Study on Soil Nutrients and Microbial Community Diversity in Ancient Tea Plantations of China


 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Soil Sampling
2.2. Soil Chemical Analysis and Indicator Selection
2.2.1. Indicators of Soil Fertility
2.2.2. Standardization for Soil Fertility Assessment
2.3. Determination of Rhizosphere Soil Microbial Community Diversity
2.3.1. Extraction of Total DNA from Microbiome
2.3.2. PCR Amplification of Target Fragments
2.3.3. Purification and Recovery of Amplified Products Using Magnetic Beads
2.3.4. Fluorescence Quantification of Amplified Products
2.3.5. Sequencing Library Preparation
2.3.6. High-Throughput Sequencing Operation
2.4. Statistical Analyses
3. Results
3.1. Soil Fertility Status
3.2. Diversity and Composition of Microbial Communities
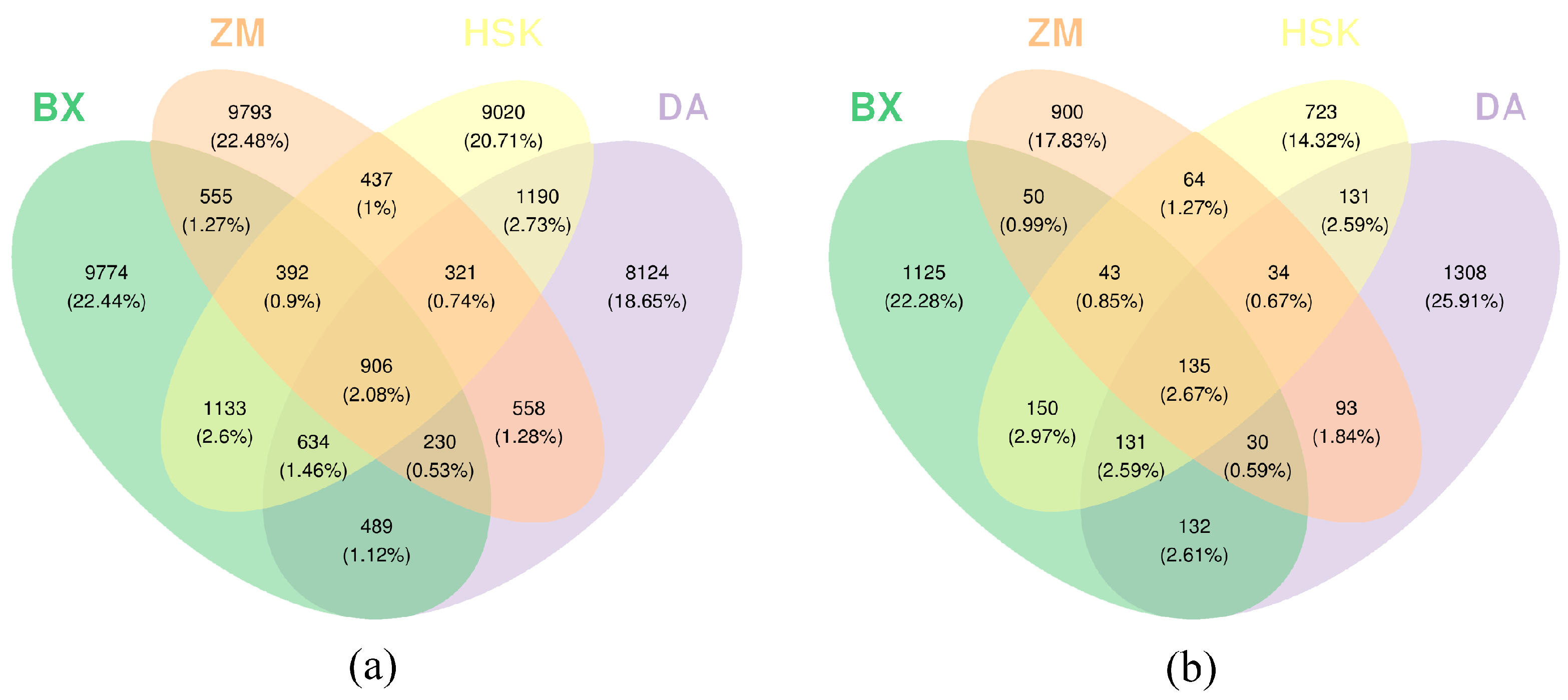
3.2.1. OTU Distribution in Rhizosphere Microbial Communities
3.2.2. Soil Microbial Alpha Diversity
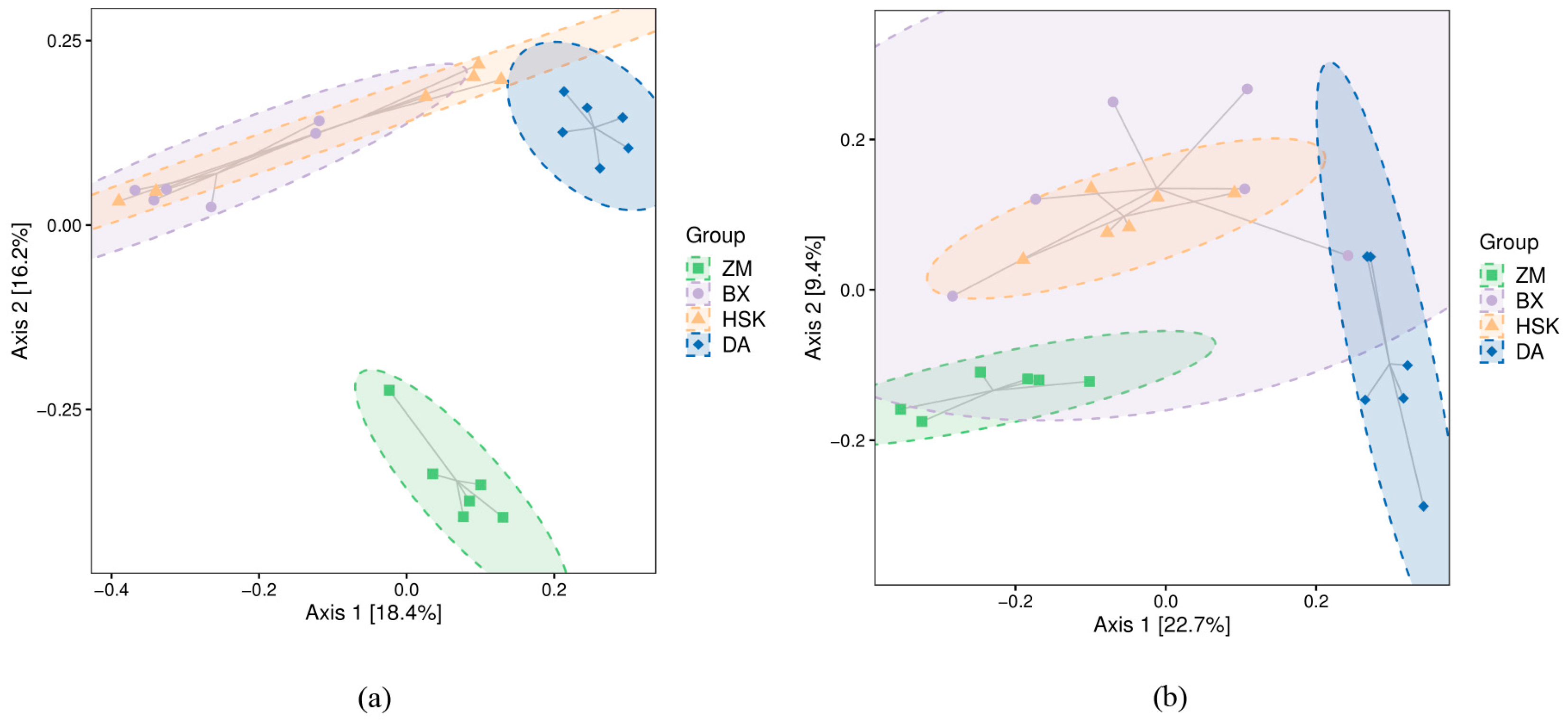
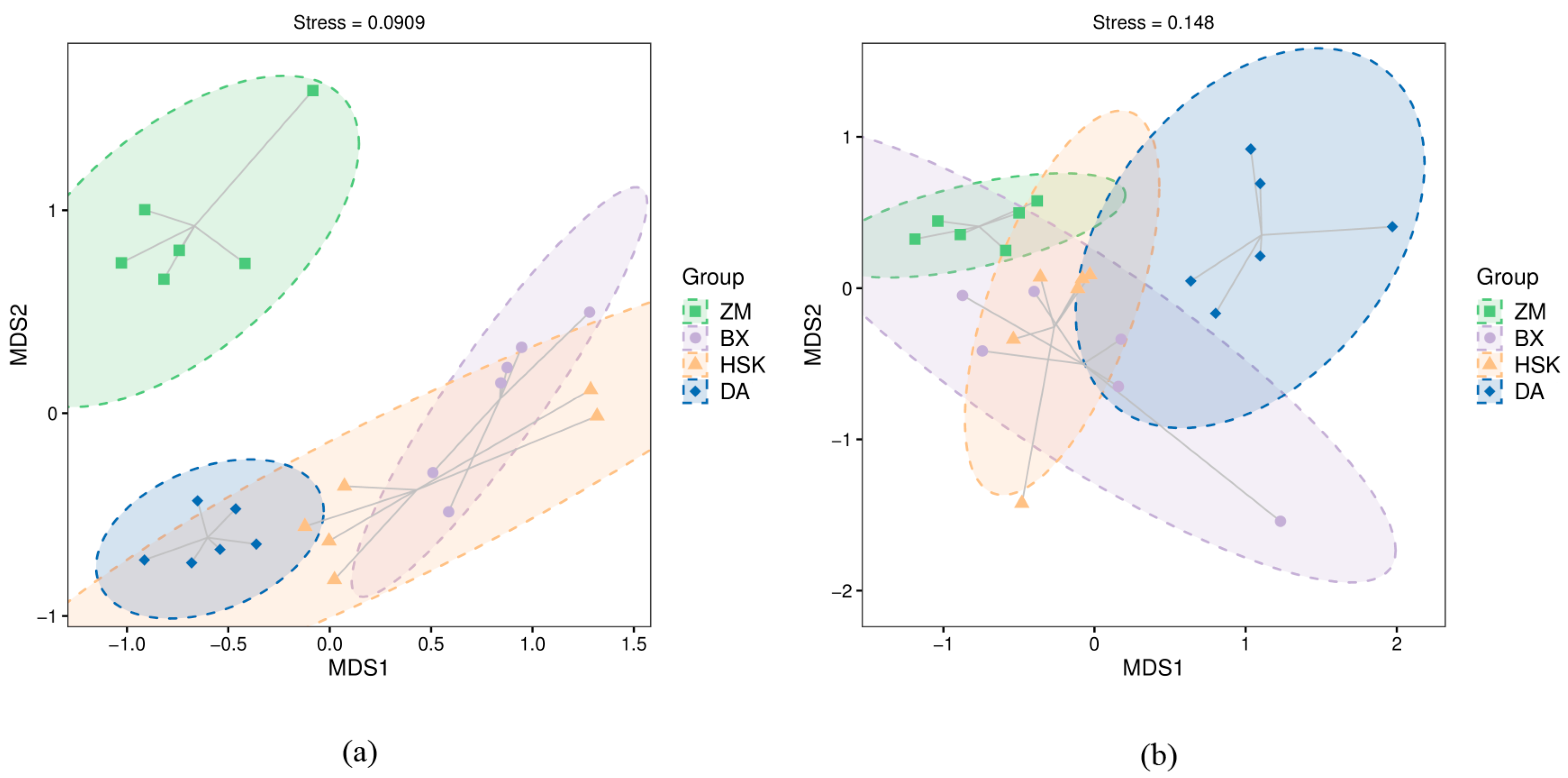
3.2.3. Soil Microbial Beta Diversity
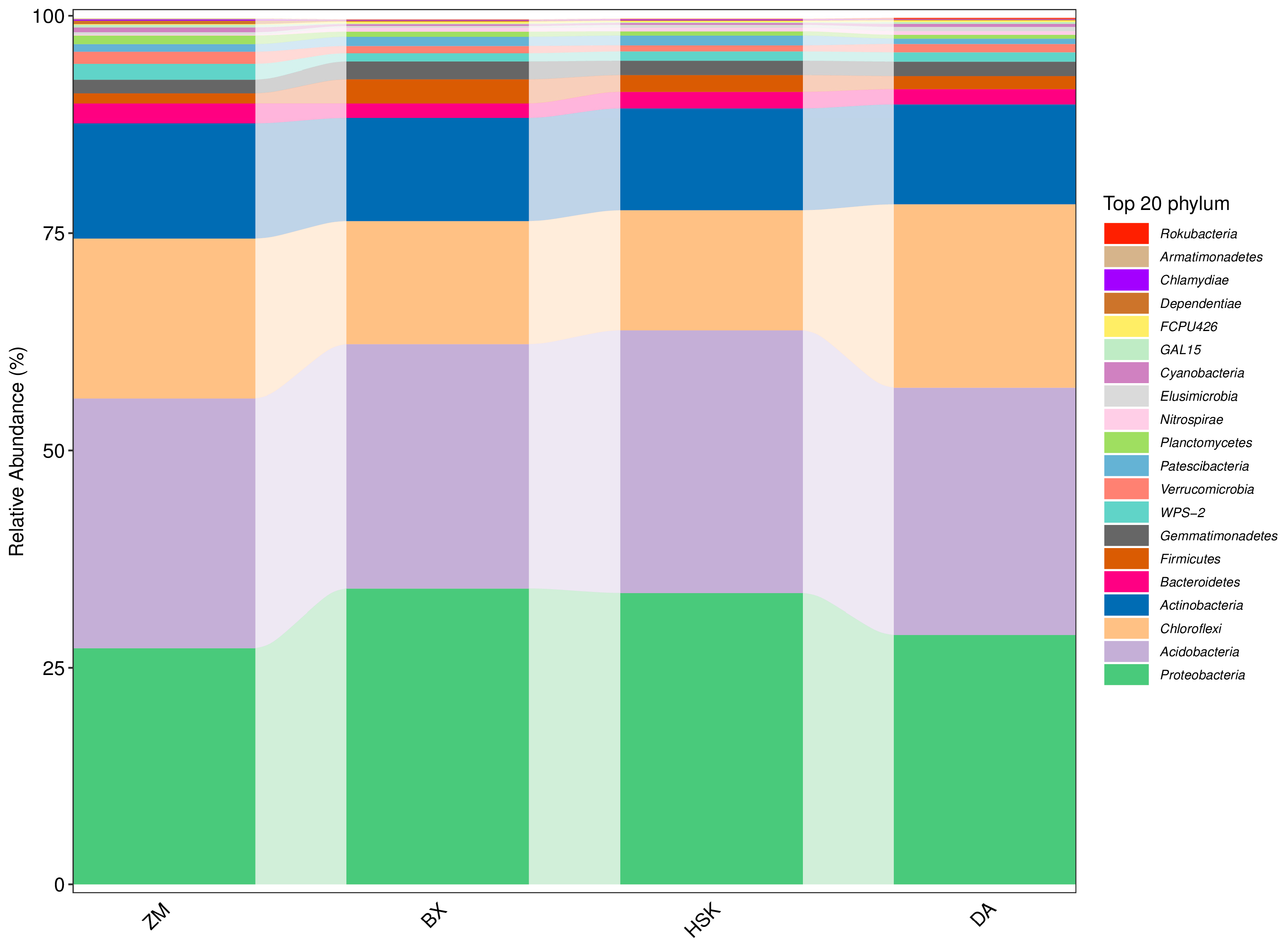
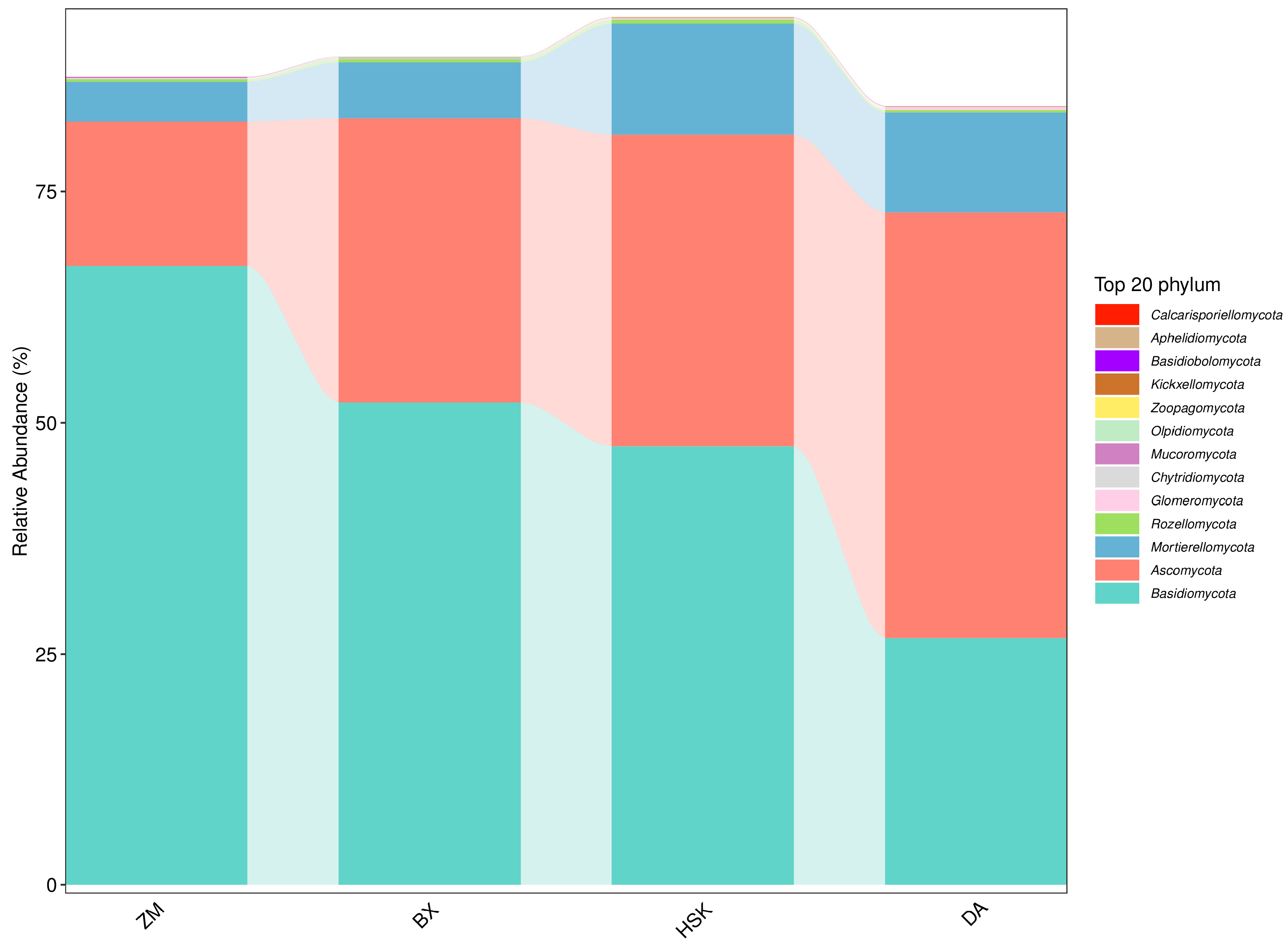
3.2.4. Structural Composition of Soil Bacteria and Fungi at the Phylum Level
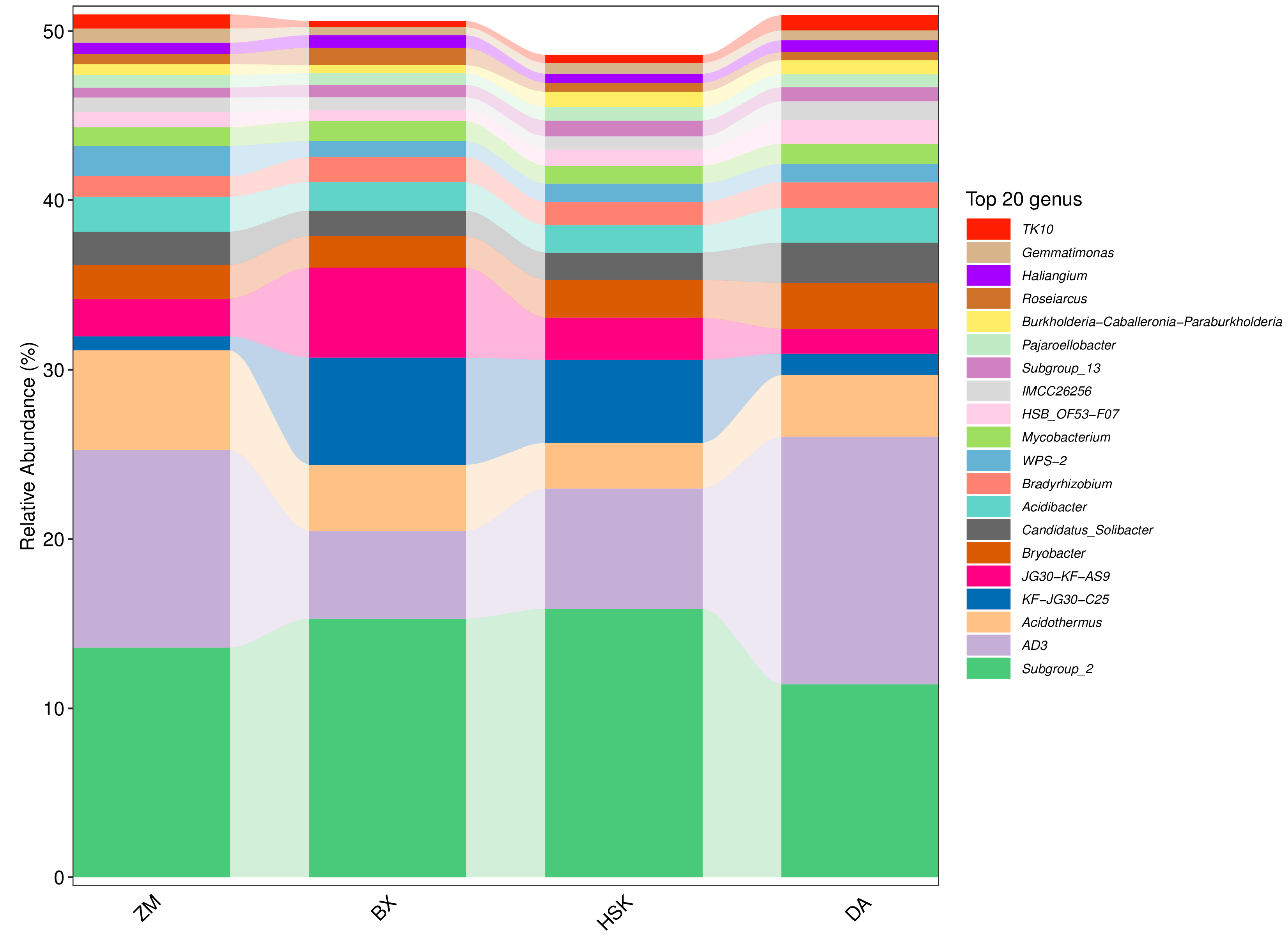
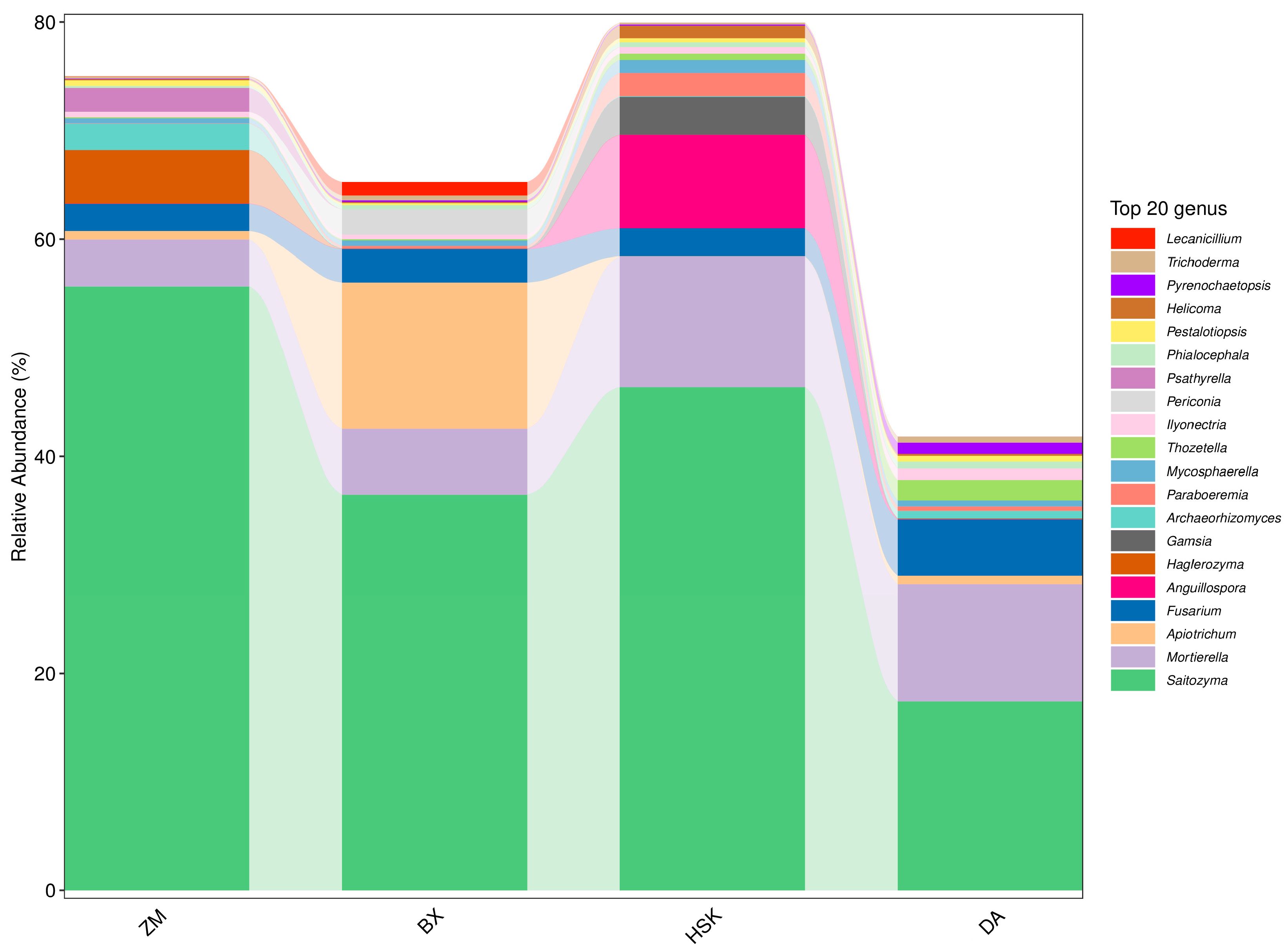
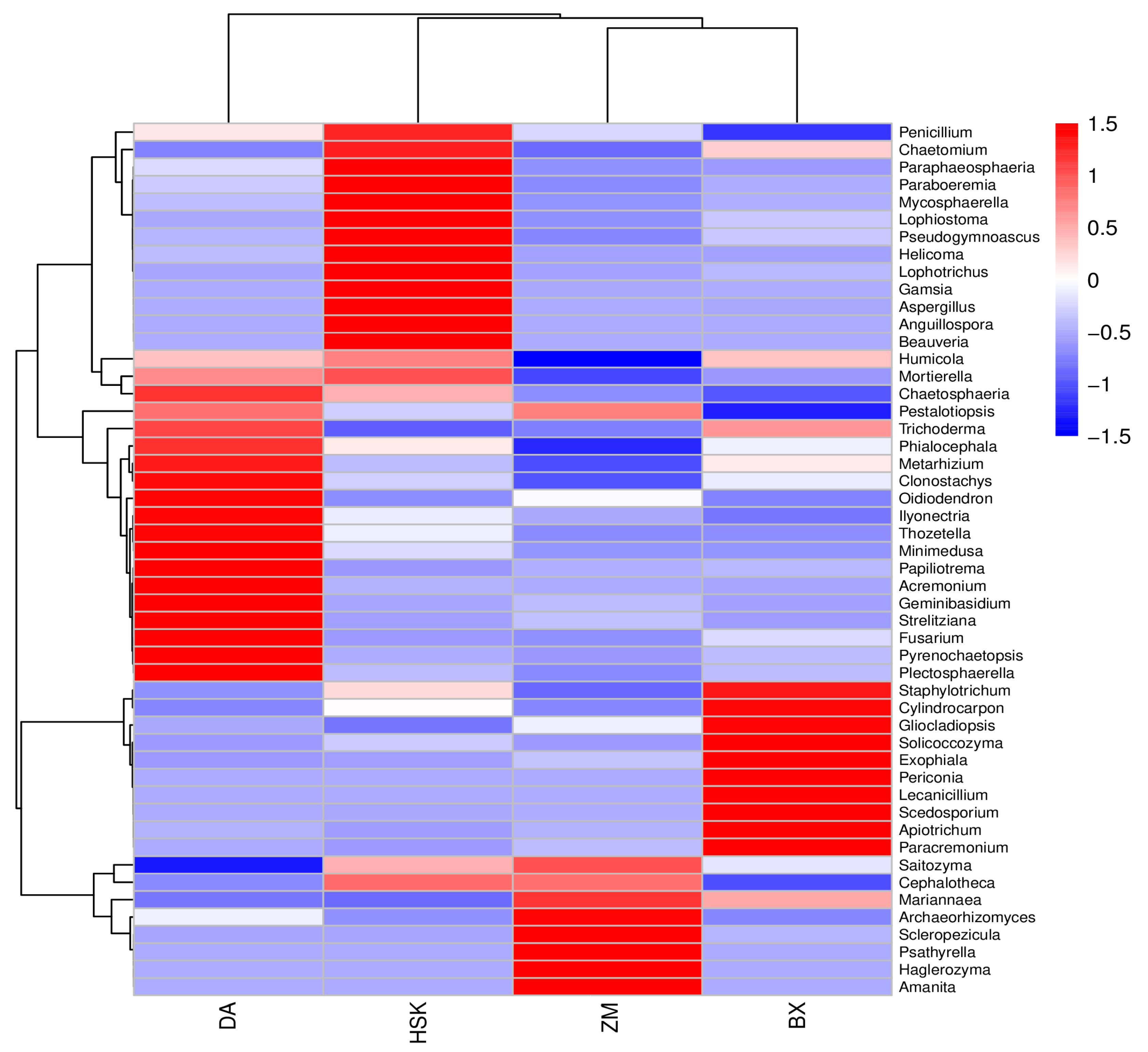
3.2.5. Structural Composition of Soil Bacteria and Fungi at the Genus Level
3.3. Correlation Analysis
3.3.1. Correlation Between Soil Fertility Indicators and Rhizosphere Microbial Diversity Metrics
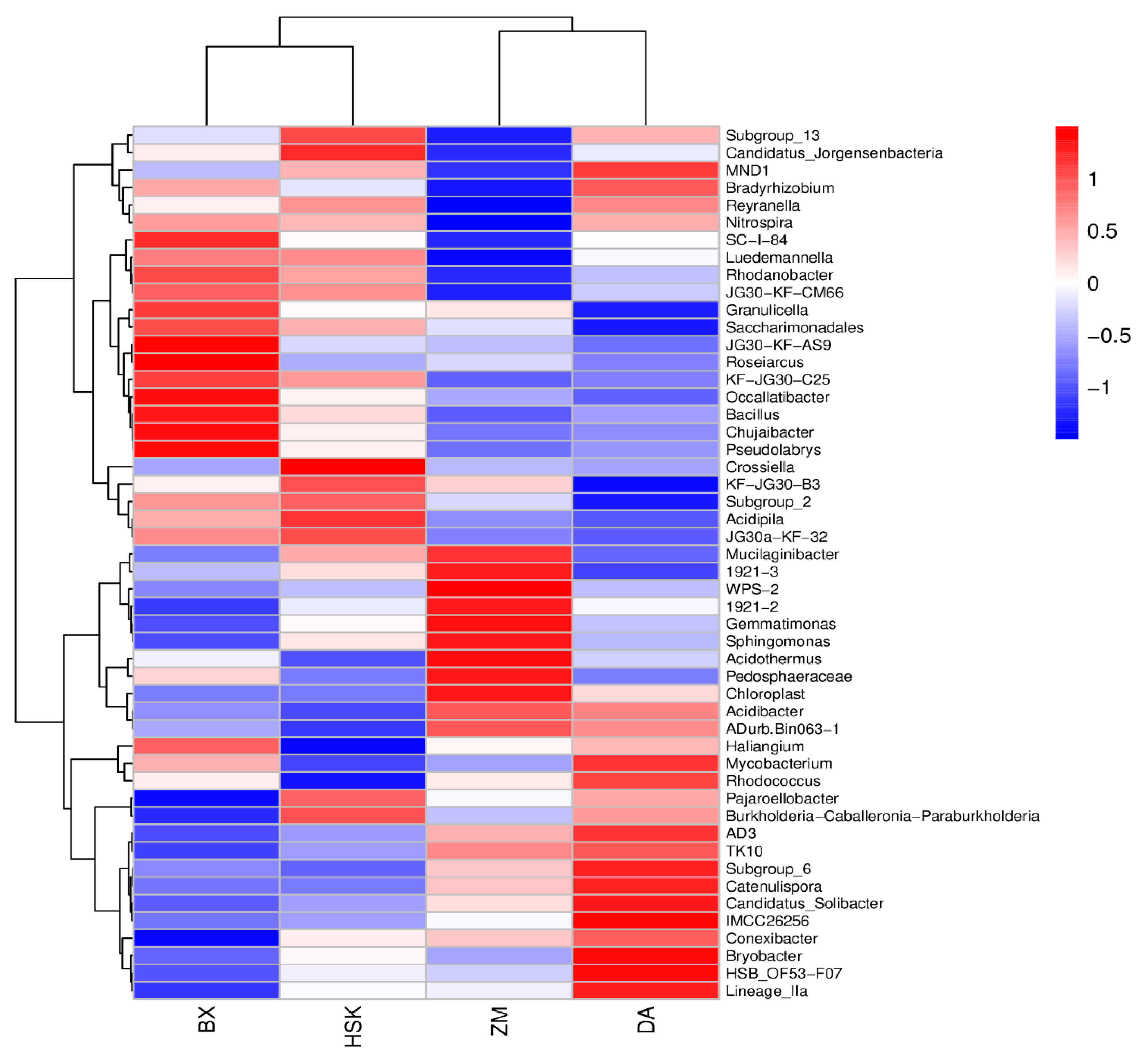
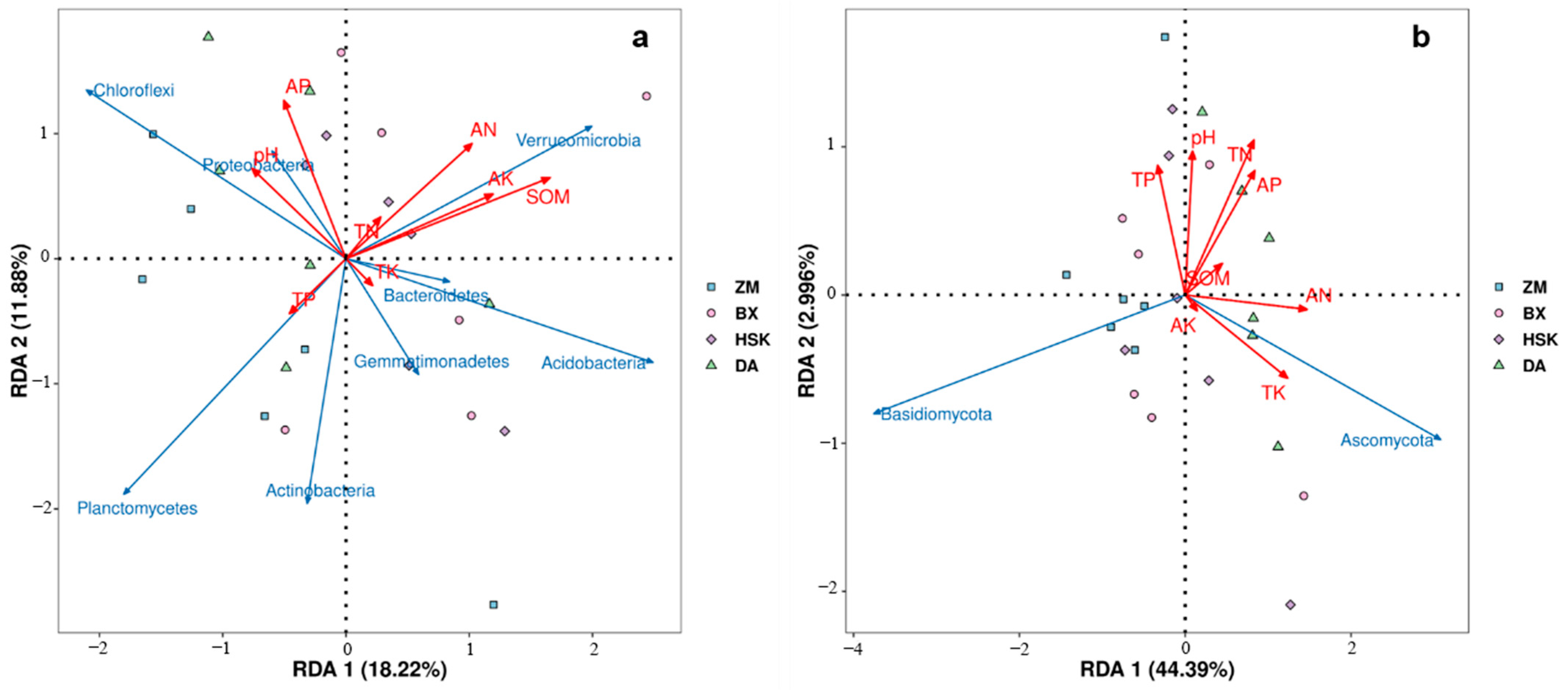
3.3.2. Redundancy Analysis of Rhizosphere Soil Microorganisms and Soil Fertility Indicators
4. Discussion
4.1. Soil pH in Dancong Ancient Tea Plantations
4.2. Soil Nutrients of Dancong Ancient Tea Plantations
4.3. Diversity of Soil Microbial Communities in Ancient Tea Plantations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tan, L.; Gu, S.; Li, S.; Ren, Z.; Deng, Y.; Liu, Z.; Gong, Z.; Xiao, W.; Hu, Q. Responses of Microbial Communities and Interaction Networks to Different Management Practices in Tea Plantation Soils. Sustainability 2019, 11, 4428. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, W.; Cheng, X.; Wang, Z.; Yuan, F.; Wu, W.; Liao, S. Evaluating the Productivity of Ancient Pu’er Tea Trees (Camellia sinensis Var. Assamica): A Multivariate Modeling Approach. Plant Methods 2022, 18, 95. [Google Scholar] [CrossRef] [PubMed]
