Agro-Morphological Evaluation of Rice (Oryza sativa L.) for Seasonal Adaptation in the Sahelian Environment


 ,
,
Abstract
:1. Introduction
2. Results
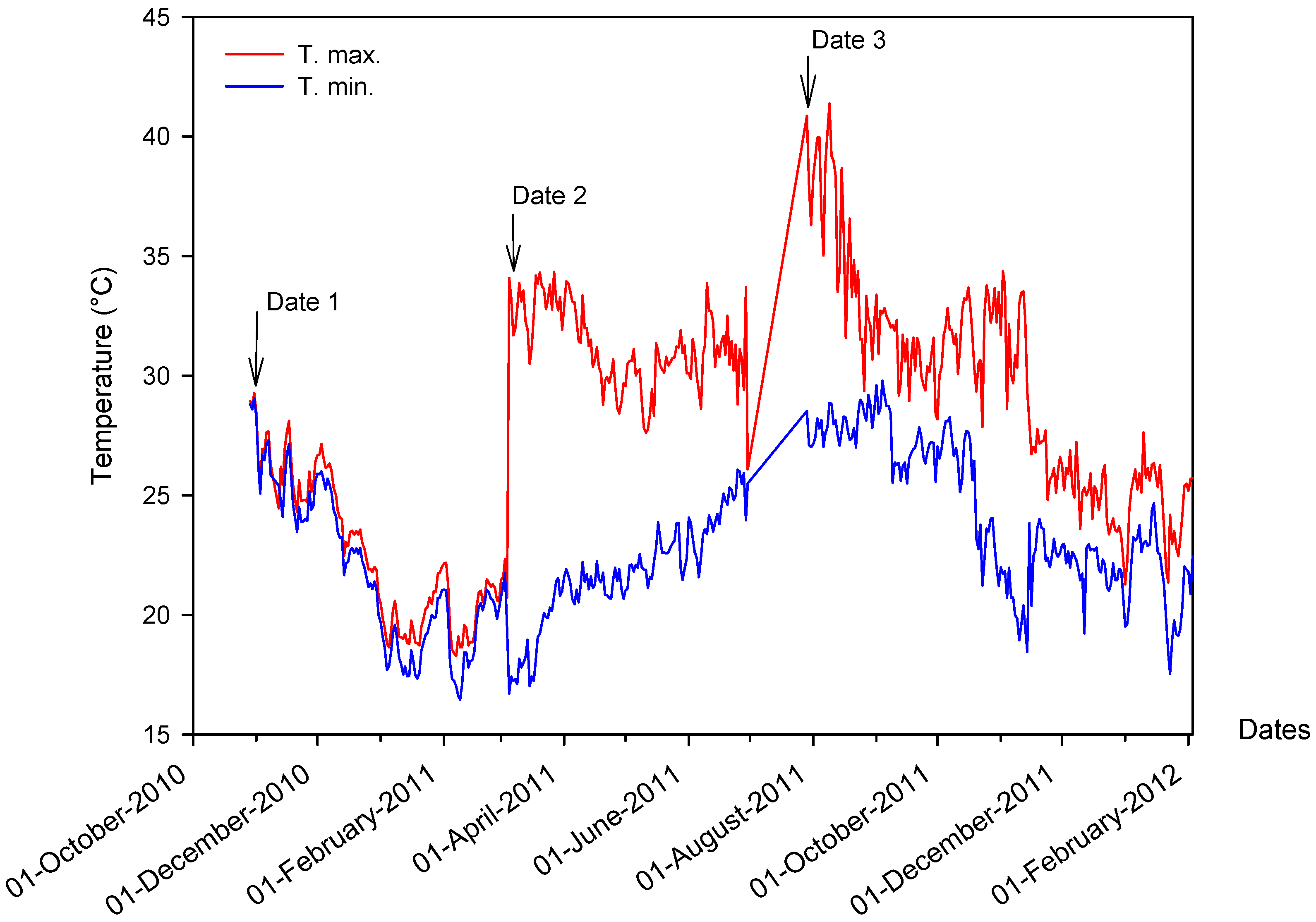
2.1. Air and Water Temperature in the Experiment Field
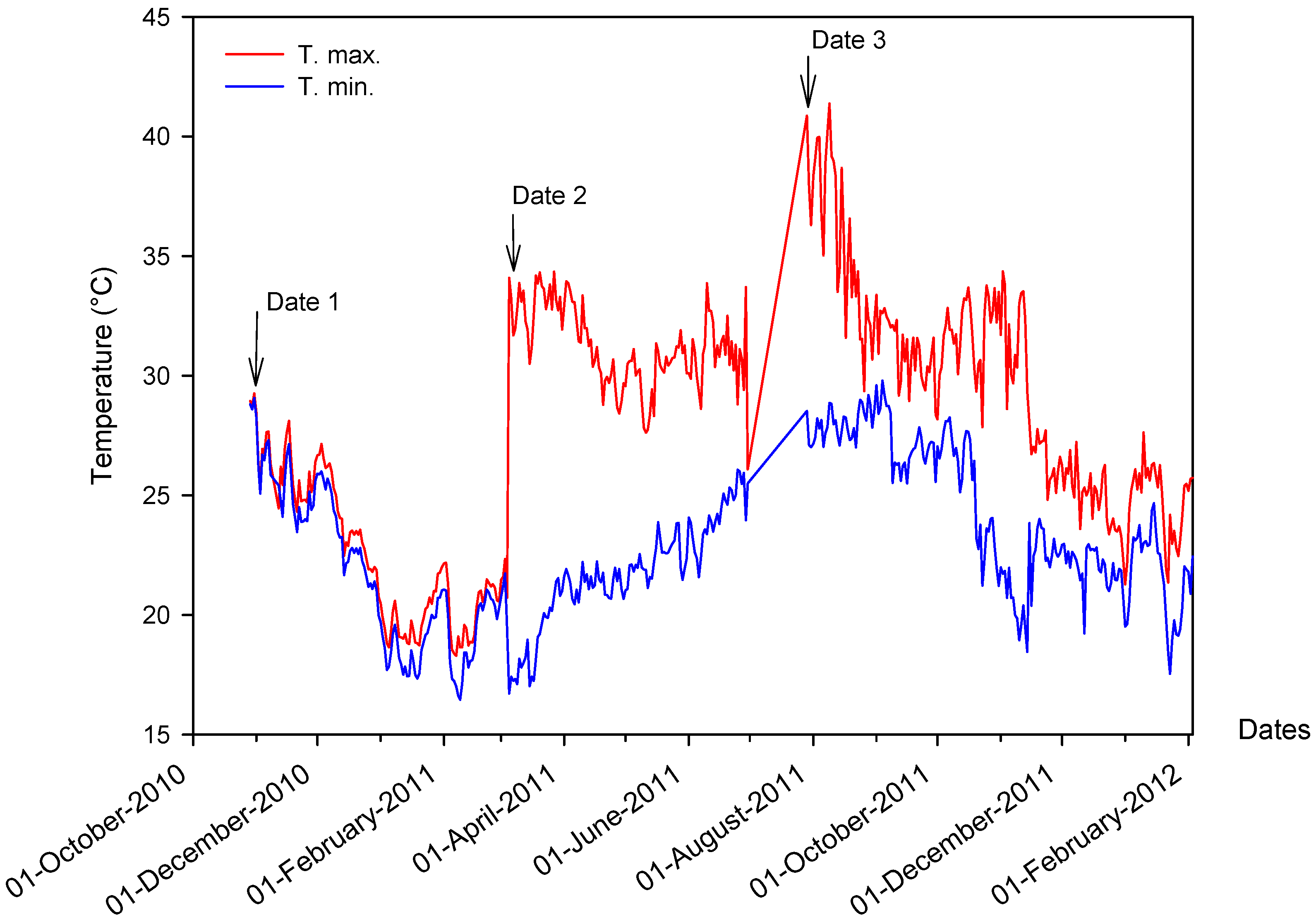
2.2. Effect of Sowing Date on Yield and Morpho-Physiological Traits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | df | Sum Square | Mean Square | F Value |
|---|---|---|---|---|
| Sterility (%) | ||||
| Date (D) | 2 | 210,311 | 105,155 | 349.52 *** |
| Genotype (G) | 223 | 118,178 | 530 | 5.31 *** |
| GXD | 418 | 189,714 | 454 | 4.55 *** |
| Panicle length | ||||
| Date (D) | 2 | 2,354.9 | 1,177.44 | 165.63 *** |
| Genotype (G) | 223 | 6,436.2 | 28.86 | 2.87 *** |
| GXD | 418 | 5,474.3 | 13.1 | 1.30 |
| Panicle exsertion | ||||
| Date (D) | 2 | 2,888.8 | 1,444.38 | 174.70 *** |
| Genotype (G) | 223 | 4,269.2 | 19.14 | 2.15 ** |
| GXD | 418 | 3,740.8 | 8.95 | 1.00 |
| Tiller number | ||||
| Date (D) | 2 | 15,294.3 | 7,647 | 115.21 *** |
| Genotype (G) | 223 | 21,584.2 | 96.8 | 4.24 *** |
| GXD | 418 | 14,093.4 | 33.7 | 1.48 |
| Days to mature | ||||
| Date (D) | 2 | 167,171 | 83,585 | 147.30 *** |
| Genotype (G) | 223 | 180,984 | 812 | 16.091 *** |
| GXD | 418 | 98,802 | 236 | 4.69 *** |
| Plant height (cm) | ||||
| Date (D) | 2 | 113,738 | 56,869 | 46.85 *** |
| Genotype (G) | 223 | 339,532 | 1,523 | 8.21 *** |
| GXD | 418 | 164,196 | 393 | 2.12 ** |
| Harvest index | ||||
| Date (D) | 2 | 4.3328 | 2.1664 | 83.65 *** |
| Genotype (G) | 223 | 5.8066 | 0.02604 | 5.67 *** |
| GXD | 418 | 5.1519 | 0.01233 | 2.69 *** |
| Yield | ||||
| Date (D) | 2 | 2,303 | 1,151.5 | 215.92 *** |
| Genotype (G) | 223 | 1,219.6 | 5.47 | 5.43 *** |
| GXD | 418 | 2,056.13 | 4.92 | 4.88 *** |
| Biomass | ||||
| Date (D) | 2 | 149,199 | 74,599 | 8.95 ** |
| Genotype (G) | 223 | 1,246,177 | 5,588 | 3.00 *** |
| GXD | 418 | 1,426,052 | 3,412 | 1.83 * |
2.3. Effect of Low Temperatures on the Morpho-Physiological Traits of Rice
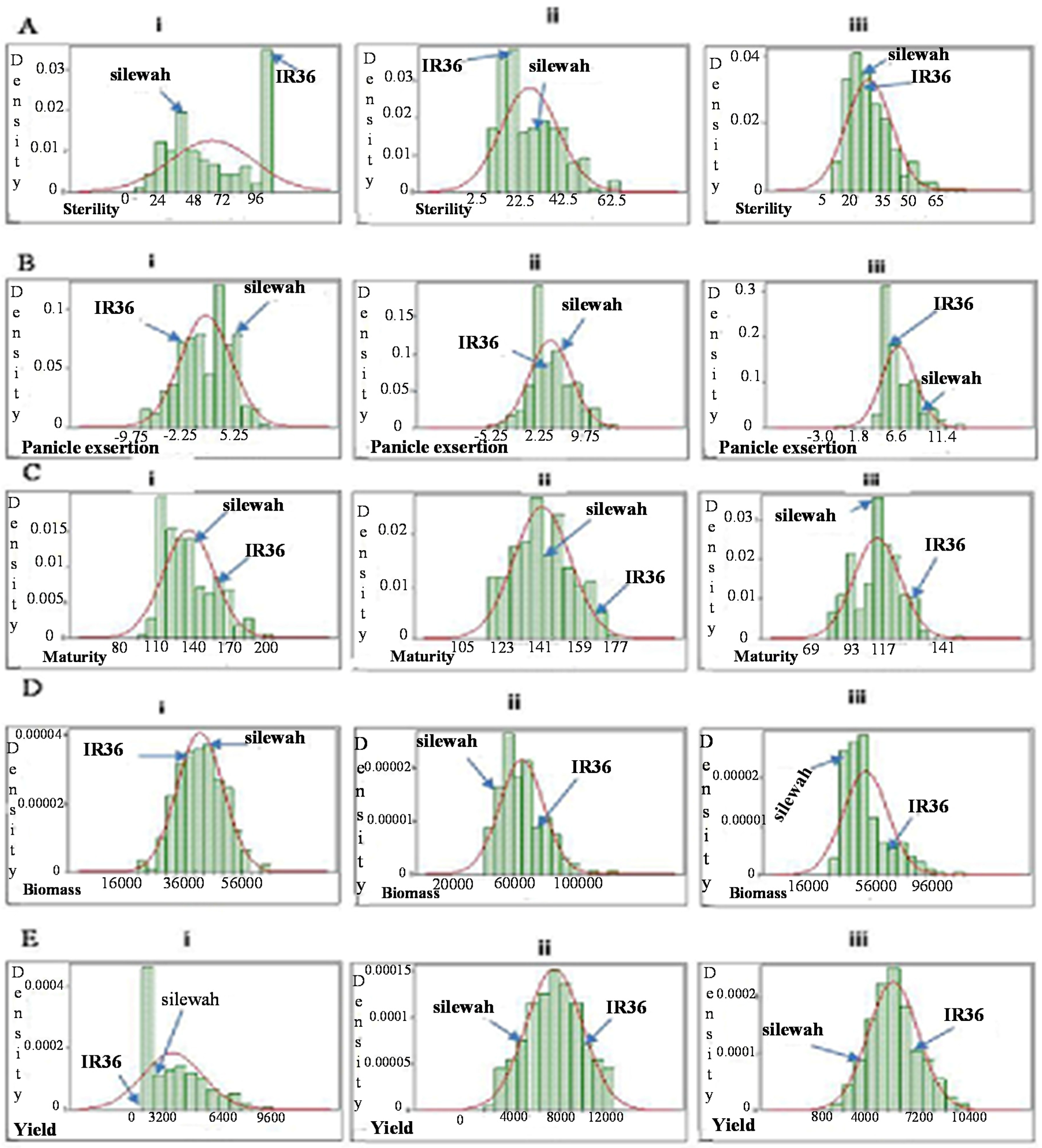
2.3.1. Spikelet Sterility
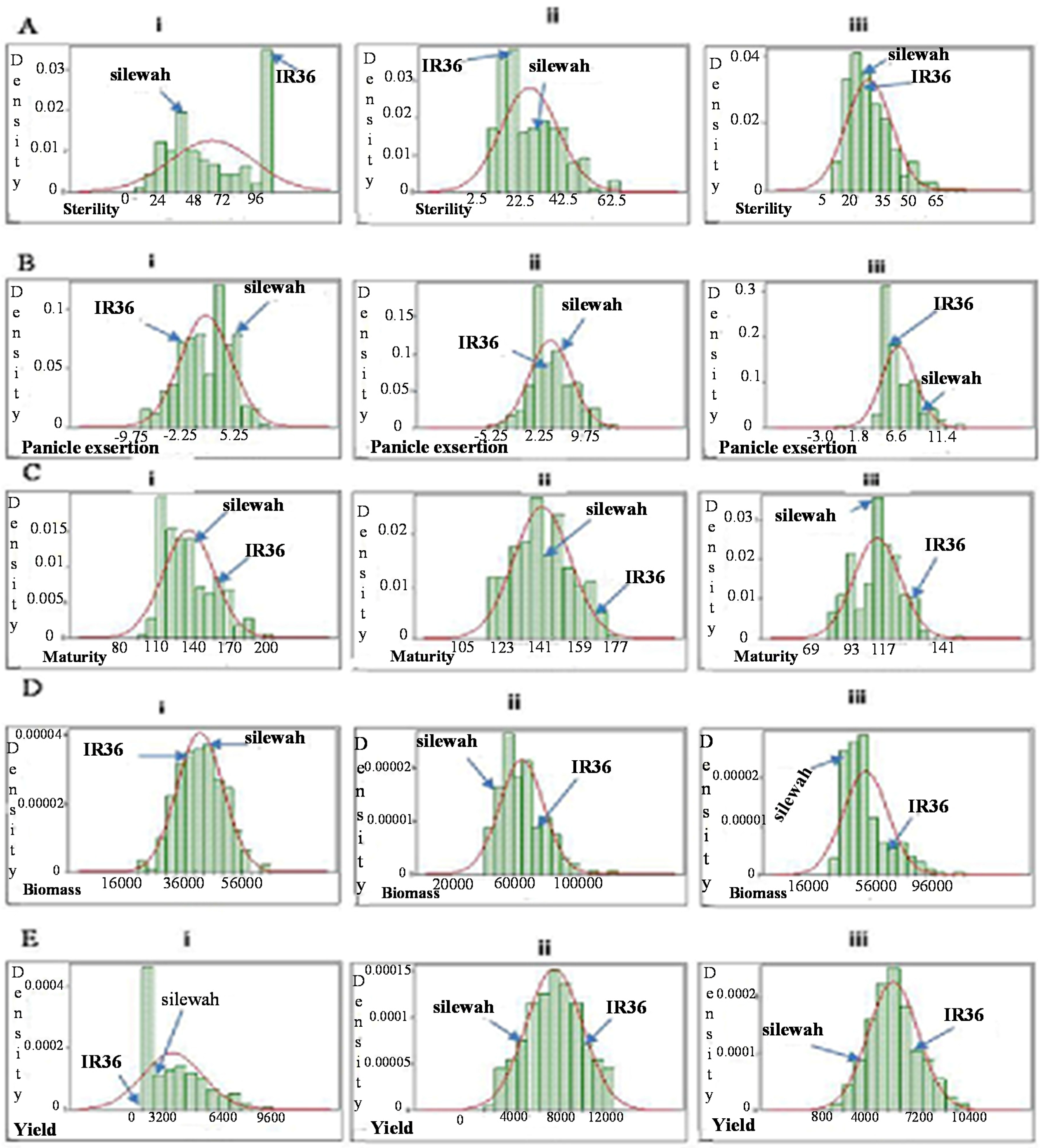
2.3.2. Panicle Exsertion
2.3.3. Crop Maturity
2.3.4. Biomass
2.3.5. Grain Yield
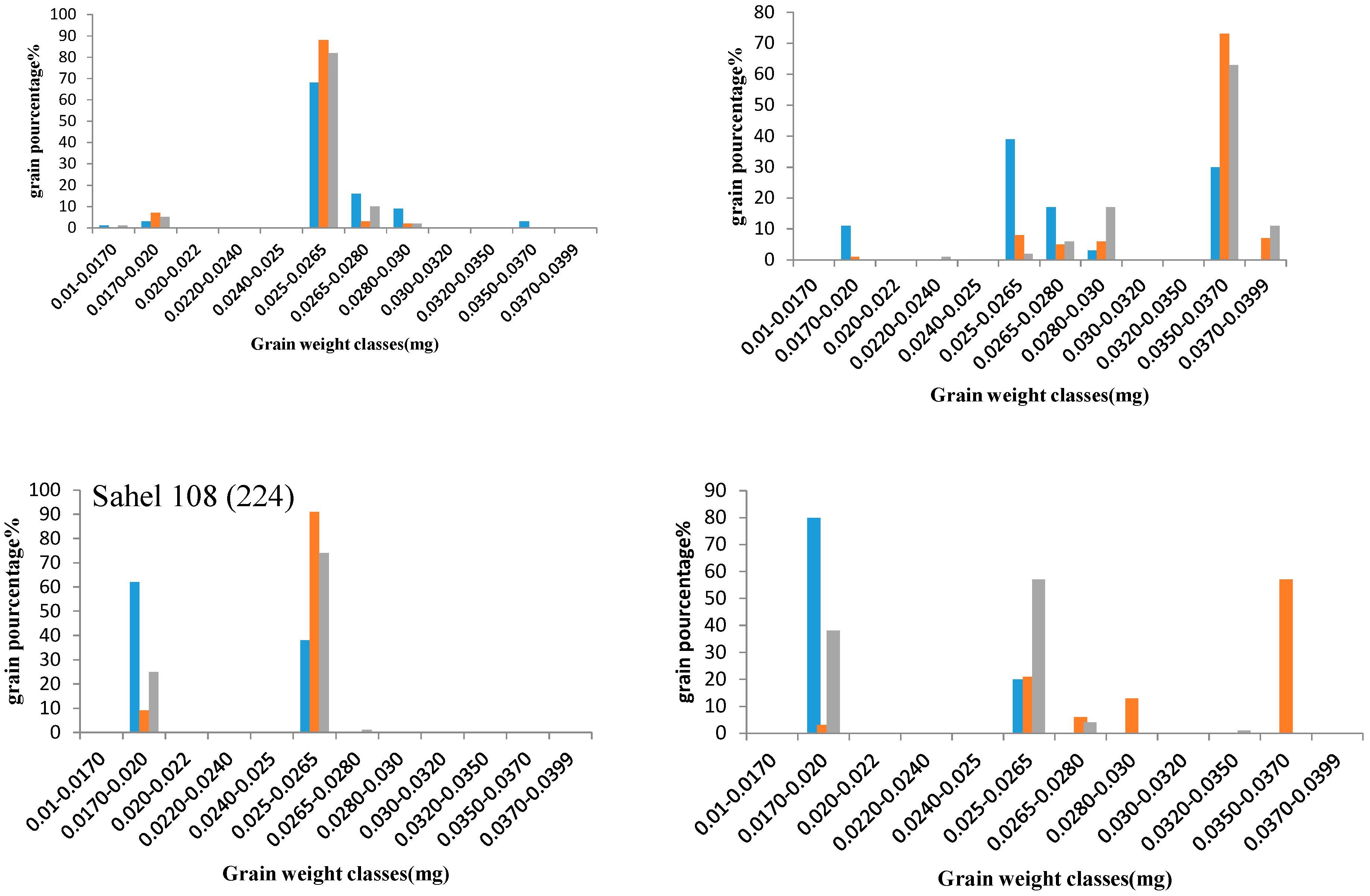
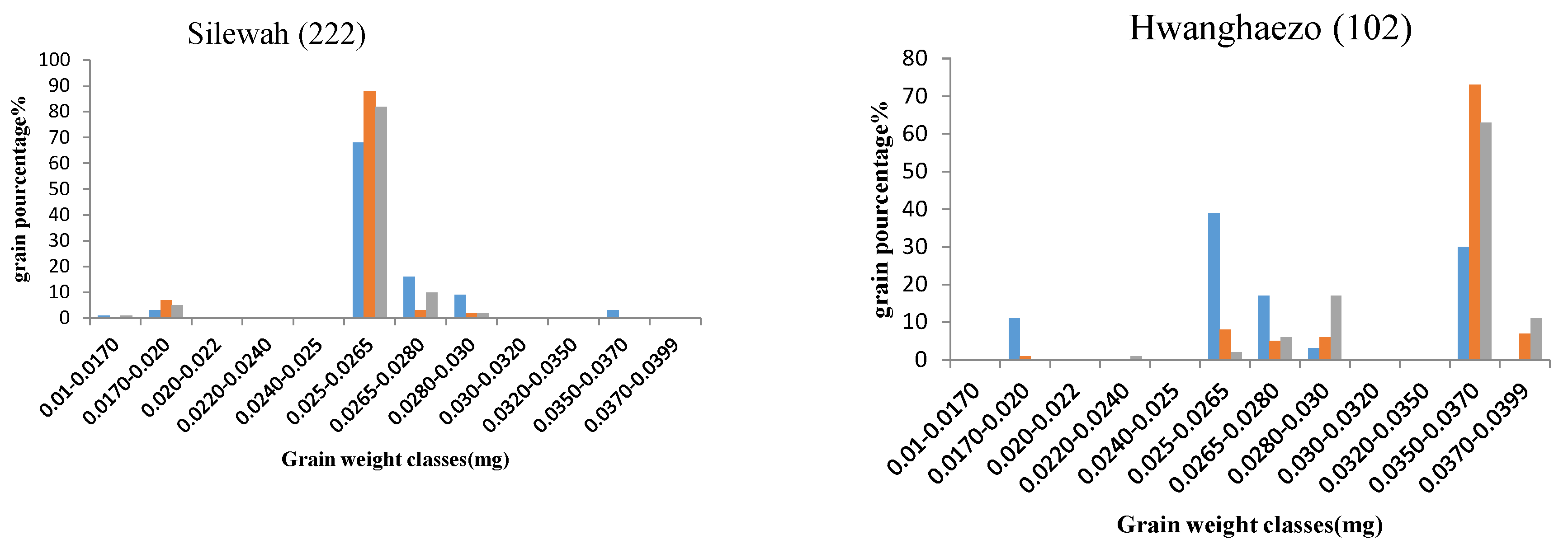
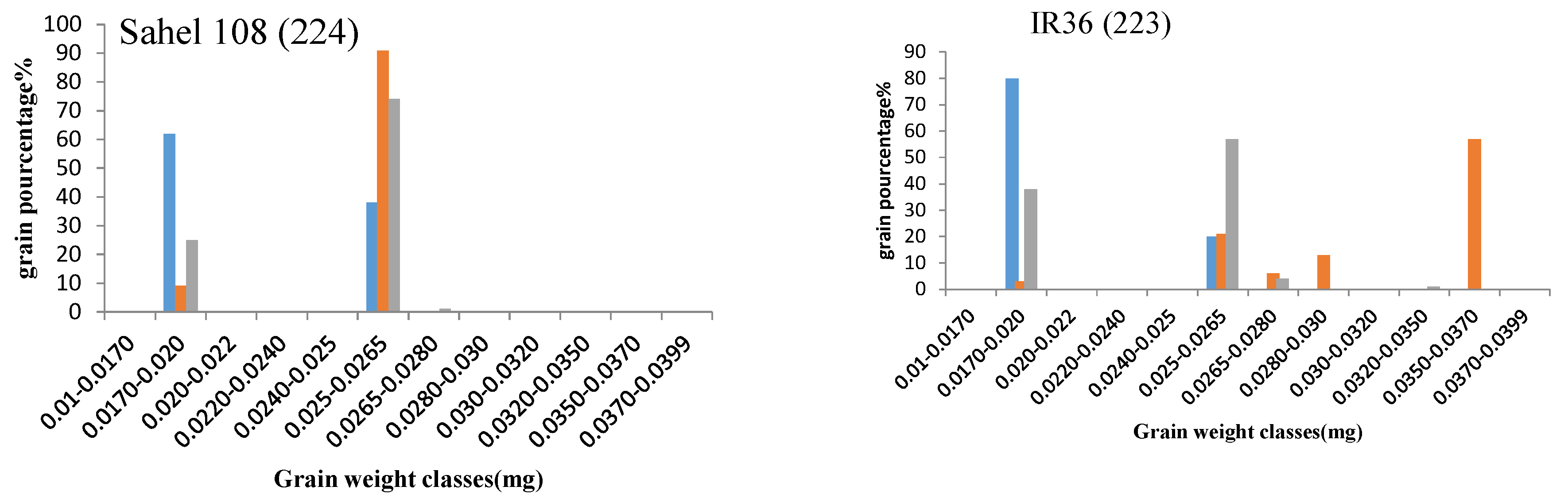
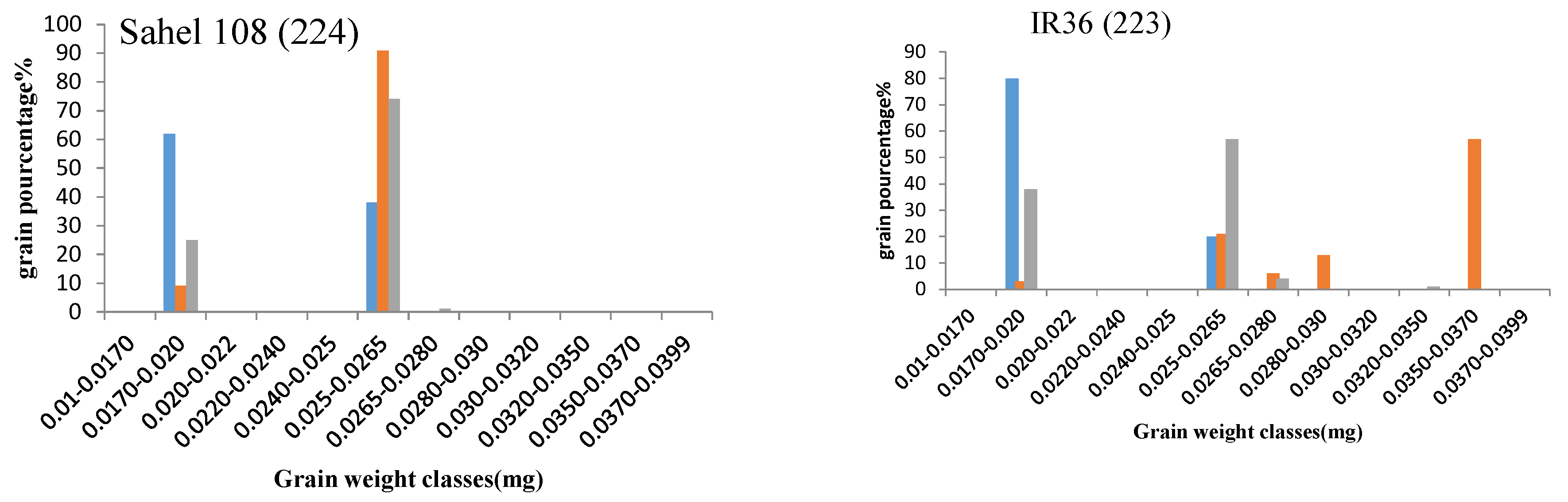
2.3.6. Grain Filling

2.3.7. Correlation between Agro-Morphological-Physiological Parameters at Different Sowing Dates
