
Development of a Statistical Crop Model to Explain the Relationship between Seed Yield and Phenotypic Diversity within the Brassica napus Genepool

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
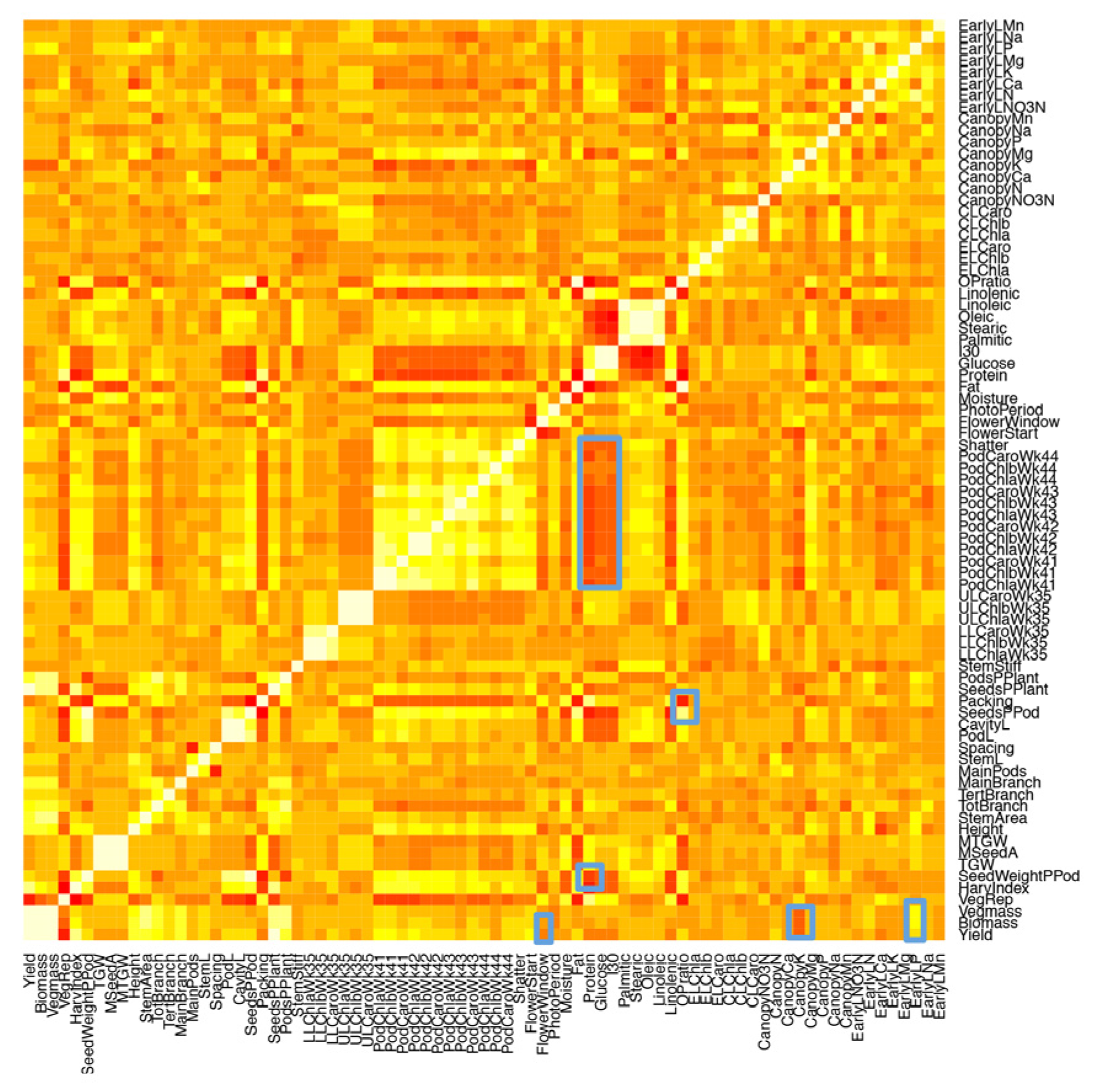
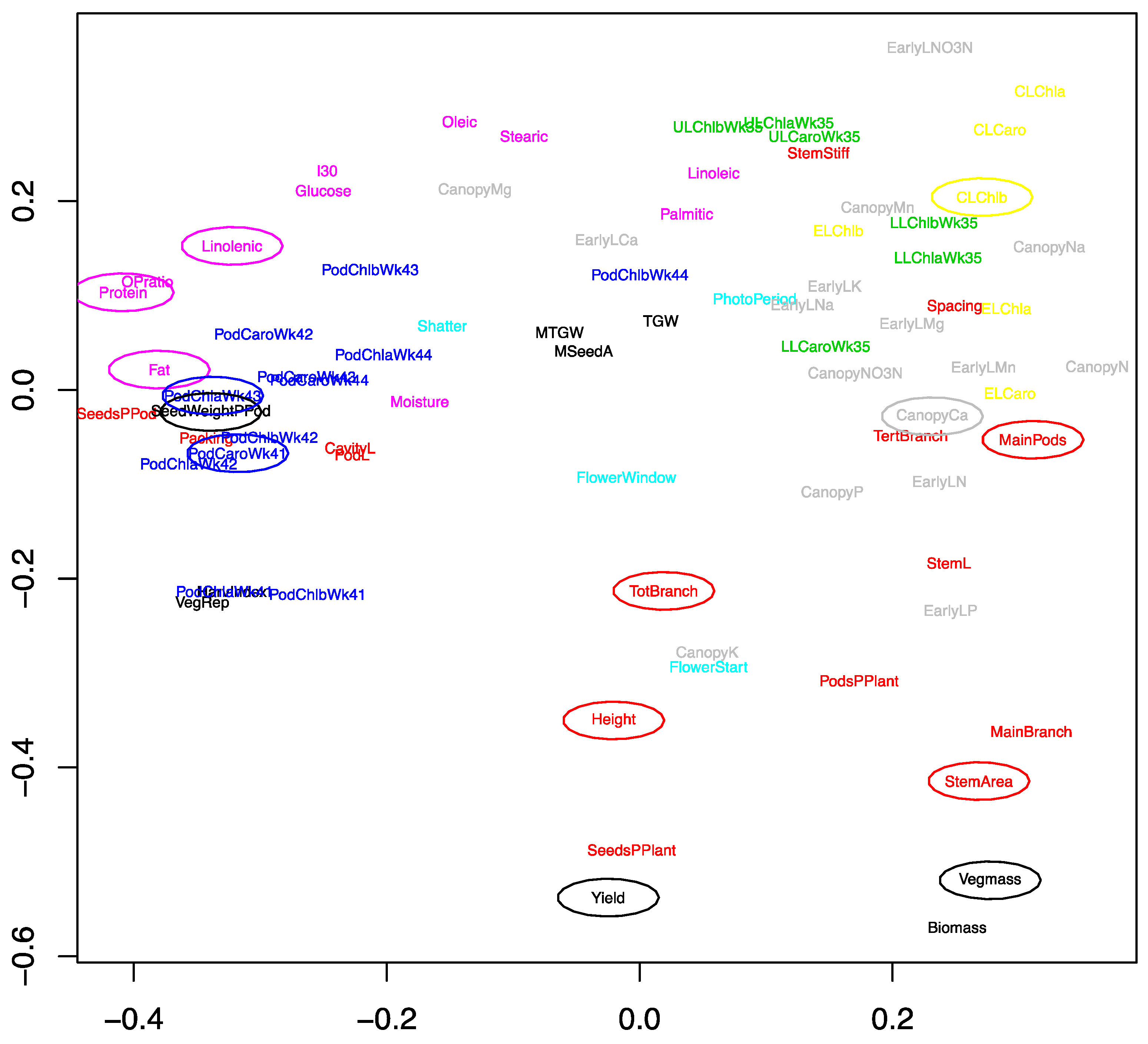
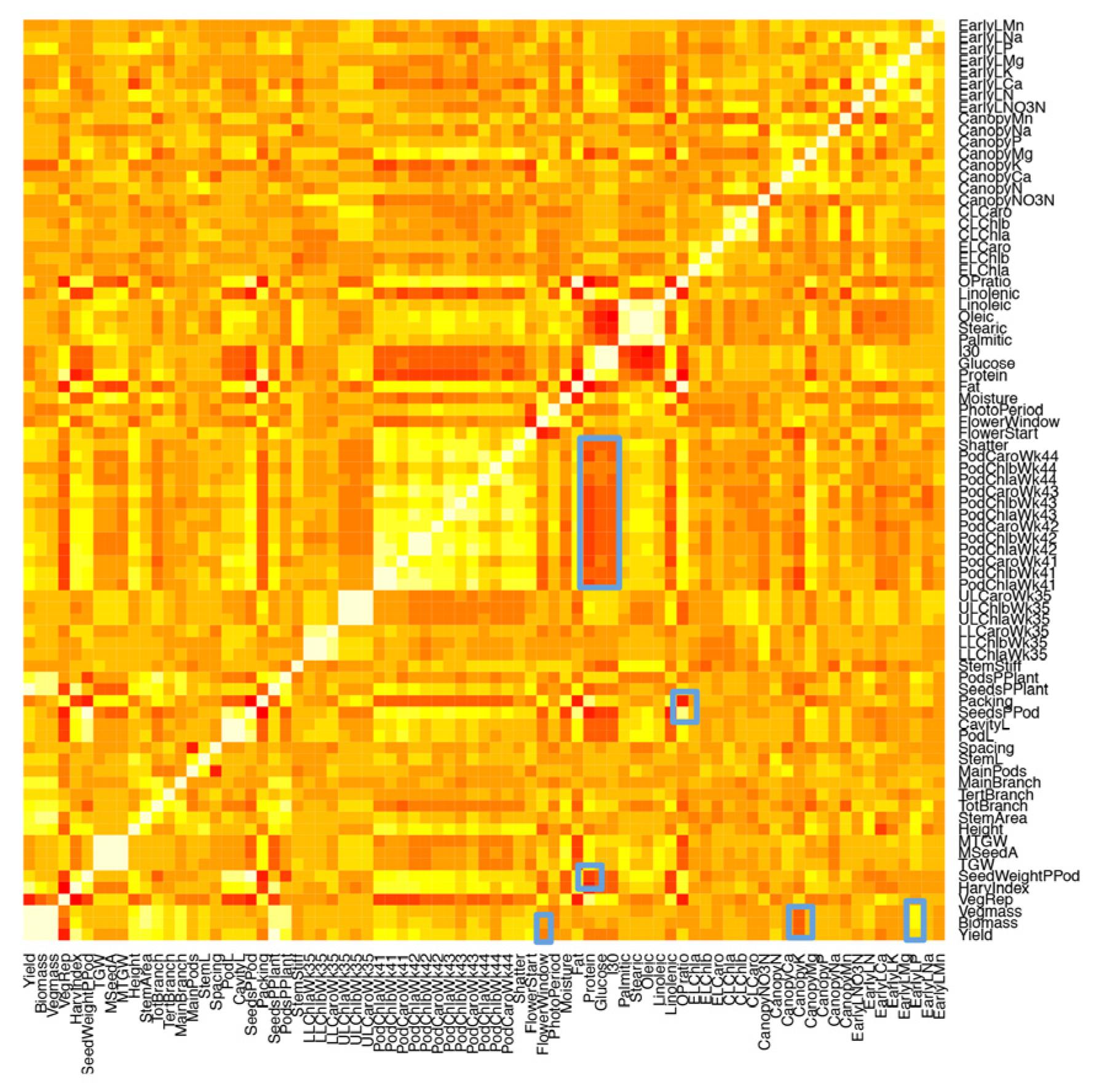
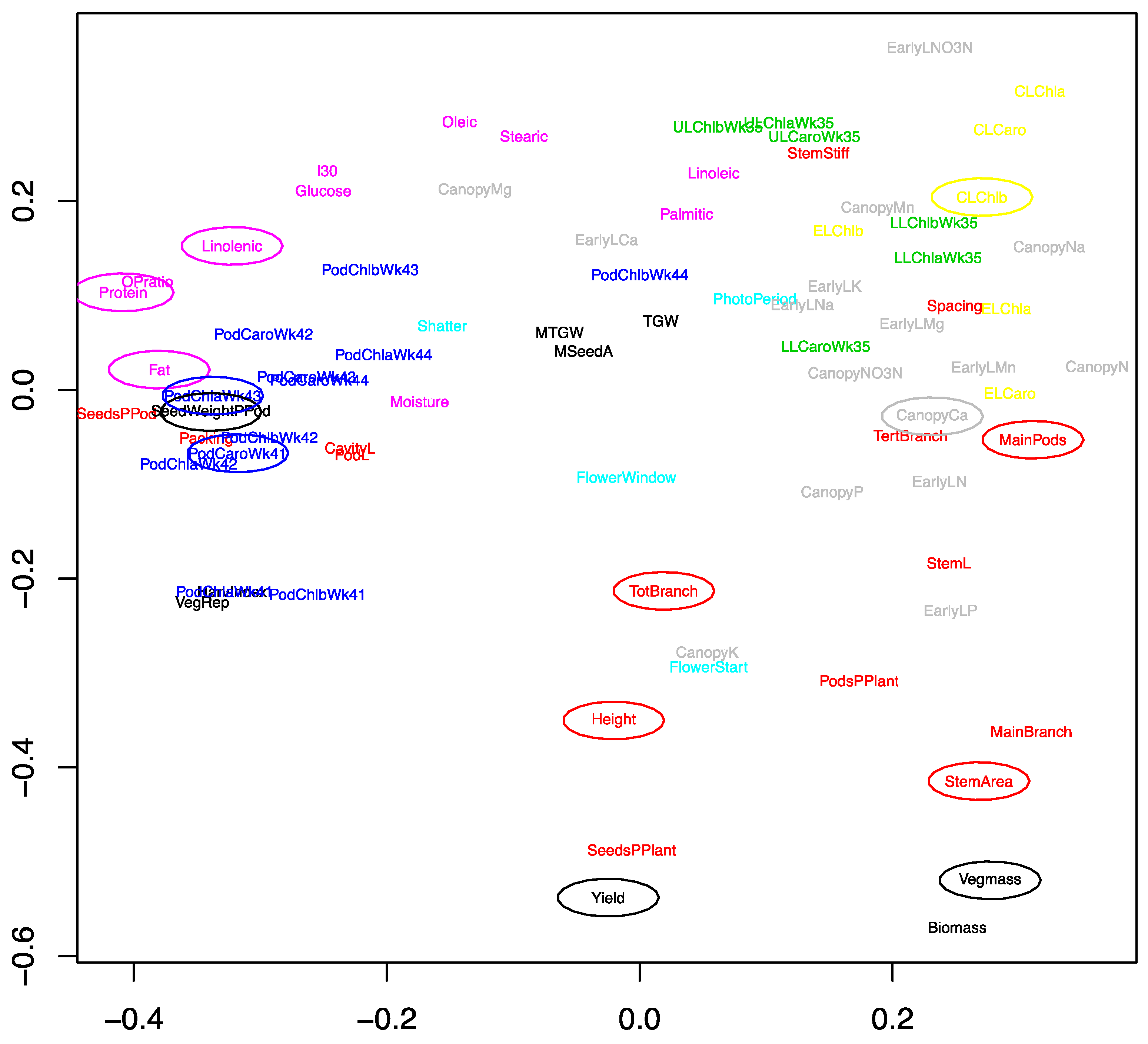
2.1. Development of an Efficient Statistical Model to Explain Yield
2.2. Genetic Factors Are Stronger Predictors of Yield than Environmental Factors within Year
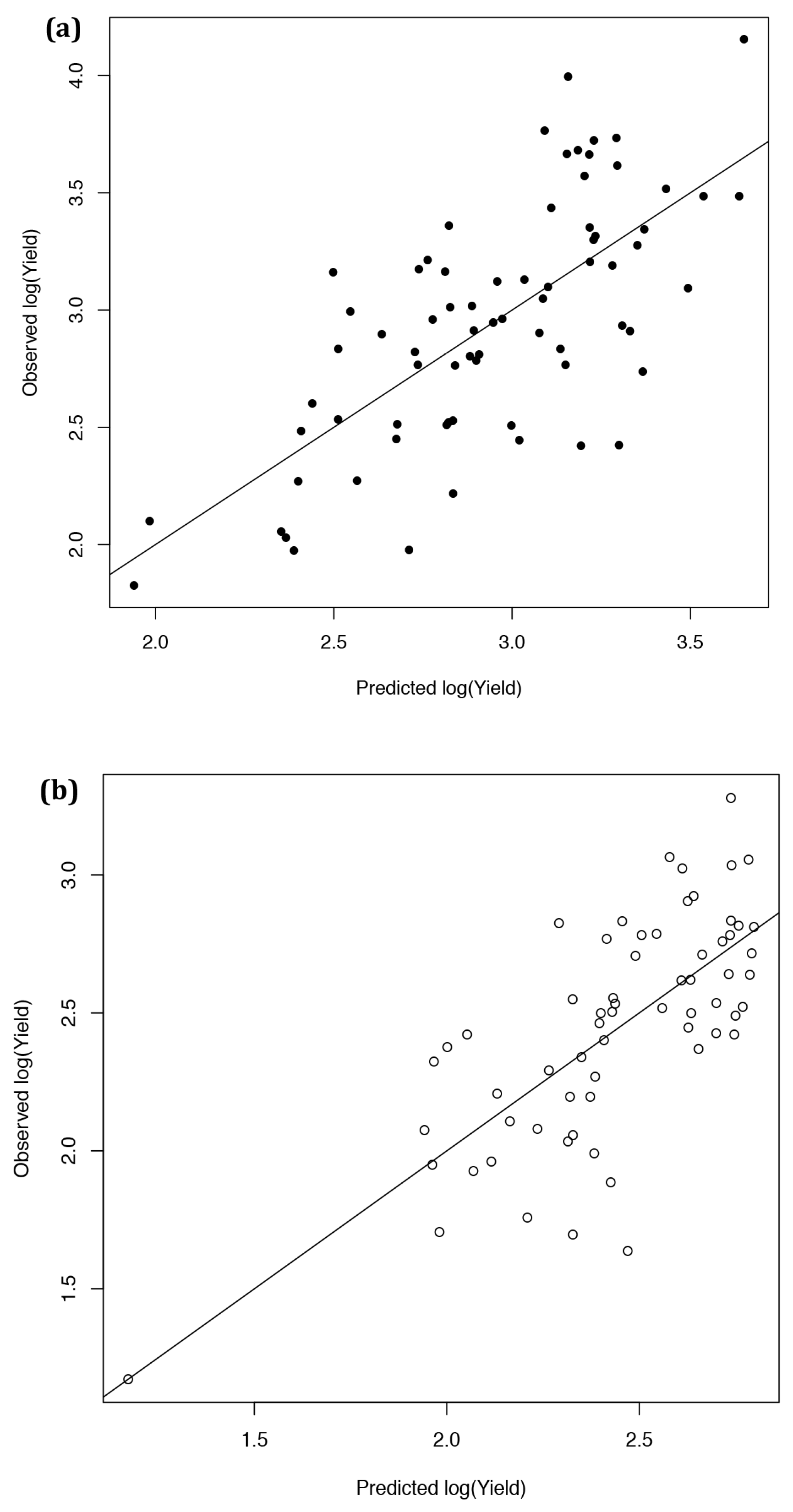
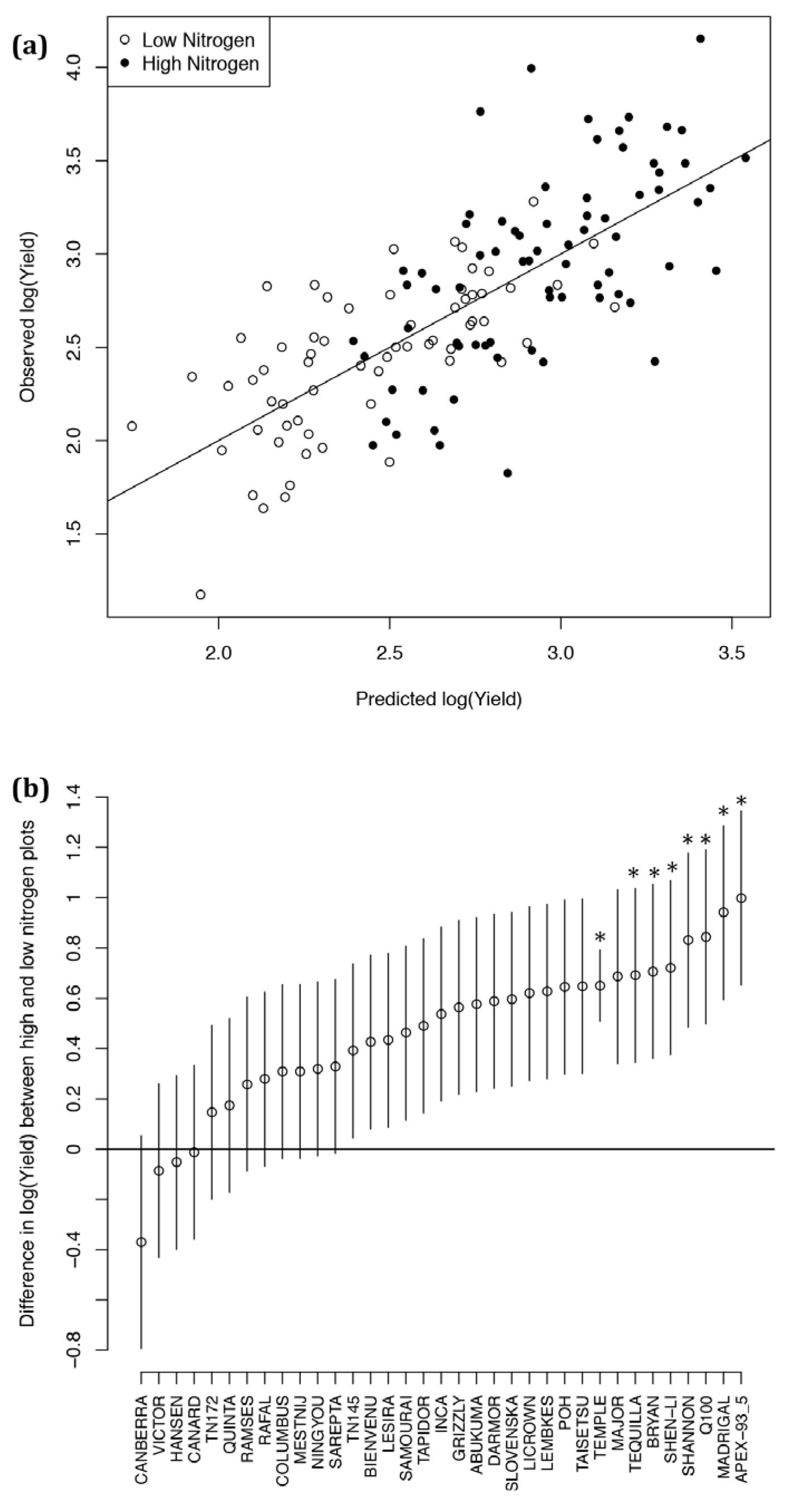
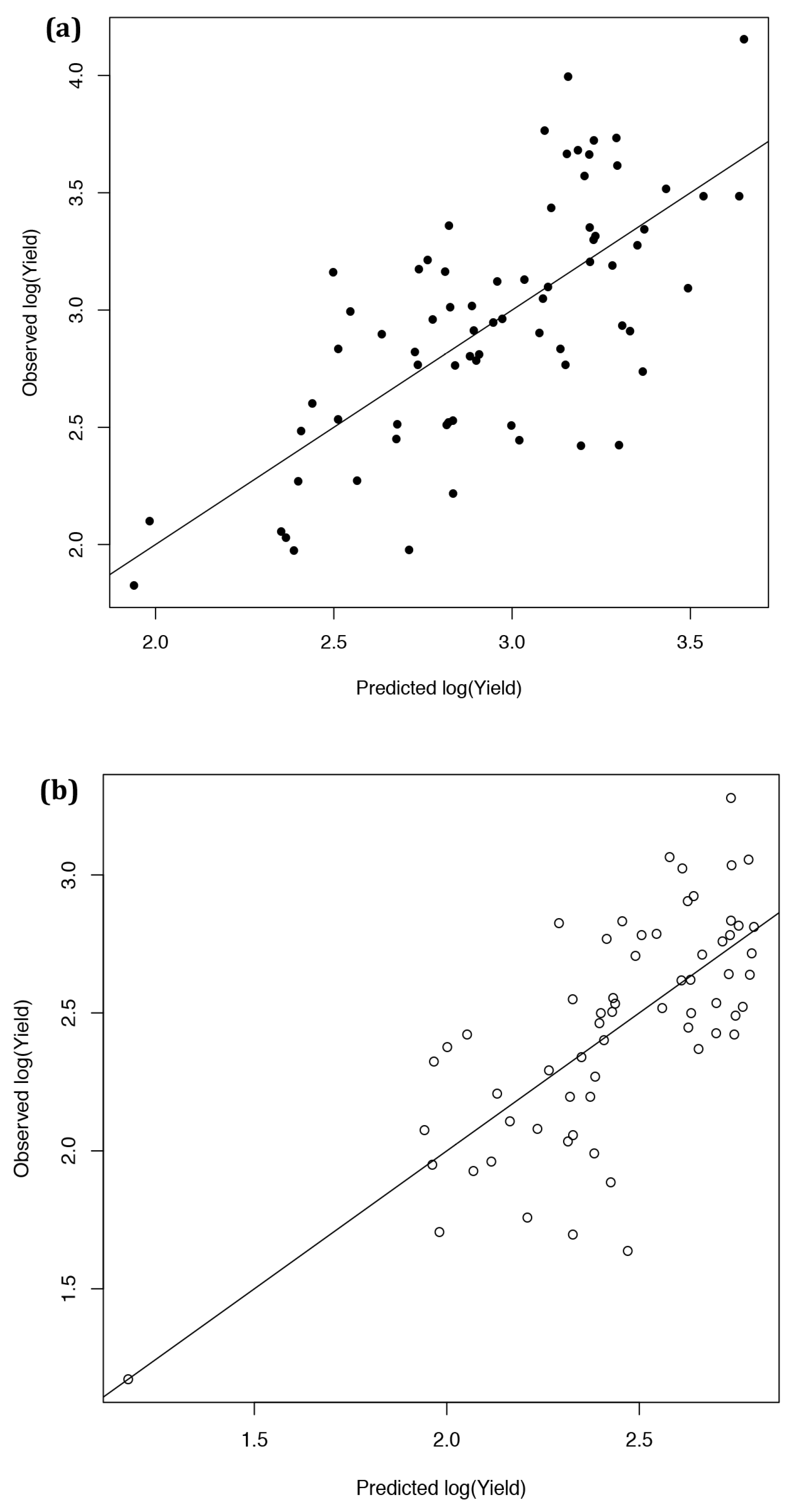
2.3. Developing an Improved Model to Explain Yield
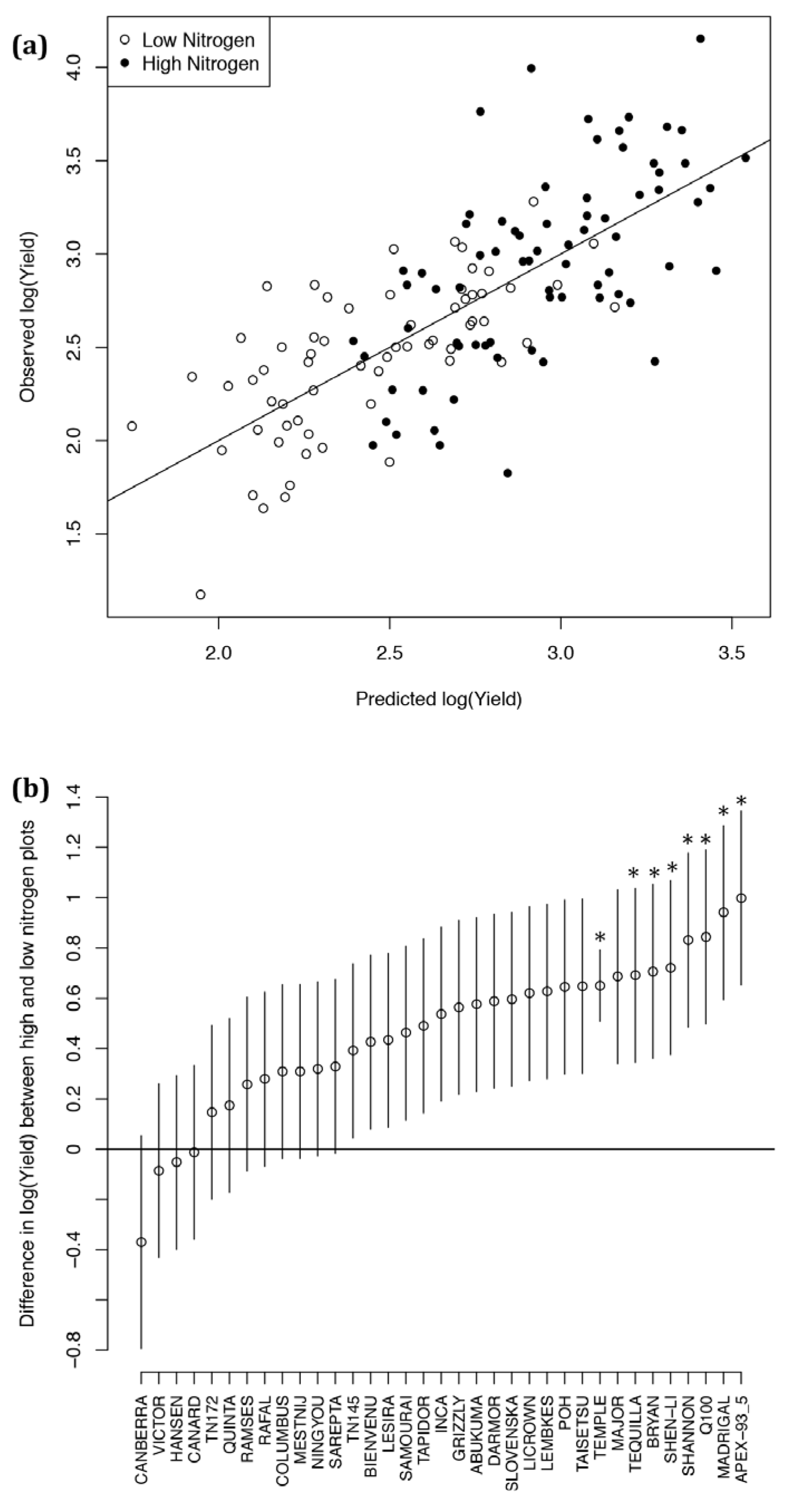
2.4. Ten-Fold Cross-Validation of the Year 2 Models
