Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB)


Abstract
1. Introduction
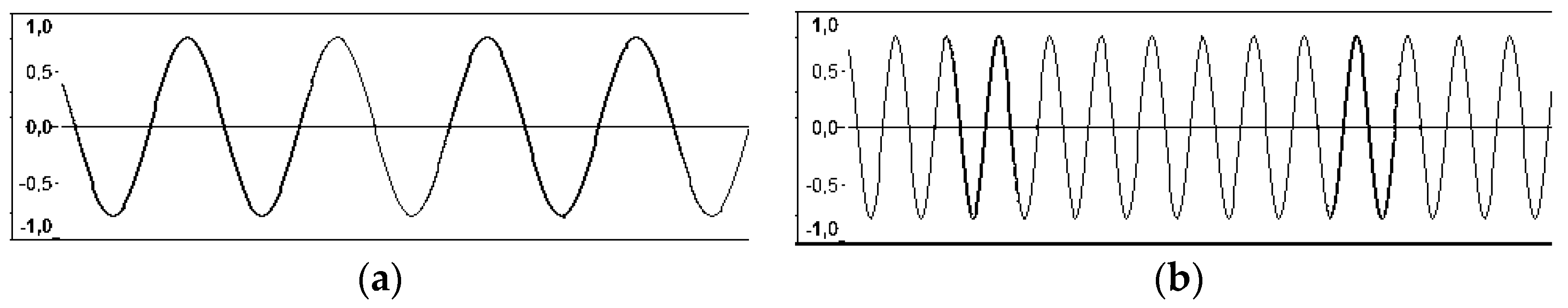
2. Sound Waves and Sound Magnitude
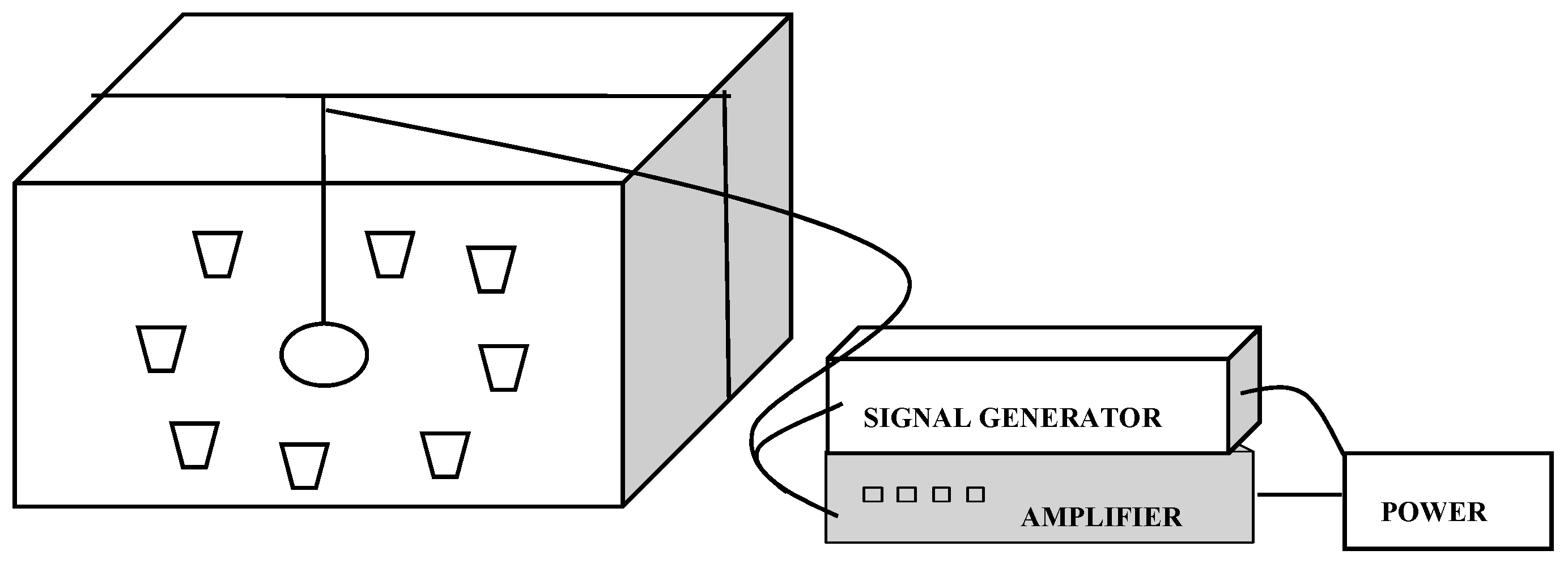
3. Materials and Methods
3.1. Measurements and Analyses in Strawberry Plants and Fruits
3.2. Determination of Total Soluble Solids (TSS) and Titratable Acidity (TTA)
3.3. Extraction of Sugars and Acids (TS and TA)
3.4. HPLC of Organic Acid and Sugars
3.5. Determination of Total Phenolic Content (TPC) and Ascorbic Acid (AA) in Strawberry Fruits
4. Results and Discussion
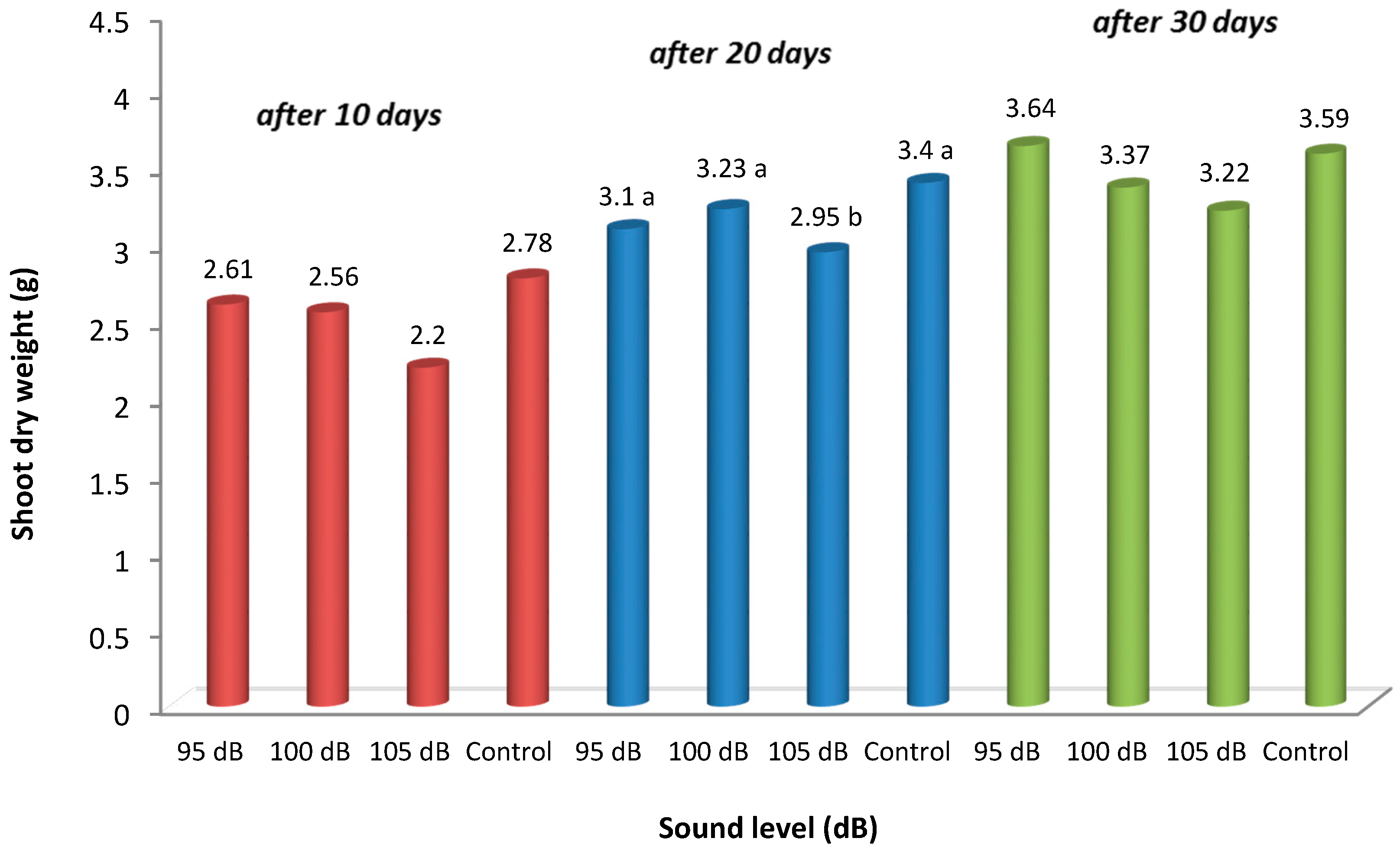
4.1. Results of the Weight Measurements in Strawberry Plants in Fresh and Dry Roots and Shoots
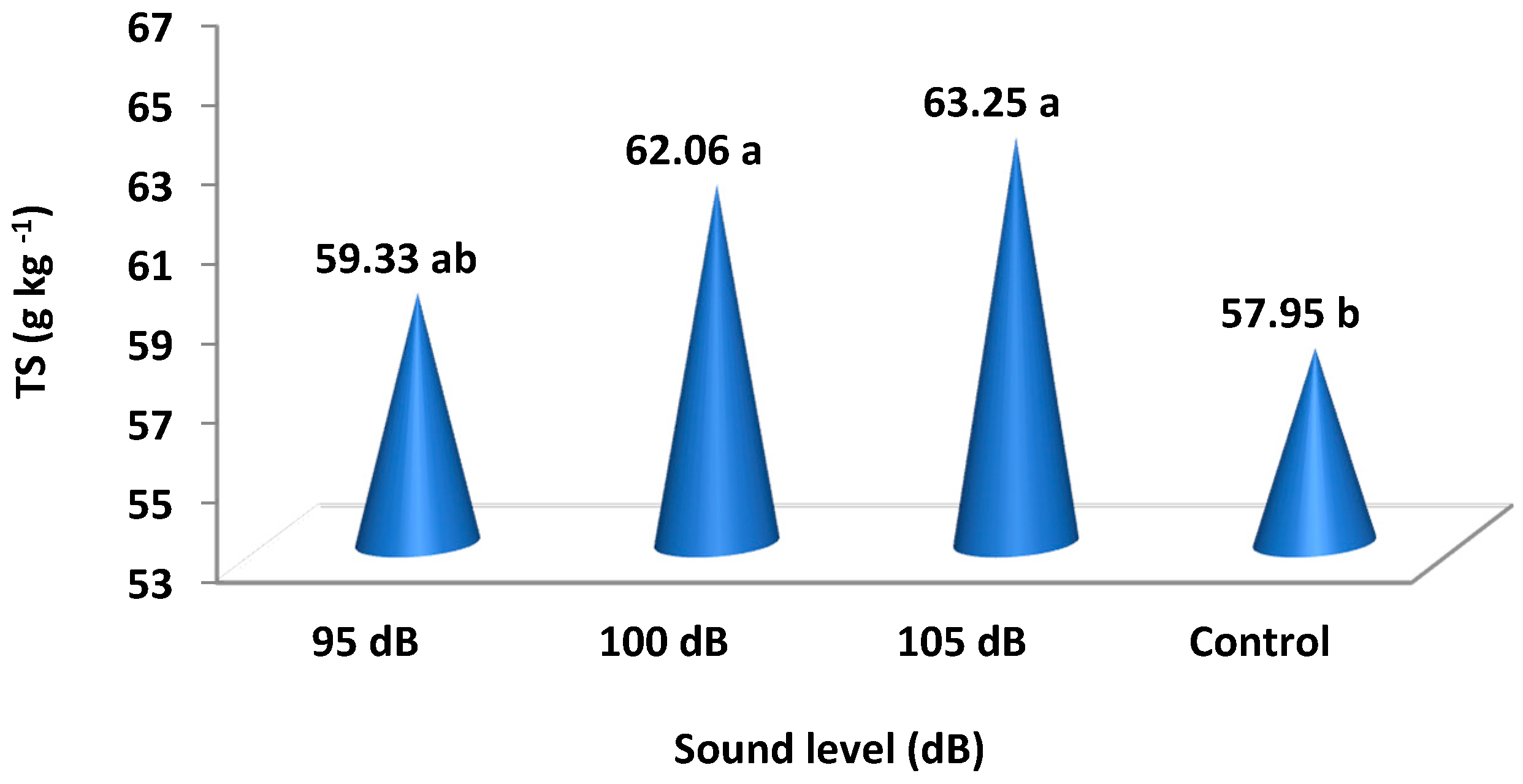
4.2. Results of the Analysis of Strawberry Plants
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Klein, B.P.; Perry, A.K. Ascorbic acid and vitamin A activity in selected vegetables from different geographical areas of the United States. J. Food Sci. 1982, 47, 941–945. [Google Scholar] [CrossRef]
- Robards, K.; Prenzler, P.D.; Tucker, G.; Swatsitang, P.; Glover, W. Phenolic compounds and their role in oxidative processes in fruits. Food Chem. 1999, 66, 401–436. [Google Scholar] [CrossRef]
- Akula, R.; Ravishankar, G.A. Influence of abiotic stress signals on secondary metabolites in plants. Plant Signal. Behav. 2011, 6, 1720–1731. [Google Scholar] [CrossRef] [PubMed]
- Ozkurt, H.; Altuntas, O. The Effect of Sound Waves at Different Frequencies upon the Plant Element Nutritional Uptake of Snake Plant (Sansevieria trifasciata) Plants. Indian J. Sci. Technol. 2016, 9, 48–55. [Google Scholar] [CrossRef]
- Ozkurt, H.; Altuntas, O.; Bozdogan, E. The Effects of Sound Waves upon Plant Nutrient Elements Uptake of Sword Fern (Nephrolepis exaltata) Plants. J. Basic Appl. Sci. Res. 2016, 6, 9–15. [Google Scholar]
