Physiological and Agronomic Strategies to Increase Mungbean Yield in Climatically Variable Environments of Northern Australia

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Mungbean Evolution and Diversity
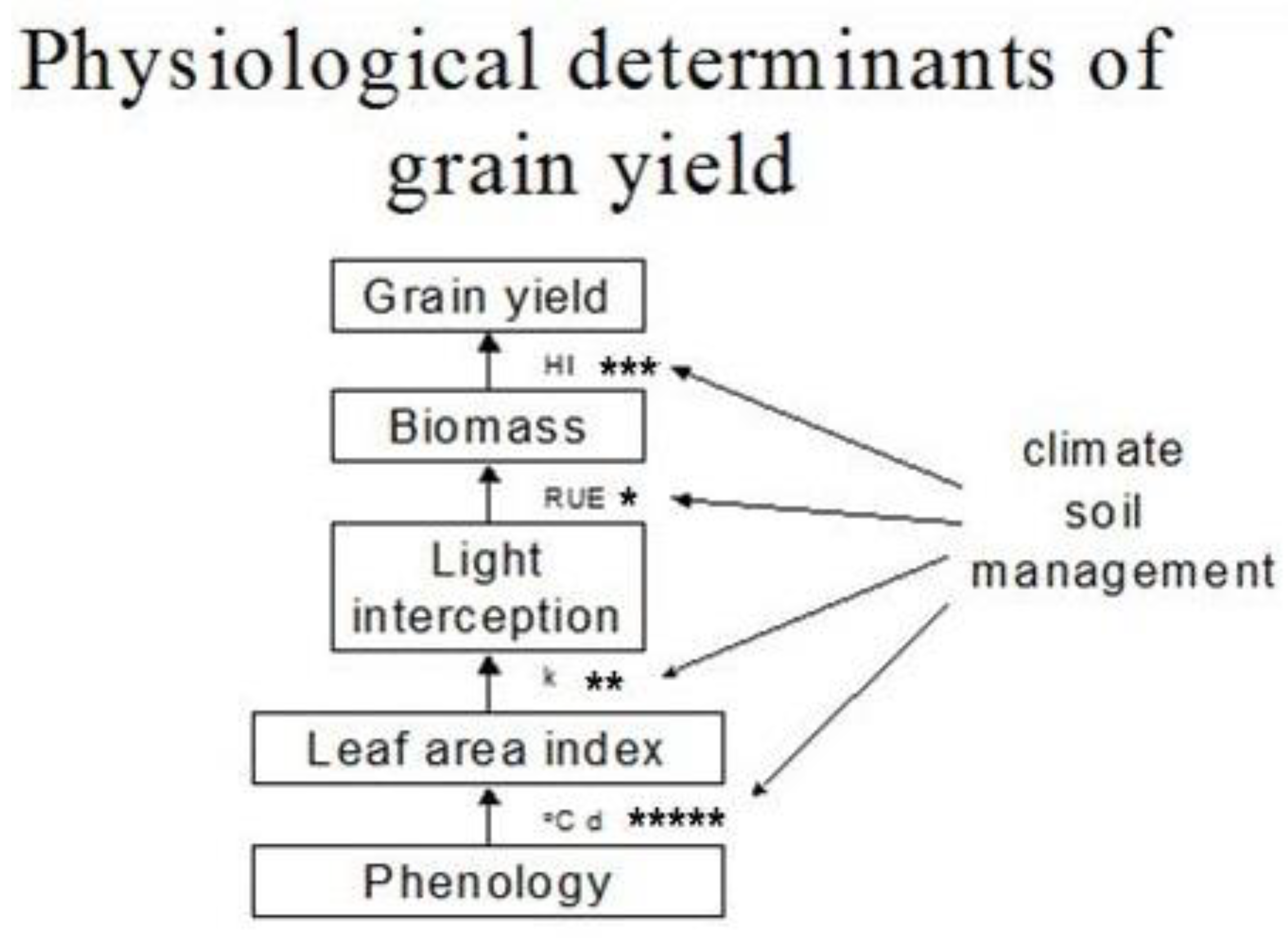
3. Physiological Framework of Yield in Mungbean
3.1. Crop Establishment and Development
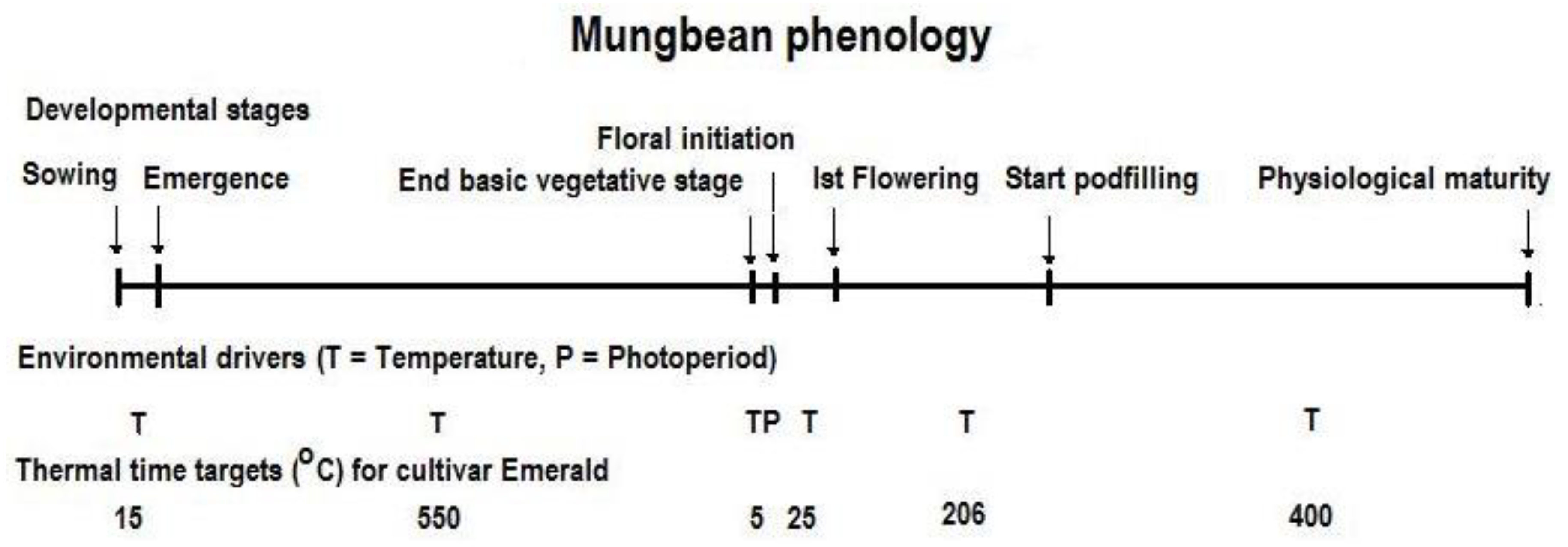
3.2. Crop Phenology
3.3. Leaf Area Development
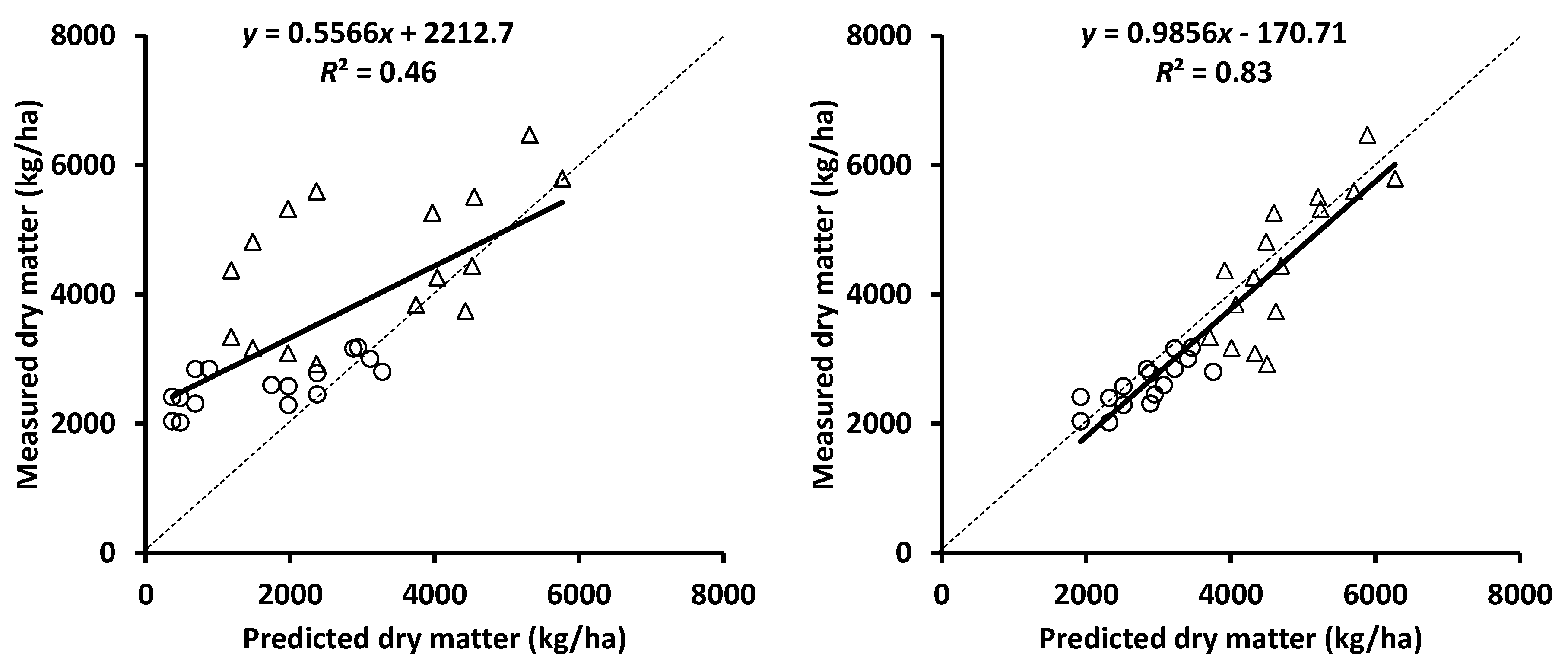
3.4. Biomass Accumulation
3.5. Biomass Partitioning
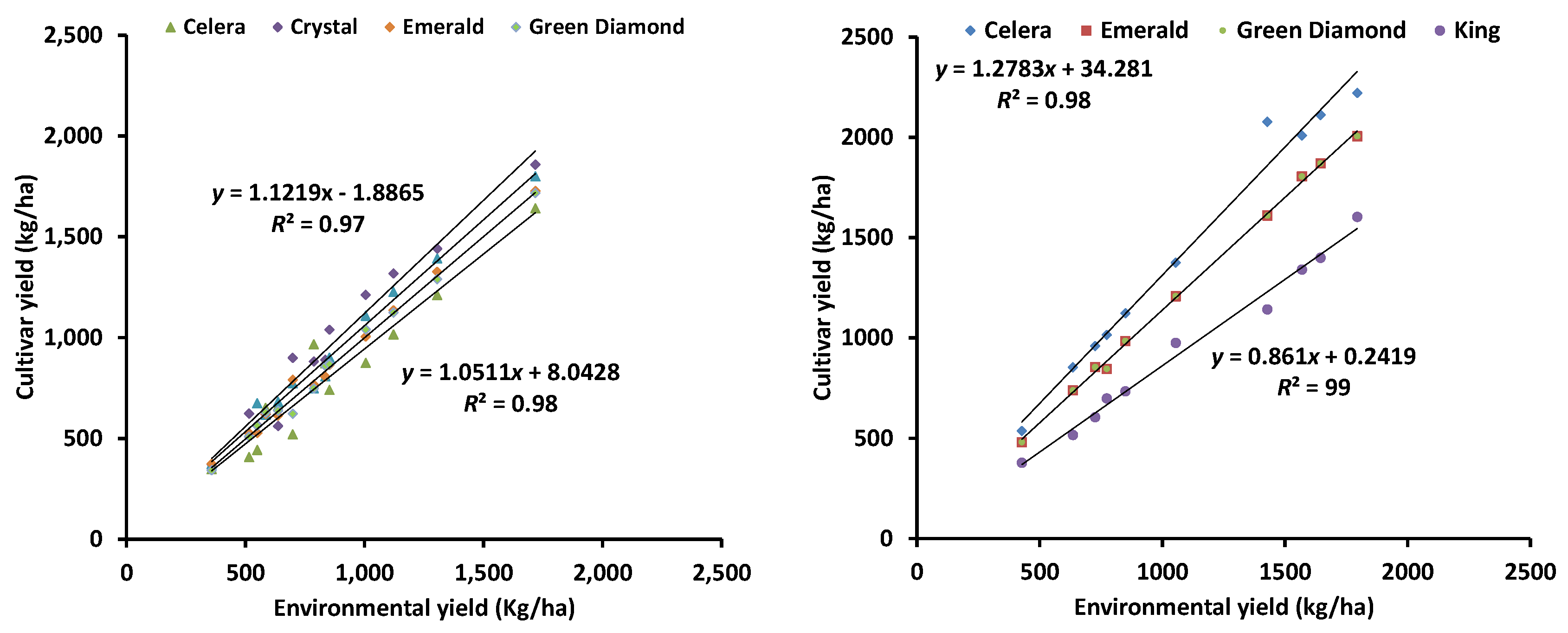
4. Genotype × Environment Interactions
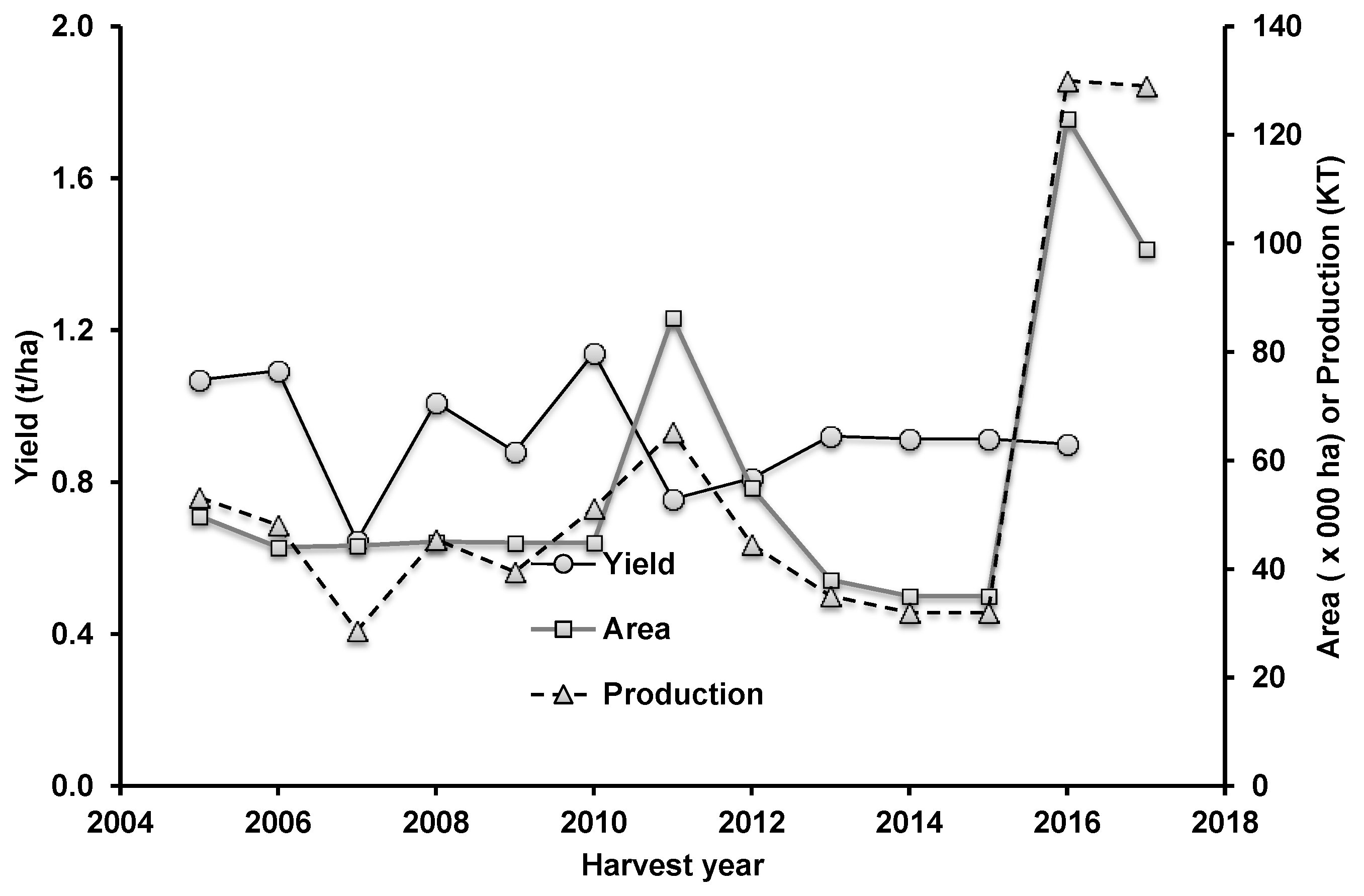
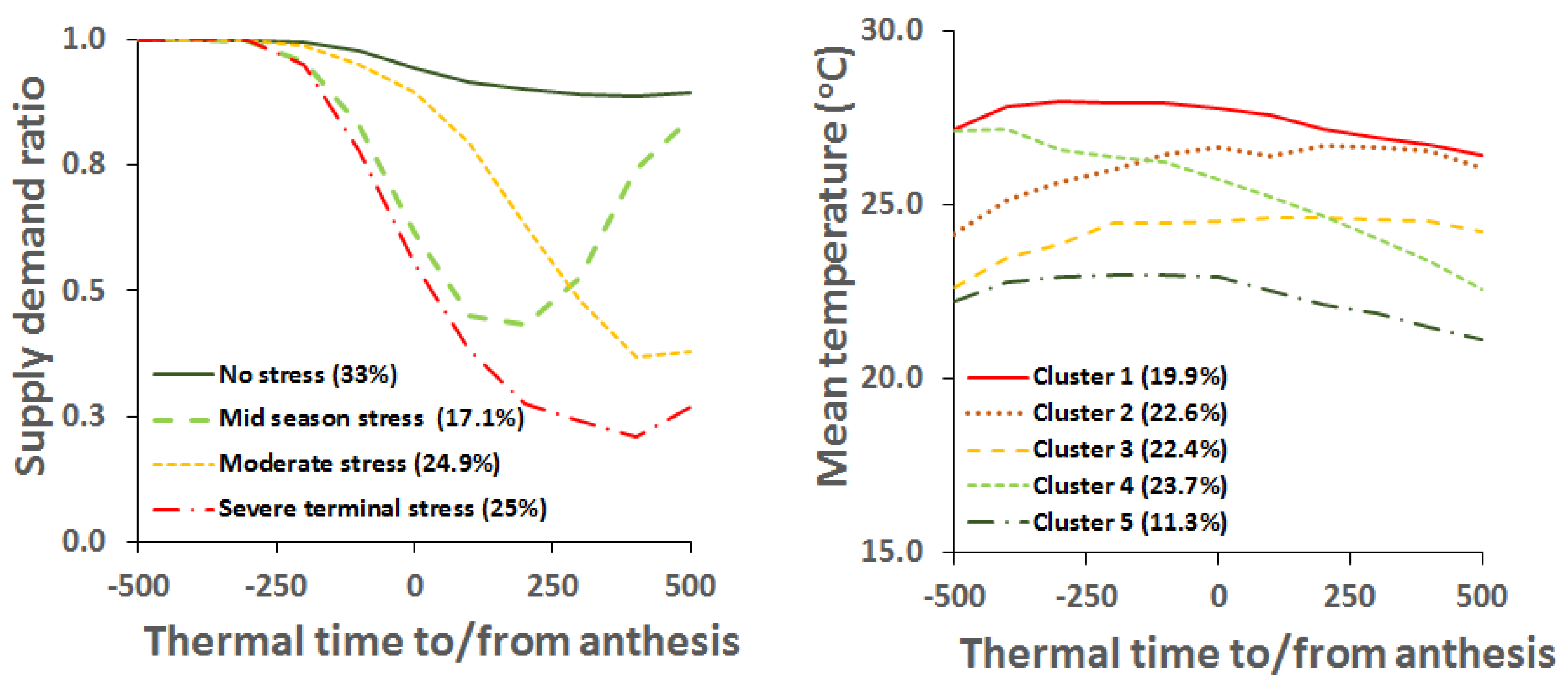
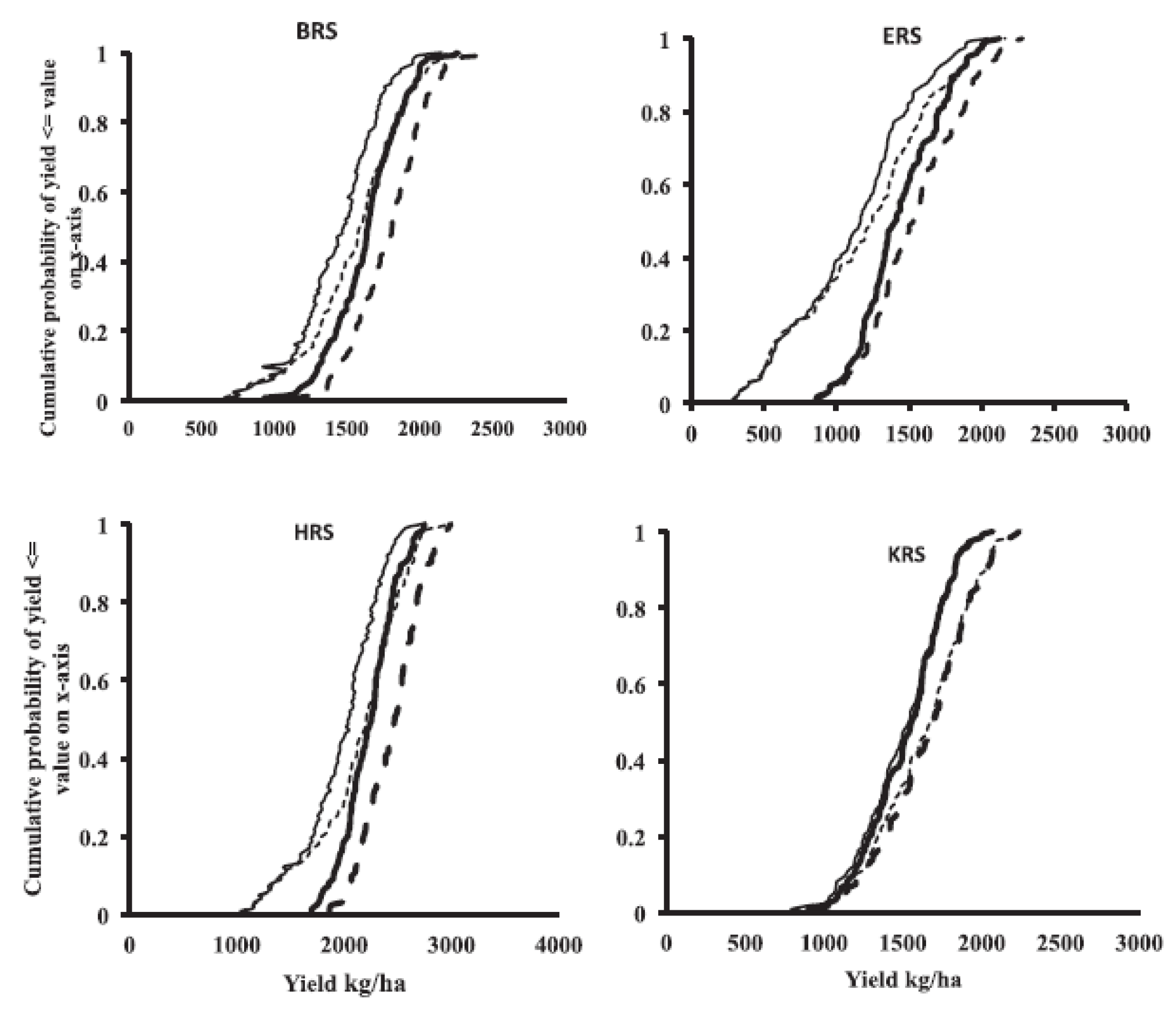
5. Growing Environments of Northern Australia—Main Features
6. Agronomic Innovations in Mungbean Production
7. Conclusions and Future Research Directions
Acknowledgments
Conflicts of Interest
References
- Nair, R.M.; Yang, R.Y.; Easdown, W.J.; Thavarajah, D.; Thavarajah, P.; Hughes, J.D.A.; Keatinge, J. Biofortification of mungbean (Vigna radiata) as a whole food to enhance human health. J. Sci. Food Agric. 2013, 93, 1805–1813. [Google Scholar] [CrossRef] [PubMed]
- Peoples, M.; Brockwell, J.; Herridge, D.; Rochester, I.; Alves, B.; Urquiaga, S.; Boddey, R.; Dakora, F.; Bhattarai, S.; Maskey, S. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Ebert, A.W. Potential of underutilized traditional vegetables and legume crops to contribute to food and nutritional security, income and more sustainable production systems. Sustainability 2014, 6, 319–335. [Google Scholar] [CrossRef]
- Keatinge, J.; Easdown, W.; Yang, R.; Chadha, M.; Shanmugasundaram, S. Overcoming chronic malnutrition in a future warming world: The key importance of mungbean and vegetable soybean. Euphytica 2011, 180, 129–141. [Google Scholar] [CrossRef]
- Tang, D.; Dong, Y.; Ren, H.; Li, L.; He, C. A review of phytochemistry, metabolite changes, and medicinal uses of the common food mung bean and its sprouts (Vigna radiata). Chem. Cent. J. 2014, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Alexandratos, N.; Bruinsma, J. World Agriculture towards 2030/2050: The 2012 Revision; ESA Working Paper; FAO: Rome, Italy, 2012. [Google Scholar]
- Lawn, R.; Russell, J. Mungbean: A grain legume for summer rainfall cropping areas of Australia. J. Aust. Inst. Agric. Sci. 1978, 44, 28–41. [Google Scholar]
- Robertson, M.; Carberry, P.; Lucy, M. Evaluation of a new cropping option using a participatory approach with on-farm monitoring and simulation: A case study of spring-sown mungbeans. Aust. J. Agric. Res. 2000, 51, 1–12. [Google Scholar] [CrossRef]