| Trait | Panicle Length (cm) | Panicle Exsertion (cm) | Tiller Number | Days to Mature | Plant Height (cm) | Yield (kg/ha) | Biomass (kg/ha) | Biological Harvest Index (%) |
|---|---|---|---|---|---|---|---|---|
| Sterility (%) | −0.136 * | −0.298 ** | 0.163 * | 0.452 ** | −0.297 ** | −0.815 ** | −0.087 | −0.675 ** |
| Panicle length (cm) | −0.152 * | −0.271 ** | −0.312 ** | 0.538 ** | 0.051 | 0.009 | 0.187 * | |
| Panicle exsertion (cm) | −0.218 * | −0.082 | 0.168 * | 0.270 ** | 0.118 | 0.209 * | ||
| Tiller number/m2 | 0.316 ** | −0.451 ** | −0.121 | 0.12 | −0.232 * | |||
| Days to mature (days) | −0.495 ** | −0.303 ** | 0.284 ** | −0.464 ** | ||||
| Plant height (cm) | 0.139 * | −0.055 | 0.210 * | |||||
| Yield (kg/ha) | 0.285 ** | 0.680 ** | ||||||
| Biomass (kg/ha) | 0.068 |
| Panicle Length (cm) | Panicle Exsertion (cm) | Tiller Number | Days to Mature (Days) | Plant Height (cm) | Yield (kg/ha) | Biomass (kg/ha) | Biological Harvest Index (%) | |
|---|---|---|---|---|---|---|---|---|
| Sterility (%) | 0.31 ** | 0.009 | 0.287 ** | 0.531 ** | 0.082 | −0.385 ** | 0.246 * | −0.438 ** |
| Panicle length (cm) | 0.261 * | 0.122 | 0.325 ** | 0.285 ** | −0.051 | 0.343 ** | −0.296 ** | |
| Panicle exsertion (cm) | 0.003 | 0.059 | 0.572 ** | −0.128 | 0.171 * | −0.368 ** | ||
| Tiller number/m2 | 0.415 ** | −0.039 | 0.229 * | 0.479 ** | −0.161 * | |||
| Days to mature (days) | −0.038 | −0.049 | 0.442 ** | −0.424 ** | ||||
| Plant height (cm) | −0.17 * | 0.193 * | −0.491 ** | |||||
| Yield (kg/ha) | 0.435 ** | 0.481 ** | ||||||
| Biomass (kg/ha) | −0.145 * |
| Panicle Length (cm) | Panicle Exsertion (cm) | Tiller Number | Days to Mature | Plant Height (cm) | Yield (kg/ha) | Biomass (kg/ha) | Biological Harvest Index (%) | |
|---|---|---|---|---|---|---|---|---|
| Sterility (%) | 0.206 | 0.008 | 0.178 | 0.379 | 0.126 | −0.388 | 0.246 | −0.463 |
| Panicle length (cm) | −0.05 | 0.247 | 0.482 | 0.229 | 0.066 | 0.532 | −0.527 | |
| Panicle exsertion (cm) | −0.111 | −0.142 | 0.084 | −0.003 | -0.003 | 0.055 | ||
| Tiller number/m2 | 0.47 | −0.082 | 0.198 | 0.343 | −0.192 | |||
| Days to mature (days) | −0.001 | 0.006 | 0.588 | −0.635 | ||||
| Plant height (cm) | −0.074 | 0.218 | −0.33 | |||||
| Yield (kg/ha) | 0.308 | 0.313 | ||||||
| Biomass (kg/ha) | −0.58 |
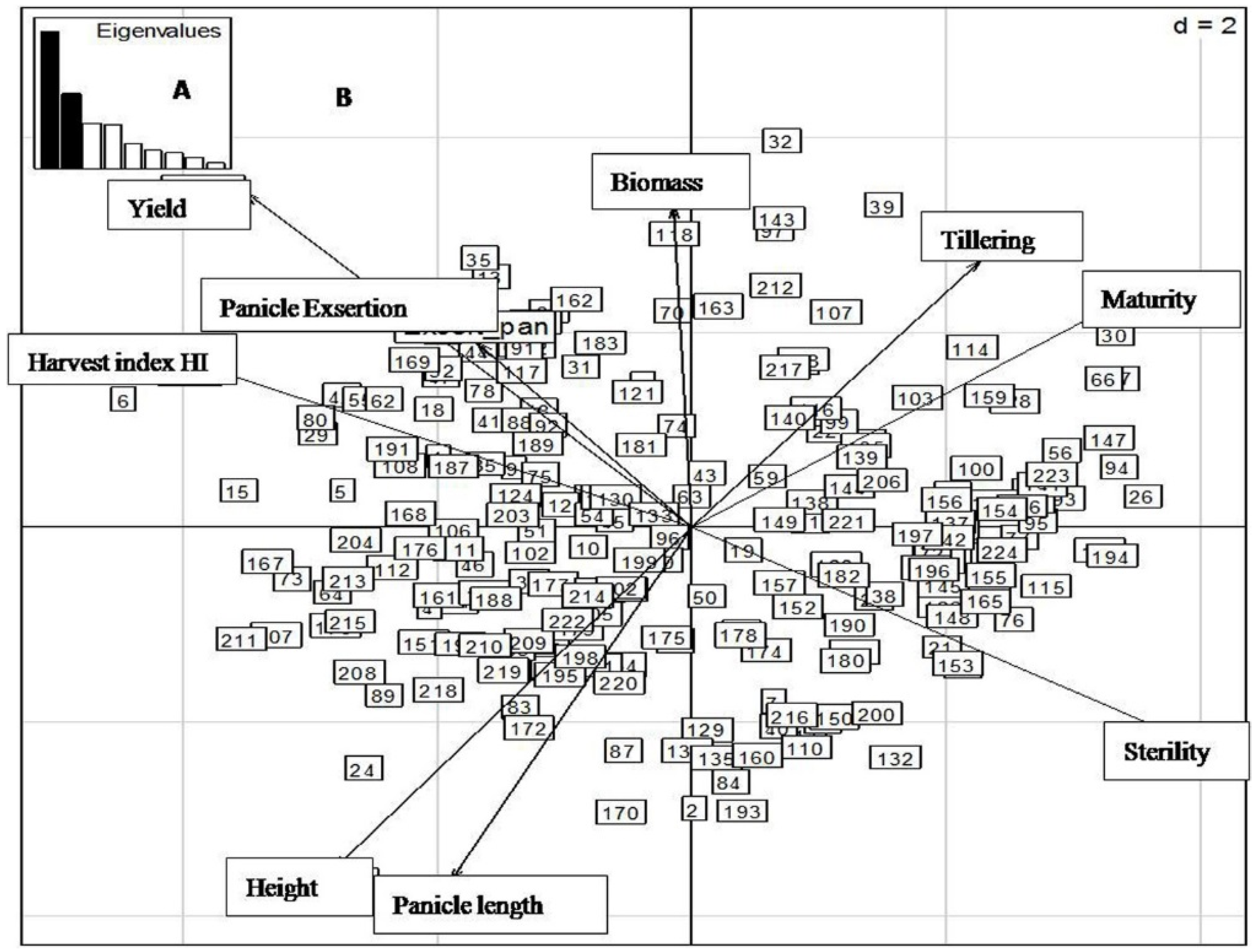
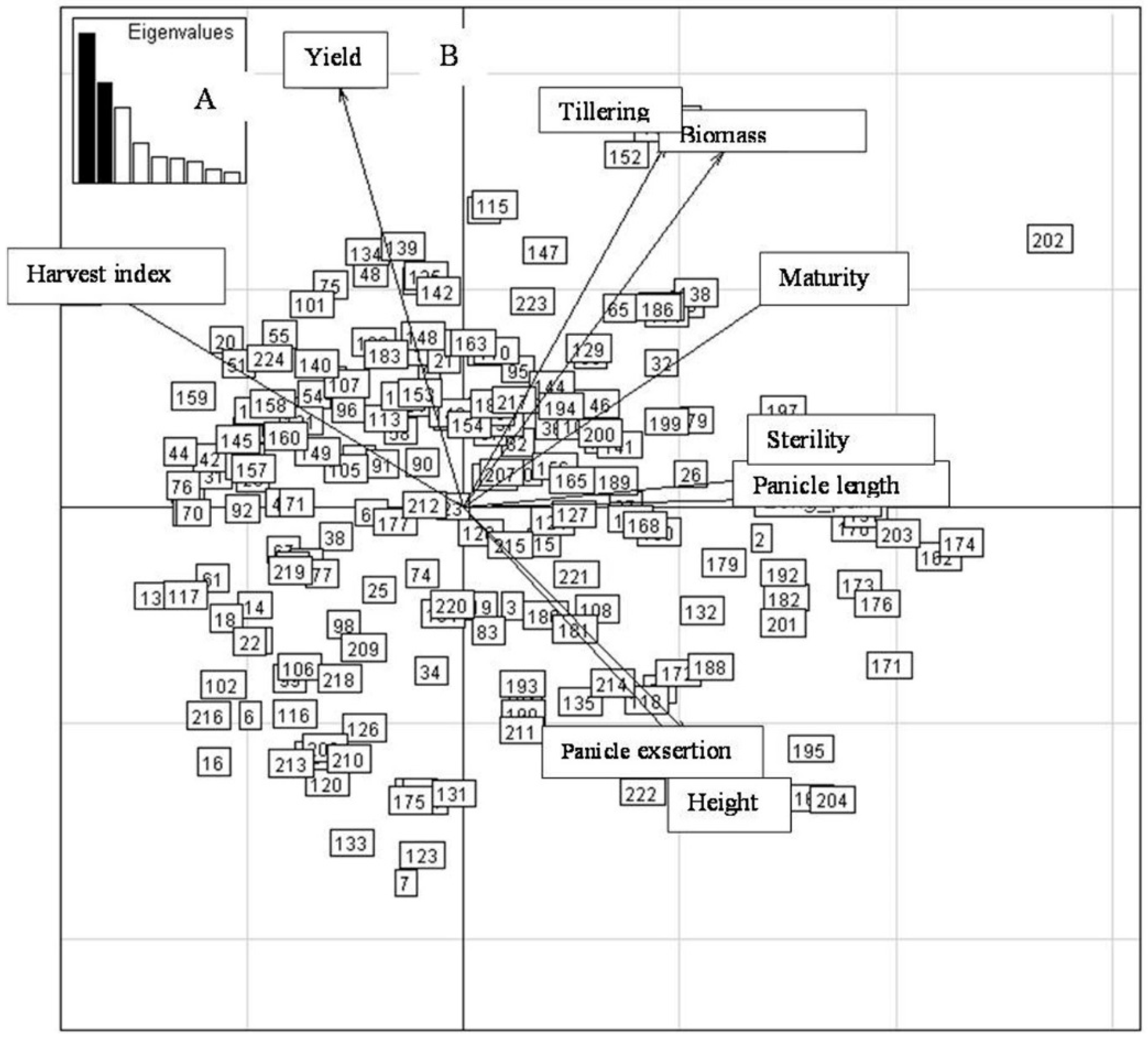
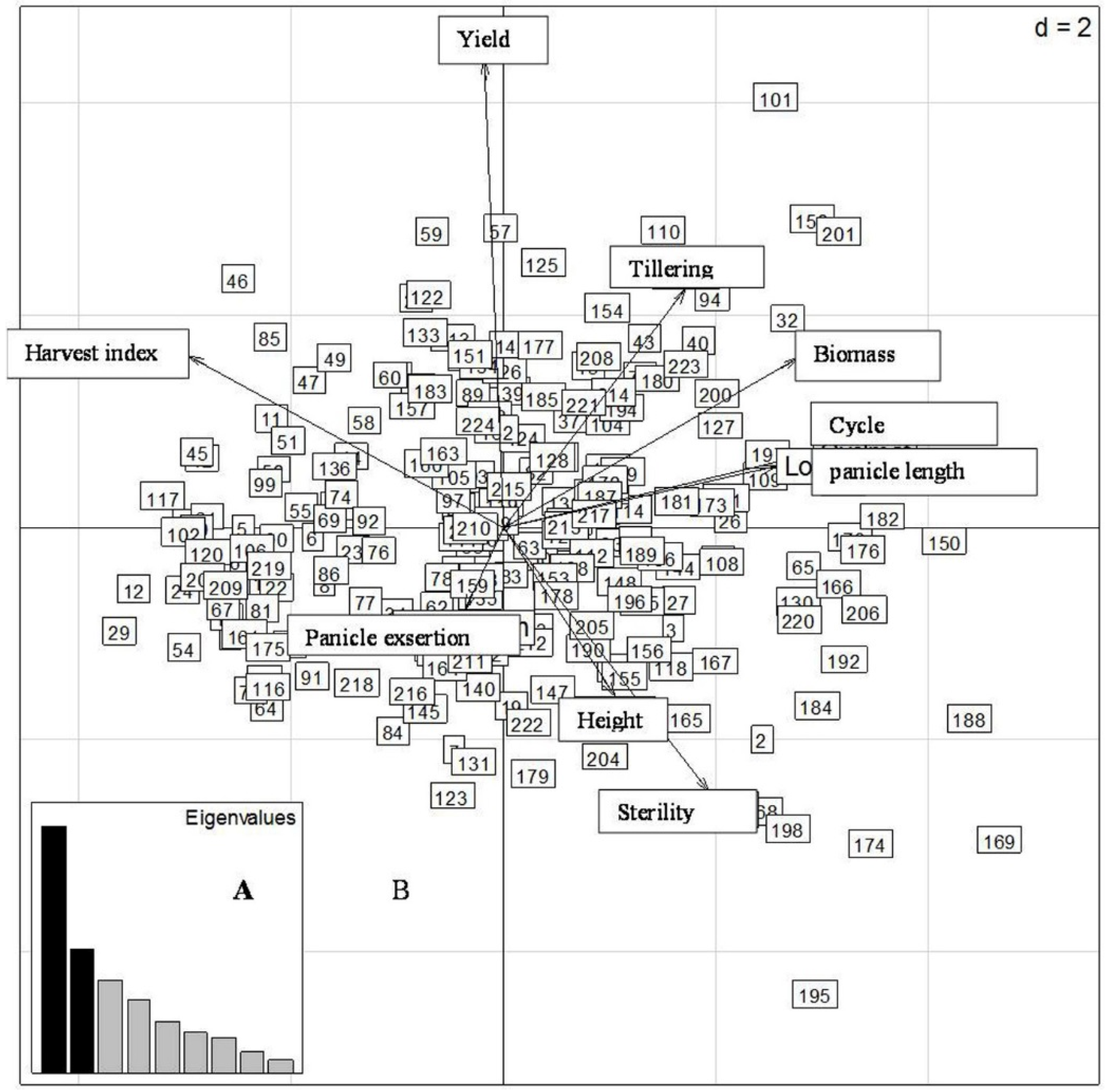
2.4. Principal Component Analysis (PCA) of Yield and Related Traits during the Different Sowing Dates
2.4.1. First Sowing Date
