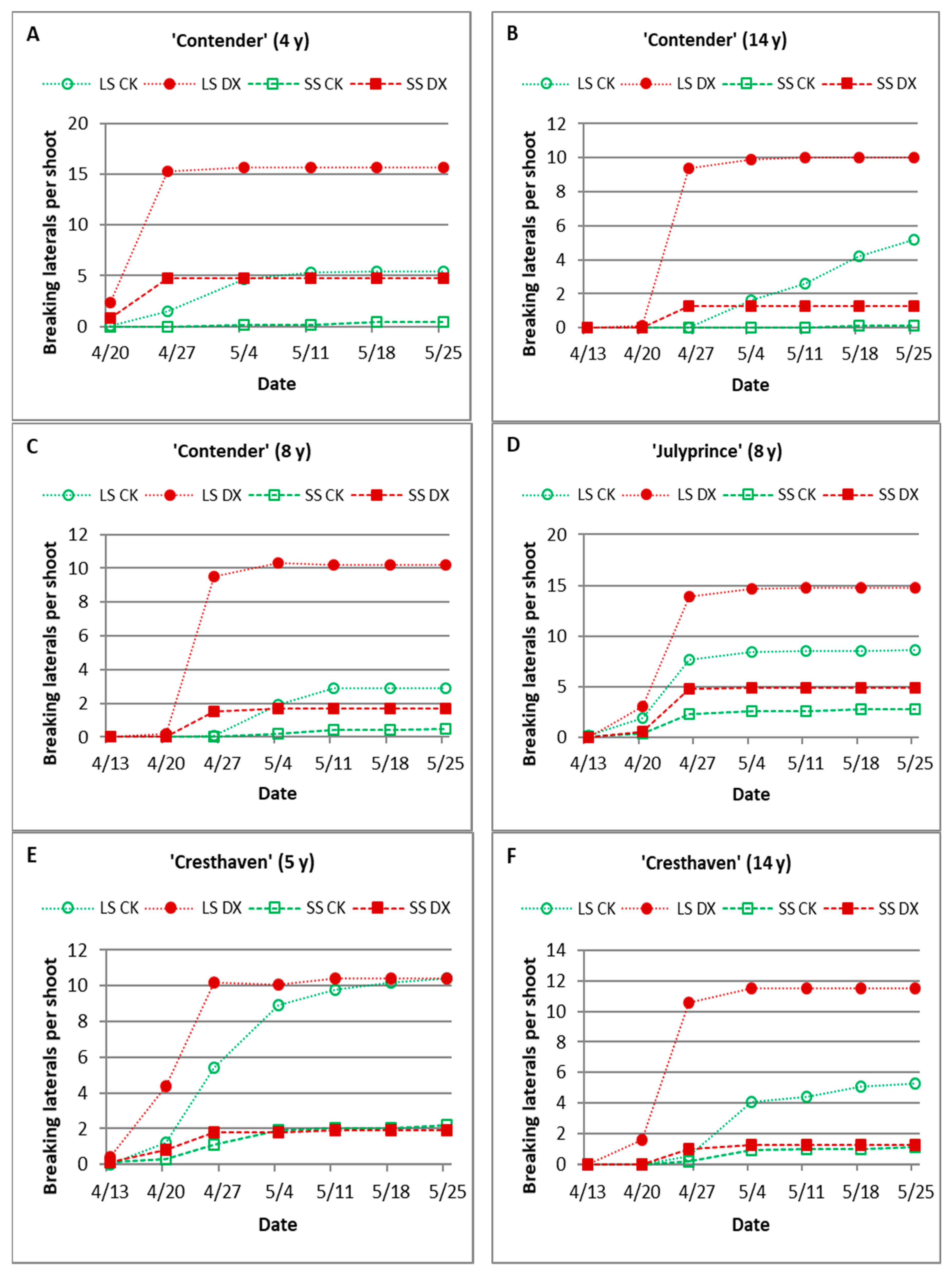
3. Results
3.1. Effect of Dormex on Budbreak
3.2. Effect of Dormex on Year-End Vegetative Growth
3.3. Effects of Dormex on Fruit Number and Yield in Following Season
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weinberger, J.H. Chilling requirement of peach varieties. Proc. Am. Soc. Hortic. Sci. 1950, 56, 122–128. [Google Scholar]
- Dozier, W.A.; Powell, A.A.; Caylor, A.W.; Mcdaniel, N.R.; Carden, E.L.; Mcguire, J.A. Hydrogen cyanamide induces budbreak of peaches and nectarines following inadequate chilling. HortScience 1990, 25, 1573–1575. [Google Scholar] [CrossRef]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for ‘Redhaven’ and ‘Elberta’ peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Anderson, J.L.; Richardson, E.A. The Utah chill unit/flower bud phenology models for deciduous fruit: Their implication for production in subtropical areas. Acta Hortic. 1987, 199, 45–50. [Google Scholar] [CrossRef]
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The dynamic model for rest completion in peach buds. Acta Hortic. 1990, 276, 165–174. [Google Scholar] [CrossRef]
- Mayles, K.M.; Kilby, M.W. Chemical regulation of budbreak of flame seedless table grapes following dormant applications of hydrogen cyanamide in a low desert climate. HortScience 1987, 22, 1039. [Google Scholar]
- Pontikis, C.A. Effects of hydrogen cyanamide on bloom advancement in female pistachio (P. vera L). Fruit Var. J. 1989, 43, 125–128. [Google Scholar]
- Wood, B.W. Hydrogen cyanamide advances pecan budbreak and harvesting. J. Am. Soc. Hortic. Sci. 1993, 118, 690–693. [Google Scholar] [CrossRef]
- Williamson, J.G.; Maust, B.E.; NeSmith, D.S. Timing and concentration of hydrogen cyanamide affect blueberry bud development and flower mortality. HortScience 2001, 36, 922–924. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, L.; Ma, C.; Xu, W.P.; Liu, Z.R.; Zhang, C.X.; Matthew, W.D.; Wang, S.P. Impact of chilling accumulation and hydrogen cyanamide on floral organ development of sweet cherry in a warm region. J. Integr. Agric. 2016, 15, 2529–2538. [Google Scholar] [CrossRef]
- George, A.P.; Lloyd, J.; Nissen, R.J. Effects of hydrogen cyanamide, paclobutrazol and pruning date on dormancy release of the low chill peach cultivar Flordaprince in subtropical Australia. Aust. J. Exp. Agric. 1992, 32, 89–95. [Google Scholar] [CrossRef]
