
Effect of Planting Density and Harvesting Age on Iris pallida Lam. Biomass, Morphology and Orris Concrete Production

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
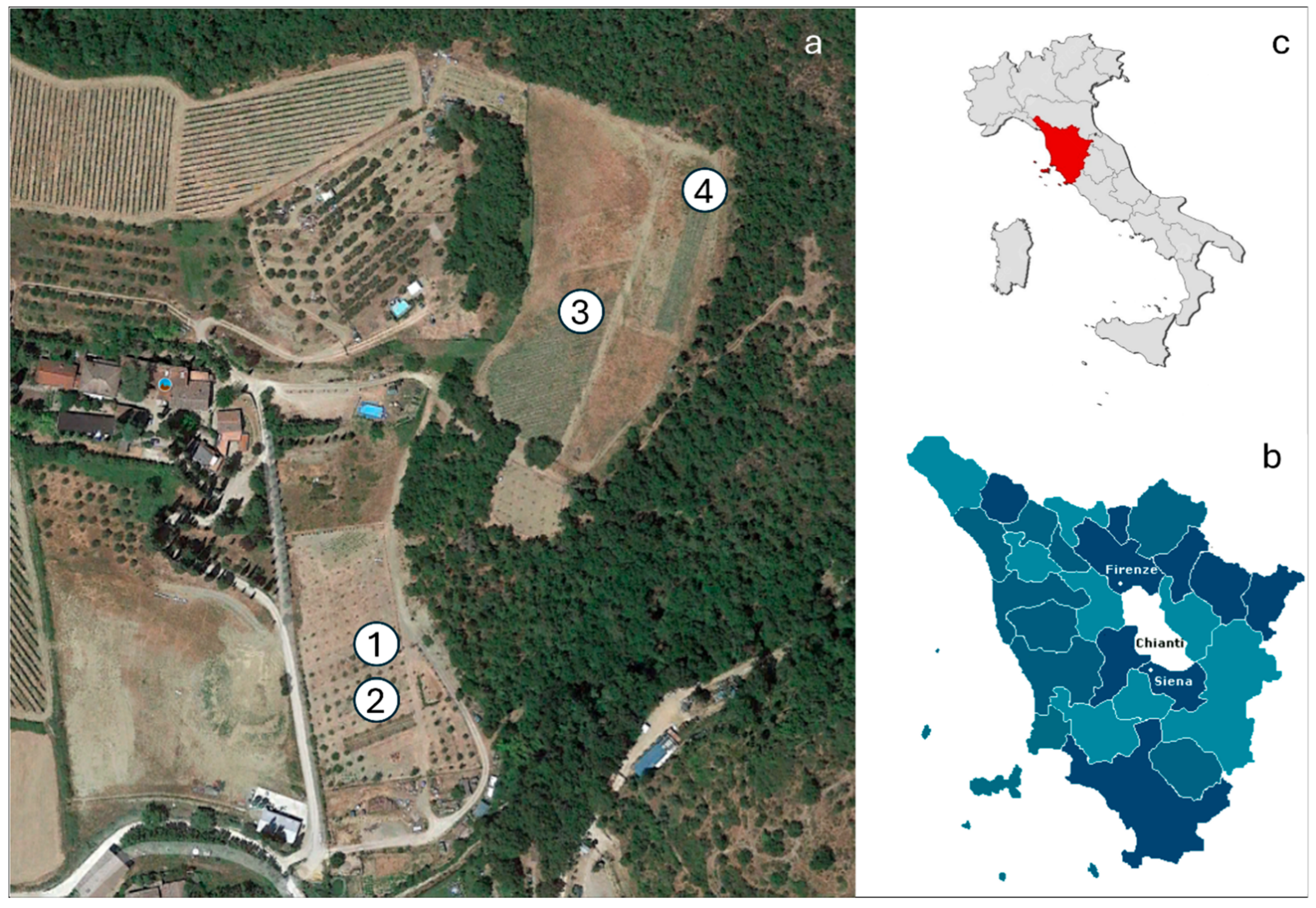
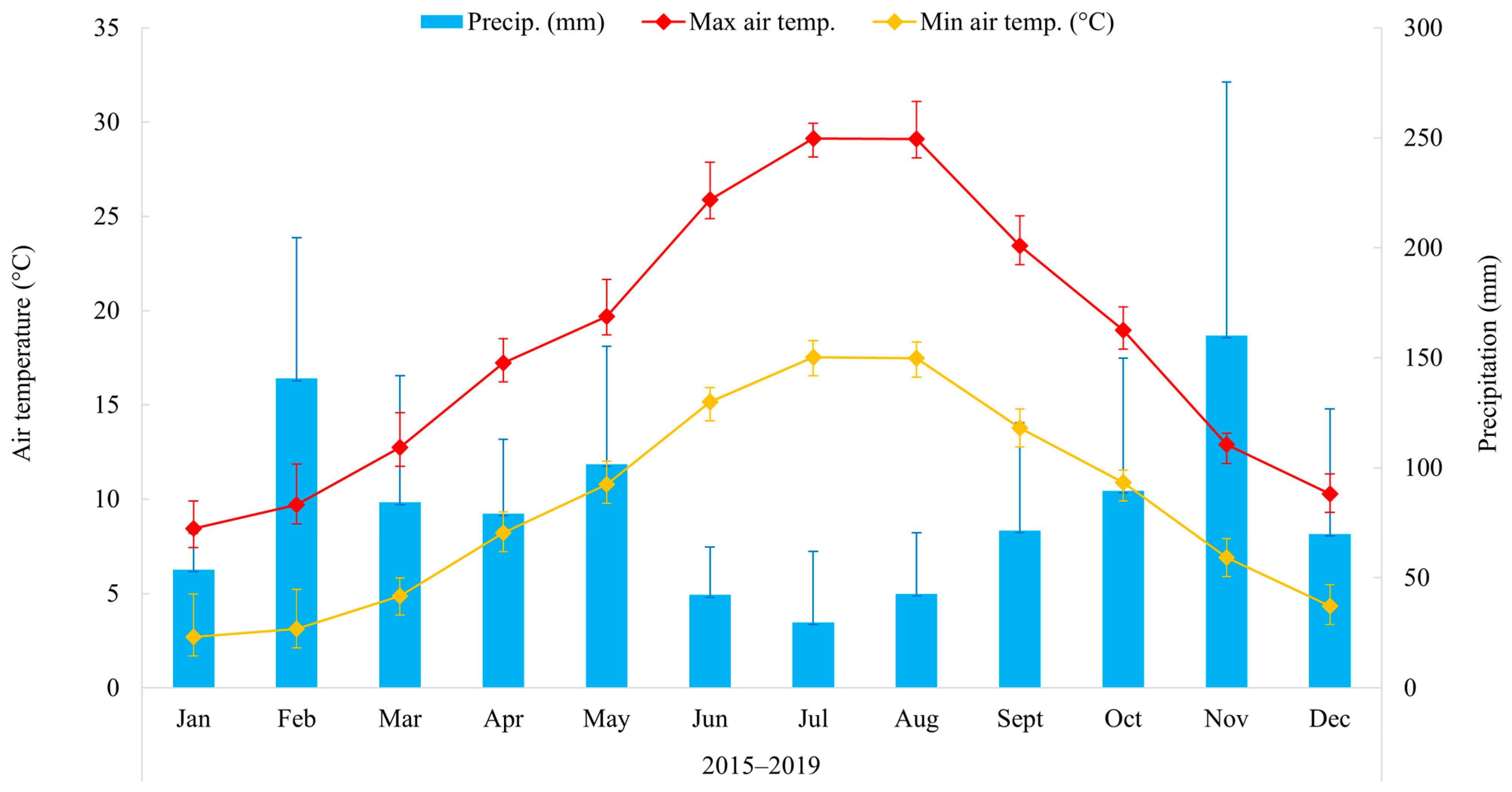
2.1. Study Area
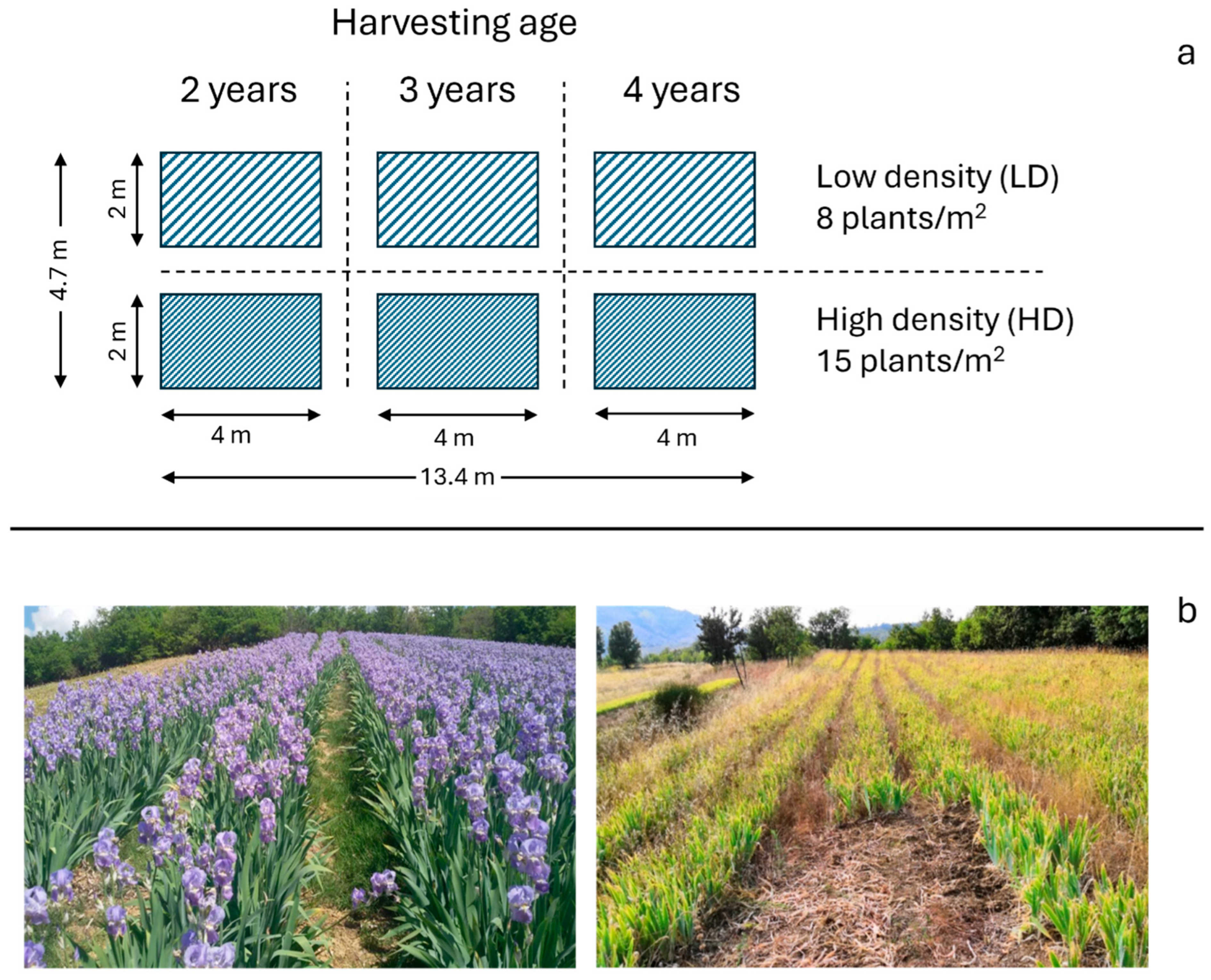
2.2. Experimental Design
2.3. Biomass Data
2.4. Orris Concrete Extraction
2.5. Statistical Analysis
3. Results
3.1. Biomass
3.2. Morphological Traits
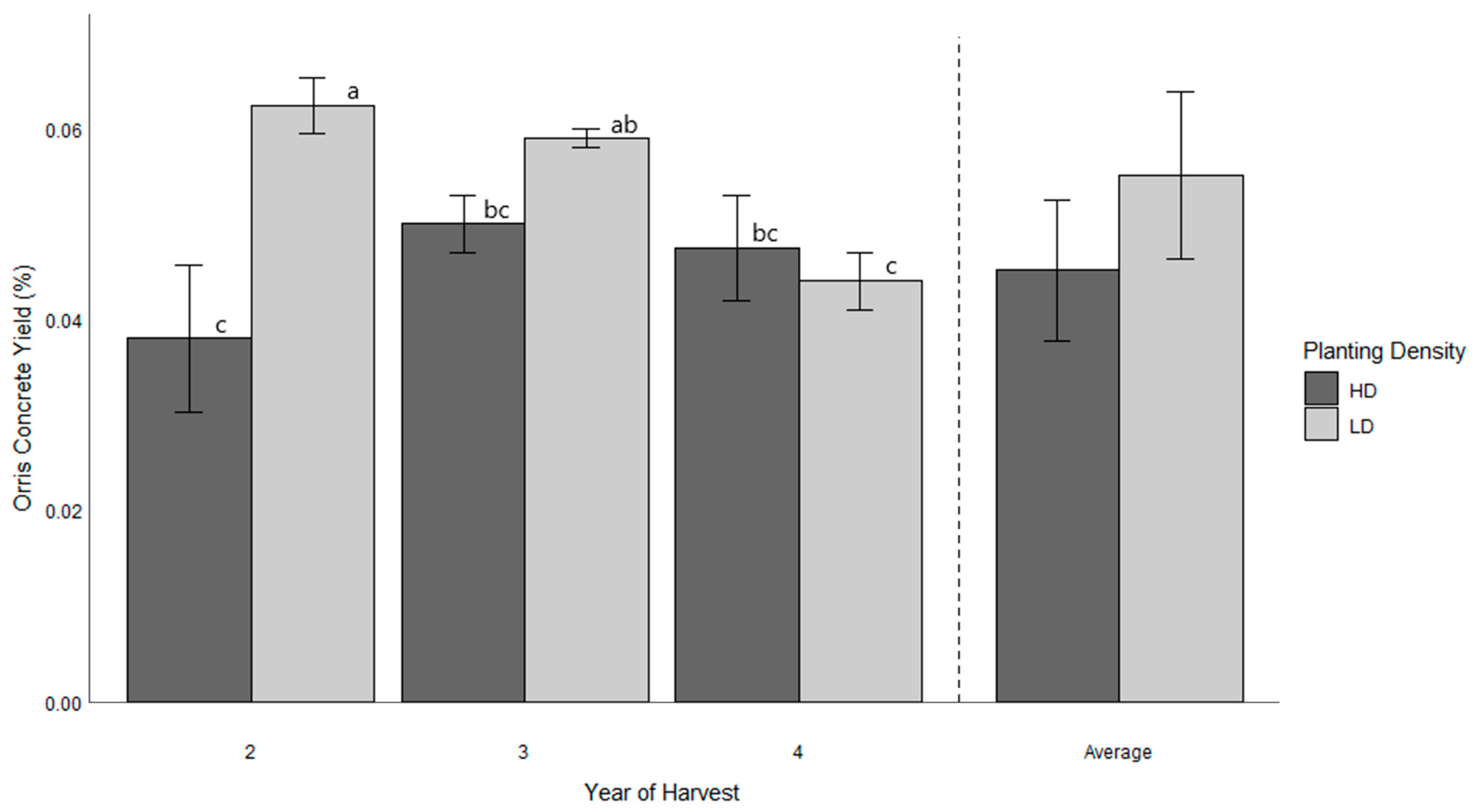
3.3. Orris Concrete Productivity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamenetsky, R.; Okubo, H. Ornamental Geophytes: From Basic Science to Sustainable Production; Kamenetsky, R., Okubo, H., Eds.; Taylor & Francis: Boca Raton, Florida, 2012; p. 597, eBook; ISBN 9780429061776. [Google Scholar] [CrossRef]
- Lucchesini, M.; Bedini, L.; Florio, E.F.; Maggini, R.; Malorgio, F.; Pezzarossa, B.; Mensuali-Sodi, A. The improvement of Iris pallida propagation by somatic embryogenesis. Acta Hortic. 2017, 1155, 127–134. [Google Scholar] [CrossRef]
- Belletti, G.; Fani, E.; Marescotti, A.; Scaramuzzi, S. The role of traditional products in the valorisation of marginal rural areas: The case of Iris pallida. In Proceedings of the 10th European IFSA Symposium, Aarhus, Danmark, 1–4 July 2012. [Google Scholar]