- Jahan, I.; Shopan, J.; Rahman, M.M.; Sarkar, A.; Baset, M.A.; Zhang, Z.; Li, X.; Ahammed, G.J.; Hasan, M.K. Long-Term Traditional Fertilization Alters Tea Garden Soil Properties and Tea Leaf Quality in Bangladesh. Agronomy 2022, 12, 2128. [Google Scholar] [CrossRef]
- Ran, W.; Li, Q.; Hu, X.; Zhang, D.; Yu, Z.; Chen, Y.; Wang, M.; Ni, D. Comprehensive Analysis of Environmental Factors on the Quality of Tea (Camellia sinensis Var. Sinensis) Fresh Leaves. Sci. Hortic. 2023, 319, 112177. [Google Scholar] [CrossRef]
- Xin, W.; Zhang, J.; Yu, Y.; Tian, Y.; Li, H.; Chen, X.; Li, W.; Liu, Y.; Lu, T.; He, B.; et al. Root Microbiota of Tea Plants Regulate Nitrogen Homeostasis and Theanine Synthesis to Influence Tea Quality. Curr. Biol. 2024, 34, 868–880.e6. [Google Scholar] [CrossRef]
- Chen, Y.; Fu, W.; Xiao, H.; Zhai, Y.; Luo, Y.; Wang, Y.; Liu, Z.; Li, Q.; Huang, J. A Review on Rhizosphere Microbiota of Tea Plant (Camellia sinensis L): Recent Insights and Future Perspectives. J. Agric. Food Chem. 2023, 71, 19165–19188. [Google Scholar] [CrossRef]
- Shang, J.; Sheng, Z.; Deng, M. A Microbial Consortium Enhances Tea Yield and Quality in a Field Study. Rhizosphere 2023, 26, 100691. [Google Scholar] [CrossRef]
- Wang, T.; Duan, Y.; Lei, X.; Cao, Y.; Liu, L.; Shang, X.; Wang, M.; Lv, C.; Ma, Y.; Fang, W.; et al. Tea Plantation Intercropping Legume Improves Soil Ecosystem Multifunctionality and Tea Quality by Regulating Rare Bacterial Taxa. Agronomy 2023, 13, 1110. [Google Scholar] [CrossRef]
- Zhang, Z.; Ge, S.; Fan, L.-C.; Guo, S.; Hu, Q.; Ahammed, G.J.; Yan, P.; Zhang, L.-P.; Li, Z.-Z.; Zhang, J.-Y.; et al. Diversity in Rhizospheric Microbial Communities in Tea Varieties at Different Locations and Tapping Potential Beneficial Microorganisms. Front. Microbiol. 2022, 13, 1027444. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, R.; Peng, W.; Yang, Y.; Ma, X.; Zhang, W.; Ji, A.; Liu, L.; Liu, P.; Yan, L.; et al. Tea Plants with Gray Blight Have Altered Root Exudates That Recruit a Beneficial Rhizosphere Microbiome to Prime Immunity against Aboveground Pathogen Infection. Front. Microbiol. 2021, 12, 774438. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, F.; Wu, Z.; Jiang, F.; Yu, W.; Yang, J.; Chen, J.; Jian, G.; You, Z.; Zeng, L. Effects of Long-Term Nitrogen Fertilization on the Formation of Metabolites Related to Tea Quality in Subtropical China. Metabolites 2021, 11, 146. [Google Scholar] [CrossRef]