- Sillercepeda, J.H.; Fuchigami, L.H.; Chen, T.H.H. Hydrogen cyanamide-Induced budbreak and phytotoxicity in Redhaven peach buds. HortScience 1992, 27, 874–876. [Google Scholar] [CrossRef]
- Fan, S.; Bielenberg, D.G.; Zhebentyayeva, T.N.; Reighard, G.L.; Okie, W.R.; Holland, D.; Abbott, A.G. Mapping quantitative trait loci associated with chilling requirement, heat requirement and bloom date in peach (Prunus persica). New Phytol. 2010, 185, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H. Regulation of bud dormancy and bud break in Japanese apricot (Prunus mume Siebold & Zucc.) and Peach [Prunus persica (L.) Batsch]: A summary of recent studies. J. Jpn. Soc. Hortic. Sci. 2014, 83, 187–202. [Google Scholar]
- Li, Z.G.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated MADS genes from the EVG locus of peach [Prunus persica (L.) Batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| LB | FB | |||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 4/13 | 5/25 | Ratio | 4/13 | 5/25 | Ratio | |||||||||||||
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Total | SE | |||||||
| TT | ||||||||||||||||||
| DX | 9.11 | 0.40 | a | 7.00 | 0.41 | a | 0.71 | 0.04 | a | 14.21 | 0.54 | a | 1.20 | 0.11 | a | 0.10 | 0.01 | A |
| CK | 7.56 | 0.36 | b | 3.51 | 0.27 | b | 0.43 | 0.03 | b | 13.66 | 0.49 | a | 0.66 | 0.08 | b | 0.06 | 0.01 | B |
| ST | ||||||||||||||||||
| LS | 12.55 | 0.34 | a | 8.57 | 0.38 | a | 0.70 | 0.03 | a | 17.45 | 0.56 | a | 1.18 | 0.11 | a | 0.08 | 0.01 | A |
| SS | 4.13 | 0.16 | b | 1.86 | 0.15 | b | 0.44 | 0.04 | b | 10.43 | 0.33 | b | 0.68 | 0.08 | b | 0.09 | 0.01 | A |
| CV | ||||||||||||||||||
| BB | 9.75 | 0.82 | ab | 4.23 | 0.84 | bc | 0.37 | 0.05 | bc | 16.13 | 0.86 | b | 0.93 | 0.19 | bc | 0.06 | 0.01 | B |
| CO | 6.81 | 0.48 | cd | 4.85 | 0.48 | b | 0.62 | 0.06 | ab | 12.78 | 0.81 | cd | 0.93 | 0.14 | bc | 0.11 | 0.02 | Ab |
| CH | 6.90 | 0.56 | cd | 5.14 | 0.60 | b | 0.63 | 0.05 | ab | 15.53 | 0.73 | bc | 1.39 | 0.19 | ab | 0.11 | 0.02 | Ab |
| EL | 9.85 | 1.20 | ab | 4.50 | 0.96 | b | 0.70 | 0.20 | a | 9.18 | 0.72 | e | 0.18 | 0.07 | c | 0.02 | 0.01 | B |
| JP | 10.35 | 0.86 | a | 7.78 | 0.89 | a | 0.65 | 0.05 | ab | 13.50 | 0.97 | bcd | 1.83 | 0.31 | a | 0.18 | 0.05 | A |
| JN | 8.03 | 0.73 | bc | 5.93 | 0.76 | ab | 0.71 | 0.05 | a | 12.48 | 0.76 | d | 0.83 | 0.21 | bc | 0.06 | 0.01 | B |