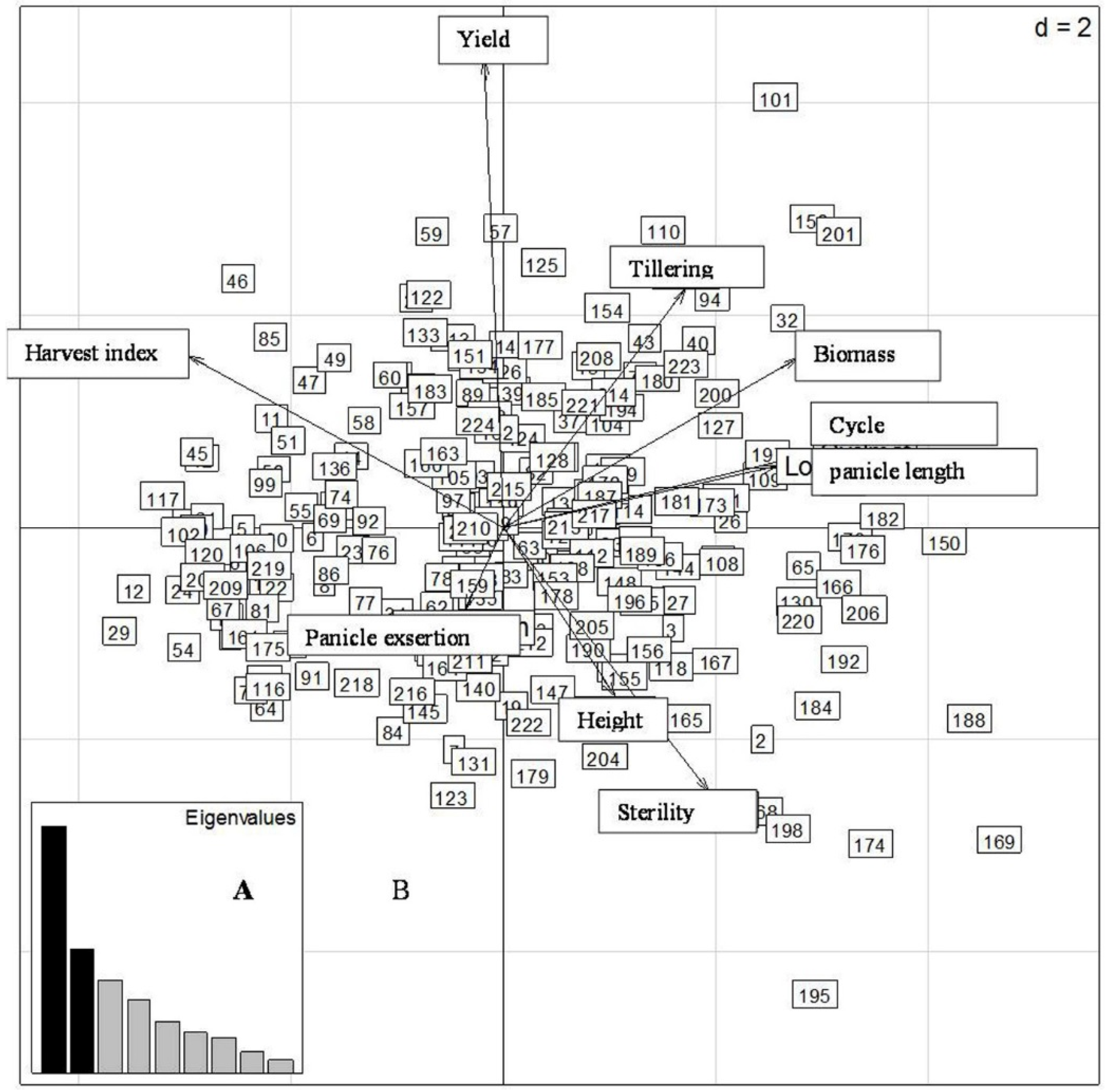
2.4.2. Second Sowing Date
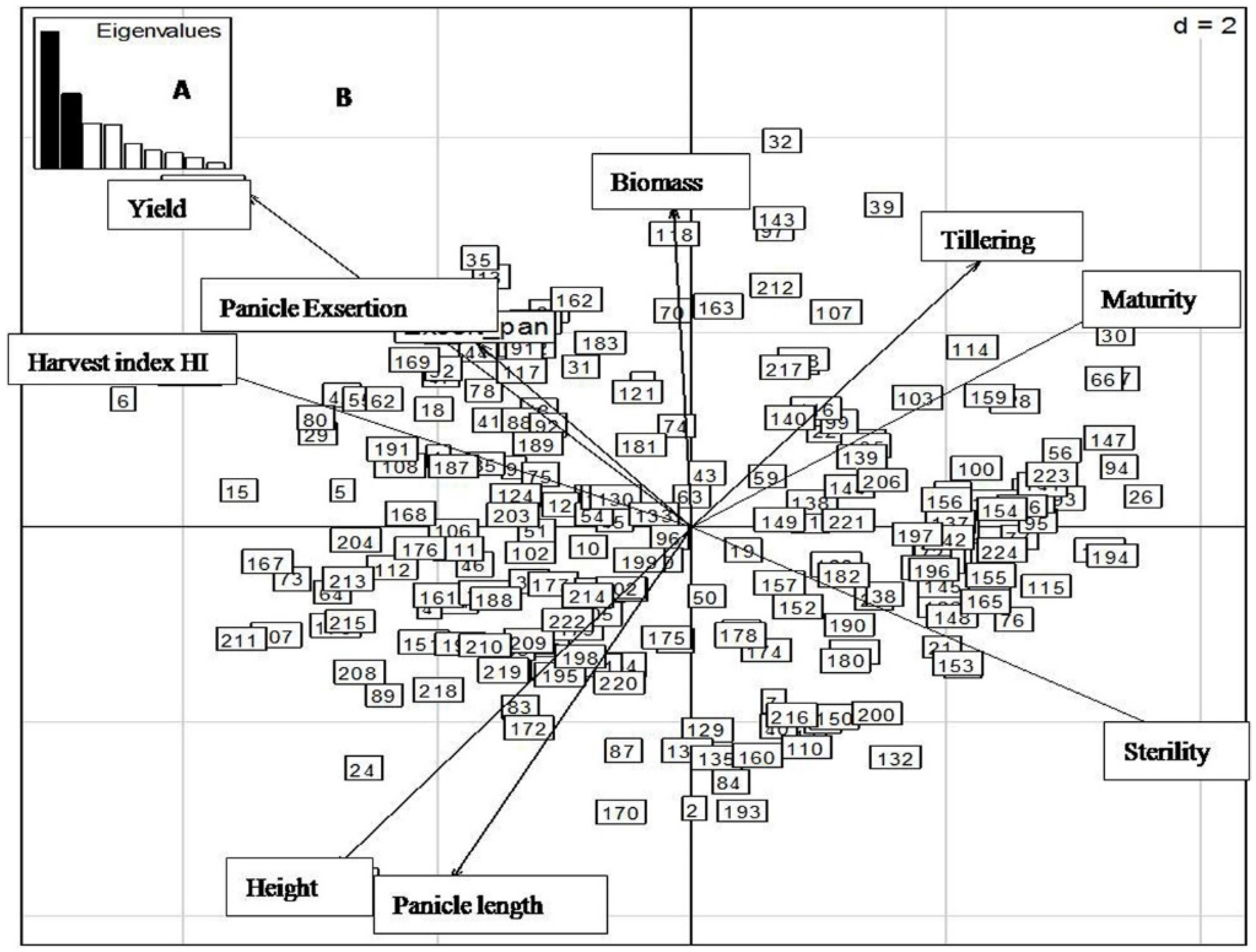
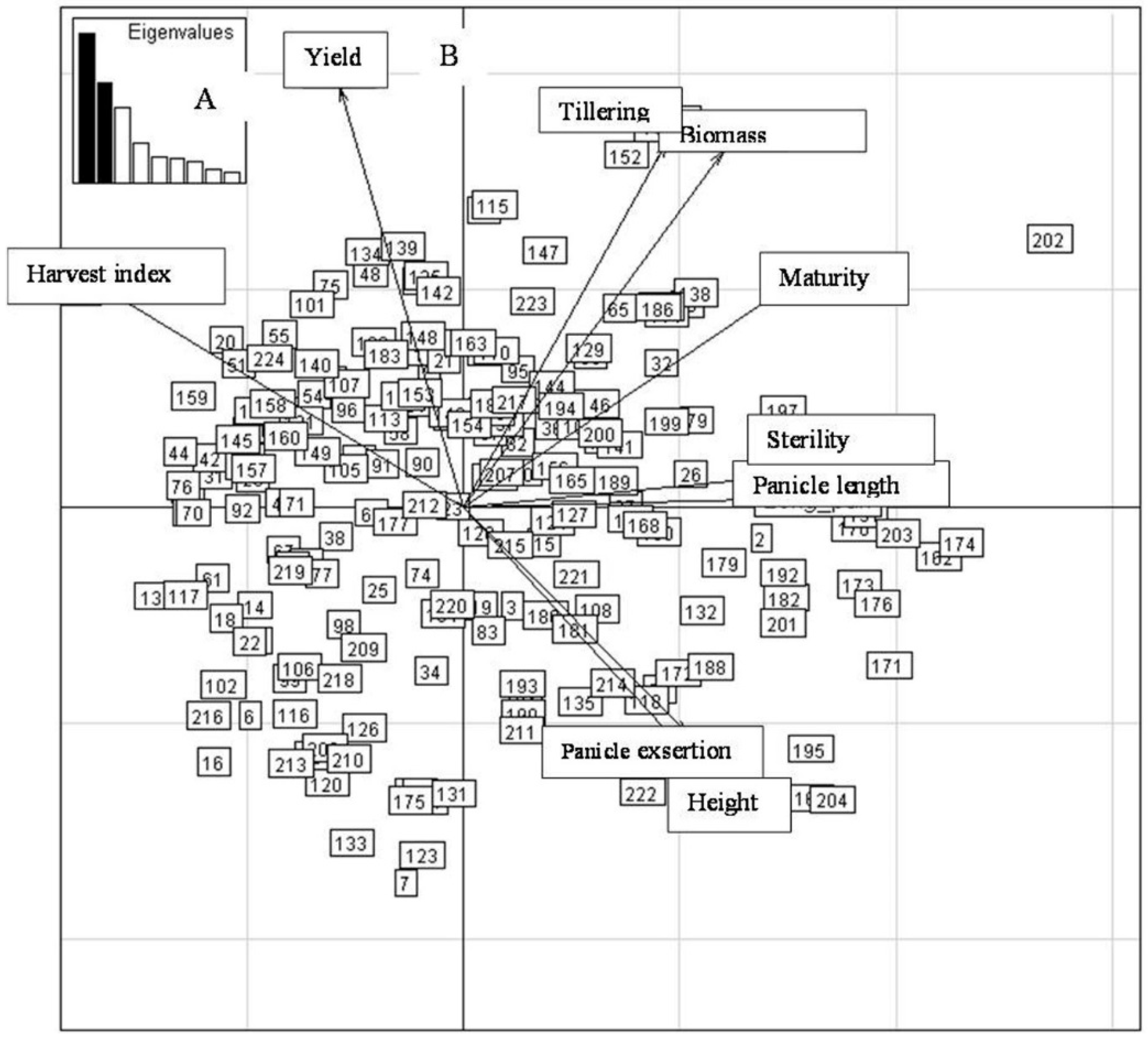
2.4.3. Third Sowing Date
3. Discussion
3.1. Genotype-by-Environment Interaction for Grain Yield and Agro-Morphological Traits
3.2. Impact of Cold Stress on Grain Yield and Agro-Morphological Traits
3.3. Association of Morpho-Physiological Traits with Grain Yield under Cold Tolerance Stress
4. Materials and Methods
4.1. Plant Material and Site Characteristics
4.2. Experimental Design
4.3. Cultural Practices
4.4. Parameters Measured
4.4.1. Air and Water Temperature
4.4.2. Crop Measurements
4.5. Statistical Analysis of Data
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix
| Number | Genotype | Origin | Subspecies | Number | Genotype | Origin | Subspecies |
|---|---|---|---|---|---|---|---|
| 1 | 82079-TR489-3-1-1 | TURKEY | Japonica | 29 | HR 17570-21-5-2-5-2-2-1-5 | PHILIPPINES | Japonica |
| 2 | 86011-TR 888-2-1-2-1 | TURKEY | Japonica | 30 | IR 776-74-3B-1-1-12-5 | IRRI | Indica |
| 3 | 87020-TR 968-1-1-1 | TURKEY | Japonica | 31 | HWASEONGBYEO | KOREA | Japonica |
| 4 | 87024-TR 972-6-3-1 | TURKEY | Japonica | 32 | IR 49830-7-1-2-3 | IRRI | Indica |
| 5 | 87041-TR 990-11-2-1 | TURKEY | Japonica | 33 | IR57107-2B-12-2-2-2 | IRRI | Indica |
| 6 | 88021-TR 1046-2-1-2-1 | TURKEY | Japonica | 34 | IR 59471-2B-20-2-1 | IRRI | Indica |
| 7 | CHANDANATH-3 | TURKEY | Japonica | 35 | IR 57257-34-1-2-1 | IRRI | Indica |
| 8 | 88024-TR 1049-6-1-2-1 | TURKEY | Japonica | 36 | IR 60059-4B-4B-4-1-1-2-1 | IRRI | Indica |
| 9 | 88076-TR 1101-9-2-1 | TURKEY | Japonica | 37 | IR 64 | IRRI | Indica |
| 10 | 88088-TR 1113-4-1-1 | TURKEY | Japonica | 38 | IR 64629-6-2-2-2 | IRRI | Indica |
| 11 | 88090-TR 1115-4-1-6 | TURKEY | Japonica | 39 | IR 65469-161-2-2-3-2-2 | IRRI | Indica |
| 12 | 89010-TR1130-8-1-1-2 | TURKEY | Japonica | 40 | IR 66097-8-1-1-1 | IRRI | Indica |
| 13 | 89014-TR 1134-2-2-3 | TURKEY | Japonica | 41 | IR 68333-R-R-B-22 | IRRI | Indica |
| 14 | 69016-TR 1136-1-1-1 | TURKEY | Japonica | 42 | IR 68349-131-2-2-3 | IRRI | Indica |
| 15 | 89018-TR 1138-4-2-1 | TURKEY | Japonica | 43 | IR68399-78-2-3-3-1 | IRRI | Indica |
| 16 | 90040-TR 1232-4-1-1 | TURKEY | Japonica | 44 | IR 71121-35-1-1-1-2 | IRRI | Indica |
| 17 | 90051-TR 1243-2-2-1 | TURKEY | Japonica | 45 | IR 71131-BF 4-B-30-5 | IRRI | Indica |
| 18 | 83025-TR 643-1-1-1-1 | TURKEY | Japonica | 46 | IR 73944-1-2-2 | IRRI | Indica |
| 19 | BR1543-9-2-1 | BANGLADESH | Indica | 47 | IR 73305-14-2-2 | IRRI | Indica |