- Jaffe, M.J.; Leopold, A.C.; Staples, R.C. Thigmo responses inplants and fungi. Am. J. Bot. 2002, 89, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Baluška, F.; Šamaj, J.; Wojtazek, P.; Volkmann, D.; Menzel, D. Cytoskeleton–plasma membrane–cell wall continuum in plants. Emerging links revisited. Plant Physiol. 2003, 133, 483–491. [Google Scholar] [CrossRef] [PubMed]
- Mary, M.; Braam, J. The Arabidopsis TCH4 xyloglucan endotransglyco-sylase. Plant Physiol. 1997, 115, 181–190. [Google Scholar]
- Sistrunk, M.L.; Antosiewicz, D.M.; Purugganan, M.M.; Braam, J. Arabidopsis TCH3 encodes a novel Ca2+ binding protein and shows environmentally induced and tissue-specific regulation. Plant Cell 1994, 6, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Kadota, A.; Wada, M. Mechanically induced avoidance response of chloroplasts in fern protonemal cells. Plant Physiol. 1999, 121, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Erner, Y.; Jaffe, M.J. Thigmomorphogenesis: The involvement of auxin and abscisic acid in growth retardation due to mechanical perturbation. Plant Cell Physiol. 1982, 23, 935–941. [Google Scholar]
- Wang, L.; Weller, C.L. Recent advances in extraction of nutraceuticals from plants. Trends Food Sci. Technol. 2006, 17, 300–312. [Google Scholar] [CrossRef]
- Hou, T.Z.; Li, M.D. Experimental evidence of a plant meridian system: IV. The effects of acupuncture on growth and metabolism of Phaseolus vulgaris L. beans. Am. J. Chin. Med. 1997, 25, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Z.; Luan, J.Y.; Wang, J.Y.; Li, M.D. Experimental evidence of a plant meridian system: III. The sound characteristics of Phylodendron (Alocasia) and effects of acupuncture on those properties. Am. J. Chin. Med. 1994, 22, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.Z.; Re, Z.W.; Li, M.D. Experimental