- Roger, B.; Jeannot, V.; Fernandez, X.; Cerantola, S.; Chahboun, J. Characterisation and quantification of flavonoids in Iris Germanica L. and Iris Pallida Lam. resinoids from Morocco. Phytochem. Anal. 2012, 23, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Kaššák, P. Secondary metabolites of the choosen genus Iris species. Acta Univ. Agric. Silvic. Mendel. Brun. 2013, 60, 269–280. [Google Scholar] [CrossRef]
- Masson, J.; Liberto, E.; Brevard, H.; Bicchi, C.; Rubiolo, P. A metabolomic approach to quality determination and authentication of raw plant material in the fragrance field. J. Chromatogr. 2014, 14, 143–154. [Google Scholar] [CrossRef]
- Kukula-Koch, W.; Sieniawska, E.; Widelski, J.; Urjin, O.; Głowniak, P.; Skalicka-Woźniak, K. Major secondary metabolites of Iris spp. Phytochem. Rev. 2015, 14, 51–80. [Google Scholar] [CrossRef]
- Mykhailenko, O. Composition of volatile oil of Iris pallida Lam. from Ukraine. Turk. J. Pharm. Sci. 2018, 15, 85–90. [Google Scholar] [CrossRef]
- Jthan, H.; Courtois, D.; Ehret, C.; Lerch, K.; Pttiard, V. Plant regeneration of Iris pallida Lam. and Iris germanica L. via somatic embryogenesis from leaves, apices and young flowers. Plant Cell Rep. 1994, 13, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Pezzarossa, B.; Borghesi, E.; Pini, R.; Bretzel, F.; Maggini, R.; Malorgio, F. Influence of pedo-climatic conditions on the quality of Iris pallida rhizomes. Eur. J. Hortic. Sci. 2020, 85, 100–109. [Google Scholar] [CrossRef]
- Pezzarossa, B.; Bretzel, F.; Malorgio, F.; Borghesi, E.; Maggini, R.; Scaramuzzi, S. La coltivazione del giaggiolo in Toscana: Dalla marginalità alla valorizzazione. Colt. Protette 2016, 7/8, 62–66. [Google Scholar]
- Landi, R.; Nicolelli, M. Tecnica colturale del giaggiolo (otto anni di sperimentazione a Caspri). In Estratto dal Bollettino Della Società Italiana Dell’iris; Società Italiana Dell’iris: Firenze, Italy, 1977; p. 28. [Google Scholar]
- Marner, F.J.; Kerp, B. Composition of iridals, unusual triterpenoids from sword-Lilies, and the seasonal dependence of their content in various parts of different iris species. Z. Naturforschung C 1992, 47, 21–25. [Google Scholar] [CrossRef]
- Belcour, B.; Courtois, D.; Petiard, V.; Ehret, C. Rapid production of irones by maturation of orris rhizomes with two bacterial strains. Phytochemistry 1993, 34, 1313–1315. [Google Scholar] [CrossRef]
- Firmin, L.; Courtois, D.; Pétiard, V.; Ehret, C.; Lerch, K. Evaluation of the natural variability in irone content and selection of Iris sp. for perfume production. HortScience 1998, 33, 1046–1047. [Google Scholar] [CrossRef]