| 20 | BR 28 | BANGLADESH | indica | 48 | IR73688-82-2-3-2-2 | IRRI | Indica |
| 21 | CHINA 1039 | INDIA | Indica | 49 | IR 73688-82-3 | IRRI | Indica |
| 22 | CT6744-F2-CA-8 | CHILE | Japonica | 50 | IR 84645-305-61-1-B | IRRI | Indica |
| 23 | GAUTAM | INDIA | Japonica | 51 | IR 73689-31-1 | IRRI | Indica |
| 24 | GWANSAN 2 | DPR KOREA | Japonica | 52 | IR 73690-7-2-1-1-3-2-2-1 | IRRI | Indica |
| 25 | H231-59-3-1 | ARGENTINA | Japonica | 53 | IR 74506-28-4-3-2-1-3-2-2 | IRRI | Indica |
| 26 | HEXI 24 | CHINA | Japonica | 54 | IR 74520-29-4-2-2-2-4-1-1 | IRRI | Indica |
| 27 | HEXI 25 | CHINA | Japonica | 55 | IR 76687-22-1-3-2-5 | IRRI | Indica |
| 28 | HR17512-11-2-3-1-4-2-3 | PILIPPINES | Japonica | 56 | IR 80098-38-3-1-2 | IRRI | Indica |
| 57 | PSB RC28 | IRRI | Indica | 92 | SR22746-68-2-3-4-2-4 | KOREA | Japonica |
| 58 | PSB RC 44 | IRRI | Indica | 93 | STEJAREE 45 | RUSSIA | Japonica |
| 59 | PSB RC 64 | IRRI | Indica | 94 | SU 98 | DPR KOREA | Indica |
| 62 | JARRAH | AUSTRALIA | Japonica | 95 | SUREX 95 | TURKEY | Japonica |
| 63 | K-39-96-1-1-1-2 | INDIA | Indica | 96 | WON 124 | DPR KOREA | Japonica |
| 64 | CHANDANATH-1 | NEPAL | Japonica | 97 | YR1076-8-4-1-2-3-1-2 | KOREA | Japonica |
| 65 | MAHSURI | MALAYSIA | Indica | 98 | ZHI 20-5 | CHINA | Japonica |
| 66 | MANAW THUKHA | MALAYSIA | Indica | 99 | CHOJANG | KOREA | Japonica |
| 67 | MILYANG15 | KOREA | Japonica | 100 | HAMNAM 14 | DPR KOREA | Japonica |
| 68 | MILYANG 55 | KOREA | Japonica | 101 | HAMNAM15 | DPR KOREA | Japonica |
| 69 | NONG 56 | DPR KOREA | Japonica | 102 | HWANGHAEZO | DPR KOREA | Japonica |
| 70 | NR 11 | VIETNAM | indica | 103 | NONG 57 | DPR KOREA | Indica |
| 71 | OLBYEO1 | KOREA | Japonica | 104 | RYONGSONG12 | KOREA | Indica |
| 72 | OM 987-1 | VIETNAM | Indica | 105 | YUNLEN 4 | CHINA | Japonica |
| 73 | PANDA | ITALY | Indica | 106 | X-JIGNA | ETHIOPIA | Japonica |
| 74 | PSB RC 4 | IRRI | Indica | 107 | WD1 DALAN KAOUYE | NIGER | Indica |
| 75 | PSB RC 92 | IRRI | Indica | 108 | TY 53 | NIGER | Indica |
| 76 | PSB RC 94 | IRRI | Indica | 109 | DS 10 | NIGER | Indica |
| 77 | PSB RC 96 | IRRI | Indica | 110 | TY 47 | NIGER | Indica |
| 78 | RCPL-3-2 | INDIA | Indica | 111 | TY 32 | NIGER | Indica |
| 79 | RCPL-3-6 | INDIA | Indica | 112 | DS 4 | NIGER | Indica |
| 80 | ROJOFOTSY 653 | MADAGASCAR | Japonica | 113 | IR 84649-97-2-B-B | IRRI | Indica |
| 81 | SANT ANDEREA | ITALY | Japonica | 114 | IR 8 | IRRI | Indica |