- Yan, P.; Wu, L.; Wang, D.; Fu, J.; Shen, C.; Li, X.; Zhang, L.; Zhang, L.; Fan, L.; Wenyan, H. Soil Acidification in Chinese Tea Plantations. Sci. Total Environ. 2020, 715, 136963. [Google Scholar] [CrossRef]
- Li, Y.C.; Li, Z.; Li, Z.W.; Jiang, Y.H.; Weng, B.Q.; Lin, W.X. Variations of Rhizosphere Bacterial Communities in Tea (Camellia sinensis L.) Continuous Cropping Soil by High-Throughput Pyrosequencing Approach. J. Appl. Microbiol. 2016, 121, 787–799. [Google Scholar] [CrossRef]
- Arafat, Y.; Ud Din, I.; Tayyab, M.; Jiang, Y.; Chen, T.; Cai, Z.; Zhao, H.; Lin, X.; Lin, W.; Lin, S. Soil Sickness in Aged Tea Plantation Is Associated with a Shift in Microbial Communities as a Result of Plant Polyphenol Accumulation in the Tea Gardens. Front. Plant Sci. 2020, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Kui, L.; Xiang, G.; Wang, Y.; Wang, Z.; Li, G.; Li, D.; Yan, J.; Ye, S.; Wang, C.; Yang, L.; et al. Large-Scale Characterization of the Soil Microbiome in Ancient Tea Plantations Using High-Throughput 16S rRNA and Internal Transcribed Spacer Amplicon Sequencing. Front. Microbiol. 2021, 12, 745225. [Google Scholar] [CrossRef] [PubMed]
- Kaiyang, L. Bacteria Distribution Diversity in Tea Rhizospheric Soil from Different Habitats at Nannuo Mountain Based on 16S rRNA Sequence Analysis. J. Kunming Univ. Sci. Technol. 2016, 41, 89–95. [Google Scholar]
- Cao, H.-L.; Cai, F.-Y.; Jiao, W.-B.; Liu, C.; Zhang, N.; Qiu, H.-Y.; Rensing, C.; Lü, J. Assessment of Tea Garden Soils at an’xi County in Southeast China Reveals a Mild Threat from Contamination of Potentially Harmful Elements. R. Soc. Open Sci. 2018, 5, 180050. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, N.-M.; Deng, H. Soil Nutrient Status and Fertility Evaluation of Tea Gardens in Menghai County. J. Agric. Resour. Environ. 2021, 38, 79–86. [Google Scholar]
- Ma, W.; Zhu, K.; Zhuo, Z. Effects of Acidification on Mineral Transformation and Potassium Supply Capacity of Tea Garden Soils. J. Tea Sci. 2023, 43, 17–26. [Google Scholar]
- Wang, Y.; Zhang, Q.; Li, J.; Lin, S.; Jia, X.; Zhang, Q.; Ye, J.; Wang, H.; Wu, Z. Study on the Effect of pH on Rhizosphere Soil Fertility and the Aroma Quality of Tea Trees and Their Interactions. Agriculture 2023, 13, 1739. [Google Scholar] [CrossRef]