- Thomas Robertson, M.J.; Fukai, S.; Peoples, M.B. The effect of timing and severity of water deficit on growth, development, yield accumulation and nitrogen fixation of mungbean. Field Crops Res. 2004, 86, 67–80. [Google Scholar] [CrossRef]
- Amarasingha, R.; Suriyagoda, L.; Marambe, B.; Rathnayake, W.; Gaydon, D.; Galagedara, L.; Punyawardena, R.; Silva, G.; Nidumolu, U.; Howden, M. Improving water productivity in moisture-limited rice-based cropping systems through incorporation of maize and mungbean: A modelling approach. Agric. Water Manag. 2017, 189, 111–122. [Google Scholar] [CrossRef]
- Col Douglas. DAF, Hermitage, Qld, Australia. Personal communication. 2017.
- Lawn, R.; Imrie, B. Crop improvement for tropical and subtropical Australia: Designing plants for difficult climates. Field Crops Res. 1991, 26, 113–139. [Google Scholar] [CrossRef]
- Singh, H.; Joshi, B.; Thomas, T. The phaseolus group. In Pulse Crops of India’; Kachroo, P., Ed.; Indian Council of Agricultural Research: New Delhi, India, 1970; pp. 136–164. [Google Scholar]
- Smartt, J. Evolution of grain legumes. III. Pulses in the genus Vigna. Exp. Agric. 1985, 21, 87–100. [Google Scholar] [CrossRef]
- Lukoki, L.; Marechal, R.; Otoul, E. Les ancetres sauvages des haricots cultives: Vigna radiata (L.) Wilczek et V. mungo (L.) Hepper. Bull. Jard. Bot. Natl. Belg./Bull. Natl. Plantentuin Belg. 1980, 50, 385–391. [Google Scholar] [CrossRef]
- Abbas, G.; Hameed, A.; Rizwan, M.; Ahsan, M.; Asghar, M.J.; Iqbal, N. Genetic confirmation of mungbean (Vigna radiata) and mashbean (Vigna mungo) interspecific recombinants using molecular markers. Front. Plant Sci. 2015, 6, 1107. [Google Scholar] [CrossRef] [PubMed]
- Yimram, T.; Somta, P.; Srinives, P. Genetic variation in cultivated mungbean germplasm and its implication in breeding for high yield. Field Crops Res. 2009, 112, 260–266. [Google Scholar] [CrossRef]
- Gwag, J.G.; Chung, J.W.; Chung, H.K.; Lee, J.H.; Ma, K.H.; Dixit, A.; Park, Y.J.; Cho, E.G.; Kim, T.S.; Lee, S.H. Characterization of new microsatellite markers in mung bean, Vigna radiata (L.). Mol. Ecol. Resour. 2006, 6, 1132–1134. [Google Scholar] [CrossRef]
- Kim, S.K.; Nair, R.M.; Lee, J.; Lee, S.-H. Genomic resources in mungbean for future breeding programs. Front. Plant Sci. 2015, 6, 626. [Google Scholar] [CrossRef] [PubMed]
- Lambrides, C.; Godwin, I. Mungbean. In Pulses, Sugar and Tuber Crops; Springer: Berlin/Heidelberg, Germany, 2007; pp. 69–90. [Google Scholar]