- Brenna, E.; Fuganti, C.; Ronzani, S.; Serra, S. Enzyme-mediated syntheses of the enantiomers of γ-irones. Helv. Chim. Acta 2001, 84, 3650–3666. [Google Scholar] [CrossRef]
- Konig, W.; Faltin, Y.; Scheffer, J.; Schoffler, H.; Braun, V. Role of Cell-Bound Hemolysin as a Pathogenicity Factor for Serratia Infections. Infect. Immun. 1987, 55, 2554–2561. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tash, K. Rahnella aquatilis bacteremia from a suspected urinary source. J. Clin. Microbiol. 2005, 43, 2526–2528. [Google Scholar] [CrossRef] [PubMed]
- Azam, M.W.; Khan, A.U. Updates on the pathogenicity status of Pseudomonas aeruginosa. Drug Discov. Today 2019, 24, 350–359. [Google Scholar] [CrossRef]
- Ieri, F.; Vignolini, P.; Urciuoli, S.; Pinelli, P.; Romani, A. The Cultivation of Iris pallida as an Opportunity for the Enhancement of Tuscan Agro-Biodiversity and a Resource for the Local Economy. In Innovation, Quality and Sustainability for a Resilient Circular Economy; Lagioia, G., Paiano, A., Amicarelli, V., Gallucci, T., Ingrao, C., Eds.; Springer: Cham, Switzerland, 2024. [Google Scholar] [CrossRef]
