

High-Resolution 3D Reconstruction of Individual Rice Tillers for Genetic Studies


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
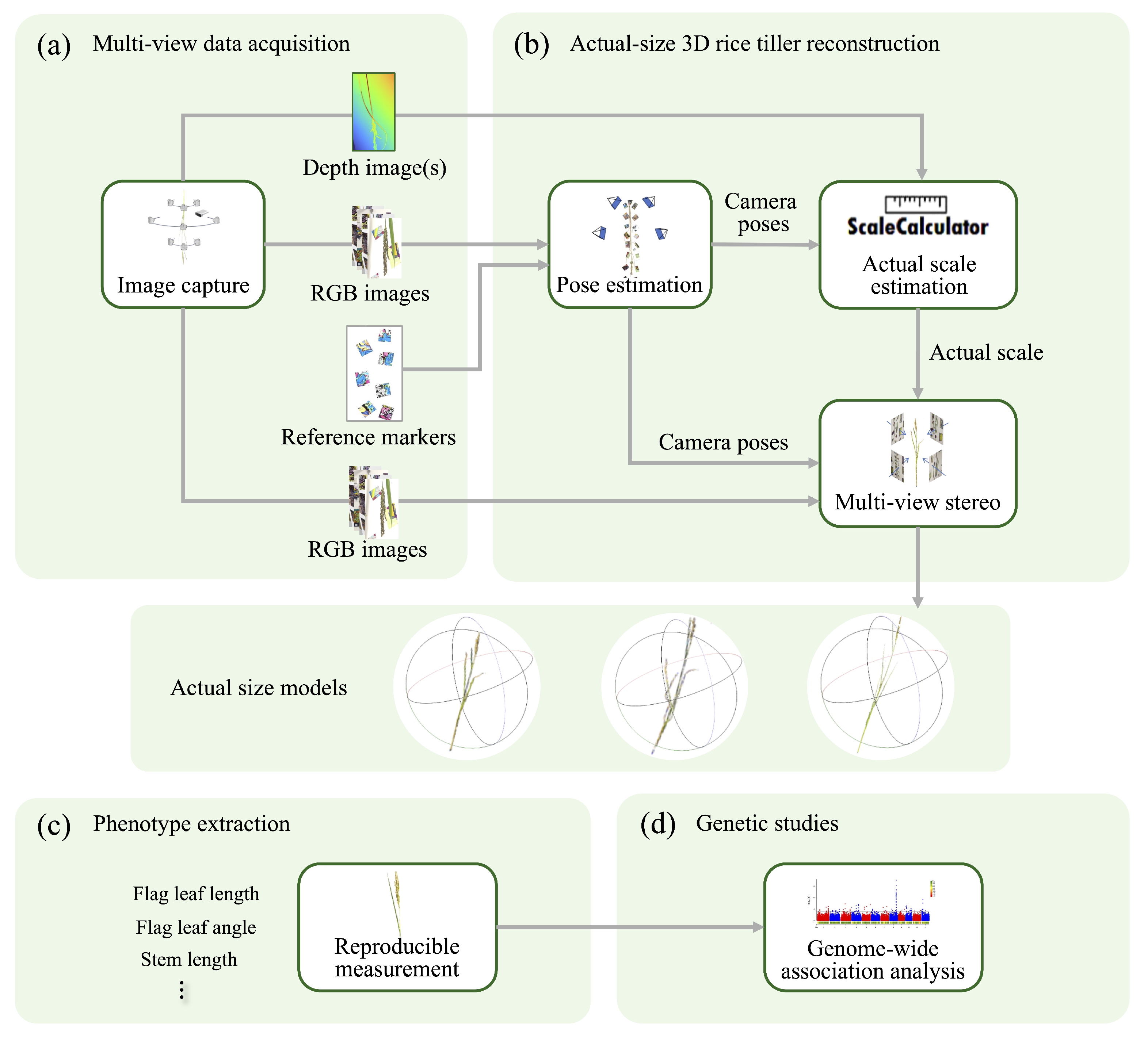
2.1. Overview of 3D Tiller Reconstruction for Genetic Study