- Yan, P.; Han, W.; Li, X.; Zhang, L.; Zhang, L. Present Situation and Analysis of Soil Acidification in Chinese Tea Garden. Sci. Agric. Sin. 2020, 53, 795–801. [Google Scholar]
- Ruan, L.; Li, X.; Song, Y.; Li, J.; Palansooriya, K.N. Effects of Tea Plant Varieties with High- and Low-Nutrient Efficiency on Nutrients in Degraded Soil. Plants 2023, 12, 905. [Google Scholar] [CrossRef]
- Tang, S.; Liu, Y.; Zheng, N.; Li, Y.; Ma, Q.; Xiao, H.; Zhou, X.; Xu, X.; Jiang, T.; He, P.; et al. Temporal Variation in Nutrient Requirements of Tea (Camellia sinensis) in China Based on QUEFTS Analysis. Sci. Rep. 2020, 10, 1745. [Google Scholar] [CrossRef] [PubMed]
- Su, N.I.U.; Qin, S.; Wei, F.A.N. Analysis on Site Soil Nutrients of Ancient Tea Trees in the Southwest State of Guizhou Province. Acta Agric. Zhejiangensis 2013, 25, 1348–1353. [Google Scholar]
- Yang, G.; Zhou, D.; Wan, R.; Wang, C.; Xie, J.; Ma, C.; Li, Y. HPLC and High-Throughput Sequencing Revealed Higher Tea-Leaves Quality, Soil Fertility and Microbial Community Diversity in Ancient Tea Plantations: Compared with Modern Tea Plantations. BMC Plant Biol. 2022, 22, 239. [Google Scholar] [CrossRef]
- Yang, G.R.; Wang, X.Q.; Li, Y.M.; Xie, J.; Lv, C.Y. Study on Soil Nutrients and Tea Quality of Ancient Tea Arboretum and Modern Terrace Tea Garden in Jingmai Mountain. J. Yunnan Agric. Univ. (Nat. Sci.) 2016, 31, 519–527. [Google Scholar]
- Yang, X.; Huang, X.; Hu, X.; Cheng, X.; Luo, Y. Changes in Rhizosphere and Bulk Soil Microbial Communities of Tableland Tea Garden and Ancient Tea Plantation in Southwest China. Agronomy 2024, 14, 1388. [Google Scholar] [CrossRef]
- Xu, L.; Wang, G.; Zhang, S.; Li, T.; Xu, X.; Gong, G.; Zhou, W.; Pu, Y.; Jia, Y.; Li, Y.; et al. Inhibition of High Sulfur on Functional Microorganisms and Genes in Slightly Contaminated Soil by Cadmium and Chromium. Environ. Pollut. 2024, 344, 123421. [Google Scholar] [CrossRef]
- Wang, S.; Li, T.; Zheng, Z.; Chen, H.Y.H. Soil Aggregate-Associated Bacterial Metabolic Activity and Community Structure in Different Aged Tea Plantations. Sci. Total Environ. 2019, 654, 1023–1032. [Google Scholar] [CrossRef]
- Yang, Y.; Wu, H.; Wang, Sh. Comparison of Microbial Communities in the Rhizosphere Soils from Two Varieties of Camellia sinensis in Yunnan Province, China. Microbiology 2021, 90, 801–804. [Google Scholar] [CrossRef]