2.5. Seed Oil Content Rather Than Protein Drives Seed Yield
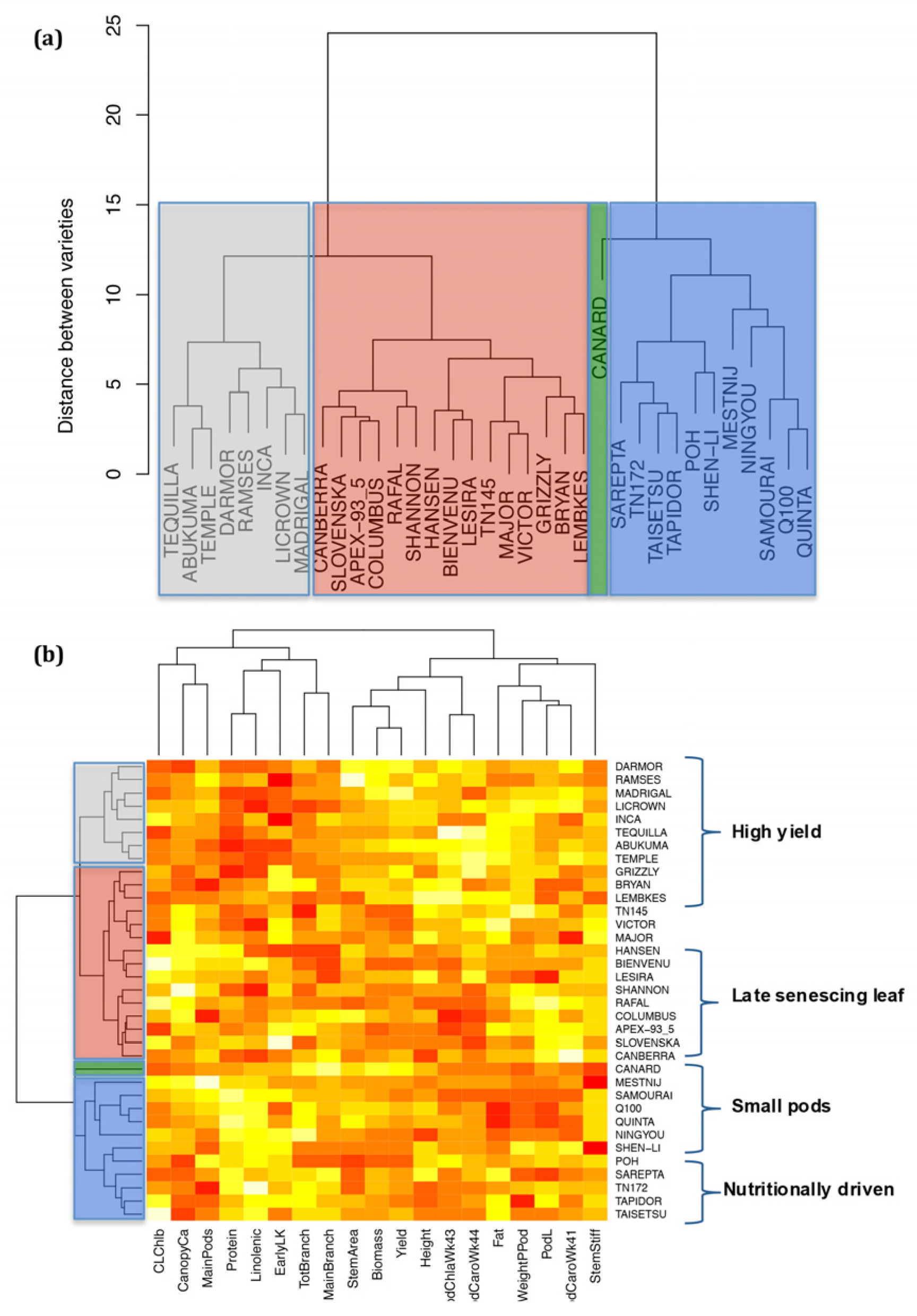
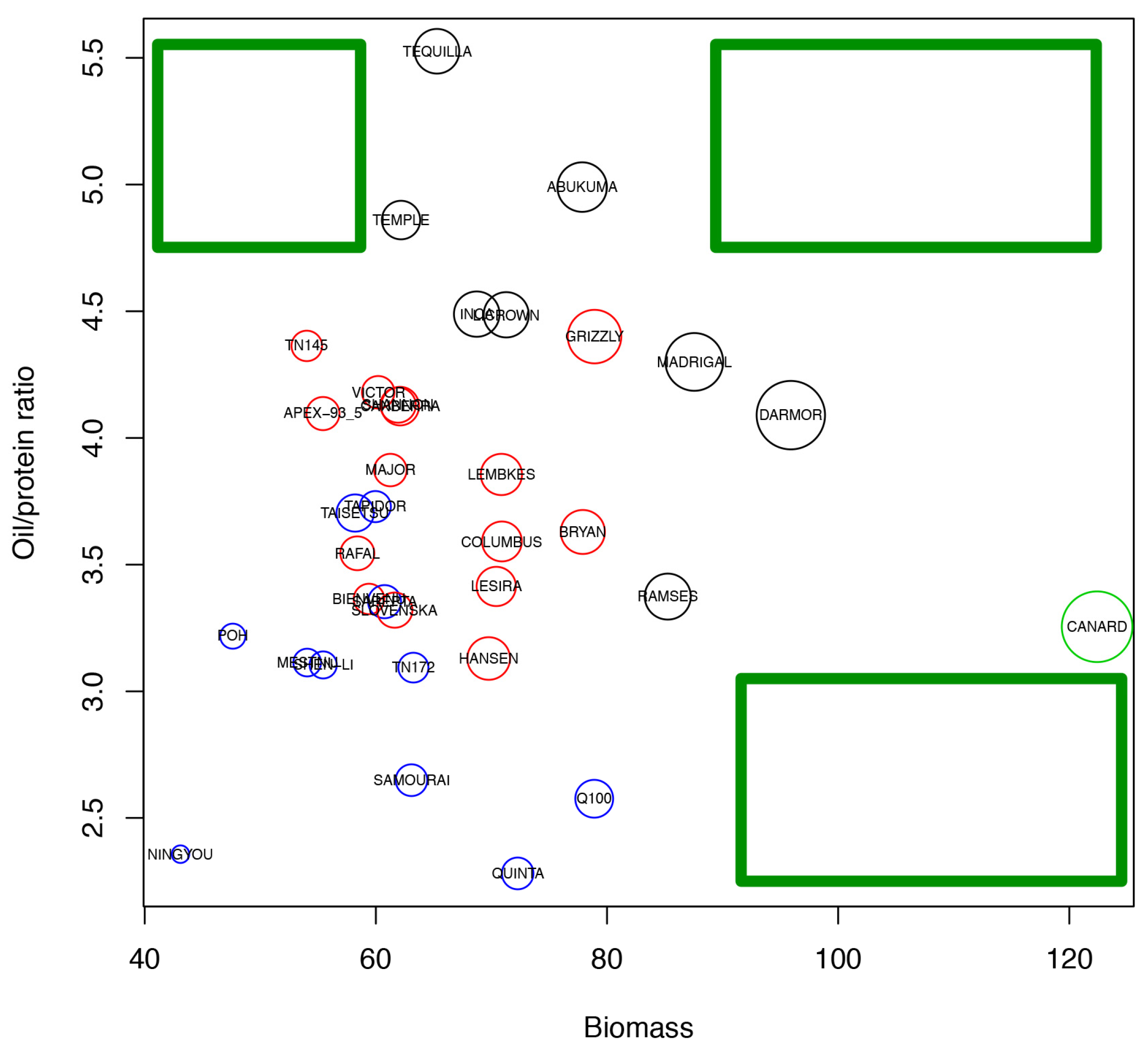
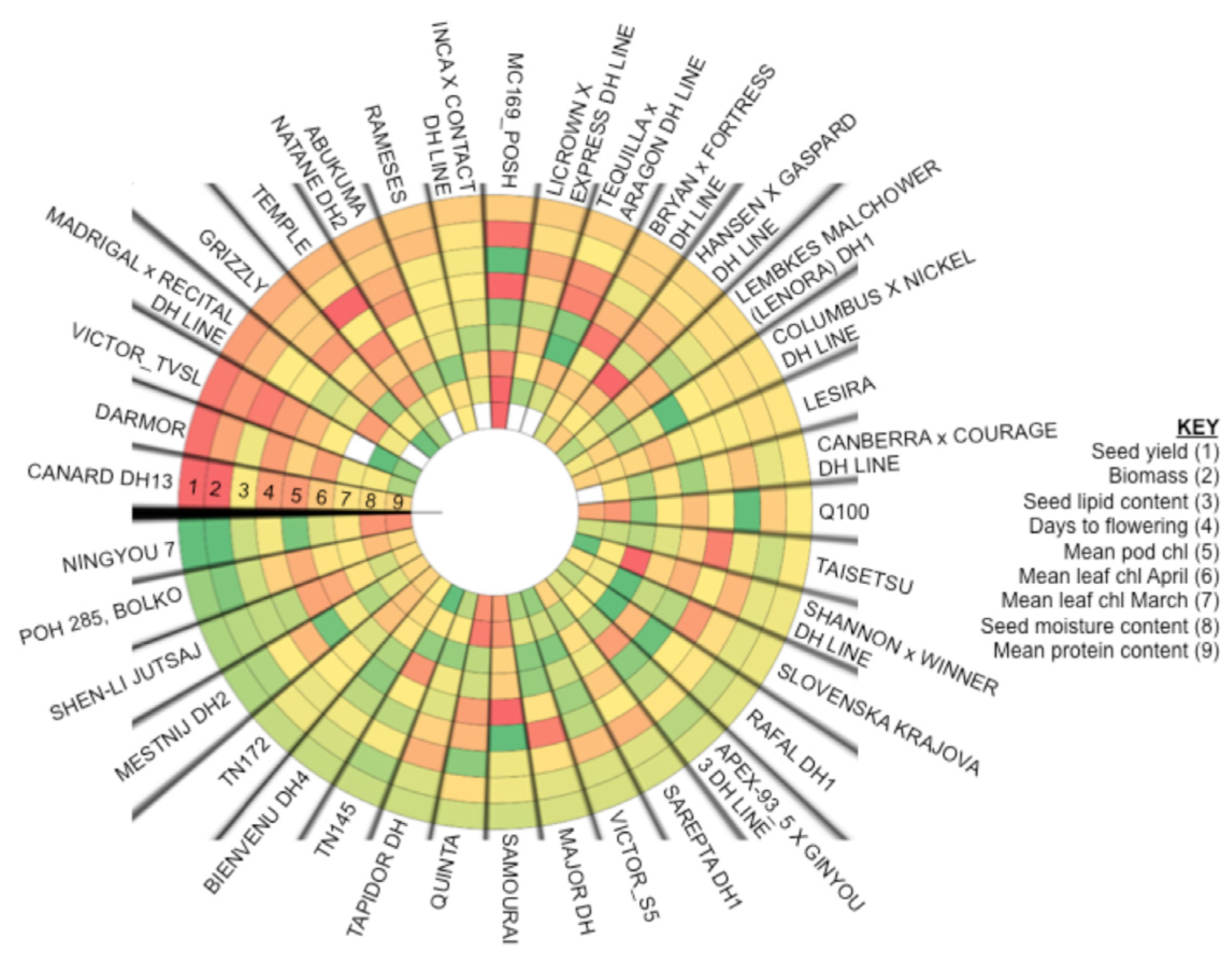
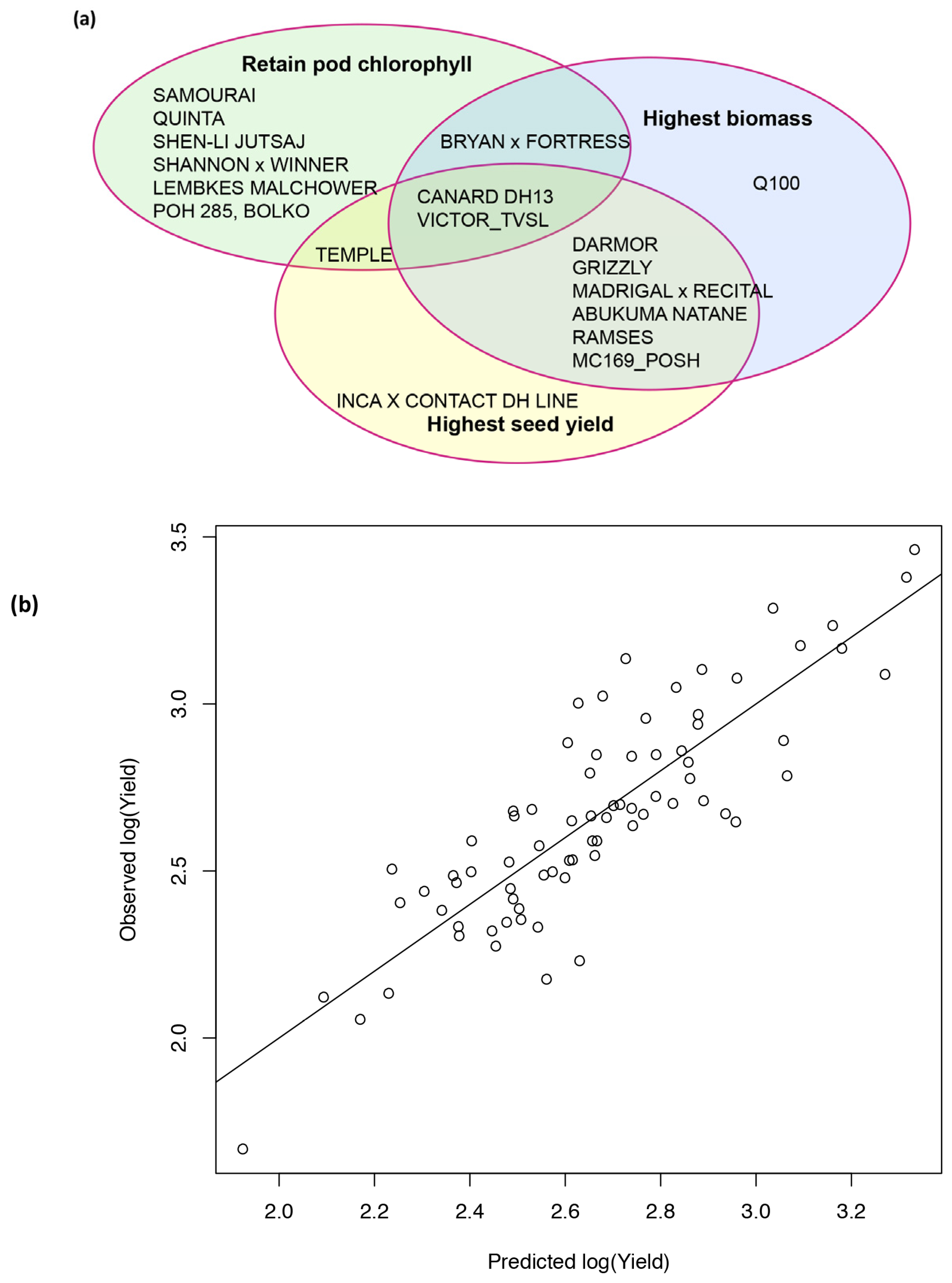
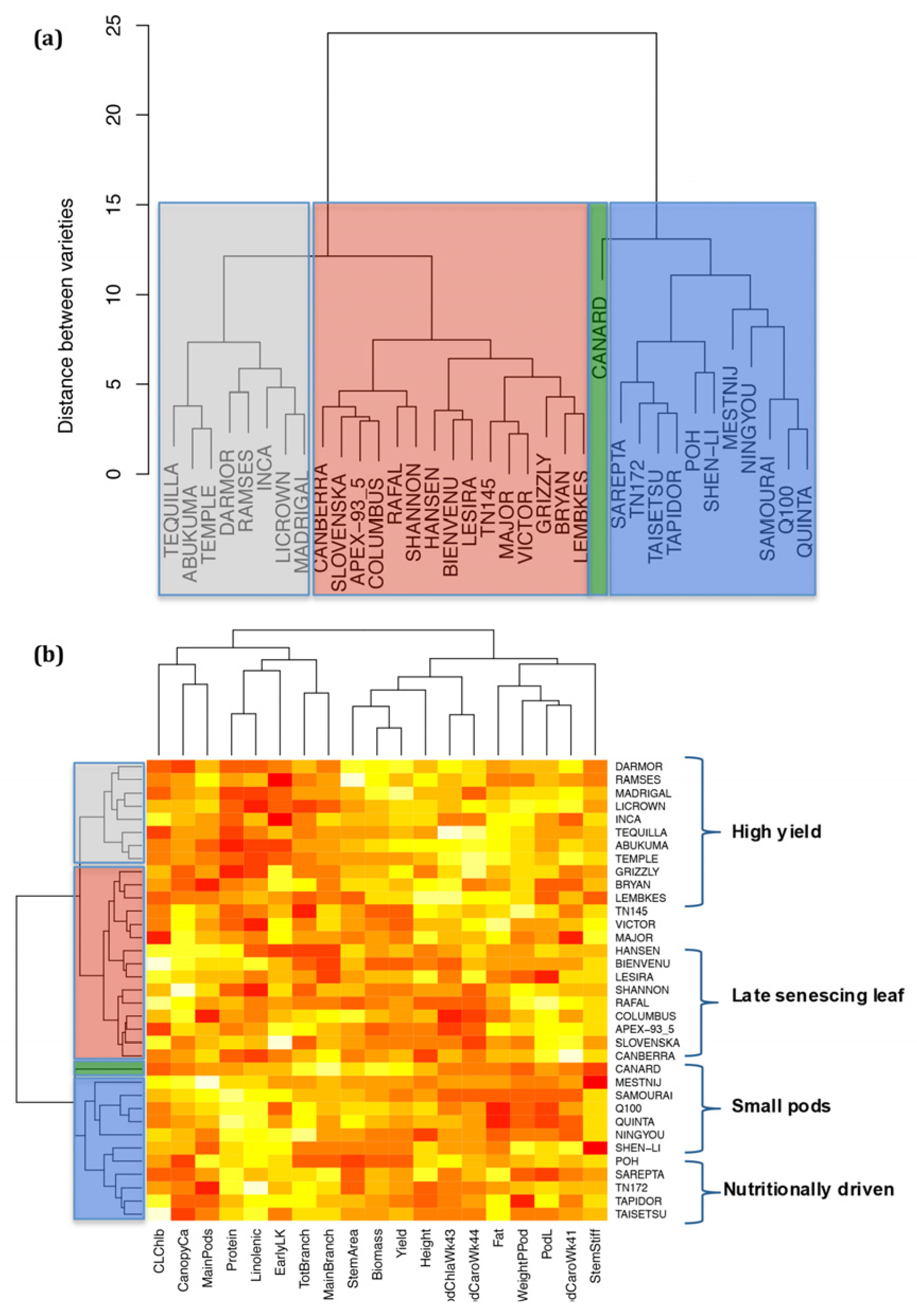
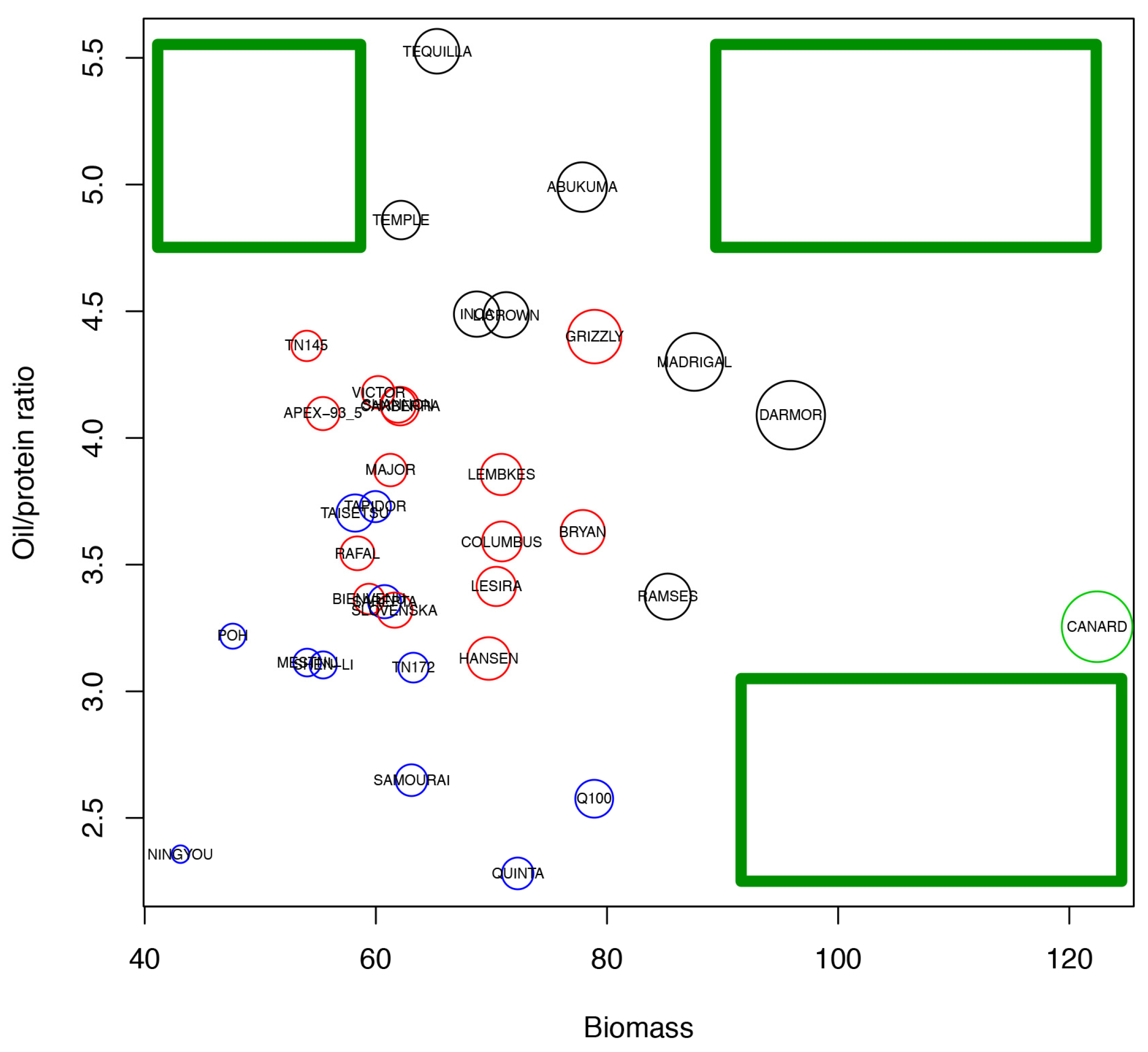
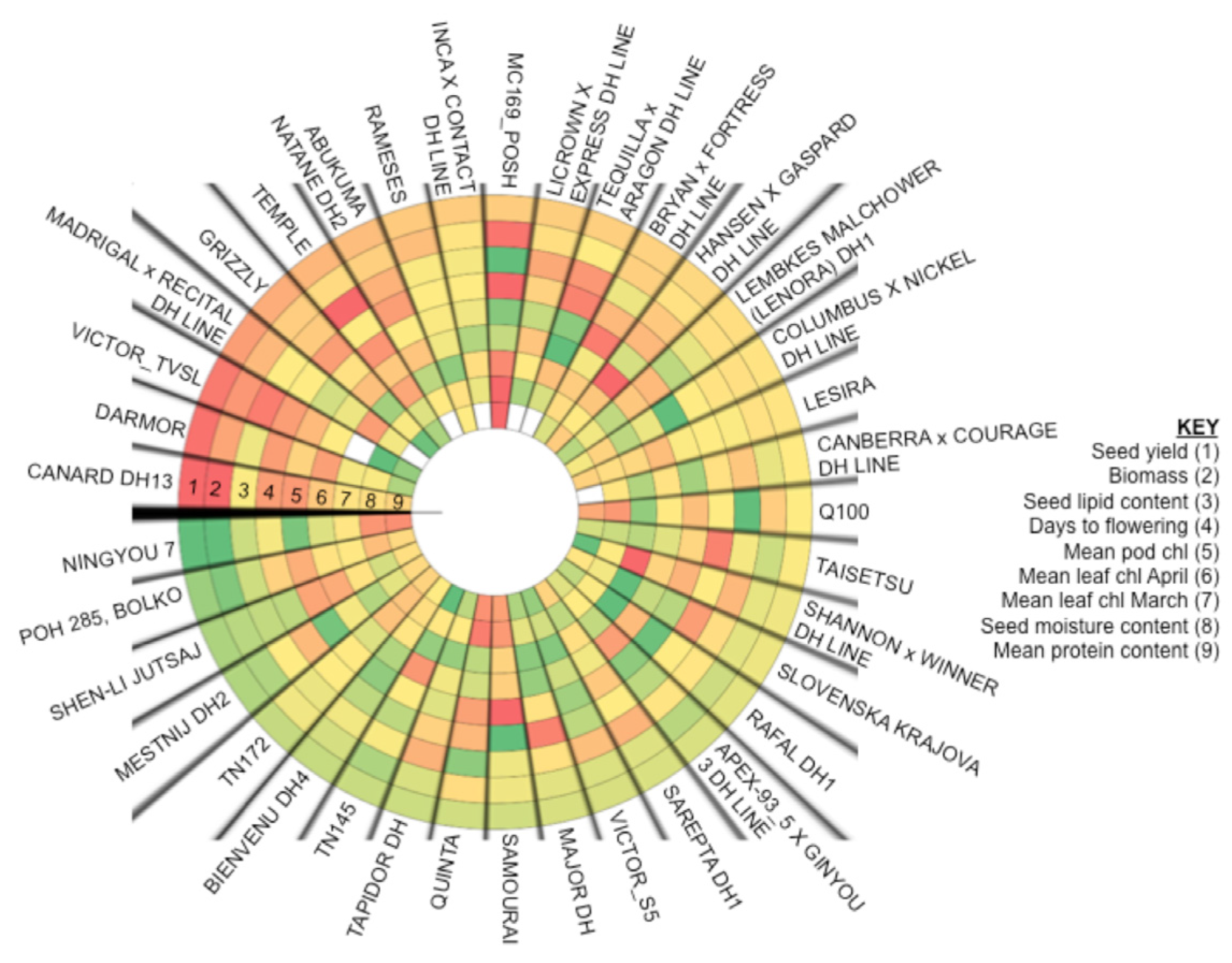
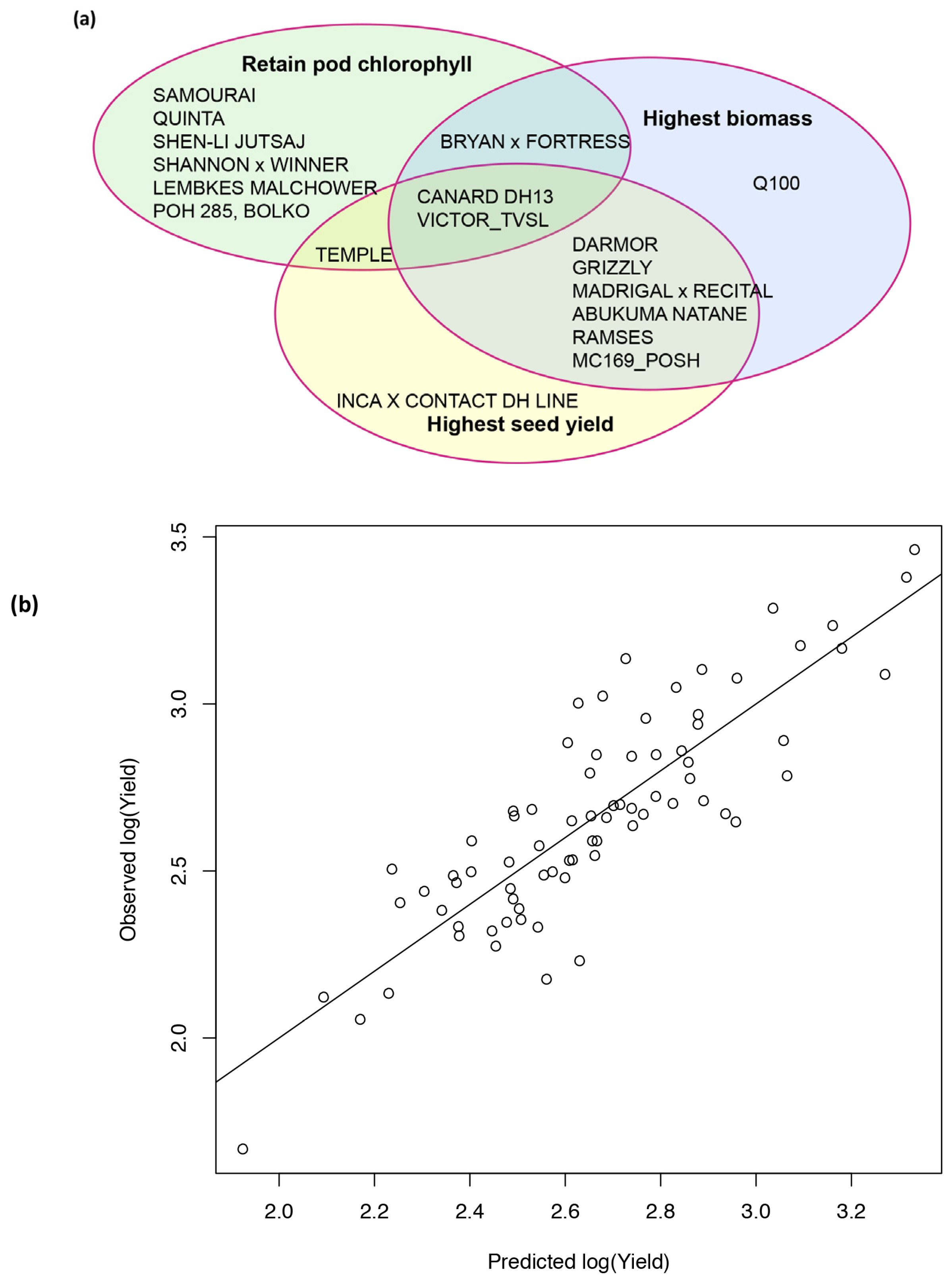
2.6. Analysis of Varieties to Determine Those Produce High Yields and Highlight Gaps in Breeding Potential
2.7. Traits Measured Early in Crop Development Are Also Good Predictors of Yield
3. Discussion
3.1. Greater Branching Density Drives Yield Improvement
3.2. Increasing Seed Oil:Protein Ratio
3.3. Early Leaf P Status Correlates with Final Yield
3.4. Number of Pods Determines Yield
3.5. Flowering Start Date and Duration Do Not Affect Seed Yield