- Shanmugasundaram, S.; Keatinge, J.; d Arros Hughes, J. The Mungbean Transformation Diversifying Crops, Defeating Malnutrition; International Food Policy Research Institute: Washington, DC, USA, 2009; Volume 922. [Google Scholar]
- Martre, P.; Quilot-Turion, B.; Luquet, D.; Memmah, M.-M.O.-S.; Chenu, K.; Debaeke, P. Model-assisted phenotyping and ideotype design. In Crop Physiology, 2nd ed.; Elsevier: New York, NY, USA, 2015; pp. 349–373. [Google Scholar]
- Kang, Y.J.; Kim, S.K.; Kim, M.Y.; Lestari, P.; Kim, K.H.; Ha, B.-K.; Jun, T.H.; Hwang, W.J.; Lee, T.; Lee, J. Genome sequence of mungbean and insights into evolution within Vigna species. Nat. Commun. 2014, 5, 5443. [Google Scholar] [CrossRef] [PubMed]
- Rachaputi, R.C.; Chauhan, Y.; Douglas, C.; Martin, W.; Krosch, S.; Agius, P.; King, K. Physiological basis of yield variation in response to row spacing and plant density of mungbean grown in subtropical environments. Field Crops Res. 2015, 183, 14–22. [Google Scholar] [CrossRef]
- Harris, D.; Breese, W.; Rao, J.K. The improvement of crop yield in marginal environments using ‘on-farm’seed priming: Nodulation, nitrogen fixation, and disease resistance. Aust. J. Agric. Res. 2005, 56, 1211–1218. [Google Scholar] [CrossRef]
- Cook, S.; Gupta, S.; Woodhead, T.; Larson, W. Soil physical constraints to establishment of mungbeans (Vigna radiata L. Wilczek) in paddy rice (Oryza sativa L.) soils. Soil Tillage Res. 1995, 33, 47–64. [Google Scholar] [CrossRef]
- Abrecht, D. No-till crop establishment on red earth soils at Katherine, Northern Territory: Effect of sowing depth and firming wheel pressure on the establishment of cowpea, mung bean, soybean and maize. Aust. J. Exp. Agric. 1989, 29, 397–402. [Google Scholar] [CrossRef]
- TeKrony, D.M.; Egli, D.B. Relationship of seed vigor to crop yield: A review. Crop Sci. 1991, 31, 816–822. [Google Scholar] [CrossRef]
- Armstrong, R.; McCosker, K.; Johnson, S.; Millar, G.; Walsh, K.; Kuskopf, B.; Probert, M.; Standley, J. Legume and opportunity cropping systems in central Queensland. 1. Legume growth, nitrogen fixation, and water use. Aust. J. Agric. Res. 1999, 50, 909–924. [Google Scholar] [CrossRef]
- Summerfield, R.; Lawn, R. Environmental modulation of flowering in mung bean (Vigna radiata): A reappraisal. Exp. Agric. 1987, 23, 461–470. [Google Scholar] [CrossRef]
- Robertson, M.; Carberry, P.; Huth, N.; Turpin, J.; Probert, M.E.; Poulton, P.; Bell, M.; Wright, G.; Yeates, S.; Brinsmead, R. Simulation of growth and development of diverse legume species in APSIM. Aust. J. Agric. Res. 2002, 53, 429–446. [Google Scholar] [CrossRef]