- Yergeau, E.; Bokhorst, S.; Kang, S.; Zhou, J.; Greer, C.W.; Aerts, R.; Kowalchuk, G.A. Shifts in Soil Microorganisms in Response to Warming Are Consistent across a Range of Antarctic Environments. ISME J. 2012, 6, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Niu, Z.; Chen, S.; Zhao, S.; Qiu, Z.; Wang, Y.; Hua, X.; Ding, Z.; Ma, Q. Effects of Pea-Tea Intercropping on Rhizosphere Soil Microbial Communities. Plant Soil 2025, 506, 125–135. [Google Scholar] [CrossRef]
- Dai, Z.; Su, W.; Chen, H.; Barberán, A.; Zhao, H.; Yu, M.; Yu, L.; Brookes, P.C.; Schadt, C.W.; Chang, S.X.; et al. Long-term Nitrogen Fertilization Decreases Bacterial Diversity and Favors the Growth of Actinobacteria and Proteobacteria in Agro-ecosystems across the Globe. Glob. Change Biol. 2018, 24, 3452–3461. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, Y.; Wu, Z.; You, Z.; Yu, W.; Yu, X.; Yang, Z. Effects of Organic Management Mode on Soil Fungal Community Structure and Functions in Tea Gardens. J. Tea Sci. 2022, 42, 672–688. [Google Scholar]
- Lim, Y.W.; Kim, B.K.; Kim, C.; Jung, H.S.; Kim, B.-S.; Lee, J.-H.; Chun, J. Assessment of Soil Fungal Communities Using Pyrosequencing. J. Microbiol. 2010, 48, 284–289. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, W.; Zhou, B.; Yang, J.; Xing, S. Effect of Biochar-Based Fertilizer on Soil Bacteria and Fungi Quantity and Community Structure in Acidified Tea Garden. J. Fujian Agric. For. Univ. 2023, 52, 247–257. Available online: https://kns.cnki.net/kcms2/article/abstract?v=UThtwiquHbf-e5kACKb-sFO5LYXiGprNU7x_D5W645hheug2KbWutw5N-n2rLVXJ9R6mjNhTb9tCOvD9-tZq8Y0qK5rXAxRR3G4subIULWaMP1lcJf7UL6CyJbWcKitEUxyitL3zcIZcw4ZpwBVTTM8XnjUEDt-GbfvIJGp9XZ6eOwwo0RD2Fw==&uniplatform=NZKPT&language=CHS (accessed on 27 June 2025).
- Fu, H.; Li, H.; Yin, P.; Mei, H.; Li, J.; Zhou, P.; Wang, Y.; Ma, Q.; Jeyaraj, A.; Thangaraj, K.; et al. Integrated Application of Rapeseed Cake and Green Manure Enhances Soil Nutrients and Microbial Communities in Tea Garden Soil. Sustainability 2021, 13, 2967. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, J.; Ye, J.; Ye, Y.; Lin, Y.; Liu, C. Effects of Biochar Application on Soil Properties and Fungi Community Structure in Acidified Tea Gardens. J. Tea Sci. 2021, 41, 419–429. [Google Scholar]
- Fu, H.P.; Zhou, P.Q.; Wang, Y.J.; Mo, Z.D.; Li, Z.H.; Ma, Q.P.; Hoang, H.N.; Li, X.H. Effects of Intercropping Different Green Manures on Fungal Community Characteristics in Rhizosphere Soil of Tea Plant. J. Tea Commun. 2020, 47, 406–415. [Google Scholar]