- Meucci, A.; Ghelardi, C.; Maggini, R.; Malorgio, F.; Pezzarossa, B.; Rosellini, I.; Mensuali, A. Insights into orris (Iris pallida Lam.) in vivo acclimatization and response to salt stress via exogenous melatonin application. Agriculture 2024, 14, 2353. [Google Scholar] [CrossRef]
- Gürbüzer, G.; Kara, N. Effect of Harvest Times on Rhizoma Yield, Essential Oil Content and Composition in Iris germanica L. Species. Turk. J. Agric. Food Sci. Technol. 2019, 7, 707–713. [Google Scholar] [CrossRef]
- Alsheikly, A.A. Effect of growing media on growth, flowering and bulbs production of Iris hollandica. Diyala Agric. Sci. J. 2010, 5, 581–592. [Google Scholar]
- Mahgoub, H.J.; Eid, R.A.; Abou Leila, B.H. Response of Iris bulbs grown in sandy soil to nitrogen and potassium fertilization. J. Appl. Sci. Res. 2006, 2, 899–903. [Google Scholar]
- Garnero, J.; Joulain, D.; Buil, P. De l’influence du stockage des rhizomes d’iris sur la composition de l’huile essentielle ou beurre d’iris et quelques constituants inédits. Riv. Ital. EPPOS 1978, 10, 568–590. [Google Scholar]
- TGSC Information System, 2022. Orris Rhizome Concrete Butter (Iris pallida). CAS Number: 8002-73-1. Available online: http://www.thegoodscentscompany.com/data/co1001091.html (accessed on 4 January 2025).
- RStudio Team. RStudio: Integrated Development Environment for R. 2021. Available online: http://www.rstudio.com/ (accessed on 17 January 2025).