| 82 | SIM 2 SUMADEL | PHILIPPINES | Indica | 115 | PAVLOVSKY | RUSSIA | Japonica |
| 83 | SKAU 105 | INDIA | Japonica | 116 | PLOVDIV 22 | RUSSIA | Japonica |
| 84 | PALUNG-2 | Japonica | 117 | CALORO | USA | Japonica | |
| 85 | SKAU 27 | INDIA | Japonica | 118 | SAHEL 177 | AFRICARICE | Indica |
| 86 | SKAU337 | INDIA | Japonica | 119 | IR84649-50-1-B | IRRI | Indica |
| 87 | SKAU 339 | INDIA | Japonica | 120 | KOSHIHIKARI | JAPAN | Japonica |
| 88 | SKAU 382 | INDIA | Japonica | 121 | L 202 | USA | Japonica |
| 89 | SKUAT 27 | INDIA | Japonica | 122 | IR 50 | IRRI | Indica |
| 90 | SR 13349-59-1 | KOREA | Japonica | 123 | JUMALI MARSHI | NEPAL | Japonica |
| 91 | SR18518-BF-4-B-12-1-2 | KOREA | Japonica | 124 | PADI LABOU ALUMBIS | MALAYSIA | Japonica |
| 125 | KANTO | MADAGASCAR | Japonica | 155 | WAS55-B-B-2-1-2-5 | AFRICARICE | Indica |
| 126 | DOONGARA | AUSTRALIA | Japonica | 157 | WAS200-B-B-1-1-1 | AFRICARICE | Indica |
| 127 | C21 | PHILIPPINES | Indica | 158 | WAS122-IDSA-11-WAS-6-3 | AFRICARICE | Indica |
| 128 | WAS187-7-WASB-1-WAS1 | AFRICARICE | Indica | 159 | WAS194-B-1 | AFRICARICE | Indica |
| 129 | LENG KWANG | CHINA | Japonica | 160 | WAS114-B-IDSA-B-WAS-1-5-FKR-1 | AFRICARICE | Indica |
| 130 | KUNMINGXIAOBEIGU | CHINA | Japonica | 161 | 87025-TR 973-3-1-1 | TURKEY | Japonica |
| 131 | DOURADO AGUILHA | BRAZIL | Japonica | 162 | FANDRAPOTSY 104 | MADAGASCAR | Japonica |
| 132 | THANGONE | LAOS | Japonica | 163 | 93-11 | BHUTAN | Indica |
| 133 | JUMALI | NEPAL | Japonica | 164 | AL CHIAO HONG | CHINA | Japonica |
| 134 | PADI SASAHAL | MALAYSIA | Indica | 165 | BETSILAIZIAN | MADAGASCAR | Japonica |
| 135 | MITAK | INDONESIA | Japonica | 166 | BODOMANO | MADAGASCAR | Japonica |
| 136 | SASANISHIKI | JAPAN | Japonica | 167 | BOTOHAVANA | MADAGASCAR | Japonica |
| 137 | SAHEL 201 | AFRICARICE | Indica | 168 | BOTRA MAITSO | MADAGASCAR | Japonica |
| 138 | SAHEL 202 | AFRICARICE | Indica | 169 | BOTRYKELY | MADAGASCAR | Japonica |
| 139 | SAHEL 134 | AFRICARICE | Indica | 170 | BOTRY 731 | MADAGASCAR | Japonica |
| 140 | SAHEL 159 | AFRICARICE | Indica | 171 | GAJPATI | NEPAL | Indica |
| 141 | SAHEL 208 | AFRICARICE | Indica | 172 | GOPAL | NEPAL | Indica |
| 142 | SAHEL 209 | AFRICARICE | Indica | 173 | JENJAR | NEPAL | Indica |
| 143 | SAHEL 210 | AFRICARICE | Indica | 174 | KATI | BHUTAN | Indica |
| 144 | IR 84649-231-1-1-B | IRRI | Indica | 175 | MACHA PUCHRE-3 | NEPAL | Japonica |
| 145 | RASSI | INDIA | Indica | 176 | KITRANA 508 | MADAGASCAR | Japonica |