| RB | 5.33 | 0.77 | d | 2.10 | 0.51 | c | 0.29 | 0.06 | c | 6.78 | 0.39 | e | 0.15 | 0.06 | c | 0.02 | 0.01 | B |
| RH | 11.85 | 1.18 | a | 7.90 | 1.20 | a | 0.59 | 0.06 | abc | 14.78 | 1.04 | bcd | 0.55 | 0.12 | bc | 0.04 | 0.01 | B |
| TQ | 10.65 | 0.94 | a | 5.18 | 0.81 | b | 0.43 | 0.06 | abc | 25.00 | 1.36 | a | 1.15 | 0.29 | ab | 0.05 | 0.01 | B |
| Age | ||||||||||||||||||
| 4 | 10.40 | 0.65 | a | 6.49 | 0.64 | ab | 0.64 | 0.07 | a | 9.22 | 0.60 | d | 0.58 | 0.10 | b | 0.10 | 0.02 | B |
| 5 | 7.03 | 0.46 | bc | 4.75 | 0.45 | c | 0.57 | 0.04 | a | 11.15 | 0.53 | d | 0.89 | 0.14 | b | 0.08 | 0.01 | B |
| 7 | 10.35 | 0.86 | a | 7.78 | 0.89 | a | 0.65 | 0.05 | a | 13.50 | 0.97 | c | 1.83 | 0.31 | a | 0.18 | 0.05 | A |
| 8 | 7.63 | 0.57 | b | 4.03 | 0.54 | c | 0.47 | 0.04 | a | 16.75 | 0.73 | b | 0.95 | 0.17 | b | 0.05 | 0.01 | B |
| 10 | 10.65 | 0.94 | a | 5.18 | 0.81 | bc | 0.43 | 0.06 | a | 25.00 | 1.36 | a | 1.15 | 0.29 | b | 0.05 | 0.01 | B |
| 14 | 5.75 | 0.50 | c | 4.10 | 0.54 | c | 0.59 | 0.07 | a | 17.06 | 0.76 | b | 0.95 | 0.15 | b | 0.07 | 0.01 | B |
| Shoot Type | Treatment | Terminal Bud Class | 4/13 | 4/20 | 4/27 | 5/4 | 5/11 | 5/18 | 5/25 |
|---|---|---|---|---|---|---|---|---|---|
| LS | DX | N | 83.33 | 63.33 | 20.83 | 15.83 | 7.50 | 3.33 | 0.83 |
| Y | 10.83 | 28.33 | 70.00 | 70.00 | 70.83 | 65.83 | 65.83 | ||
| D | 0.00 | 2.50 | 3.33 | 8.33 | 15.83 | 25.00 | 27.50 | ||
| B | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | ||
| CK | N | 84.17 | 74.17 | 48.33 | 17.50 | 10.00 | 5.00 | 3.33 | |
| Y | 7.50 | 16.67 | 37.50 | 63.33 | 68.33 | 71.67 | 73.33 | ||
| D | 2.50 | 3.33 | 8.33 | 13.33 | 15.83 | 17.50 | 17.50 | ||
| B | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | 5.83 | ||
| SS | DX | N | 91.67 | 70.83 | 23.33 | 14.17 | 5.00 | 2.50 | 2.50 |
| Y | 7.50 | 25.83 | 69.17 | 70.83 | 70.00 | 66.67 | 65.83 | ||
| D | 0.00 | 2.50 | 6.67 | 14.17 | 24.17 | 30.00 | 30.83 | ||
| B | 0.83 | 0.83 | 0.83 | 0.83 | 0.83 | 0.83 | 0.83 | ||
| CK | N | 90.00 | 80.00 | 46.67 | 17.50 | 11.67 | 8.33 | 3.33 | |
| Y | 5.00 | 13.33 | 44.17 | 72.50 | 75.83 | 78.33 | 81.67 | ||
| D | 1.67 | 3.33 | 5.83 | 6.67 | 9.17 | 10.00 | 11.67 | ||
| B | 3.33 | 3.33 | 3.33 | 3.33 | 3.33 | 3.33 | 3.33 |
| Shoots | Fruit | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Count | Length (cm) | Number | Yield (kg) | |||||||||