- Summerfield, R.; Lawn, R. Environmental modulation of flowering in mung bean (Vigna radiata): Further reappraisal for diverse genotypes and photothermal regimes. Exp. Agric. 1988, 24, 75–88. [Google Scholar] [CrossRef]
- Carberry, P.; Muchow, R.; Williams, R.; Sturtz, J.; McCown, R. A simulation model of kenaf for assisting fibre industry planning in northern Australia. I. General introduction and phenological model. Aust. J. Agric. Res. 1992, 43, 1501–1513. [Google Scholar] [CrossRef]
- Rebetzke, G.; Lawn, R. Adaptive responses of wild mungbean (Vigna radiata ssp. sublobata) to photo-thermal environment. II.* Growth, biomass, and seed yield. Aust. J. Agric. Res. 2006, 57, 929–937. [Google Scholar] [CrossRef]
- Imrie, B.; Lawn, R. Time to flowering of mung bean (Vigna radiata) genotypes and their hybrids in response to photoperiod and temperature. Exp. Agric. 1990, 26, 307–318. [Google Scholar] [CrossRef]
- Ellis, R.; Lawn, R.; Summerfield, R.; Qi, A.; Roberts, E.; Chay, P.; Brouwer, J.; Rose, J.; Yeates, S.; Sandover, S. Towards the reliable prediction of time to flowering in six annual crops. IV. Cultivated and wild mung bean. Exp. Agric. 1994, 30, 31–43. [Google Scholar] [CrossRef]
- Chauhan, Y.; Rachaputi, R.C. Defining agro-ecological regions for field crops in variable target production environments: A case study on mungbean in the northern grains region of Australia. Agric. For. Meteorol. 2014, 194, 207–217. [Google Scholar] [CrossRef]
- Rawson, H.; Craven, C. Variation between short duration mungbean cultivars (Vigna radiata (L.) Wilczek) in response to temperature and photoperiod. Indian J. Plant Physiol. 1979, 22, 127–136. [Google Scholar]
- Wallace, D.; Zobel, R.; Yourstone, K. A whole-system reconsideration of paradigms about photoperiod and temperature control of crop yield. Theor. Appl. Genet. 1993, 86, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Lee, J.Y.; Kim, D.K.; Yoon, C.Y.; Bak, G.C.; Park, I.J.; Bang, G.P.; Moon, J.K.; Oh, Y.J.; Min, K.S. A new high-yielding mungbean cultivar,” Samgang” with lobed leaflet. Kor. Breed. J. 2004, 36, 183–184. [Google Scholar]
- Muchow, R.; Robertson, M.; Pengelly, B. Accumulation and partitioning of biomass and nitrogen by soybean, mungbean and cowpea under contrasting environmental conditions. Field Crops Res. 1993, 33, 13–36. [Google Scholar] [CrossRef]
- Muchow, R. Canopy development in grain legumes grown under different soil water regimes in a semi-arid tropical environment. Field Crops Res. 1985, 11, 99–109. [Google Scholar] [CrossRef]