evidence of a plant meridian system: II. The effects of needle acupuncture on the; temperature changes of soybean (Glycine max). Am. J. Chin. Med. 1994, 22, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.C.; Wang, B.C.; Duan, C.R.; Dai, C.Y.; Jia, Y.; Wang, X.J. Brief study on physiological effects of sound field on actinidia Chinese callus. J. Chongqing Univ. 2002, 25, 79–84. (In Chinese) [Google Scholar]
- Yang, X.C.; Wang, B.C.; Duan, C.R. Effects of sound stimulation on energy metabolism of Actinidia chinensis callus. Colloids Surf. B Biointerfaces 2003, 30, 67–72. [Google Scholar]
- Yang, X.C.; Wang, B.C.; Ye, M. Effects of different sound intensities on root development of Actinidia Chinese plantlet. Chin. J. Appl. Environ. Biol. 2004, 10, 274–276. (In Chinese) [Google Scholar]
- Taş, F. Internal Acoustic Research and Improvement Methods in a Commercial Vehicle. Ph.D. Thesis, Istanbul Technical University, Istanbul, Turkey, 2010. [Google Scholar]
- Hoagland, D.R.; Arnon, D.I. The water culture method for growing plants without soil. In Circular California Agricultural Experiment Station; University of California: Berkeley, CA, USA, 1938; pp. 347–461. [Google Scholar]
- Kafkas, E.; Koşar, M.; Paydaş, S.; Kafkas, S.; Başer, K.H.C. Quality characteristics of strawberry genotypes at different maturation stages. Food Chem. 2007, 100, 1229–1236. [Google Scholar] [CrossRef]
- Pinheiro, C.; Chaves, M.M. Photosynthesis and drought: Can we make metabolic connections from available data? J. Exp. Bot. 2010, 62, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Cramer, G.R.; Ergül, A.; Grimplet, J.; Tillett, R.L.; Tattersall, E.A.; Bohlman, M.C.; Quilici, D. Water and salinity stress in grapevines: Early and late changes in transcript and metabolite profiles. Funct. Integr. Genom. 