3.6. The Relationship between Plant Senescence and Seed Yield
4. Materials and Methods
4.1. Site Description and Meteorological Conditions
4.2. Crop Management and Field Trial Design
4.3. Chlorophyll Measurements
4.4. Leaf Mineral Analysis
4.5. Plant Development
4.6. Plant Architecture
4.7. Pod Physiology
4.8. Yield
4.9. Seed Composition
4.10. Statistics
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wang, E.; Engel, T. Simulation of phenological development of wheat crops. Agric. Syst. 1998, 58, 1–24. [Google Scholar] [CrossRef]
- Yang, H.S.; Dobermann, A.; Lindquist, J.L.; Walters, D.T.; Arkebauer, T.J.; Cassman, K.G. Hybrid-maize—A maize simulation model that combines two crop modeling approaches. Field Crops Res. 2004, 87, 131–154. [Google Scholar] [CrossRef]
- Son, N.T.; Chen, C.F.; Chen, C.R.; Minh, V.Q.; Trung, N.H. A comparative analysis of multitemporal MODIS EVI and NDVI data for large-scale rice yield estimation. Agric. For. Meteorol. 2014, 197, 52–64. [Google Scholar] [CrossRef]
- Habekotte, B. Evaluation of seed yield determining factors of winter oilseed rape (Brassica napus L.) by means of crop growth modelling. Field Crops Res. 1997, 54, 137–151. [Google Scholar] [CrossRef]
- Habekotte, B. A model of the phenological development of winter oilseed rape (Brassica napus L.). Field Crops Res. 1997, 54, 127–136. [Google Scholar] [CrossRef]
- Habekotte, B. Options for increasing seed yield of winter oilseed rape (Brassica napus L.): A simulation study. Field Crops Res. 1997, 54, 109–126. [Google Scholar] [CrossRef]