- Muchow, R.; Charles-Edwards, D. An analysis of the growth of mung beans at a range of plant densities in tropical Australia. I. Dry matter production. Aust. J. Agric. Res. 1982, 33, 41–51. [Google Scholar] [CrossRef]
- Kuo, C.; Wang, L.; Cheng, A.; Chou, M. Physiological basis for mungbean yield improvement. In Proceedings of the International Mungbean Symposium, Los Banos, Philippines, 16–19 August 1977. [Google Scholar]
- Lawn, R. Response of four grain legumes to water stress in south-eastern Queensland. II. Plant growth and soil water extraction patterns. Aust. J. Agric. Res. 1982, 33, 497–509. [Google Scholar] [CrossRef]
- Robertson, M.; Carberry, P.; Wright, G.; Singh, D. Using models to assess the value of traits of food legumes from a cropping systems perspective. In Linking Research and Marketing Opportunities for Pulses in the 21st Century; Springer: Berlin/Heidelberg, Germany, 2000; pp. 265–278. [Google Scholar]
- Muchow, R. An analysis of the effects of water deficits on grain legumes grown in a semi-arid tropical environment in terms of radiation interception and its efficiency of use. Field Crops Res. 1985, 11, 309–323. [Google Scholar] [CrossRef]
- Chauhan, Y.; Douglas, C.; Rachaputi, R.; Agius, P.; Martin, W.; King, K.; Skerman, A. Physiology of mungbean and development of the mungbean crop model. In Proceedings of the 1st Australian Summer Grains Conference, Gold Coas, Australia, 21–24 June 2010. [Google Scholar]
- Muchow, R.; Robertson, M.; Pengelly, B. Radiation-use efficiency of soybean, mugbean and cowpea under different environmental conditions. Field Crops Res. 1993, 32, 1–16. [Google Scholar] [CrossRef]
- Lawn, R. Agronomic and physiological constraints to the productivity of tropical grain legumes and prospects for improvement. Exp. Agric. 1989, 25, 509–528. [Google Scholar] [CrossRef]
- Lawn, R. Response of four grain legumes to water stress in south-eastern Queensland. III. Dry matter production, yield and water use efficiency. Aust. J. Agric. Res. 1982, 33, 511–521. [Google Scholar] [CrossRef]
- Dodt, M.G. Characterisation of Root Architectural Responses of Mungbean to Water Deficit; Queensland University of Technology: Brisbane, Australia, 2017. [Google Scholar]
- Singh, D.; Singh, B. Breeding for tolerance to abiotic stresses in mungbean. J. Food Legum. 2011, 24, 83–90. [Google Scholar]