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Razali, N.M.; Wah, Y.B. Power comparisons of Shapiro–Wilk, Kolmogorov–Smirnov, Lilliefors and Anderson–Darling tests. J. Stat. Model. Anal. 2011, 2, 21–33. [Google Scholar]
- Gastwirth, J.L.; Gel, Y.R.; Miao, W. The impact of Levene’s test of equality of variances on statistical theory and practice. Stat. Sci. 2009, 24, 343–360. [Google Scholar] [CrossRef]
- Graves, S.; Piepho, H.P.; Selzer, L.; Dorai-Raj, S. multcompView: Visualizations of Paired Comparisons. 2023. Available online: https://CRAN.R-project.org/package=multcompView (accessed on 20 February 2025).
- Han-Wu-Shuang, B. bruceR: Broadly Useful Convenient and Efficient R Functions. 2023. Available online: https://CRAN.R-project.org/package=bruceR (accessed on 28 March 2025).
- Kratky, B.; Bernabe, C.I. Outdoor growing of clean edible ginger seed by a pot-in-pot-in-pot sub-irrigation method. In Proceedings of the 35th National Agricultural Plastics Congress, American Society for Plasticulture, Bellafonte, PA, USA, 13–16 July 2009; Available online: https://www.ctahr.hawaii.edu/hawaii/downloads/outdoor_growing_of_clean_edible_ginger_seed_by_a_pot-in-pot-in-pot_sub-irrigation_method.pdf (accessed on 13 February 2023).
- Oztas, F.; Turkmen, A.; Oztas, H.; Turkmen, M. The medical properties of Iris and its usage in pharmaceutical, perfumery and cosmetic industries. In Medical Research and Its Applications; Blue Pen International: Chennai, India, 2024; Volume 4, pp. 114–124. [Google Scholar] [CrossRef]
- Retana-Cordero, M.; Flores, S.J.; Fisher, P.R.; Freyre, R.; Gómez, C. Effect of container volume and planting density on ginger and turmeric growth and yield. HortTechnology 2022, 32, 425–434. [Google Scholar] [CrossRef]
- Landi, R. Effetti della popolazione coltivata, della concimazione e dei fattori ambientali sulla produzione del giaggiolo (Iris pallida Lam.). In Proceedings of the Atti Convegno Internazionale “Coltivazione e Miglioramento di Piante Officinali”, Trento, Italy, 2–3 June 1994; pp. 47–70. [Google Scholar]
- Mahender, B.; Reddy, P.S.S.; Sivaram, G.T.; Balakrishna, M.; Prathap, B. Effect of seed rhizome size and plant spacing on growth, yield and quality of ginger (Zingiber officinale Rosc.) under coconut cropping system. Plant Arch. 2015, 15, 769–774. Available online: https://plantarchives.org/pdf%2015-2/769-774%20(3001).pdf (accessed on 14 March 2025).
- Verma, T.; Sharma, B.P.; Thakur, M. Effect of Different Nitrogen Levels and Spacing on Growth and Flowering of Iris (Iris orientalis Mill.) cv.‘Frigia’. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 2101–2109. [Google Scholar] [CrossRef]
- Gerami, F.; Rezvani Moghaddam, P.; Ghorbani, R.; Hassani, A. Influence of planting date and plant density on morphological characteristics, seed yield, and essential oil percentage of oregano (Origanum vulgare L.). J. Appl. Hortic. 2018, 20, 171–176. [Google Scholar] [CrossRef]
- Berimavandi, A.R.; Hashemabadi, D.; Facouri Ghaziani, M.V.; Kaviani, B. Effects of plant density and sowing date on the growth, flowering and quantity of essential oil of Calendula officinalis L. J. Med. Plants Res. 2011, 5, 5110–5115. Available online: http://www.academicjournals.org/JMPR (accessed on 14 March 2025).