- Deligios, P.A.; Farci, R.; Sulas, L.; Hoogenboom, G.; Ledda, L. Predicting growth and yield of winter rapeseed in a Mediterranean environment: Model adaptation at a field scale. Field Crops Res. 2013, 144, 100–112. [Google Scholar] [CrossRef]
- Husson, F.; Wallach, D.; Vandeputte, B. Evaluation of CECOL, a model of winter rape (Brassica napus L.). Eur. J. Agron. 1998, 8, 205–214. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Jauhiainen, L.; Hyovela, M.; Nissila, E. Trade-off between oil and protein in rapeseed at high latitudes: Means to consolidate protein crop status? Field Crops Res. 2011, 121, 248–255. [Google Scholar] [CrossRef]
- Nesi, N.; Delourme, R.; Bregeon, M.; Falentin, C.; Renard, M. Genetic and molecular approaches to improve nutritional value of Brassica napus L. seed. Comptes Rendus Biol. 2008, 331, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Smooker, A.M.; Wells, R.; Morgan, C.; Beaudoin, F.; Cho, K.; Fraser, F.; et al. The identification and mapping of candidate genes and QTL involved in the fatty acid desaturation pathway in Brassica napus. Theor. Appl. Genet. 2011, 122, 1075–1090. [Google Scholar] [CrossRef] [PubMed]
- Chalhoub, B.; Denoeud, F.; Liu, S.; Parkin, I.A.P.; Tang, H.; Wang, X.; Chiquet, J.; Belcram, H.; Tong, C.; Samans, B.; et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Gressel, J. Genetic Glass Ceilings: Transgenics for Crop Biodiversity, 1st ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2008; p. 488. [Google Scholar]
- FAOSTAT. Available online: http://faostat3.fao.org/faostat-gateway/go/to/download/Q/QC/E (accessed on 31 October 2015).