- Bushby, H.; Lawn, R. Accumulation and partitioning of nitrogen and dry matter by contrasting genotypes of mungbean (Vigna radiata (L.) Wilczek). Aust. J. Agric. Res. 1992, 43, 1609–1628. [Google Scholar] [CrossRef]
- Cui, S.; Yu, D. Estimates of relative contribution of biomass, harvest index and yield components to soybean yield improvements in China. Plant Breed. 2005, 124, 473–476. [Google Scholar] [CrossRef]
- Duncan, W.; McCloud, D.; McGraw, R.; Boote, K. Physiological aspects of peanut yield improvement. Crop Sci. 1978, 18, 1015–1020. [Google Scholar] [CrossRef]
- Gifford, R.M.; Thorne, J.; Hitz, W. Crop productivity and photoassimilate partitioning. Science 1984, 225, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Hay, R. Harvest index: A review of its use in plant breeding and crop physiology. Ann. Appl. Biol. 1995, 126, 197–216. [Google Scholar] [CrossRef]
- Lawn, R.J. Contributions to Crop Improvement and Sustainable Agriculture in Tropical and Subtropical Australia; The University of Queensland: Brisbane, Australia, 2004. [Google Scholar]
- Lawn, R. Response of four grain legumes to water stress in south-eastern Queensland. I. Physiological response mechanisms. Aust. J. Agric. Res. 1982, 33, 481–496. [Google Scholar] [CrossRef]
- Lawn, R.; Williams, J. Limits imposed by climatological factors, Food Legume improvement for Asian farming systems. proceedings of an international workshop, Khon Kaen, Thailand, 1987; Wallis, E., Byth, D., Eds.; pp. 83–89. [Google Scholar]
- Muchow, R.; Charles-Edwards, D. An analysis of the growth of mung beans at a range of plant densities in tropical Australia. II. Seed production. Aust. J. Agric. Res. 1982, 33, 53–61. [Google Scholar] [CrossRef]
- Imrie, B.; Butler, K. An analysis of variability and genotype× environment interaction in mung bean (Vigna radiata) in south-eastern Queensland. Aust. J. Agric. Res. 1982, 33, 523–530. [Google Scholar] [CrossRef]
- Lawn, R. Agronomic studies on Vigna spp. in south-eastern Queensland. I. Phenological response of cultivars to sowing date. Aust. J. Agric. Res. 1979, 30, 855–870. [Google Scholar] [CrossRef]
- Lawn, R. Agronomic studies on Vigna spp. in south-eastern Queensland. II. Vegetative and reproductive response of cultivars to sowing date. Aust. J. Agric. Res. 1979, 30, 871–882. [Google Scholar] [CrossRef]
- Rebetzke, G.; Lawn, R. Adaptive responses of wild mungbean (Vigna radiata ssp. sublobata) to photo-thermal environment. I.* Phenology. Aust. J. Agric. Res. 2006, 57, 917–928. [Google Scholar] [CrossRef]