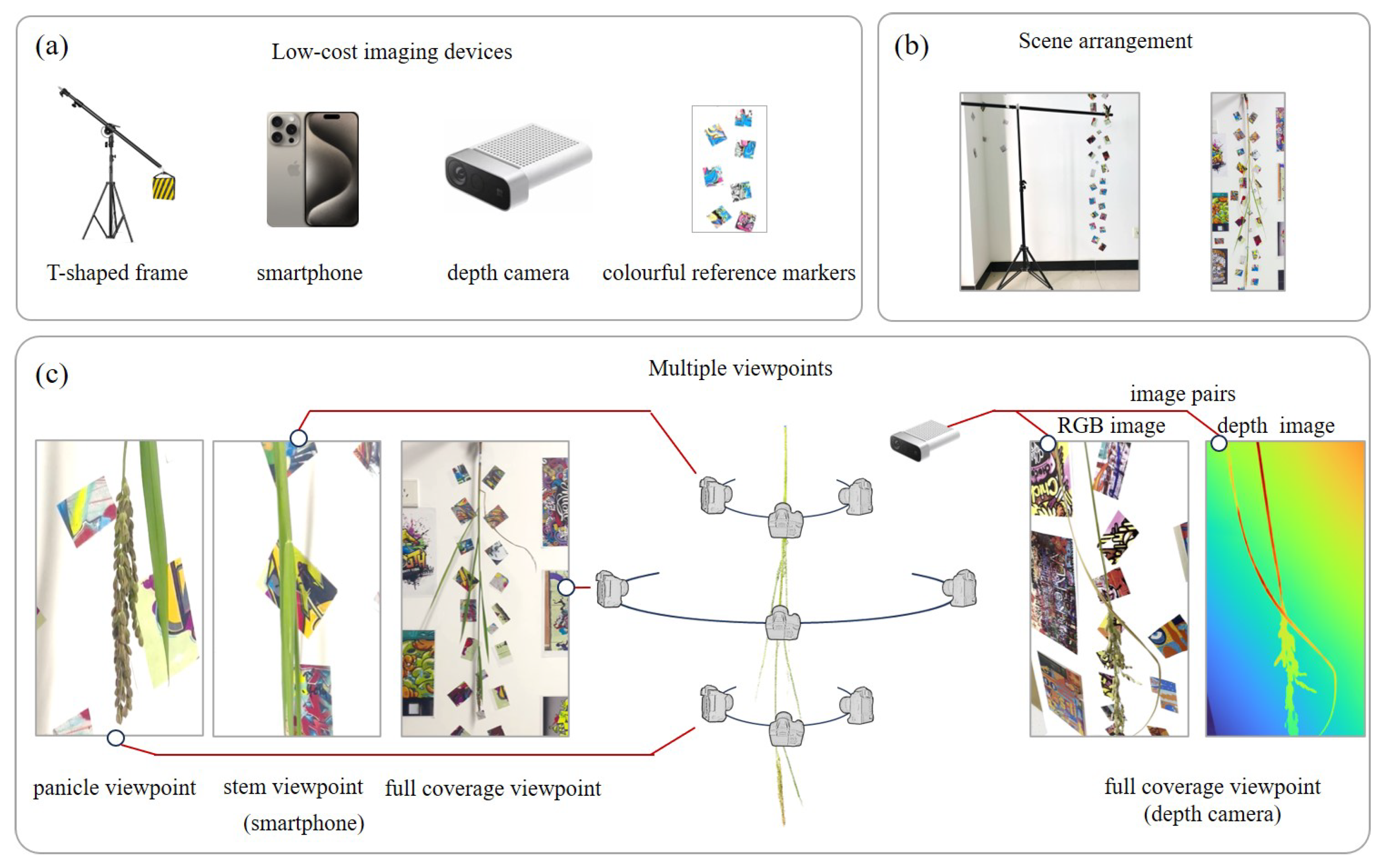
2.2. Reference Markers for Feature Points Matching Enhancement
2.3. Plant Materials
2.4. Multi-View Image Capturing
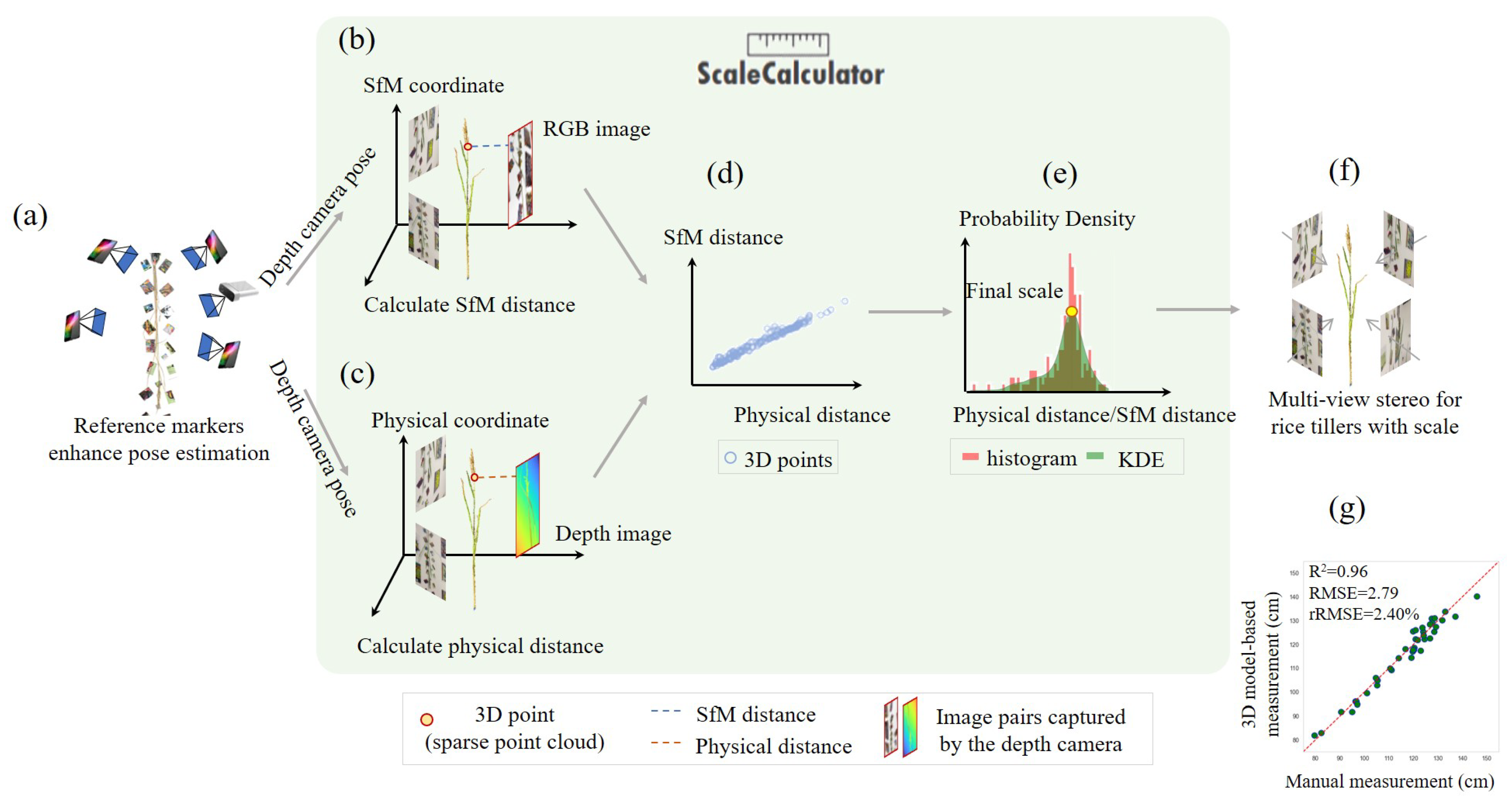
2.5. Recovering the Physical Scale for 3D Models