2007, 7, 111–134. [Google Scholar] [CrossRef] [PubMed]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; Harter, K. The AtGenExpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Munns, R. Comparative physiology of salt and water stress. Plant Cell Environ. 2002, 25, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Rosa, S.B.; Caverzan, A.; Teixeira, F.K.; Lazzarotto, F.; Silveira, J.A.; Ferreira-Silva, S.L.; Abreu-Neto, J.; Margis, R.; Margis-Pinheiro, M. Cytosolic APx knockdown indicates an ambiguous redox responses in rice. Phytochemistry 2010, 71, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Britt, A.B. Repair of DNA damage induced by solar UV. Photosynth. Res. 2004, 81, 105–112. [Google Scholar] [CrossRef]
- Hilal, M.; Parrado, M.F.; Rosa, M.; Gallardo, M.; Orce, L.; Massa, E.M.; Prado, F.E. Epidermal lignin deposition in quinoa cotyledons in response to UV-B radiation. Photochem. Photobiol. 2004, 79, 205–210. [Google Scholar] [CrossRef]
- Ibañez, S.; Rosa, M.; Hilal, M.; González, J.A.; Prado, F.E. Leaves of Citrus aurantifolia exhibit a different sensibility to solar UV-B radiation according to development stage in relation to photosynthetic pigments and UV-B absorbing compounds production. J. Photochem. Photobiol. B Biol. 2008, 90, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Bartwal, A.; Mall, R.; Lohani, P.; Guru, S.K.; Arora, S. Role of secondary metabolites and brassinosteroids in plant defense against environmental stresses. J. Plant Growth Regul. 2013, 32, 216–232. [Google Scholar] [CrossRef]
- Seigler, D.S. Plant Secondary Metabolism; Kluwer Academic Publishers: Boston, MA, USA, 1998. [Google Scholar]
- DiCosmo, F.; Misawa, M. Eliciting secondary metabolism in plant cell cultures. Trends Biotechnol. 1985, 3, 318. [Google Scholar] [CrossRef]