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | |||||
| TT | ||||||||||||
| DX | 1.90 | 0.20 | a | 9.51 | 0.94 | a | 91.00 | 10.33 | a | 16.75 | 1.48 | a |
| CK | 2.31 | 0.20 | a | 8.08 | 0.70 | a | 115.31 | 19.83 | a | 16.90 | 2.76 | a |
| ST | ||||||||||||
| LS | 2.01 | 0.19 | a | 9.09 | 0.79 | a | - | - | ||||
| SS | 2.23 | 0.21 | a | 8.4 | 0.86 | a | - | - | ||||
| CV | ||||||||||||
| BB | 2.13 | 0.45 | a | 7.36 | 2.8 | b | - | - | ||||
| CO | 2.23 | 0.28 | a | 8.39 | 0.87 | b | 134.55 | 10.47 | a | 20.41 | 1.59 | ab |
| CH | 1.40 | 0.25 | a | 3.71 | 0.73 | b | 31.50 | 14.20 | c | 8.66 | 3.37 | ab |
| EL | 2.89 | 0.63 | a | 8.63 | 1.54 | b | - | - | ||||
| JP | 1.81 | 0.39 | a | 8.77 | 2.13 | b | 132.75 | 33.51 | ab | 26.88 | 6.26 | a |
| JN | 1.43 | 0.40 | a | 7.43 | 2.05 | b | 38.75 | 29.65 | bc | 3.22 | 2.30 | b |
| RB | 1.93 | 0.56 | a | 10.58 | 2.14 | b | - | - | ||||
| RH | 2.70 | 0.53 | a | 9.85 | 1.66 | b | - | - | ||||
| TQ | 2.71 | 0.49 | a | 18.83 | 3.00 | a | 150.00 | 13.74 | a | 20.71 | 1.52 | ab |
| Age | ||||||||||||
| 4 | 2.74 | 0.32 | a | 8.85 | 0.89 | b | 134.50 | 22.19 | a | 22.16 | 3.89 | ab |
| 5 | 1.59 | 0.28 | a | 7.69 | 1.16 | b | 43.71 | 22.64 | b | 7.06 | 3.83 | ab |
| 7 | 1.81 | 0.39 | a | 8.77 | 2.13 | b | 132.75 | 33.51 | a | 26.88 | 6.26 | a |
| 8 | 1.71 | 0.28 | a | 6.97 | 1.53 | b | 80.71 | 5.03 | b | 13.50 | 1.12 | b |
| 10 | 2.71 | 0.49 | a | 18.83 | 3.00 | a | 150.00 | 13.74 | a | 20.71 | 1.52 | ab |
| 14 | 2.13 | 0.33 | a | 6.97 | 1.01 | b | 136.14 | 11.37 | a | 22.79 | 1.30 | a |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.; Beckman, T.G. Effect of a Late Spring Application of Hydrogen Cyanamide on High-Chill Peaches. Agronomy 2019, 9, 726. https://doi.org/10.3390/agronomy9110726
Chen C, Beckman TG. Effect of a Late Spring Application of Hydrogen Cyanamide on High-Chill Peaches. Agronomy. 2019; 9(11):726. https://doi.org/10.3390/agronomy9110726
Chicago/Turabian StyleChen, Chunxian, and Thomas G. Beckman. 2019. "Effect of a Late Spring Application of Hydrogen Cyanamide on High-Chill Peaches" Agronomy 9, no. 11: 726. https://doi.org/10.3390/agronomy9110726
APA StyleChen, C., & Beckman, T. G. (2019). Effect of a Late Spring Application of Hydrogen Cyanamide on High-Chill Peaches. Agronomy, 9(11), 726. https://doi.org/10.3390/agronomy9110726