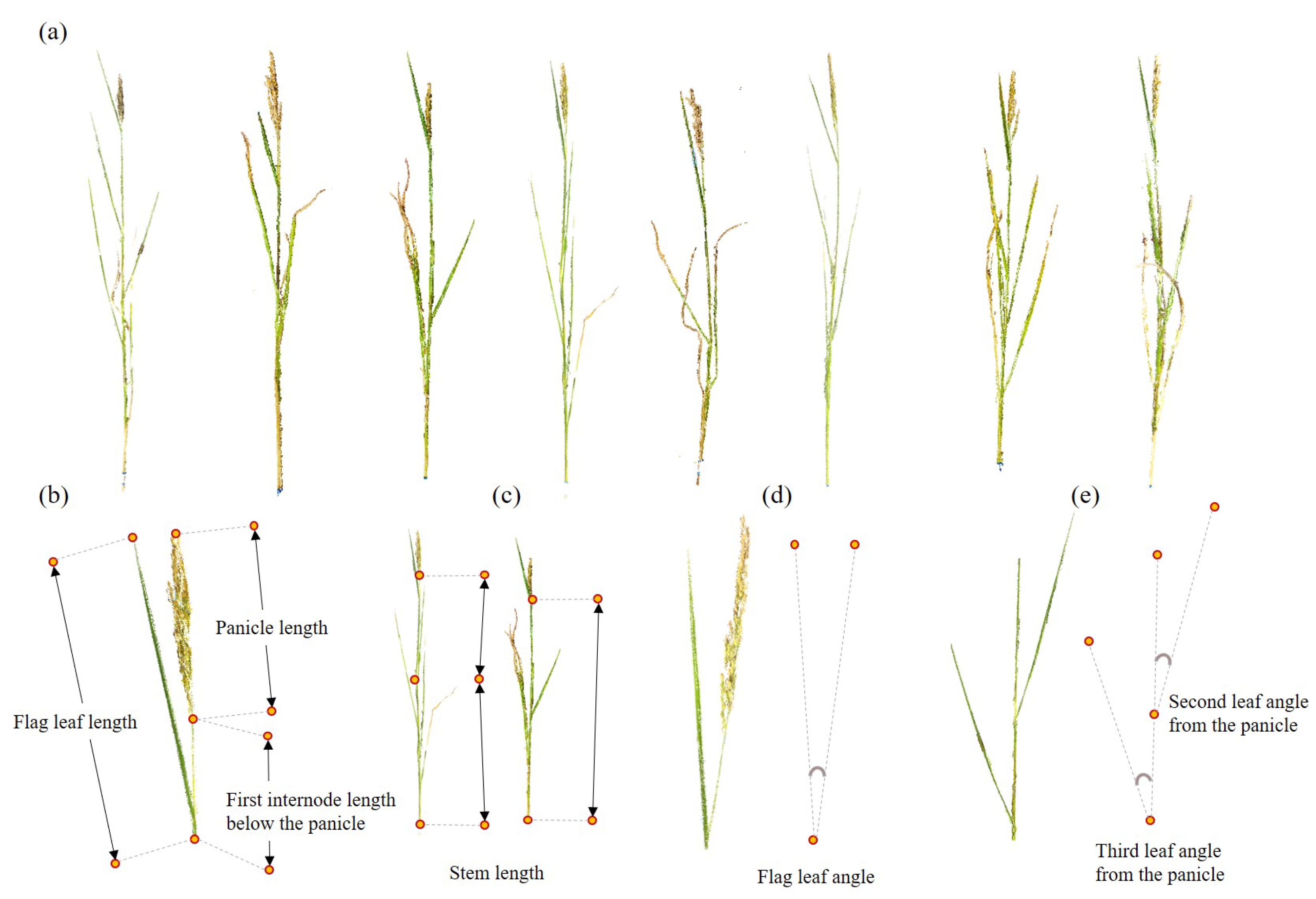
2.6. Three-Dimensional Model-Based Phenotypic Trait Extraction
2.7. Genome-Wide Association Study
3. Results
3.1. High-Resolution 3D Rice Tiller Models
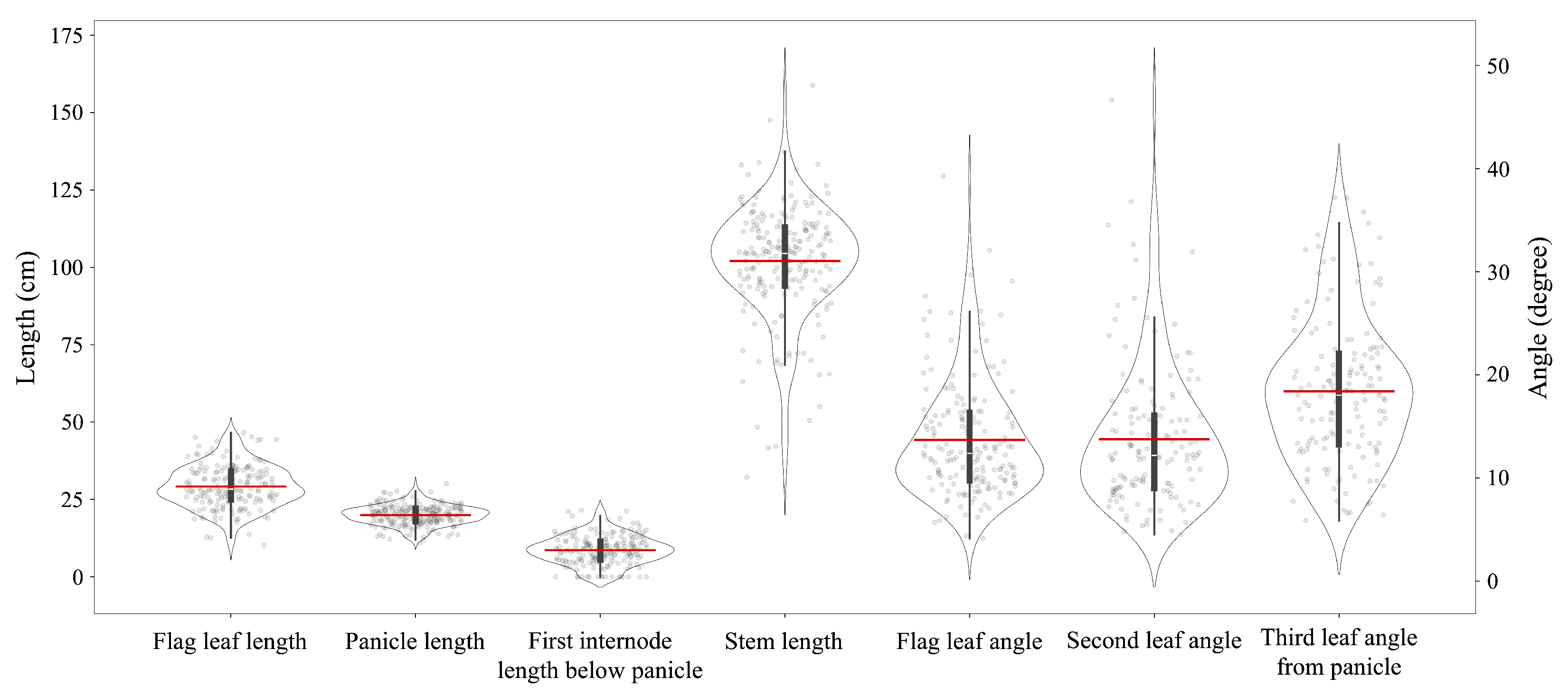
