Genetic Analysis and Gene Mapping for a Short-Petiole Mutant in Soybean (Glycine max (L.) Merr.)




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Genetic Analysis of dsp
2.3. DNA Extraction and SSR Markers Analysis
2.4. Bulked Segregant Analysis (BSA) and Target Gene Mapping of the dsp Mutant
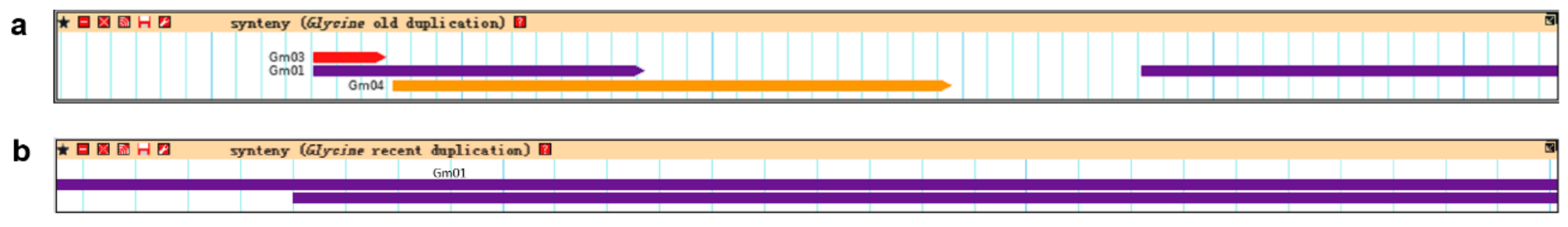
2.5. Synteny Information and Homologous Protein Information Retrieval
3. Results and Discussion
3.1. Characteristics of Short-Petiole Trait in the dsp Mutant
3.2. Genetic Analysis of Petiole Length in the dsp Mutant
3.3. Mapping Genes dsp1 and dsp2 with SSR Markers
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Chavarria, G.; Caverzan, A.; Müller, M.; Rakocevic, M. Soybean architecture plants: From solar radiation interception to crop protection. In Soybean-The Basis of Yield, Biomass and Productivity; InTechOpen: London, UK; p. 15.
- Mwanamwenge, J.; Siddique, K.H.M.; Sedgley, R.H. Canopy development and light absorption of grain legume species in a short season mediterranean-type environment. J. Agron. Crop Sci. 1997, 179, 1–7. [Google Scholar] [CrossRef]
- Isoda, A.; Mori, M.; Matsumoto, S.; Li, Z.; Wang, P. High yielding performance of soybean in Northern Xinjiang, China. Plant Prod. Sci. 2015, 9, 401–407. [Google Scholar] [CrossRef]
- Liu, X.; Jin, J.; Wang, G.; Herbert, S.J. Soybean yield physiology and development of high-yielding practices in Northeast China. Field Crop Res. 2008, 105, 157–171. [Google Scholar] [CrossRef]
- Muraoka, H.; Takenaka, A.; Tang, Y.; Koizumi, H.; Washitani, I. Flexible leaf orientations of Arisaema heterophyllum maximize light capture in a forest understorey and avoid excess irradiance at a deforested site. Ann. Bot. 1998, 82, 297–307. [Google Scholar] [CrossRef]
- Junior, C.P.; Kawakami, J.; Bridi, M.; Conte, M.V.D.; Michalovicz, L. Phenological and quantitative plant development changes in soybean cultivars caused by sowing date and their relation to yield. Afr. J. Agric. Res. 2015, 10, 515–523. [Google Scholar] [CrossRef]
- Kokubun, M. Design and evaluation of soybean ideotypes. Bull. Tohoku Natl. Agric. Exp. Stn. 1988, 77, 77–142. [Google Scholar]
- Niinemets, Ü.; Fleck, S. Petiole mechanics, leaf inclination, morphology and investment in support in relation to light availability in the canopy of Liriodendron tulipifera. Oecologia 2002, 132, 21–33. [Google Scholar] [CrossRef]
- Thomas, J.F.; Raper, C.D. Internode and petiole elongation of soybean in response to photoperiod and end-of-day light quality. Bot. Gaz. 1985, 146, 495–500. [Google Scholar] [CrossRef]
- Jun, T.H.; Kang, S.T.; Moon, J.K.; Seo, M.J.; Yun, H.T.; Lee, S.K.; Lee, Y.H.; Kim, S.J. Genetic analysis of new short petiole gene in soybean. J. Crop Sci. Biotechnol. 2009, 12, 87–89. [Google Scholar] [CrossRef]
- Kilen, T. Inheritance of a short petiole trait soybean. Crop Sci. 1983, 23, 1208–1210. [Google Scholar] [CrossRef]
- You, M.; Zhao, T.; Gai, J.; Yen, Y. Genetic analysis of short petiole and abnormal pulvinus in soybean. Euphytica 1998, 102, 329–333. [Google Scholar] [CrossRef]
- Jun, T.H.; Kang, S.T. Genetic map of lps3: A new short petiole gene in soybeans. Genome 2012, 55, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Cary, T.R.; Nickell, C.D. Genetic analysis of a short petiole-type soybean, LN89-3502TP. J. Hered. 1999, 90, 300–301. [Google Scholar] [CrossRef]