- Sudha, G.; Ravishankar, G.A. Influence of methyl jasmonate and salicylic acid in the enhancement of capsaicin production in cell suspension cultures of Capsicum frutescens Mill. Curr. Sci. 2003, 85, 1212–1217. [Google Scholar]
- Karuppusamy, S. A review on trends in production of secondary metabolites from higher plants by in vitro tissue, organ and cell cultures. J. Med. Plants Res. 2009, 3, 1222–1239. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Cook, D.; Fowler, S.; Fiehn, O.; Thomashow, M.F. A prominent role for the CBF cold response pathway in configuring the low-temperature metabolome of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 15243–15248. [Google Scholar] [CrossRef] [PubMed]
- Chalker-Scott, L.; Fnchigami, L.H. The role of phenolic compounds in plant stress responses. In Low Temperature Stress Physiology in Crops; Paul, H.L., Ed.; CRC Press Inc.: Boca Raton, FL, USA, 1989; p. 40. [Google Scholar]
- Kaplan, F.; Kopka, J.; Sung, D.Y.; Zhao, W.; Popp, M.; Porat, R.; Guy, C.L. Transcript and metabolite profiling during cold acclimation of Arabidopsis reveals an intricate relationship of cold-regulated gene expression with modifications in metabolite content. Plant J. 2007, 50, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Wingler, A.; Roitsch, T. Metabolic regulation of leaf senescence: Interactions of sugar signalling with biotic and abiotic stress responses. Plant Biol. 2008, 10, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Hare, P.D.; Cress, W.A.; Van Staden, J. Dissecting the roles of osmolyte accumulation during stress. Plant Cell Environ. 1998, 21, 535–553. [Google Scholar] [CrossRef]
- Avonce, N.; Leyman, B.; Mascorro-Gallardo, J.O.; Van Dijck, P.; Thevelein, J.M.; Iturriaga, G. The Arabidopsis trehalose-6-P synthase AtTPS1 gene is a regulator of glucose, abscisic acid, and stress signalling. Plant Physiol. 2004, 136, 3649–3659. [Google Scholar] [CrossRef] [PubMed]