- Heuer, B.; Nadler, A. Growth and development of potatoes under salinity and water deficit. Aust. J. Agric. Res. 1995, 46, 1477–1486. [Google Scholar] [CrossRef]
- Häkkinen, S.T.; Oksman-Caldentey, K.M. Progress and prospects of hairy root research. In Hairy Roots: An Effective Tool of Plant Biotechnology; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2018; Chapter 1; pp. 3–19. ISBN 978-981-13-2562-5. [Google Scholar]
- Bahl, J.R.; Bansal, R.P.; Garg, S.N.; Gupta, M.M.; Singh, V.; Goel, R.; Kumar, S. Variation in yield of curcumin and yield and quality of leaf and rhizome essential oils among Indian landraces of turmeric Curcuma longa L. Proc. Indian. Natl Sci. Acad. 2014, 80, 143–156. [Google Scholar] [CrossRef]
- Liu, F.; Shahnazari, A.; Andersen, M.N.; Jacobsen, S.E.; Jensen, C.R. Physiological responses of potato (Solanum tuberosum L.) to partial root-zone drying: ABA signalling, leaf gas exchange, and water use efficiency. J. Exp. Bot. 2006, 57, 3727–3735. [Google Scholar] [CrossRef]
- Meyer, C.J.; Seago, J.L.; Peterson, C.A. Environmental effects on the maturation of the endodermis and multiseriate exodermis of Iris germanica roots. Ann. Bot. 2009, 103, 687–702. [Google Scholar] [CrossRef]
- Bicchi, C.; Joulain, D. A comprehensive review on essential oils and extracts from Iris rhizomes. Phytochem. Rev. 2024, 24, 1629–1665. [Google Scholar] [CrossRef]
- Almaarri, K.; Abou Zedan, T.; Albatal, N. Chemical analysis of essential oils of some Syrian wild Iris species. Am. J. Biochem. Mol. Biol. 2013, 3, 38–49. [Google Scholar] [CrossRef]
- Falster, D.S.; Westoby, M. Leaf size and angle vary widely across species: What consequences for light interception? New Phytol. 2003, 158, 509–525. [Google Scholar] [CrossRef]
- Wright, I.J.; Dong, N.; Maire, V.; Prentice, I.C.; Westoby, M.; Díaz, S.; Gallagher, R.V.; Jacobs, B.F.; Kooyman, R.; Law, E.A.; et al. Functional traits explain variation in plant growth and response to climate across 2000 species from around the globe. Ecol. Res. 2010, 25, 707–723. [Google Scholar] [CrossRef]
- Wollinger, A. Application of a Supercritical Carbon Dioxide Extraction Unit. Ph.D. Thesis, University of Regensburg, Regensburg, Germany, 2016. (In German). Available online: https://epub.uni-regensburg.de/33710/7/Dissertation_Wollinger_18042016.pdf (accessed on 12 February 2025).
- Spadi, A.; Angeloni, G.; Guerrini, L.; Corti, F.; Parenti, A.; Innocenti, M.; Bellumori, M.; Masella, P. Hydrodistillation of Coffee By-Products to Recover Bioactive Compounds: The Spent Coffee Ground and Coffee Silverskin Case-Study. Chem. Eng. Trans. 2021, 87, 313–318. [Google Scholar] [CrossRef]
- Mehalaine, S.; Chenchouni, H. Quantifying how climatic factors influence essential oil yield in wild-growing plants. Arab. J. Geosci. 2021, 14, 1257. [Google Scholar] [CrossRef]
- Zuzarte, M.R.; Dinis, A.M.; Cavaleiro, C.; Salgueiro, L.R.; Canhoto, J.M. Trichomes, essential oils and in vitro propagation of Lavandula pedunculata (Lamiaceae). Ind. Crops Prod. 2010, 32, 580–587. [Google Scholar] [CrossRef]
- Bettaieb, I.; Hamrouni-Sellami, I.; Bourgou, S.; Limam, F.; Marzouk, B. Drought effects on polyphenol composition and antioxidant activities in aerial parts of Salvia officinalis L. Acta Physiol. Plant. 2011, 33, 1103–1111. [Google Scholar] [CrossRef]
- Sun, H.; Wang, Y.; Yang, C.; Liu, J.; Li, J.; Chen, S. Genetic and environmental influences on essential oil composition in Atractylodes lancea. PLoS ONE 2019, 14, e0217522. [Google Scholar] [CrossRef]
- Rawat, S.; Bhatt, I.D.; Rawal, R. Variation in essential oil composition in rhizomes of natural populations of Hedychium spicatum in different environmental conditions and habitats. J. Essent. Oil Res. 2020, 32, 348–360. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Plant Density (D) | Harvest Age (H) | Interaction (D H) | Plot (Random Effect) |