- Pearson, K.X. On the criterion that a given system of deviations from the probable in the case of a correlated system of variables is such that it can be reasonably supposed to have arisen from random sampling. Philos. Mag. 1900, 50, 157–175. [Google Scholar] [CrossRef]
- Yates, F. Contingency tables involving small numbers and the χ2 test. J. R. Stat. Soc. 1934, 1, 217–235. [Google Scholar]
- Michelmore, R.W.; Paran, I.; Kesseli, R.V. Identification of markers linked to disease-resistance genes by bulked segregant analysis: A rapid method to detect markers in specific genomic regions by using segregating populations. Proc. Natl. Acad. Sci. USA 1991, 88, 9828–9832. [Google Scholar] [CrossRef]
- Lander, E.S.; Green, P.; Abrahamson, J.; Barlow, A.; Daly, M.J.; Lincoln, S.E.; Newburg, L. MAPMAKER: An interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics 1987, 1, 174–181. [Google Scholar] [CrossRef]
- Kosambi, D.D. The estimation of ma distance from recombination values. Ann. Eugen. 1944, 12, 172–175. [Google Scholar] [CrossRef]
- Song, Q.J.; Marek, L.F.; Shoemaker, R.C.; Lark, K.G.; Concibido, V.C.; Delannay, X.; Specht, J.E.; Cregan, P.B. A new integrated genetic linkage map of the soybean. Theor. Appl. Genet. 2004, 109, 122–128. [Google Scholar] [CrossRef]
- Grant, D.; Nelson, R.T.; Cannon, S.B.; Shoemaker, R.C. SoyBase, the USDA-ARS soybean genetics and genomics database. Nucleic Acids Res. 2010, 38, D843–D846. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Jia, G.; Zhu, Y.; Grant, D.; Nelson, R.T.; Hwang, E.Y.; Hyten, D.L.; Cregan, P.B. Abundance of SSR motifs and development of candidate polymorphic SSR markers (BARCSOYSSR_1.0) in soybean. Crop Sci. 2010, 50, 1950–1960. [Google Scholar] [CrossRef]
- Chan, C.; Qi, X.; Li, M.W.; Wong, F.L.; Lam, H.M. Recent developments of genomic research in soybean. J. Genet. Genom. 2012, 39, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Schlueter, J.A.; Lin, J.Y.; Schlueter, S.D.; Vasylenko-Sanders, I.F.; Deshpande, S.; Yi, J.; O’bleness, M.; Roe, B.A.; Nelson, R.T.; Scheffler, B.E.; et al. Gene duplication and paleopolyploidy in soybean and the implications for whole genome sequencing. BMC Genom. 2007, 8, 330. [Google Scholar] [CrossRef]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.; Thelen, J.J.; Cheng, J.; et al. Genome sequence of the palaeopolyploid soybean. Nature 2010, 463, 178. [Google Scholar] [CrossRef]
- Shoemaker, R.C.; Schluete, J.; Doyle, J.J. Paleopolyploidy and gene duplication in soybean and other legumes. Curr. Opin. Plant Biol. 2006, 9, 104–109. [Google Scholar] [CrossRef]
- Innan, H.; Kondrashov, F. The evolution of gene duplications: Classifying and distinguishing between models. Nat. Rev. Genet. 2010, 11, 97–108. [Google Scholar] [CrossRef]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef]
- Tian, Z.; Wang, X.; Lee, R.; Li, Y.; Specht, J.E.; Nelson, R.L.; McClean, P.E.; Qiu, L.; Ma, J. Artificial selection for determinate growth habit in soybean. Proc. Natl. Acad. Sci. USA 2010, 107, 8563–8568. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, W.; Zhang, Y.; Liu, M.; Kong, J.; Yu, Z.; Jaffer, A.M.; Gai, J.; Zhao, T. Identification of two duplicated loci controlling a disease-like rugose leaf phenotype in soybean. Crop Sci. 2016, 56, 1611. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population | Year | No. of Wild-type Plants | No. of Mutant-type Plants | Total Number | Expected Ratio | χ2 | p-Value |