- Hu, Z.; Ji, L.; Wan, Q.; Li, H.; Li, R.; Yang, Y. Short-Term Effects of Bio-Organic Fertilizer on Soil Fertility and Bacterial Community Composition in Tea Plantation Soils. Agronomy 2022, 12, 2168. [Google Scholar] [CrossRef]
- Bhattacharyya, C.; Imchen, M.; Mukherjee, T.; Haldar, S.; Mondal, S.; Mukherji, S.; Haldar, A.; Kumavath, R.; Ghosh, A. Rhizosphere Impacts Bacterial Community Structure in the Tea (Camellia sinensis (L.) O. Kuntze.) Estates of Darjeeling, India. Environ. Microbiol. 2022, 24, 2716–2731. [Google Scholar] [CrossRef]
- Gui, H.; Fan, L.; Wang, D.; Yan, P.; Li, X.; Pang, Y.; Zhang, L.; Zamanian, K.; Shi, L.; Xu, J.; et al. Variations in Soil Nutrient Dynamics and Bacterial Communities after the Conversion of Forests to Long-Term Tea Monoculture Systems. Front. Microbiol. 2022, 13, 896530. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Thomson, B.C.; James, P.; Bell, T.; Bailey, M.; Whiteley, A.S. The Bacterial Biogeography of British Soils. Environ. Microbiol. 2011, 13, 1642–1654. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Xiu, W.; Tan, B.; Li, G.; Zhao, J.; Yang, D.; Zhang, G.; Zhang, Y. Responses of Soil Microbial and Nematode Communities to Various Cover Crop Patterns in a Tea Garden of China. Int. J. Environ. Res. Public Health 2022, 19, 2695. [Google Scholar] [CrossRef]
- Wang, H.B.; Chen, X.T.; Ding, L.; Qiu, F.; Ye, J.; Jia, X.; Kong, X.; He, H. Effect of Soil Acidity on Microbial Diversity in Rhizospheric Soils of Tea Plants. Chin. J. Trop. Crops 2018, 39, 448–454. [Google Scholar]
- Kong, C.; Li, Y.; Sun, Y.; Shang, W.; Liu, B.; Yang, T. Variance of Nitrogen, Phosphorus and Potassium Content in New Shoots of Different Tea Tree Varieties and Diversity of Rhizosphere Soil Bacterial Communities. Southwest China J. Agric. Sci. 2023, 12, 2763–2770. [Google Scholar]
- Arafat, Y.; Tayyab, M.; Khan, M.U.; Chen, T.; Amjad, H.; Awais, S.; Lin, X.; Lin, W.; Lin, S. Long-Term Monoculture Negatively Regulates Fungal Community Composition and Abundance of Tea Orchards. Agronomy 2019, 9, 466. [Google Scholar] [CrossRef]
- Yan, P.; Shen, C.; Zou, Z.; Fan, L.; Li, X.; Zhang, L.; Zhang, L.; Dong, C.; Fu, J.; Han, W.; et al. Increased Soil Fertility in Tea Gardens Leads to Declines in Fungal Diversity and Complexity in Subsoils. Agronomy 2022, 12, 1751. [Google Scholar] [CrossRef]