- Mawson, R.; Heaney, R.K.; Zdunczyk, Z.; Kozlowska, H. Rapeseed meal-glucosinolates and their antinutritional effects. Part II. Flavor and palatability. Mol. Nutr. Food Res. 1993, 37, 336–344. [Google Scholar]
- Quijada, P.A.; Udall, J.A.; Lambert, B.; Osborn, T.C. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L.): 1. Identification of genomic regions from winter germplasm. Theor. Appl. Genet. 2006, 113, 549–561. [Google Scholar] [CrossRef] [PubMed]
- Masclaux-Daubresse, C.; Chardon, F. Exploring nitrogen remobilization for seed filling using natural variation in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 2131–2142. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. The role of the pod in seed development: Strategies for manipulating yield. New Phytol. 2011, 190, 838–853. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. Manipulating resource allocation in plants. J. Exp. Bot. 2012, 63, 3391–3400. [Google Scholar] [CrossRef] [PubMed]
- Byzova, M.; Verduyn, C.; Brouwer, D.; Block, M. Transforming petals into sepaloid organs in Arabidopsis and oilseed rape: Implementation of the hairpin RNA-mediated gene silencing technology in an organ-specific manner. Planta 2004, 218, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Buchanan-Wollaston, V.; Earl, S.; Harrison, E.; Mathas, E.; Navabpour, S.; Page, T.; Pink, D. The molecular analysis of leaf senescence—A genomics approach. Plant Biotechnol. J. 2003, 1, 3–22. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S.; Parlitz, S.; Mueller-Roeber, B.; Meyer, R.C. Natural developmental variations in leaf and plant senescence in Arabidopsis thaliana. Plant Biol. 2008, 10, 136–147. [Google Scholar] [CrossRef] [PubMed]
- Levey, S.; Wingler, A. Natural variation in the regulation of leaf senescence and relation to other traits in Arabidopsis. Plant Cell Environ. 2005, 28, 223–231. [Google Scholar] [CrossRef]
- Flood, P.J.; Harbinson, J.; Aarts, M.G.M. Natural genetic variation in plant photosynthesis. Trends Plant Sci. 2011, 16, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, C.; Yang, T.J.W.; Stead, A.D.; Buchanan-Wollaston, V.; Roberts, J.A. A molecular and structural characterization of senescing Arabidopsis siliques and comparison of transcriptional profiles with senescing petals and leaves. Plant J. 2009, 57, 690–705. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.J.; Thurling, N. Growth, development, and yield of three oilseed Brassica species in a water-limited environment. Aust. J. Exp. Agric. 1994, 34, 93–103. [Google Scholar] [CrossRef]
- Robinson, C.K.; Hill, S.A. Altered resource allocation during seed development in Arabidopsis caused by the abi3 mutation. Plant Cell Environ. 1999, 22, 117–123. [Google Scholar] [CrossRef]
- Diaz, C.; Lemaitre, T.; Christ, A.; Azzopardi, M.; Kato, Y.; Sato, F.; Morot-Gaudry, J.-F.; Le Dily, F.; Masclaux-Daubresse, C. Nitrogen recycling and remobilization are differentially controlled by leaf senescence and development stage in Arabidopsis under low nitrogen nutrition. Plant Physiol. 2008, 147, 1437–1449. [Google Scholar] [CrossRef] [PubMed]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. Use of mutants to dissect the role of ethylene signalling in organ senescence and the regulation of yield in Arabidopsis thaliana. J. Plant Growth Regul. 2014, 33, 56–65. [Google Scholar] [CrossRef]
- Hopkins, C.J.; Welham, S.J.; Teakle, G.R.; Peplow, K.-S.; Pink, D.; Carion, P.W.C.; King, G.J.; Barker, J. Oilseed Rape Genetic Improvement Network Diversity Demonstration Trial—Year 1; Rothamsted Research: Harpenden, UK, 2009–2010. [Google Scholar]
- Hopkins, C.J.; Welham, S.J.; Teakle, G.R.; Peplow, K.-S.; Pink, D.; Carion, P.W.C.; King, G.J.; Barker, J. Oilseed Rape Genetic Improvement Network Diversity Demonstration Trial—Year 2; Rothamsted Research: Harpenden, UK, 2010–2011. [Google Scholar]
- Ali, N.; Javidfar, F.; Elmira, J.Y.; Mirza, A.M.Y. Relationship among yield components and selection criteria for yield improvement in winter rapeseed (Brassica napus L.). Pak. J. Bot. 2003, 35, 167–174. [Google Scholar]
- Allen, E.J.; Morgan, D.G. A quantitative analysis of the effects of nitrogen on the growth, development and yield of oilseed rape. J. Agric. Sci. 1972, 78, 315–324. [Google Scholar] [CrossRef]
- Lu, G.-Y.; Zhang, F.; Zheng, P.-Y.; Cheng, Y.; Liu, F.-I.; Fu, G.-P.; Zhang, X.-K. Relationship among yield components and selection criteria for yield improvement in early rapeseed (Brassica napus L.). Agric. Sci. China 2011, 10, 997–1003. [Google Scholar] [CrossRef]
- Berry, P.M.; Spink, J.H. A physiological analysis of oilseed rape yields: Past and future. J. Agric. Sci. 2006, 144, 381–392. [Google Scholar] [CrossRef]
- Gabrielle, B.; Denoroy, P.; Gosse, G.; Justes, E.; Andersen, M.N. A model of leaf area development and senescence for winter oilseed rape. Field Crops Res. 1998, 57, 209–222. [Google Scholar] [CrossRef]
- Diepenbrock, W. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crops Res. 2000, 67, 35–49. [Google Scholar] [CrossRef]
- Bennett, E.J.; Gawthrop, F.; Yao, C.; Boniface, C.; Ishihara, H.; Roberts, J.A.; Wagstaff, C. Effects of planting density and nitrogen application on seed yield and other morphological traits of the leafy vegetable kale (Brassica oleracea). Ann. Appl. Biol. 2013, 119, 201–216. [Google Scholar]
- Navabi, A.; Iqbal, M.; Strenzke, K.; Spaner, D. The relationship between lodging and plant height in a diverse wheat population. Can. J. Plant Sci. 2006, 86, 723–726. [Google Scholar] [CrossRef]
- Islam, N.; Evans, E.J. Influence of lodging and nitrogen rate on the yield and yield attributes of oilseed rape (Brassica napus L.). Theor. Appl. Genet. 1994, 88, 530–534. [Google Scholar] [CrossRef] [PubMed]
- Al-Ahmad, H.; Dwyer, J.; Moloney, M.; Gressel, J. Mitigation of establishment of Brassica napus transgenes in volunteers using a tandem construct containing a selectively unfit gene. Plant Biotechnol. J. 2006, 4, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Leyser, O. Auxin, self-organisation, and the colonial nature of plants. Curr. Biol. 2011, 21, R331–R337. [Google Scholar] [CrossRef] [PubMed]
- Sachs, T.; Novoplansky, A.; Cohen, D. Plants as competing populations of redundant organs. Plant Cell Environ. 1993, 16, 765–770. [Google Scholar] [CrossRef]
- Grami, B.; Stefansson, B.R.; Baker, R.J. Genetics of protein and oil content in summer rape: Heritability, number of effective factors, and correlations. Can. J. Plant Sci. 1977, 57, 937–943. [Google Scholar] [CrossRef]
- Chardon, F.; Jasinski, S.; Durandet, M.; Lecureuil, A.; Soulay, F.; Bedu, M.; Guerche, P.; Masclaux-Daubresse, C. QTL meta-analysis in Arabidopsis reveals an interaction between leaf senescence and resource allocation to seeds. J. Exp. Bot. 2014, 65, 3949–3962. [Google Scholar] [CrossRef] [PubMed]
- Specht, J.E.; Hume, D.J.; Kumudini, S.V. Soybean yield potential—A genetic and physiological perspective. Crop Sci. 1999, 39, 1560–1570. [Google Scholar] [CrossRef]
- Petersson, A.; Thomsen, M.H.; Hauggaard-Nielsen, H.; Thomsen, A.-B. Potential bioethanol and biogas production using lignocellulosic biomass from winter rye, oilseed rape and faba bean. Biomass Bioenergy 2007, 31, 812–819. [Google Scholar] [CrossRef]
- Pandey, A.; Soccol, C.R.; Nigam, P.; Soccol, V.T. Biotechnological potential of agro-industrial residues. I: Sugarcane bagasse. Bioresour. Technol. 2000, 74, 69–80. [Google Scholar] [CrossRef]
- Orson, J.; Booth, E.; Merritt, C.; Lea, C. Growing High Oleic Low Linolenic (HOLL) Oilseed Rape for Specialised Markets; HGCA Project Report 442; AHDB Cereals & Oilseeds: Kenilworth, UK, 2008. [Google Scholar]
- Venegas-Caleron, M.; Sayanova, O.; Napier, J.A. An alternative to fish oils: Metabolic engineering of oil-seed crops to produce omega-3 long chain polyunsaturated fatty acids. Prog. Lipid Res. 2010, 49, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Betancor, M.B.; Sprague, M.; Usher, S.; Sayanova, O.; Campbell, P.J.; Napier, J.A.; Tocher, D.R. A nutritionally-enhanced oil from transgenic Camelina sativa effectively replaces fish oil as a source of eicosapentaenoic acid for fish. Sci. Rep. 2015, 5, 8104. [Google Scholar] [CrossRef] [PubMed]
- Vigeolas, H.; Waldeck, P.; Zank, T.; Geigenberger, P. Increasing seed oil content in oil-seed rape (Brassica napus L.) by over-expression of a yeast glycerol-3-phosphate dehydrogenase under the control of a seed-specific promoter. Plant Biotechnol. J. 2007, 5, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Brennan, R.F.; Bolland, M.D.A. Effect of fertiliser phosphorus and nitrogen on the concentrations of oil and protein in grain and the grain yield of canola (Brassica napus L.) grown in south-western Australia. Aust. J. Exp. Agric. 2007, 47, 984–991. [Google Scholar] [CrossRef]
- Brennan, R.F.; Bolland, M.D.A. Comparing the nitrogen and phosphorus requirements of canola and wheat for grain yield and quality. Crop Pasture Sci. 2009, 60, 566–577. [Google Scholar] [CrossRef]
- Krueger, K.; Goggi, A.S.; Mallarino, A.P.; Mullen, R.E. Phosphorus and potassium fertilization effects on soybean seed quality and composition. Crop Sci. 2013, 53, 602–610. [Google Scholar] [CrossRef]
- Lickfett, T.; Matthäus, B.; Velasco, L.; Möllers, C. Seed yield, oil and phytate concentration in the seeds of two oilseed rape cultivars as affected by different phosphorus supply. Eur. J. Agron. 1999, 11, 293–299. [Google Scholar] [CrossRef]
- Mourtzinis, S.; Arriaga, F.J.; Balkcom, K.S.; Ortiz, B.V. Corn grain and stover yield prediction at R1 growth stage. Agron. J. 2013, 105, 1045–1050. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, P.; Hong, D.; Huang, A.; Li, S.; He, Q.; Yang, G. Inheritance of seeds per silique in Brassica napus L. using joint segregation analysis. Field Crops Res. 2010, 116, 58–67. [Google Scholar] [CrossRef]