|---|---|---|---|---|---|---|---|
| F2(HDS-1 × dsp) | 2014 | 681 | 22 | 703 | 15:1 | 11.16 | 0.00 |
| F2(BW-2 × dsp) | 2014 | 230 | 8 | 238 | 15:1 | 2.91 | 0.09 |
| No. of Segregating Lines | No. of Non-segregating Lines | Total Number | Expected Ratio | χ2 | p-Value | ||
| F2:3(HDS-1 × dsp) | 2015 | 87 | 97 | 184 | 8:7 | 2.47 | 0.12 |
| F3:4(HDS-1 × dsp)-Dsp1_ dsp2dsp2 | 2016 | 54 | 27 | 81 | 2:1 | 0.01 | 0.91 |
| F3:4(HDS-1 × dsp)-dsp1dsp1 Dsp2_ | 2016 | 59 | 32 | 91 | 2:1 | 0.07 | 0.80 |
| Glyma.Wm82. a1.v1.1 Gene Models | Glyma.Wm82. a2.v1 Gene Models | Description |
|---|---|---|
| Glyma07g16731 | Glyma.07g139000 | Mitochondrial carrier protein |
| Glyma07g16740 | Glyma.07g139100 | Major Facilitator Superfamily |
| Glyma07g16750 | Glyma.07g139200 | Uncharacterized protein |
| Glyma07g16760 | Glyma.07g139300 | Uncharacterized protein |
| Glyma07g16770 | Glyma.07g139400 | Probable lipid transfer |
| Glyma07g16790 | Glyma.07g139500 | PRONE (Plant-specific Rop nucleotide exchanger) |
| Glyma07g16800 | Glyma.07g139600 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16810 | Glyma.07g139700 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16830 | Glyma.07g139800 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16840 | Glyma.07g139900 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16850 | Glyma.07g140000 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16860 | Glyma.07g140200 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16876 | Glyma.07g140300 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16893 | - | Cytochrome P450 CYP4/CYP19/CYP26 subfamilies |
| Glyma07g16910 | Glyma.07g140400 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16925 | Glyma.07g140500 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16940 | Glyma.07g140700 | Glutathione S-transferase, C-terminal domain |
| Glyma07g16950 | Glyma.07g140800 | Universal stress protein family |
| Glyma07g16970 | Glyma.07g141000 | Uncharacterized protein |
| Glyma07g16980 | Glyma.07g141100 | Myb-like DNA-binding domain |
| Glyma07g16990 | Glyma.07g141200 | Ctr copper transporter family |
| Glyma07g17000 | Glyma.07g141300 | Probable lipid transfer |
| Glyma07g17010 | Glyma.07g141400 | Non-specific serine/threonine protein kinase. |
| Glyma07g17030 | Glyma.07g141500 | Probable lipid transfer |
| Glyma07g17060 | Glyma.07g141600 | Ctr copper transporter family |
| Glyma07g17080 | Glyma.07g141700 | Hsp20/alpha crystallin family |
| Glyma07g17090 | Glyma.07g141800 | Uncharacterized protein |
| Glyma07g17101 | Glyma.07g141900 | Domain of unknown function (DUF3527) |
| Glyma07g17110 | Glyma.07g142000 | PAP2 superfamily C-terminal |
| Glyma07g17116 | Glyma.07g142100 | Ubiquitinyl hydrolase 1. |
| Glyma07g17130 | Glyma.07g142300 | RING-variant domain |
| Glyma07g17140 | Glyma.07g142400 | Multicopper oxidase |
| Glyma07g17150 | Glyma.07g142500 | Multicopper oxidase |
| Glyma07g17170 | Glyma.07g142600 | Multicopper oxidase |
| Glyma07g17180 | Glyma.07g142700 | Fructose-1-6-bisphosphatase |
| Glyma07g17190 | Glyma.07g142800 | Uncharacterized protein |
| Glyma.Wm82. a1.v1.1 Gene Models | Glyma.Wm82. a2.v1 Gene Models | Description |
|---|---|---|
| Glyma11g01300 | Glyma.11G011000 | RNA-binding proteins |