3.2. Phenotypic Traits by 3D Rice Tiller Models
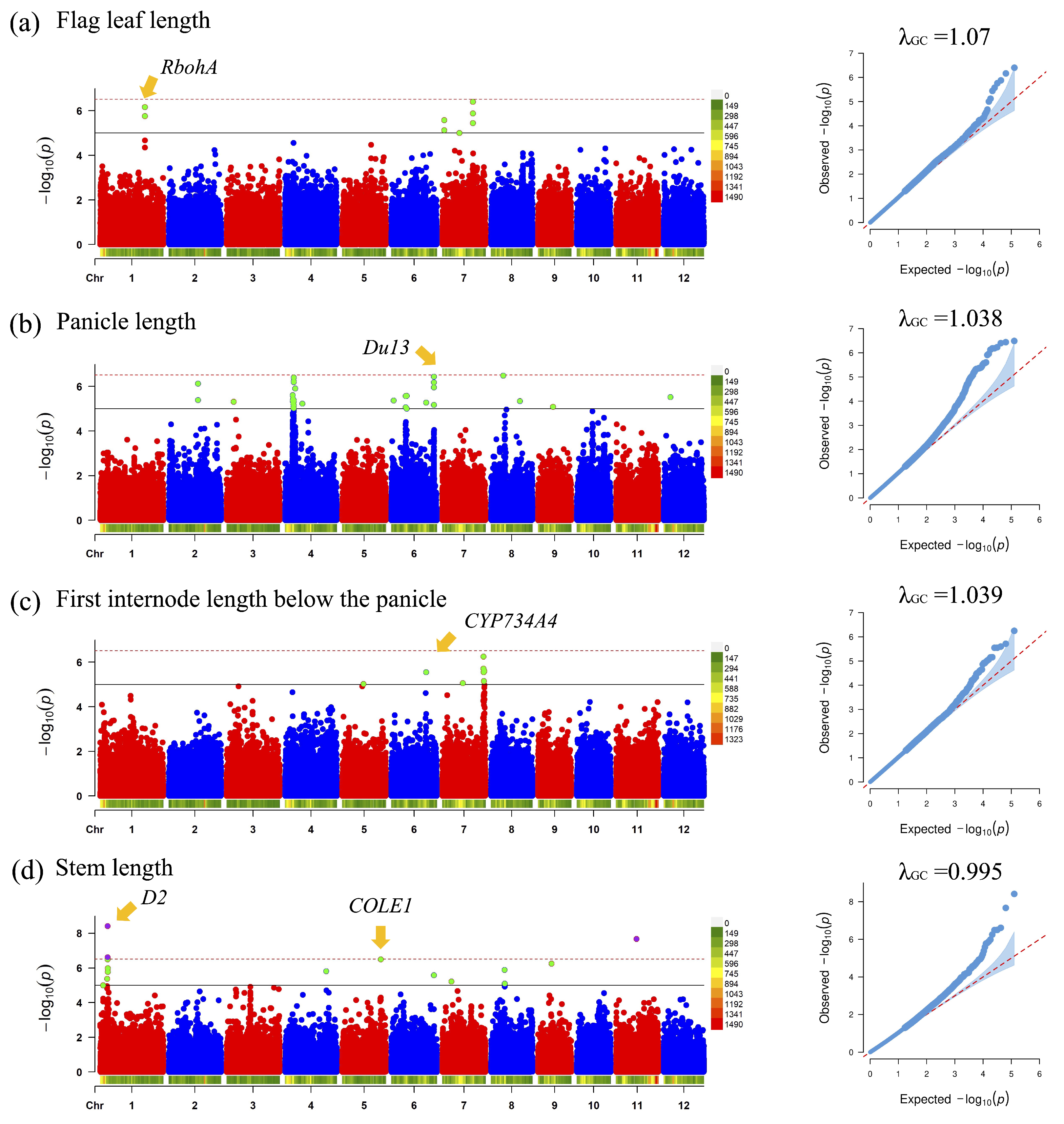
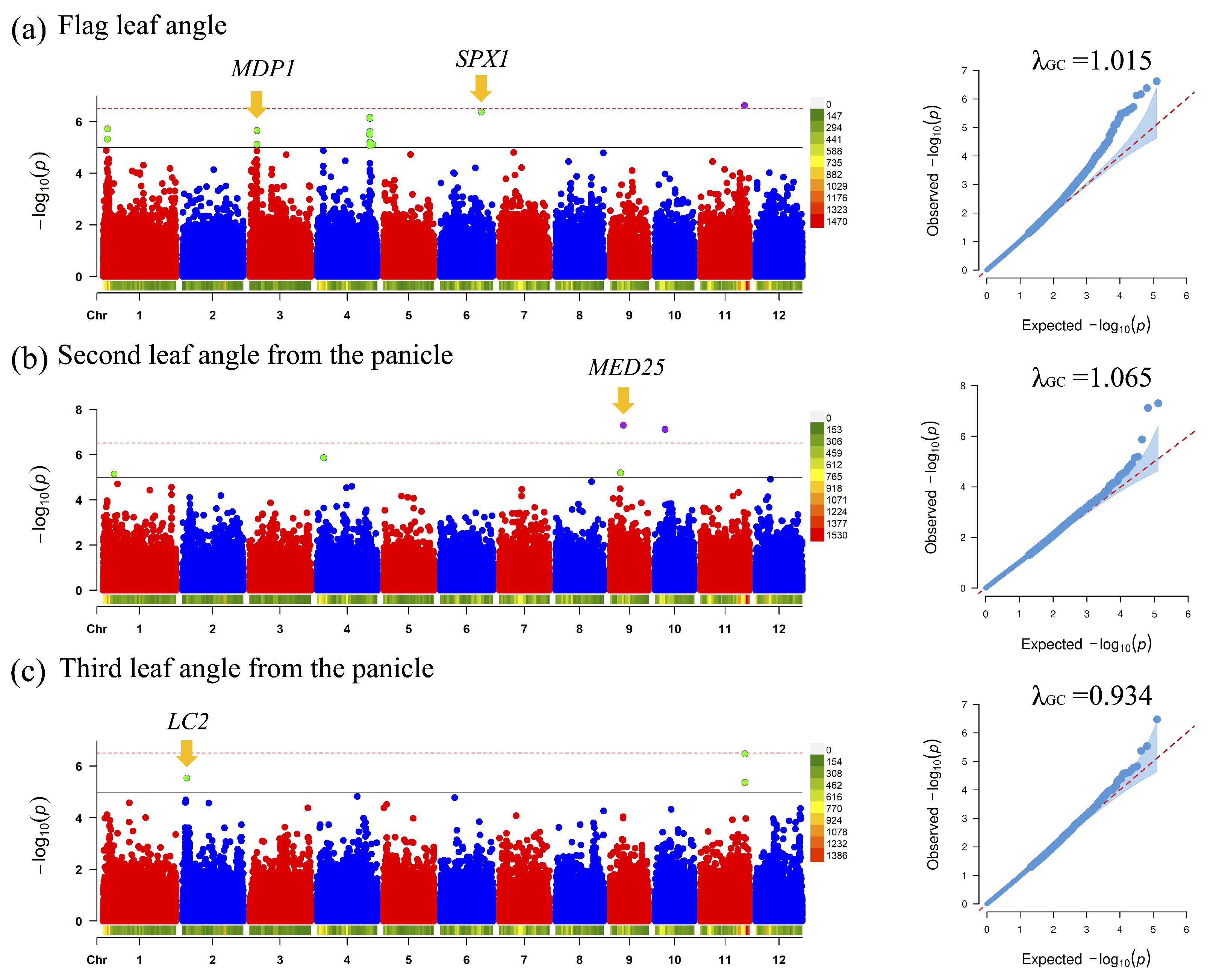
3.3. GWAS for 3D Rice Tiller Phenotypes
4. Discussion
4.1. Three-Dimensional Models for Rice Production
4.2. Applicability to Other Species
4.3. Challenges and Future Directions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT: Production Quantity (Crops). Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 5 July 2025).