- Pego, J.V.; Kortstee, A.J.; Huijser, C.; Smeekens, S.C.M. Photosynthesis, sugars and the regulation of gene expression. J. Exp. Bot. 2000, 51, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Ramon, M.; Rolland, F.; Sheen, J. Sugar sensing and signaling. The Arabidopsis Book. Am. Soc. Plant Biol. 2008. [Google Scholar] [CrossRef]
- Van den Ende, W.; Valluru, R. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging? J. Exp. Bot. 2009, 60, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef] [PubMed]
- Kalt, W.; Ryan, D.A.J.; Duy, J.C.; Prior, R.L.; Ehlenfeldt, M.K.; Vander Kloet, S.P. Interspecific variation in anthocyanins, phenolics, and antioxidant capacity among genotypes of highbush and lowbush blueberries (Vaccinium section cyanococcus spp.). J. Agric. Food Chem. 2001, 49, 4761–4767. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kader, A.A. Preharvest and postharvest factors influencing vitamin C content of horticultural crops. Postharvest Biol. Technol. 2000, 20, 207–220. [Google Scholar] [CrossRef]
- Dumas, Y.; Dadomo, M.; Di Lucca, G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidantcontent of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Bartoli, C.G.; Yu, J.; Gomez, F.; Fernández, L.; McIntosh, L.; Foyer, C.H. Inter-relationships between light and respiration in the control of ascorbic acid synthesis and accumulation in Arabidopsis thaliana leaves. J. Exp. Bot. 2006, 57, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Conklin, P.L.; Loewus, F.A. Biosynthesis of ascorbic acid in plants: A renaissance. Annu. Rev. Plant Biol. 2001, 52, 437–467. [Google Scholar] [CrossRef] [PubMed]