- Inouye, J.; Shanmugasundaram, S. Photoperiod and temperature effects on the growth and reproductive behaviour of less photoperiod sensitive soybeans. Soybeans Trop. Sub-Trop. Crop. Syst. 1984, 354–360. [Google Scholar]
- Rowell, T.; Mortley, D.; Loretan, P.; Bonsi, C.; Hill, W. Continuous daily light period and temperature influence peanut yield in nutrient film technique. Crop Sci. 1999, 39, 1111–1114. [Google Scholar] [CrossRef]
- Chauhan, Y.; Venkataratnam, N.; Sheldrake, A. Factors affecting growth and yield of short-duration pigeonpea and its potential for multiple harvests. J. Agric. Sci. 1987, 109, 519–529. [Google Scholar] [CrossRef]
- Saxena, K.; Gupta, S.; Sharma, D.; Reddy, L.; Chauhan, Y.; Kannaiyan, J.; Green, J.; Nene, Y.; Faris, D. Registration of ICPL 87 pigeonpea. Crop Sci. 1989, 29, 237–237. [Google Scholar] [CrossRef]
- Chauhan, Y.; Johansen, C.; Moon, J.-K.; Lee, Y.-H.; Lee, S.-H. Photoperiod responses of extra-short-duration pigeonpea lines developed at different latitudes. Crop Sci. 2002, 42, 1139–1146. [Google Scholar] [CrossRef]
- Lawn, R.; James, A. Application of physiological understanding in soybean improvement. I. Understanding phenological constraints to adaptation and yield potential. Crop Pasture Sci. 2011, 62, 1–11. [Google Scholar] [CrossRef]
- James, A.; Lawn, R. Application of physiological understanding in soybean improvement. II. Broadening phenological adaptation across regions and sowing dates. Crop Pasture Sci. 2011, 62, 12–24. [Google Scholar] [CrossRef]
- Bashandi, M.M.; Poehlman, J.T. Photoperiod response in mungbeans (Vigna radiata (L.) Wilczek). Euphytica 1974, 23, 691–697. [Google Scholar] [CrossRef]
- Major, D.; Kiniry, J.; Hodges, T. Predicting day length effects on phenological processes. In Predicting Crop Phenology; CRC Press: Boca Raton, FL, USA, 1991; pp. 15–28. [Google Scholar]
- Vergara, B.S.; Chang, T.-T. The Flowering Response of the Rice Plant to Photoperiod: A Review of the Literature; International Rice Reseach Institute: Los Banos, Philippines, 1985. [Google Scholar]
- Takeno, K. Epigenetic regulation of photoperiodic flowering. Plant Signal. Behav. 2010, 5, 788–791. [Google Scholar] [CrossRef] [PubMed]
- Alam Mondal, M.M.; Ali Fakir, M.S.; Juraimi, A.S.; Hakim, M.; Islam, M.; Shamsuddoha, A. Effects of flowering behavior and pod maturity synchrony on yield of mungbean [‘Vigna radiata’ (L.) Wilczek]. Aust. J. Crop Sci. 2011, 5, 945. [Google Scholar]
- Clifford, P. Control of reproductive sink yield in mung beans. Z. Pflanzenphysiol. 1981, 102, 173–181. [Google Scholar] [CrossRef]
- H. Brier. DAF, Kingaroy, Qld, Australia. Personal communication. 2017.