| 146 | ITA 212 | AFRICARICE | Indica | 177 | LAL AMAN | MADAGASCAR | Japonica |
| 147 | JAYA | INDIA | Indica | 178 | LATSIBOZAKA-112-1 | MADAGASCAR | Japonica |
| 148 | NERICA-L19 | AFRICARICE | Indica | 179 | LOHAMBITRO 224 | MADAGASCAR | Japonica |
| 149 | NERICA-L40 | AFRICARICE | Indica | 180 | MACAN BINUNDOK | MADAGASCAR | Japonica |
| 150 | GAMBIAKA KOKUM | MALI | Indica | 181 | MADINIKA 1329 | MADAGASCAR | Japonica |
| 151 | WAS127-12-1-2-1 | AFRICARICE | Indica | 182 | MAKALIOKA 34 | MADAGASCAR | Japonica |
| 152 | WAS21-B-B-20-4-3-3 | AFRICARICE | Indica | 183 | MALADY | MADAGASCAR | Japonica |
| 153 | WAS62-B-B-14-1 | AFRICARICE | Indica | 184 | MAMORIAKA | MADAGASCAR | Japonica |
| 154 | WAS62-B-B-17-1-1-3 | AFRICARICE | Indica | 185 | MENAHODITRA 1234 | MADAGASCAR | Japonica |
| 186 | NGAJA | BHUTAN | Indica | 206 | VARY VATO 154 | MADAGASCAR | Japonica |
| 187 | PA TOU HUNG | CHINA | Indica | 207 | SUPARICA 1 | ETHIOPIA | Japonica |
| 188 | PURBIA | NEPAL | Indica | 208 | AD01 | ETHIOPIA | Japonica |
| 189 | RAY JAZAYKAYZ | BHUTAN | Indica | 209 | DEMWOZE | ETHIOPIA | Japonica |
| 190 | RAY NABJA | BHUTAN | Indica | 210 | NERICA 6 | AFRICARICE | Japonica |
| 191 | ROJOFOTSY 693 | MADAGASCAR | Japonica | 211 | AD 012 | ETHIOPIA | Japonica |
| 192 | ROJOMENA 1034 | MADAGASCAR | Japonica | 212 | MTU-1001 | ETHIOPIA | Indica |
| 193 | SHORT GRAIN | THAILAND | Japonica | 213 | AURAT-7 | ETHIOPIA | Indica |
| 194 | SONA | IRAN | 214 | AD 048 | ETHIOPIA | Japonica | |
| 195 | TELOVOLANA 177 | MADAGASCAR | Japonica | 215 | GUMARA (IAC 164) | ETHIOPIA | Japonica |
| 196 | THAPACHINIYA | NEPAL | Indica | 216 | FOFIFA 3730 | MADAGASCAR | Japonica |
| 197 | TOKAMBANY 663 | MADAGASCAR | Japonica | 217 | BG 90-2 | ETHIOPIA | Indica |
| 198 | TOKAMBANY 669 | MADAGASCAR | Japonica | 218 | WAB 189-B-B-B-HB | ETHIOPIA | Japonica |
| 199 | TSAKA | MADAGASCAR | Japonica | 219 | KOKIT (IRAT-209) | ETHIOPIA | Japonica |
| 200 | TSIPALA 1231 | MADAGASCAR | Japonica | 220 | FOFIFA 3737 | ETHIOPIA | Japonica |
| 201 | TSIPALA B160 | MADAGASCAR | Japonica | 221 | DIAMANTE | CHILE | Japonica |
| 202 | TSIPALAFOTSY 1883 | MADAGASCAR | Japonica | 222 | SILEWAH | INDONESIA | Japonica |
| 203 | TSIPALA MENA 626 | MADAGASCAR | Japonica | 223 | IR 36 | IRRI | Indica |
| 204 | VARY LAVA DE MAROVATO | MADAGASCAR | Japonica | 224 | SAHEL 108 | AFRICARICE | Indica |
| 205 | VARY MADINIKA 3494 | MADAGASCAR | Japonica |
References
- Food and Agriculture Organization (FAO) FAOSTAT. Annual Report 2010–2011; FAO: Rome, Italy, 2011; Available online: http://www.fao.org (accessed on 11 March 2013).