- Yuan, R.; Mao, Y.; Zhang, D.; Wang, S.; Zhang, H.; Wu, M.; Ye, M.; Zhang, Z. The formation of rice tillers and factors influencing it. Agronomy 2024, 14, 2904. [Google Scholar] [CrossRef]
- Huang, M.; Shan, S.; Cao, J.; Fang, S.; Tian, A.; Liu, Y.; Cao, F.; Yin, X.; Zou, Y. Primary-tiller panicle number is critical to achieving high grain yields in machine-transplanted hybrid rice. Sci. Rep. 2020, 10, 2811. [Google Scholar] [CrossRef]
- Tsugawa, S.; Shima, H.; Ishimoto, Y.; Ishikawa, K. Thickness-stiffness trade-off improves lodging resistance in rice. Sci. Rep. 2023, 13, 10828. [Google Scholar] [CrossRef]
- Wu, D.; Guo, Z.; Ye, J.; Feng, H.; Liu, J.; Chen, G.; Zheng, J.; Yan, D.; Yang, X.; Xiong, X.; et al. Combining high-throughput micro-CT-RGB phenotyping and genome-wide association study to dissect the genetic architecture of tiller growth in rice. J. Exp. Bot. 2019, 70, 545–561. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xia, P.; Gong, L.; Chen, B.; Li, Y.; Liu, C. Designing an interactively cognitive humanoid field-phenotyping robot for in-field rice tiller counting. Agriculture 2022, 12, 1966. [Google Scholar] [CrossRef]
- Van Crombrugge, I.; Sels, S.; Ribbens, B.; Steenackers, G.; Penne, R.; Vanlanduit, S. Accuracy assessment of joint angles estimated from 2D and 3D camera measurements. Sensors 2022, 22, 1729. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, I.; Shaheen, E.; Aly, M.; Shujaat, S.; Gallo, G.; Coucke, W.; Politis, C.; Jacobs, R. Accuracy and reliability of 2-dimensional photography versus 3-dimensional soft tissue imaging. Imaging Sci. Dent. 2020, 50, 15–22. [Google Scholar] [CrossRef]
- Kenchanmane Raju, S.K.; Adkins, M.; Enersen, A.; Santana de Carvalho, D.; Studer, A.J.; Ganapathysubramanian, B.; Schnable, P.S.; Schnable, J.C. Leaf Angle eXtractor: A high-throughput image processing framework for leaf angle measurements in maize and sorghum. Appl. Plant Sci. 2020, 8, e11385. [Google Scholar] [CrossRef]
- Tross, M.C.; Gaillard, M.; Zwiener, M.; Miao, C.; Grove, R.J.; Li, B.; Benes, B.; Schnable, J.C. 3D reconstruction identifies loci linked to variation in angle of individual sorghum leaves. PeerJ 2021, 9, e12628. [Google Scholar] [CrossRef]
- Xu, J. GrainShape: A landmark-annotated image dataset of japonica rice grains for geometric morphometric analysis. Data Brief 2025, 61, 111781. [Google Scholar] [CrossRef]
- Paturkar, A.; Sen Gupta, G.; Bailey, D. Making use of 3D models for plant physiognomic analysis: A review. Remote Sens. 2021, 13, 2232. [Google Scholar] [CrossRef]
- Wang, K.; Pu, X.; Li, B. Automated phenotypic trait extraction for rice plant using terrestrial laser scanning data. Sensors 2024, 24, 4322. [Google Scholar] [CrossRef]
- Pound, M.P.; French, A.P.; Fozard, J.A.; Murchie, E.H.; Pridmore, T.P. A patch-based approach to 3D plant shoot phenotyping. Mach. Vis. Appl. 2016, 27, 767–779. [Google Scholar] [CrossRef]
- Schonberger, J.L.; Frahm, J.M. Structure-from-motion revisited. In Proceedings of the IEEE Conference on Computer Vision and Pattern Recognition, Las Vegas, NV, USA, 27–30 June 2016; pp. 4104–4113. [Google Scholar]
- Schönberger, J.L.; Zheng, E.; Frahm, J.M.; Pollefeys, M. Pixelwise view selection for unstructured multi-view stereo. In Proceedings of the Computer Vision-ECCV 2016: 14th European Conference, Amsterdam, The Netherlands, 11–14 October 2016; pp. 501–518. [Google Scholar]
- Cernea, D. OpenMVS: Multi-View Stereo Reconstruction Library. Available online: https://github.com/cdcseacave/openMVS (accessed on 28 December 2024).