- Roberts, E.; Hadley, P.; Summerfield, R. Effects of temperature and photoperiod on flowering in chickpeas (Cicer arietinum L.). Ann. Bot. 1985, 55, 881–892. [Google Scholar] [CrossRef]
- Saxena, K.; Sultana, R.; Mallikarjuna, N.; Saxena, R.; Kumar, R.; Sawargaonkar, S.; Varshney, R. Male-sterility systems in pigeonpea and their role in enhancing yield. Plant Breed. 2010, 129, 125–134. [Google Scholar] [CrossRef]
- Imrie, B.; Drake, D.; Delacy, I.; Byth, D. Analysis of genotypic and environmental variation in international mungbean trials. Euphytica 1981, 30, 301–311. [Google Scholar] [CrossRef]
- Swindell, R.E.; Poehlman, J. Inheritance of photoperiod response in mungbean (Vigna radiata [L.] Wilczek). Euphytica 1978, 27, 325–333. [Google Scholar] [CrossRef]
- Imrie, B.; Shanmugasundaram, S. Source of variation in yield in international mungbean trials. Field Crops Res. 1987, 16, 197–208. [Google Scholar] [CrossRef]
- Chapman, S.C.; Cooper, M.; Hammer, G.L. Using crop simulation to generate genotype by environment interaction effects for sorghum in water-limited environments. Aust. J. Agric. Res. 2002, 53, 379–389. [Google Scholar] [CrossRef]
- Williams, R. A Study of the Causes of, and Selection for Resistance to, Weather Damage in Mungbean (Vigna radiata (L.) Wilczek; V. mungo (L.) Hepper). Ph.D. Thesis, The University of Queensland, Brisbane, Australia, 1989. [Google Scholar]
- Poehlman, J. The Mung Bean; Oxford and IBH Publication Co. Pvt. Ltd.: New Delhi, India, 1991; pp. 27–29. [Google Scholar]
- Kaur, R.; Bains, T.; Bindumadhava, H.; Nayyar, H. Responses of mungbean (Vigna radiata L.) genotypes to heat stress: Effects on reproductive biology, leaf function and yield traits. Sci. Hortic. 2015, 197, 527–541. [Google Scholar] [CrossRef]
- Sharma, L.; Priya, M.; Bindumadhava, H.; Nair, R.; Nayyar, H. Influence of high temperature stress on growth, phenology and yield performance of mungbean [Vigna radiata (L.) Wilczek] under managed growth conditions. Sci. Hortic. 2016, 213, 379–391. [Google Scholar] [CrossRef]
- Pookpakdi, A.; Pataradilok, H. Response of genotypes of mungbean and blackgram to planting dates and plant population densities. Kasetsart J. Nat. Sci 1993, 27, 395–400. [Google Scholar]
- Patriyawaty, N.R.; Rachaputi, R.C.; George, D. Physiological mechanisms underpinning tolerance to high temperature stress during reproductive phase in mungbean (Vigna radiata (L.) Wilczek). Environ. Exp. Bot. 2018, 150, 188–197. [Google Scholar] [CrossRef]
- Ali, M.; Kumar, S. Prospects of mungbean in rice-wheat cropping systems in Indo-Gangetic Plains of India. In Improving Income and Nutrition by Incorporating Mungbean in the Cereal Fallows of the Indo-Gangetic Plains of South Asia; Proceedings of the Final Workshop and Planning Meeting; AVRDC—The World Vegetable Center: Tainan, Taiwan, 2004; pp. 246–254. [Google Scholar]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chauhan, Y.S.; Williams, R. Physiological and Agronomic Strategies to Increase Mungbean Yield in Climatically Variable Environments of Northern Australia. Agronomy 2018, 8, 83. https://doi.org/10.3390/agronomy8060083
Chauhan YS, Williams R. Physiological and Agronomic Strategies to Increase Mungbean Yield in Climatically Variable Environments of Northern Australia. Agronomy. 2018; 8(6):83. https://doi.org/10.3390/agronomy8060083
Chicago/Turabian StyleChauhan, Yashvir S., and Rex Williams. 2018. "Physiological and Agronomic Strategies to Increase Mungbean Yield in Climatically Variable Environments of Northern Australia" Agronomy 8, no. 6: 83. https://doi.org/10.3390/agronomy8060083
APA StyleChauhan, Y. S., & Williams, R. (2018). Physiological and Agronomic Strategies to Increase Mungbean Yield in Climatically Variable Environments of Northern Australia. Agronomy, 8(6), 83. https://doi.org/10.3390/agronomy8060083