- Mackill, D.J.; Lei, X. Genetic variation for traits related to temperate adaptation of rice cultivars. Crop Sci. 1997, 37, 1340–1346. [Google Scholar] [CrossRef]
- Andaya, V.C.; Mackill, D.J. Mapping of QTLs associated with cold tolerance during the vegetative stage in rice. J. Exp. Bot. 2003, 54, 2579–2585. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Andaya, V.C.; Tai, T.H. Cold sensitivity in rice (Oryza sativa L.) is strongly correlated with a naturally occurring I99V mutation in the multifunctional glutathione transferase isoenzyme GSTZ2. Biochem. J. 2011, 435, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Shimono, H.; Hasegawa, T.; Iwam, K. Response of growth and grain yield in paddy rice to cool water at different growth stages. Field Crops Res. 2002, 73, 67–79. [Google Scholar] [CrossRef]
- Huang, X.; Kurata, N.; Wei, X.; Wang, Z.X.; Wang, A.; Zhao, Q.; Zhao, Y.; Liu, K.; Lu, H.; Li, W.; et al. A map of rice genome variation reveals the origin of cultivated rice. Nature 2012, 490, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, P. Evaluation on sixty-eight rice germplasms in cold tolerance at germination stage. Rice Sci. 2010, 17, 77–81. [Google Scholar] [CrossRef]
- Kotaka, S.; Abe, N. The varietal difference of germinability at low-temperature in rice varieties and the testing method for the percentage establishment of seedlings. J. Agric. Sci. Tokyo 1988, 43, 165–168. [Google Scholar]
- Bosetti, F.; Montebell, C.; Novembre, A.D.; Chamma, H.P.; Pinheiro, J.B. Genetic variation of germination cold tolerance in Japanese rice germplasm. Breed. Sci. 2012, 62, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Ñanculao, G.D.; Cárcamo, M.P.; de los Santos, O.A.; Velásquez, V.B. Cold tolerance evaluation in Chilean rice genotypes at the germination stage. Chil. J. Agric. Res. 2013, 73, 3–8. [Google Scholar] [CrossRef]
- Zeng, Y.W.; Shen, S.Q.; Xu, F.R. Ecological diversity of cold-tolerant rice in Yunnan, China. Plant Genet. Res. Newsl. 1999, 117, 43–47. [Google Scholar]
- Li, S.C.; Zeng, Y.W.; Shen, S.Q.; Pu, X.Y. Cold Tolerance of core collection at booting stage associated with eco-geographic distribution in Yunnan rice landrace (Oryza sativa), China. Rice Sci. 2004, 11, 261–268. [Google Scholar]
- Dingkuhn, M.; Miézan, K.M. Stérilité des épillets de riz induite par la température dans le Sahel. Rapp. Annu. ADRAO 1992, 68, 40–41. [Google Scholar]