- Sandhu, J.; Zhu, F.; Paul, P.; Gao, T.; Dhatt, B.K.; Ge, Y.; Staswick, P.; Yu, H.; Walia, H. PI-Plat: A high-resolution image-based 3D reconstruction method to estimate growth dynamics of rice inflorescence traits. Plant Methods 2019, 15, 162. [Google Scholar] [CrossRef]
- Dharni, J.S.; Dhatt, B.K.; Paul, P.; Gao, T.; Awada, T.; Bacher, H.; Peleg, Z.; Staswick, P.; Hupp, J.; Yu, H.; et al. A non-destructive approach for measuring rice panicle-level photosynthetic responses using 3D-image reconstruction. Plant Methods 2022, 18, 126. [Google Scholar] [CrossRef] [PubMed]
- Mildenhall, B.; Srinivasan, P.P.; Tancik, M.; Barron, J.T.; Ramamoorthi, R.; Ng, R. NeRF: Representing scenes as neural radiance fields for view synthesis. In Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2020; pp. 405–421. [Google Scholar]
- Yang, X.; Lu, X.; Xie, P.; Guo, Z.; Fang, H.; Fu, H.; Hu, X.; Sun, Z.; Cen, H. PanicleNeRF: Low-cost, high-precision in-field phenotyping of rice panicles with smartphone. Plant Phenomics 2024, 6, 0279. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Du, X.; Zhu, K.; Lin, K.; Lou, Q.; Yuan, Z.; Huang, G.; Liu, C. Panicle-3D: Efficient phenotyping tool for precise semantic segmentation of rice Panicle point cloud. Plant Phenomics 2021, 2021, 9838929. [Google Scholar] [CrossRef]
- Cui, D.; Liu, P.; Liu, Y.; Zhao, Z.; Feng, J. Automated Phenotypic Analysis of Mature Soybean Using Multi-View Stereo 3D Reconstruction and Point Cloud Segmentation. Agriculture 2025, 15, 175. [Google Scholar] [CrossRef]
- Yang, D.; Yang, H.; Liu, D.; Wang, X. Research on Automatic 3D Reconstruction of Plant Phenotype Based on Multi-View Images. Comput. Electron. Agric. 2024, 220, 108866. [Google Scholar] [CrossRef]
- He, W.; Ye, Z.; Li, M.; Yan, Y.; Lu, W.; Xing, G. Extraction of soybean plant trait parameters based on SfM–MVS algorithm combined with GRNN. Front. Plant Sci. 2023, 14, 1181322. [Google Scholar] [CrossRef]
- Wu, S.; Wen, W.; Gou, W.; Lu, X.; Zhang, W.; Zheng, C.; Xiang, Z.; Chen, L.; Guo, X. A miniaturized phenotyping platform for individual plants using multi-view stereo 3D reconstruction. Front. Plant Sci. 2022, 13, 897746. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Yang, T.; A, X.; Liu, J. Incremental Structure from Motion for Small-Scale Scenes Based on Auxiliary Calibration. Sensors 2025, 25, 415. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, W.; Tang, Z.; Gan, X. A One Stop 3D Target Reconstruction and multilevel Segmentation Method. arXiv 2023, arXiv:2308.06974. [Google Scholar] [CrossRef]
- Laina, I.; Rupprecht, C.; Belagiannis, V.; Tombari, F.; Navab, N. Deeper depth prediction with fully convolutional residual networks. In Proceedings of the 2016 4th International Conference on 3D Vision (3DV), Stanford, CA, USA, 25–28 October 2016; pp. 239–248. [Google Scholar]
- Tölgyessy, M.; Dekan, M.; Chovanec, Ľ.; Hubinský, P. Evaluation of the Azure Kinect and its comparison to Kinect V1 and Kinect V2. Sensors 2021, 21, 413. [Google Scholar] [CrossRef] [PubMed]
- CloudCompare (Version 2.14.alpha): 3D Point Cloud and Mesh Processing Software. Available online: https://www.danielgm.net/cc/ (accessed on 6 July 2025).