| Glyma11g01310 | Glyma.11G011100 | Uncharacterized protein |
| Glyma11g01320 | Glyma.11G011200 | NADH: ubiquinone oxidoreductase, B17.2 subunit |
| Glyma11g01330 | Glyma.11G011300 | E3 ubiquitin ligase |
| Glyma11g01340 | Glyma.11G011400 | Translin-associated protein X |
| Glyma11g01350 | Glyma.11G011500 | Chalcone and stilbene synthases |
| Glyma11g01360 | Glyma.11G011600 | PPR repeat |
| Glyma11g01370 | Glyma.11G011700 | Nuclear transport receptor CRM1/MSN5 (importin beta superfamily) |
| Glyma11g01380 | Glyma.11G011800 | GTP-binding ADP-ribosylation factor Arf1 |
| Glyma11g01390 | Glyma.11G011900 | Plant protein of unknown function (DUF946) |
| Glyma11g01405 | Glyma.11G012000 | Guanosine-3′,5′-bis(diphosphate)3′-pyrophosphohydrolase |
| Glyma11g01420 | Glyma.11G012100 | Ribonuclease P |
| Glyma11g01430 | Glyma.11G012200 | DEAD/DEAH box helicase/Helicase conserved C-terminal domain |
| Glyma11g01441 | Glyma.11G012300 | Pwwp domain-containing protein |
| Glyma11g01450 | Glyma.11G012400 | Cell division cycle 20 (CDC20) (Fizzy)-related |
| Glyma11g01460 | Glyma.11G012500 | Putative u4/u6 small nuclear ribonucleoprotein |
| Glyma11g01470 | Glyma.11G012700 | Mitochondrial outer membrane protein |
| Glyma11g01480 | Glyma.11G012800 | Galactosyltransferases |
| Glyma11g01491 | Glyma.11G012900 | Aspartyl proteases |
| Glyma11g01501 | Glyma.11G013000 | Aspartyl proteases |
| Glyma11g01510 | Glyma.11G013100 | Aspartyl proteases |
| Glyma11g01520 | Glyma.11G013200 | Uncharacterized protein |
| Glyma11g01530 | Glyma.11G013300 | PLAC8 family |
| Glyma11g01536 | Glyma.11G013400 | DYW family of nucleic acid deaminases (DYW_deaminase) |
| Glyma11g01543 | Glyma.11G013500 | PPR repeat (PPR) |
| Glyma11g01550 | Glyma.11G013600 | PPR repeat (PPR) |
| Glyma11g01570 | Glyma.11G013700 | leucine-rich PPR motif-containing protein, mitochondrial (LRPPRC) |
| Glyma11g01580 | Glyma.11G013800 | Complex 1 protein (LYR family) |
| Glyma11g01595 | Glyma.11G013900 | KH domain containing RNA binding protein |
| Glyma11g01610 | Glyma.11G014000 | Protein Phosphatase methylesterase-1 related |
| Glyma11g01620 | Glyma.11G014100 | Cytochrome c |
| Glyma11g01640 | Glyma.11G014200 | Ethylene-responsive transcription factor ERF021 |
| Glyma11g01650 | Glyma.11G014300 | Nuclear transport factor 2 (NTF2) domain |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, M.; Wang, Y.; Gai, J.; Bhat, J.A.; Li, Y.; Kong, J.; Zhao, T. Genetic Analysis and Gene Mapping for a Short-Petiole Mutant in Soybean (Glycine max (L.) Merr.). Agronomy 2019, 9, 709. https://doi.org/10.3390/agronomy9110709
Liu M, Wang Y, Gai J, Bhat JA, Li Y, Kong J, Zhao T. Genetic Analysis and Gene Mapping for a Short-Petiole Mutant in Soybean (Glycine max (L.) Merr.). Agronomy. 2019; 9(11):709. https://doi.org/10.3390/agronomy9110709
Chicago/Turabian StyleLiu, Meifeng, Yaqi Wang, Junyi Gai, Javaid Akhter Bhat, Yawei Li, Jiejie Kong, and Tuanjie Zhao. 2019. "Genetic Analysis and Gene Mapping for a Short-Petiole Mutant in Soybean (Glycine max (L.) Merr.)" Agronomy 9, no. 11: 709. https://doi.org/10.3390/agronomy9110709
APA StyleLiu, M., Wang, Y., Gai, J., Bhat, J. A., Li, Y., Kong, J., & Zhao, T. (2019). Genetic Analysis and Gene Mapping for a Short-Petiole Mutant in Soybean (Glycine max (L.) Merr.). Agronomy, 9(11), 709. https://doi.org/10.3390/agronomy9110709