- Pallanca, J.E.; Smirnoff, N. The control of ascorbic acid synthesis and turnover in pea seedlings. J. Exp. Bot. 2000, 51, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Green, M.A.; Fry, S.C. Vitamin C degradation in plant cells via enzymatic hydrolysis of 4-O-oxalyl-l-threonate. Nature 2005, 433, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Horemans, N.; Foyer, C.H.; Asard, H. Transport and action of ascorbate at the plant plasma membrane. Trends Plant Sci. 2000, 5, 263–267. [Google Scholar] [CrossRef]
- Ferrandino, A.; Lovisolo, C. Abiotic stress effects on grapevine (Vitis vinifera L.): Focus on abscisic acid-mediated consequences on secondary metabolism and berry quality. Environ. Exp. Bot. 2014, 103, 138–147. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Poiroux-Gonord, F.; Bidel, L.P.; Fanciullino, A.L.; Gautier, H.; Lauri-Lopez, F.; Urban, L. Health benefits of vitamins and secondary metabolites of fruits and vegetables and prospects to increase their concentrations by agronomic approaches. J. Agric. Food Chem. 2010, 58, 12065–12082. [Google Scholar] [CrossRef] [PubMed]
- Grassmann, J.; Hippeli, S.; Elstner, E.F. Plant’s defence and its benefits for animals and medicine: Role of phenolics and terpenoids in avoiding oxygen stress. Plant Physiol. Biochem. 2002, 40, 471–478. [Google Scholar] [CrossRef]
- Kunz, D.A.; Chen, J.L.; Pan, G. Accumulation of α-keto acids as essential components in cyanide assimilation by Pseudomonas fluorescens NCIMB 11764. Appl. Environ. Microbiol. 1998, 64, 4452–4459. [Google Scholar] [PubMed]
- English-Loeb, G.; Stout, M.J.; Duffey, S.S. Drought stress in tomatoes: Changes in plant chemistry and potential nonlinear consequences for insect herbivores. Oikos 1997, 79, 456–468. [Google Scholar] [CrossRef]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 10 Days after the Sound Treatment | 20 Days after the Sound Treatment | 30 Days after the Sound Treatment | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sound Intensity Treatments | RFW (g) | RDW (g) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | SFW (g) | SDW (g) | RFW (g) | RDW (g) | SFW (g) | SDW (g) |