- Zheng, H.; Tang, W.; Yang, T.; Zhou, M.; Guo, C.; Cheng, T.; Cao, W.; Zhu, Y.; Zhang, Y.; Yao, X. Grain protein content phenotyping in rice via hyperspectral imaging technology and a genome-wide association study. Plant Phenomics 2024, 6, 0200. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Stephens, M. Genome-wide efficient mixed-model analysis for association studies. Nat. Genet. 2012, 44, 821–824. [Google Scholar] [CrossRef] [PubMed]
- Uffelmann, E.; Huang, Q.Q.; Munung, N.S.; De Vries, J.; Okada, Y.; Martin, A.R.; Martin, H.C.; Lappalainen, T.; Posthuma, D. Genome-wide association studies. Nat. Rev. Methods Prim. 2021, 1, 59. [Google Scholar] [CrossRef]
- Alamin, M.; Sultana, M.H.; Lou, X.; Jin, W.; Xu, H. Dissecting Complex Traits Using Omics Data: A Review on the Linear Mixed Models and Their Application in GWAS. Plants 2022, 11, 3277. [Google Scholar] [CrossRef]
- Castel, S.E.; Tluway, F.D.; Emde, A.K.; Smyth, N.; Karim, M.; Sengupta, D.; Gray, O.A.; Hendershott, M.; LeBaron von Baeyer, S.; Burke, E.E.; et al. A map of blood regulatory variation in South Africans enables GWAS interpretation. Nat. Genet. 2025, 57, 1628–1637. [Google Scholar] [CrossRef]
- Xu, X.; Ye, J.; Yang, Y.; Zhang, M.; Xu, Q.; Feng, Y.; Yuan, X.; Yu, H.; Wang, Y.; Yang, Y.; et al. Genome-wide association study of rice rooting ability at the seedling stage. Rice 2020, 13, 59. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.M.; Wang, Y.J.; Gao, Y.T.; Li, R.; Wang, G.F.; Li, W.Q.; Liu, W.T.; Chen, K.M. The plasma membrane NADPH oxidase OsRbohA plays a crucial role in developmental regulation and drought-stress response in rice. Physiol. Plant. 2016, 156, 421–443. [Google Scholar] [CrossRef]
- Cai, Y.; Zhang, W.; Fu, Y.; Shan, Z.; Xu, J.; Wang, P.; Kong, F.; Jin, J.; Yan, H.; Ge, X.; et al. Du13 encodes a C2 H2 zinc-finger protein that regulates Wxb pre-mRNA splicing and microRNA biogenesis in rice endosperm. Plant Biotechnol. J. 2022, 20, 1387–1401. [Google Scholar] [CrossRef]
- Zhan, C.; Hu, J.; Pang, Q.; Yang, B.; Cheng, Y.; Xu, E.; Zhu, P.; Li, Y.; Zhang, H.; Cheng, J. Genome-wide association analysis of panicle exsertion and uppermost internode in rice (Oryza sativa L.). Rice 2019, 12, 72. [Google Scholar] [CrossRef]
- Hong, Z.; Ueguchi-Tanaka, M.; Umemura, K.; Uozu, S.; Fujioka, S.; Takatsuto, S.; Yoshida, S.; Ashikari, M.; Kitano, H.; Matsuoka, M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell 2003, 15, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, L.; Luo, Y.; Xu, M.; Fan, Y.; Wang, L. Interactions of Oryza sativa Os CONTINUOUS VASCULAR RING-LIKE 1 (Os COLE1) and Os COLE1-INTERACTING PROTEIN reveal a novel intracellular auxin transport mechanism. New Phytol. 2016, 212, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Duan, K.; Li, L.; Hu, P.; Xu, S.P.; Xu, Z.H.; Xue, H.W. A brassinolide-suppressed rice MADS-box transcription factor, OsMDP1, has a negative regulatory role in BR signaling. Plant J. 2006, 47, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Guo, M.; Xu, L.; Wang, X.; Zhao, H.; Wang, J.; Yi, K. An SPX-RLI1 module regulates leaf inclination in response to phosphate availability in rice. Plant Cell 2018, 30, 853–870. [Google Scholar] [CrossRef]
- Karkute, S.G.; Kumar, V.; Tasleem, M.; Mishra, D.C.; Chaturvedi, K.K.; Rai, A.; Sevanthi, A.M.; Gaikwad, K.; Sharma, T.R.; Solanke, A.U. Genome-wide analysis of von Willebrand factor A gene family in rice for its role in imparting biotic stress resistance with emphasis on rice blast disease. Rice Sci. 2022, 29, 375–384. [Google Scholar] [CrossRef]
- Zhao, S.Q.; Hu, J.; Guo, L.B.; Qian, Q.; Xue, H.W. Rice leaf inclination2, a VIN3-like protein, regulates leaf angle through modulating cell division of the collar. Cell Res. 2010, 20, 935–947. [Google Scholar] [CrossRef]
- Organisation for Economic Co-operation and Development; Food and Agriculture Organization of the United Nations (OECD/FAO). OECD Agriculture Statistics (Database): OECD-FAO Agricultural Outlook 2022–2031. Available online: https://www.oecd.org/en/publications/oecd-fao-agricultural-outlook-2022-2031_f1b0b29c-en.html (accessed on 5 July 2025).

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Lee, J.; Jiang, G.; Gan, X. High-Resolution 3D Reconstruction of Individual Rice Tillers for Genetic Studies. Agronomy 2025, 15, 1803. https://doi.org/10.3390/agronomy15081803
Xu J, Lee J, Jiang G, Gan X. High-Resolution 3D Reconstruction of Individual Rice Tillers for Genetic Studies. Agronomy. 2025; 15(8):1803. https://doi.org/10.3390/agronomy15081803
Chicago/Turabian StyleXu, Jiexiong, Jiyoung Lee, Gang Jiang, and Xiangchao Gan. 2025. "High-Resolution 3D Reconstruction of Individual Rice Tillers for Genetic Studies" Agronomy 15, no. 8: 1803. https://doi.org/10.3390/agronomy15081803
APA StyleXu, J., Lee, J., Jiang, G., & Gan, X. (2025). High-Resolution 3D Reconstruction of Individual Rice Tillers for Genetic Studies. Agronomy, 15(8), 1803. https://doi.org/10.3390/agronomy15081803