- Yang, P.; Shu, C.; Chen, L.; Xu, J.; Wu, J.; Liu, K. Identification of a major QTL for silique length and seed weight in oilseed rape (Brassica napus L.). Theor. Appl. Genet. 2012, 125, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Chay, P.; Thurling, N. Identification of genes controlling pod length in spring rapeseed, Brassica napus L.; and their utilization for yield improvement. Plant Breed. 1989, 103, 54–62. [Google Scholar] [CrossRef]
- Bommarco, R.; Marini, L.; Vaissière, B. Insect pollination enhances seed yield, quality, and market value in oilseed rape. Oecologia 2012, 169, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.M.; Rasmussen, H.B.; Birkett, M.A.; Murray, D.A.; Pye, B.J.; Watts, N.P. Behavioural and chemical ecology underlying the success of turnip rape (Brassica rapa) trap crops in protecting oilseed rape (Brassica napus) from the pollen beetle (Meligethes aeneus). Arthropod-Plant Interact. 2007, 1, 57–67. [Google Scholar] [CrossRef]
- Gan, S.S.; Amasino, R.M. Inhibition of leaf senescence by autoregulated production of cytokinin. Science 1995, 270, 1986–1988. [Google Scholar] [CrossRef] [PubMed]
- Verma, V.; Foulkes, M.J.; Worland, A.J.; Sylvester-Bradley, R.; Caligari, P.D.S.; Snape, J.W. Mapping quantitative trait loci for flag leaf senescence as a yield determinant in winter wheat under optimal and drought-stressed environments. Euphytica 2004, 135, 255–263. [Google Scholar] [CrossRef]
- Makino, A.; Osmond, B. Effects of nitrogen nutrition on nitrogen partitioning between chloroplasts and mitochondria in pea and wheat. Plant Physiol. 1991, 96, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Gifford, R.M.; Evans, L.T. Photosynthesis, carbon partitioning, and yield. Ann. Rev. Plant Physiol. 1981, 32, 485–509. [Google Scholar] [CrossRef]
- Brar, G.; Thies, W. Contribution of leaves, stem, siliques and seeds to dry matter accumulation in ripening seeds of rapeseed, Brassica napus L. Z. Pflanzenphysiol. 1977, 82, 1–13. [Google Scholar] [CrossRef]
- Nakano, H.; Muramatsu, S.; Makino, A.; Mae, T. Relationship between the suppression of photosynthesis and starch accumulation in the pod-removed bean. Aust. J. Plant Physiol. 2000, 27, 167–173. [Google Scholar]
- Hay, R.; Kirby, E. Convergence and synchrony-a review of the coordination of development in wheat. Aust. J. Agric. Res. 1991, 42, 661–700. [Google Scholar] [CrossRef]
- Price, J.S.; Hobson, R.N.; Neale, M.A.; Bruce, D.M. Seed losses in commercial harvesting of oilseed rape. J. Agric. Eng. Res. 1996, 65, 183–191. [Google Scholar] [CrossRef]
- Meakin, P.J.; Roberts, J.A. Anatomical and biochemical changes associated with the induction of oilseed rape (Brassica napus) pod dehiscence by Dasineura brassicae (Winn.). Ann. Bot. 1991, 67, 193–197. [Google Scholar] [CrossRef]
- Hensel, L.L.; Grbic, V.; Baumgarten, D.A.; Bleecker, A.B. Developmental and age-related processes that influence the longevity and senescence of photosynthetic tissues in Arabidopsis. Plant Cell 1993, 5, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Nooden, L.D.; Penney, J.P. Correlative controls of senescence and plant death in Arabidopsis thaliana (Brassicaceae). J. Exp. Bot. 2001, 52, 2151–2159. [Google Scholar] [CrossRef] [PubMed]
- Marbà, N.; Duarte, C.M.; Agustí, S. Allometric scaling of plant life history. Proc. Natl. Acad. Sci. USA 2007, 104, 15777–15780. [Google Scholar] [CrossRef] [PubMed]
- Broadley, M.R.; Hammond, J.P.; King, G.J.; Astley, D.; Bowen, H.C.; Meacham, M.C.; Mead, A.; Pink, D.A.; Teakle, G.R.; Hayden, R.M.; et al. Shoot calcium and magnesium concentrations differ between subtaxa, are highly heritable, and associate with potentially pleiotropic loci in Brassica oleracea. Plant Physiol. 2008, 146, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Lancashire, P.; Bleiholder, H.; Boom, T.; van den Bout-van den Beukel, C.J.P.; Langelüddeke, P.; Stauss, R.; Weber, E.; Velde, T.; Lancashire, P.D.; Boom, T.V.D.; et al. A uniform decimal code for growth stages of crops and weeds. Ann. Appl. Biol. 1991, 119, 561–601. [Google Scholar] [CrossRef]
- Kelly, A.A.; Shaw, E.; Powers, S.J.; Kurup, S.; Eastmond, P.J. Suppression of the SUGAR-DEPENDENT1 triacylglycerol lipase family during seed development enhances oil yield in oilseed rape (Brassica napus L.). Plant Biotechnol. J. 2013, 11, 355–361. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Coefficients | Estimate | Standard Error | t-Value | Pr (>|t|) | Significance | Cumulative % Variance Explained |
|---|---|---|---|---|---|---|
| (Intercept) | 2.472 | 0.285 | 8.672 | 1.42 × 10−14 | *** | - |
| Log(canopy leaf Chl a) | 0.491 | 0.179 | 2.740 | 0.0070 | ** | 28.81 |
| Seed linolenic acid | −0.058 | 0.016 | −3.635 | 0.0004 | *** | 44.05 |
| Seeds per pod | 0.019 | 0.008 | 2.287 | 0.0238 | * | 48.70 |
| Nitrogen applied to plot | 0.329 | 0.088 | 3.722 | 0.0003 | *** | 52.91 |
| Log(seed glucosinolate) | −0.067 | 0.031 | −2.200 | 0.0296 | * | 54.59 |
| High Nitrogen Coefficients | Estimate | Standard Error | t-Value | Pr (>|t|) | Significance | Cumulative % Variance Explained |
|---|---|---|---|---|---|---|
| (Intercept) | 1.649 | 0.833 | 1.980 | 0.0518 | . | - |
| Seed protein content | −0.051 | 0.018 | −2.767 | 0.0073 | ** | 33.77 |
| Log[early leaf Mn] | 0.632 | 0.223 | 2.836 | 0.0060 | ** | 40.51 |
| Pod dehiscence Low | 0.672 | 0.282 | 2.379 | 0.0202 | * | - |
| Pod dehiscence Medium | 0.731 | 0.278 | 0.268 | 0.0107 | * | 46.17 |
| Log[seed glucosinolate] | −0.108 | 0.054 | −2.019 | 0.0475 | * | 49.26 |
| Low Nitrogen Coefficients | ||||||
| (Intercept) | 2.113 | 0.342 | 6.180 | 7.28 × 10−8 | *** | - |
| Seed linolenic acid | −0.033 | 0.021 | −1.597 | 0.1159 | - | 35.59 |
| Seeds per pod | 0.028 | 0.009 | 2.995 | 0.0041 | ** | 41.66 |
| Pod development: Yellow stage | −0.276 | 0.243 | −1.137 | 0.2602 | - | - |
| Pod development: Yellow-green stage | −0.048 | 0.216 | −0.222 | 0.8253 | - | 52.67 |
| Year 1 Model On Year 2 Data | ||||||
|---|---|---|---|---|---|---|
| Coefficients | Estimate | Standard Error | t-Value | Pr (>|t|) | Significance | Cumulative % Variance Explained |
| (Intercept) | 2.186 | 0.325 | 6.730 | 3.65 × 10−9 | *** | - |
| Linolenic acid | −0.008 | 0.023 | −0.357 | 0.7224 | - | - |
| Seeds per pod | 0.012 | 0.007 | 1.777 | 0.0798 | . | - |
| Pod development: Yellow stage | 0.192 | 0.103 | 1.863 | 0.0666 | . | - |
| Pod development: Yellow-green stage | 0.332 | 0.103 | 3.219 | 0.0019 | ** | - |
| Year 2 Model on Year 2 Data | ||||||
| (Intercept) | −2.844 | 0.494 | −5.757 | 2.42 × 10−7 | *** | - |
| Log[Vegmass] | 1.262 | 0.042 | 30.377 | <2 × 10−16 | *** | 73.70% |
| Total Protein | −0.019 | 0.010 | −1.952 | 0.0551 | . | 89.16% |
| Canopy leaf Chlorophyll b | 0.337 | 0.072 | 4.666 | 1.55 × 10−5 | *** | 91.41% |
| Total Lipid | 0.015 | 0.006 | 2.736 | 0.0204 | * | 92.04% |
| Seed weight per pod | 0.001 | 0.000 | 4.036 | 0.0001 | *** | 92.80% |
| No. pods on primary raceme | 0.007 | 0.001 | 4.427 | 3.67 × 10−5 | *** | 93.50% |
| Canopy leaf calcium | −0.125 | 0.029 | −4.336 | 5.07 × 10−5 | *** | 94.40% |
| Linolenic acid | −0.021 | 0.008 | −2.517 | 0.0143 | * | 94.81% |
| log(Pod Chlorophyll a content Wk43) | −0.024 | 0.012 | −2.046 | 0.0448 | * | 95.12% |
| Log[Vegmass] Model on Year 2 Data | ||||||
| (Intercept) | 2.423 | 0.108 | 22.435 | <2 × 10−16 | *** | - |
| Stem area | 0.001 | 0.000 | 3.407 | 0.0011 | ** | 47.16% |
| Log[No. branches on primary stem] | 0.029 | 0.003 | 9.574 | 2.06 × 10−14 | *** | 59.93% |
| Height | 0.005 | 0.001 | 7.373 | 2.42 × 10−10 | *** | 77.74% |
| Total branch No. | 0.034 | 0.004 | 9.793 | 2.04 × 10−14 | *** | 75.63% |
| Pod carotenoid content (Wk41) | 0.021 | 0.007 | 2.857 | 0.005 | ** | 80.04% |
| Coefficients | Estimate | Standard Error | t-Value | Pr (>|t|) | Significance | Cumulative % Variance Explained |
|---|---|---|---|---|---|---|
| (Intercept) | −0.377 | 0.359 | 1.050 | 0.2975 | - | - |
| Stem area | 0.002 | 0.001 | 3.150 | 0.00241 | ** | 36.64% |
| Log[No. branches on primary stem] | 0.724 | 0.151 | 4.811 | 8.51 × 10−6 | *** | 44.22% |
| Length of flower set | 0.009 | 0.003 | 3.294 | 0.0016 | ** | 51.73% |
| Canopy leaf potassium | −0.188 | 0.057 | −3.293 | 0.0016 | ** | 56.92% |
| Canopy leaf chlorophyll b | 0.550 | 0.185 | 2.972 | 0.0041 | ** | 60.19% |
| Canopy leaf Mg | 1.594 | 0.617 | 2.582 | 0.0120 | * | 63.69% |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennett, E.J.; Brignell, C.J.; Carion, P.W.C.; Cook, S.M.; Eastmond, P.J.; Teakle, G.R.; Hammond, J.P.; Love, C.; King, G.J.; Roberts, J.A.; et al. Development of a Statistical Crop Model to Explain the Relationship between Seed Yield and Phenotypic Diversity within the Brassica napus Genepool. Agronomy 2017, 7, 31. https://doi.org/10.3390/agronomy7020031
Bennett EJ, Brignell CJ, Carion PWC, Cook SM, Eastmond PJ, Teakle GR, Hammond JP, Love C, King GJ, Roberts JA, et al. Development of a Statistical Crop Model to Explain the Relationship between Seed Yield and Phenotypic Diversity within the Brassica napus Genepool. Agronomy. 2017; 7(2):31. https://doi.org/10.3390/agronomy7020031
Chicago/Turabian StyleBennett, Emma J., Christopher J. Brignell, Pierre W. C. Carion, Samantha M. Cook, Peter J. Eastmond, Graham R. Teakle, John P. Hammond, Clare Love, Graham J. King, Jeremy A. Roberts, and et al. 2017. "Development of a Statistical Crop Model to Explain the Relationship between Seed Yield and Phenotypic Diversity within the Brassica napus Genepool" Agronomy 7, no. 2: 31. https://doi.org/10.3390/agronomy7020031
APA StyleBennett, E. J., Brignell, C. J., Carion, P. W. C., Cook, S. M., Eastmond, P. J., Teakle, G. R., Hammond, J. P., Love, C., King, G. J., Roberts, J. A., & Wagstaff, C. (2017). Development of a Statistical Crop Model to Explain the Relationship between Seed Yield and Phenotypic Diversity within the Brassica napus Genepool. Agronomy, 7(2), 31. https://doi.org/10.3390/agronomy7020031