| 95 dB | 7.94 a | 1.10 a | 10.92 | 2.61 | 9.50 ab | 1.38 a | 17.84 ab | 3.10 a | 12.50 a | 1.72 a | 21.25 a | 3.64 |
| 100 dB | 5.77 b | 0.85 b | 10.33 | 2.56 | 8.98 b | 1.32 a | 16.32 b | 3.23 a | 9.32 b | 1.40 ab | 19.18 b | 3.37 |
| 105 dB | 4.74 b | 0.64 b | 9.13 | 2.20 | 6.30 c | 0.78 b | 15.05 b | 2.95 b | 8.86 c | 0.96 b | 18.52 b | 3.22 |
| Control | 7.34 a | 1.15 a | 11.15 | 2.78 | 10.57 a | 1.43 a | 19.52 a | 3.40 a | 13.03 a | 1.77 a | 22.92 a | 3.59 |
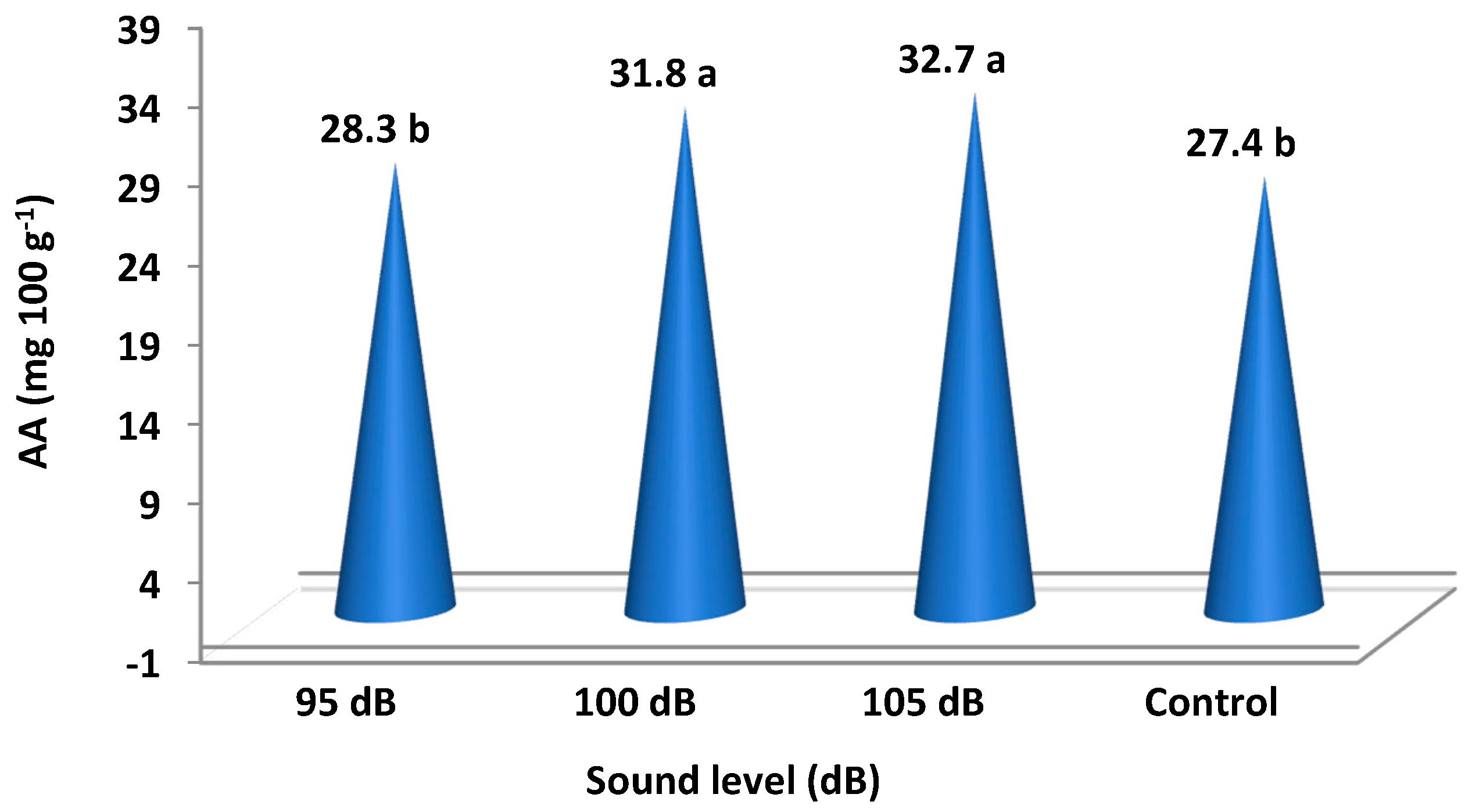
| Sound Intensity Treatments | pH | TSS (%) | TTA (g kg−1) | TS (g kg−1) | TA (g kg−1) | TPC (mg 100 g−1 Gallic Acid) | AA (mg 100 g−1) |
|---|---|---|---|---|---|---|---|
| 95 dB | 3.36 | 8.4 | 7.37 | 59.33 ab | 19.82 | 279.5 b | 28.3 b |
| 100 dB | 3.38 | 8.3 | 7.41 | 62.06 a | 20.62 | 282.0 ab | 31.8 a |
| 105 dB | 3.39 | 8.4 | 7.42 | 63.25 a | 21.36 | 288.0 a | 32.7 a |
| Control | 3.36 | 8.2 | 7.35 | 57.95 b | 19.55 | 275.5 b | 27.4 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ozkurt, H.; Altuntas, O. Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB). Agronomy 2018, 8, 127. https://doi.org/10.3390/agronomy8070127
Ozkurt H, Altuntas O. Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB). Agronomy. 2018; 8(7):127. https://doi.org/10.3390/agronomy8070127
Chicago/Turabian StyleOzkurt, Halil, and Ozlem Altuntas. 2018. "Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB)" Agronomy 8, no. 7: 127. https://doi.org/10.3390/agronomy8070127
APA StyleOzkurt, H., & Altuntas, O. (2018). Quality Parameter Levels of Strawberry Fruit in Response to Different Sound Waves at 1000 Hz with Different dB Values (95, 100, 105 dB). Agronomy, 8(7), 127. https://doi.org/10.3390/agronomy8070127