- Jiang, W.; Lee, J.; Chu, S.H.; Ham, T.H.; Woo, M.O. Genotype environment interactions for chilling tolerance of rice recombinant inbred lines under different low temperature environments. Field Crops Res. 2010, 117, 226–236. [Google Scholar] [CrossRef]
- Suh, J.P.; Cho, Y.C.; Lee, J.H.; Lee, S.B.; Jung, J.Y.; Choi, I.S.; Kim, M.K.; Kim, C.K.; Jena, K.K. SSR analysis of genetic diversity and cold tolerance in temperate rice germplasm. Plant Breed. Biotech. 2013, 1, 103–110. Available online: http://dx.doi.org/10.9787/PBB.2013.1.2.103 (accessed on 23 June 2013). [Google Scholar]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoter. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Mori, M.; Onishi, K.; Tokizono, Y.; Shinada, H.; Yoshimura, T.; Numao, Y.; Miura, H.; Sato, T. Detection of a novel quantitative trait locus for cold tolerance at the booting stage derived from a tropical japonica rice variety Silewah. Breed. Sci. 2011, 61, 61–68. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, C.; Beachell, H.M. Response of indica-japonica rice hybrids to low temperatures. SABRAO J. 1974, 6, 17–32. [Google Scholar]
- Zenna, N.; Luzi-Kihupi, A.; Manneh, B.; Raymond, R.; Gasore, E.R.; Traoré, K. Weathering the cold: Africa develops rice that thrive in the region’s cooler zones. Rice Today 2010, 27, 26–27. [Google Scholar]
- R Development Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011; Available online: http://www.R-project.org (accessed on 22 December 2011).
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ndour, D.; Diouf, D.; Bimpong, I.K.; Sow, A.; Kanfany, G.; Manneh, B. Agro-Morphological Evaluation of Rice (Oryza sativa L.) for Seasonal Adaptation in the Sahelian Environment. Agronomy 2016, 6, 8. https://doi.org/10.3390/agronomy6010008
Ndour D, Diouf D, Bimpong IK, Sow A, Kanfany G, Manneh B. Agro-Morphological Evaluation of Rice (Oryza sativa L.) for Seasonal Adaptation in the Sahelian Environment. Agronomy. 2016; 6(1):8. https://doi.org/10.3390/agronomy6010008
Chicago/Turabian StyleNdour, Daba, Diaga Diouf, Isaac Kofi Bimpong, Abdoulaye Sow, Ghislain Kanfany, and Baboucarr Manneh. 2016. "Agro-Morphological Evaluation of Rice (Oryza sativa L.) for Seasonal Adaptation in the Sahelian Environment" Agronomy 6, no. 1: 8. https://doi.org/10.3390/agronomy6010008
APA StyleNdour, D., Diouf, D., Bimpong, I. K., Sow, A., Kanfany, G., & Manneh, B. (2016). Agro-Morphological Evaluation of Rice (Oryza sativa L.) for Seasonal Adaptation in the Sahelian Environment. Agronomy, 6(1), 8. https://doi.org/10.3390/agronomy6010008