|---|---|---|---|---|
| Total biomass (kg/m2) | *** | *** | ns | 0.228 (moderate) |
| Rhizome biomass (kg/m2) | *** | *** | * | 0.172 (moderate) |
| Root biomass (kg/m2) | ns | *** | ** | 0.035 (negligible) |
| Bud biomass (kg/m2) | *** | *** | ns | 0.000 (negligible) |
| Floral stem biomass (kg/m2) | ** | *** | ** | 0.032 (negligible) |
| Rhizome dry biomass (kg/m2) | *** | ** | ns | 0.015 (negligible) |
| Floral stem number (n/plant) | *** | *** | *** | 0.005 (negligible) |
| Bud number (n/m2) | *** | *** | *** | 0.450 (high) |
| Leaves length/width ratio | ** | *** | ns | 0.003 (negligible) |
| Orris concrete yield (%) | *** | *** | *** | 0.000 (negligible) |
| P. Density | Year | Harvesting Age | Total Fresh Biomass | Rhizome | Roots Fresh | Buds | Floral Stems |
|---|---|---|---|---|---|---|---|
| (D) | (Y) | (H) | (kg/m2) | (kg/m2) | (kg/m2) | (kg/m2) | (kg/m2) |
| Low density (LD) | 2017 | 2 | 3.99 ± 0.36 b | 0.76 ± 0.05 bc | 0.24 ± 0.06 ab | 2.72 ± 0.33 bc | 0.26 ± 0.03 a |
| 2018 | 3 | 6.26 ± 1.07 a | 1.3 ± 0.04 a | 0.32 ± 0.07 a | 4.40 ± 0.97 a | 0.20 ± 0.07 ab | |
| 2019 | 4 | 6.17 ± 0.34 a | 1.70 ± 0.10 a | 0.22 ± 0.04 abc | 4.11 ± 0.48 ab | 0.12 ± 0.03 bc | |
| High density (HD) | 2017 | 2 | 1.09 ± 0.27 c | 0.34 ± 0.08 c | 0.11 ± 0.02 c | 0.56 ± 0.12 d | 0.06 ± 0.03 c |
| 2018 | 3 | 1.96 ± 0.43 c | 0.65 ± 0.18 bc | 0.22 ± 0.02 abc | 1.01 ± 0.20 d | 0.07 ± 0.03 c | |
| 2019 | 4 | 2.4 ± 0.92 bc | 0.82 ± 0.32 b | 0.14 ± 0.02 bc | 1.28 ± 0.56 cd | 0.14 ± 0.05 abc | |
| LD | - - - | Average | 5.48 ± 0.59 | 1.27 ± 0.06 | 0.26 ± 0.05 | 3.74 ± 0.62 | 0.2 ± 0.04 |
| HD | - - - | 1.82 ± 0.54 | 0.60 ± 0.19 | 0.16 ± 0.02 | 0.95 ± 0.3 | 0.09 ± 0.04 |
| P. Density | Year | Harvesting Age | Rhizome Dry Biomass | Floral Stems | Buds | Leaves |
|---|---|---|---|---|---|---|
| (D) | (Y) | (H) | (kg/m2) | (n°/Plant) | (n°/m2) | (Length/Width) |
| Low density (LD) | 2017 | 2 | 0.23 ± 0.01 bc | 1.07 ± 0.10 b | 39.38 ± 2.76 bc | 17.34 ± 1.31 b |
| 2018 | 3 | 0.4 ± 0.01 a | 1.82 ± 0.09 a | 56.46 ± 9.85 ab | 19.74 ± 0.25 a | |
| 2019 | 4 | 0.51 ± 0.03 a | 0.53 ± 0.16 c | 61.42 ± 2.47 a | 18.97 ± 0.64 ab | |
| High density (HD) | 2017 | 2 | 0.10 ± 0.02 c | 0.15 ± 0.09 d | 12.33 ± 2.78 d | 13.87 ± 0.76 c |
| 2018 | 3 | 0.19 ± 0.05 bc | 0.40 ± 0.17 cd | 24.38 ± 4.20 cd | 14.59 ± 0.36 c | |
| 2019 | 4 | 0.24 ± 0.09 b | 0.36 ± 0.03 cd | 31.38 ± 13.10 cd | 13.98 ± 0.86 c | |
| LD | - - - | Average | 0.38 ± 0.02 | 1.14 ± 0.12 | 52.42 ± 5.0 | 18.68 ± 0.73 |
| HD | - - - | 0.18 ± 0.05 | 0.3 ± 0.1 | 22.7 ± 6.7 | 14.14 ± 0.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palchetti, E.; Brilli, L.; Padovan, G.; Mariani, G.; Marini, L.; Moretta, M. Effect of Planting Density and Harvesting Age on Iris pallida Lam. Biomass, Morphology and Orris Concrete Production. Agronomy 2025, 15, 1719. https://doi.org/10.3390/agronomy15071719
Palchetti E, Brilli L, Padovan G, Mariani G, Marini L, Moretta M. Effect of Planting Density and Harvesting Age on Iris pallida Lam. Biomass, Morphology and Orris Concrete Production. Agronomy. 2025; 15(7):1719. https://doi.org/10.3390/agronomy15071719
Chicago/Turabian StylePalchetti, Enrico, Lorenzo Brilli, Gloria Padovan, Gregorio Mariani, Lorenzo Marini, and Michele Moretta. 2025. "Effect of Planting Density and Harvesting Age on Iris pallida Lam. Biomass, Morphology and Orris Concrete Production" Agronomy 15, no. 7: 1719. https://doi.org/10.3390/agronomy15071719
APA StylePalchetti, E., Brilli, L., Padovan, G., Mariani, G., Marini, L., & Moretta, M. (2025). Effect of Planting Density and Harvesting Age on Iris pallida Lam. Biomass, Morphology and Orris Concrete Production. Agronomy, 15(7), 1719. https://doi.org/10.3390/agronomy15071719