- Jia, M.; Wang, Y.; Zhang, Q.; Lin, S.; Zhang, Q.; Chen, Y.; Hong, L.; Jia, X.; Ye, J.; Wang, H. Effect of Soil pH on the Uptake of Essential Elements by Tea Plant and Subsequent Impact on Growth and Leaf Quality. Agronomy 2024, 14, 1338. [Google Scholar] [CrossRef]
- Ye, J.; Wang, Y.; Wang, Y.; Hong, L.; Jia, X.; Kang, J.; Lin, S.; Wu, Z.; Wang, H. Improvement of Soil Acidification in Tea Plantations by Long-Term Use of Organic Fertilizers and Its Effect on Tea Yield and Quality. Front. Plant Sci. 2022, 13, 1055900. [Google Scholar] [CrossRef]
- Jia, X.; Zhang, Q.; Wang, Y.; Zhang, Y.; Li, M.; Cheng, P.; Chen, M.; Lin, S.; Zou, J.; Ye, J.; et al. Changes of Physiological Characteristics, Element Accumulation and Hormone Metabolism of Tea Leaves in Response to Soil pH. Front. Plant Sci. 2023, 14, 1266026. [Google Scholar] [CrossRef]
- Song, M. Effect of Biogeochemical Cycle in Tea Garden on the Soil Acidification. J. Tea Sci. 1990, 10, 19–26. [Google Scholar]
- Dang, M.V. Soil–Plant Nutrient Balance of Tea Crops in the Northern Mountainous Region, Vietnam. Agric. Ecosyst. Environ. 2005, 105, 413–418. [Google Scholar] [CrossRef]
- Zhang, W.; Ni, K.; Long, L.; Ruan, J. Nitrogen Transport and Assimilation in Tea Plant (Camellia sinensis): A Review. Front. Plant Sci. 2023, 14, 1249202. [Google Scholar] [CrossRef]
- Liu, X.; Tian, J.; Liu, G.; Sun, L. Multi-Omics Analysis Reveals Mechanisms of Strong Phosphorus Adaptation in Tea Plant Roots. Int. J. Mol. Sci. 2023, 24, 12431. [Google Scholar] [CrossRef]
- Huang, W.; Lin, M.; Liao, J.; Li, A.; Tsewang, W.; Chen, X.; Sun, B.; Liu, S.; Zheng, P. Effects of Potassium Deficiency on the Growth of Tea (Camelia sinensis) and Strategies for Optimizing Potassium Levels in Soil: A Critical Review. Horticulturae 2022, 8, 660. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, X.; Nie, C.; Wu, T.; Dai, W.; Liu, H.; Yang, R. Analysis of Unculturable Bacterial Communities in Tea Orchard Soils Based on Nested PCR-DGGE. World J. Microbiol. Biotechnol. 2012, 28, 1967–1979. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Zhou, J.; Pan, W.; Tang, R.; Ma, Q.; Xu, M.; Qi, T.; Ma, Z.; Fu, H.; Wu, L. Impact of N Application Rate on Tea (Camellia sinensis) Growth and Soil Bacterial and Fungi Communities. Plant Soil 2022, 475, 343–359. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent Understanding of Soil Acidobacteria and Their Ecological Significance: A Critical Review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Zheng, N.; Yu, Y.; Wang, J.; Chapman, S.J.; Yao, H.; Zhang, Y. The Conversion of Subtropical Forest to Tea Plantation Changes the Fungal Community and the Contribution of Fungi to N2O Production. Environ. Pollut. 2020, 265, 115106. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality Name | Site ID | Longitude (°E) | Latitude (°N) | Altitude (m) |
|---|---|---|---|---|
| Zimao | ZM | 116.68 | 23.95 | 578.13 |
| Baixiang | BX | 116.65 | 23.96 | 1131.63 |
| Xialiao | HSK | 116.65 | 23.96 | 1066.77 |
| Daan | DA | 116.64 | 23.97 | 965.91 |
| ZM | BX | HSK | DA | |
|---|---|---|---|---|
| pH | 4.91 ± 0.00 a | 4.89 ± 0.11 a | 5.03 ± 0.27 a | 5.01 ± 0.08 a |
| TN/(g/kg) | 1.47 ± 0.31 a | 1.82 ± 0.10 a | 1.74 ± 0.12 a | 1.75 ± 0.22 a |
| TP/(g/kg) | 0.89 ± 0.10 a | 0.83 ± 0.03 a | 0.97 ± 0.09 a | 0.84 ± 0.03 a |
| TK/(g/kg) | 9.67 ± 0.88 c | 10.50 ± 0.76 bc | 15.00 ± 0.76 a | 12.83 ± 0.33 ab |
| SOM/(g/kg) | 24.97 ± 1.67 b | 36.51 ± 1.57 a | 36.51 ± 2.32 a | 30.20 ± 0.97 b |
| AN/(mg/kg) | 89.95 ± 2.94 b | 139.30 ± 4.13 a | 170.68 ± 17.57 a | 149.22 ± 2.94 a |
| AP/(mg/kg) | 36.52 ± 11.58 ab | 54.32 ± 28.00 ab | 21.22 ± 7.54 b | 82.66 ± 9.23 a |
| AK/(mg/kg) | 46.67 ± 7.69 a | 79.33 ± 20.00 a | 47.00 ± 9.29 a | 62.00 ± 5.29 a |
| ZM | BX | HSK | DA | |
|---|---|---|---|---|
| TN | 1.56 | 1.95 | 2.10 | 2.19 |
| TP | 1.49 | 1.33 | 1.46 | 1.39 |
| TK | 1.12 | 0.93 | 1.42 | 1.27 |
| SOM | 1.93 | 2.70 | 2.46 | 2.51 |
| AN | 1.04 | 1.47 | 1.88 | 1.48 |
| AP | 3.80 | 2.54 | 2.92 | 6.51 |
| AK | 0.58 | 0.95 | 0.46 | 0.57 |
| ZM | BX | HSK | DA | |
|---|---|---|---|---|
| pH | 4.91 ± 0.00 a | 4.89 ± 0.11 a | 5.03 ± 0.27 a | 5.01 ± 0.08 a |
| TN/(g/kg) | 1.47 ± 0.31 a | 1.82 ± 0.10 a | 1.74 ± 0.12 a | 1.75 ± 0.22 a |
| TP/(g/kg) | 0.89 ± 0.10 a | 0.83 ± 0.03 a | 0.97 ± 0.09 a | 0.84 ± 0.03 a |
| TK/(g/kg) | 9.67 ± 0.88 c | 10.50 ± 0.76 bc | 15.00 ± 0.76 a | 12.83 ± 0.33 ab |
| SOM/(g/kg) | 24.97 ± 1.67 b | 36.51 ± 1.57 a | 36.51 ± 2.32 a | 30.20 ± 0.97 b |
| AN/(mg/kg) | 89.95 ± 2.94 b | 139.30 ± 4.13 a | 170.68 ± 17.57 a | 149.22 ± 2.94 a |
| AP/(mg/kg) | 36.52 ± 11.58 ab | 54.32 ± 28.00 ab | 21.22 ± 7.54 b | 82.66 ± 9.23 a |
| AK/(mg/kg) | 46.67 ± 7.69 a | 79.33 ± 20.00 a | 47.00 ± 9.29 a | 62.00 ± 5.29 a |
| ZM | BX | HSK | DA | |
|---|---|---|---|---|
| TN | 1.47 | 1.82 | 1.74 | 1.75 |
| TP | 1.48 | 1.38 | 1.61 | 1.40 |
| TK | 0.97 | 1.05 | 1.50 | 1.28 |
| SOM | 1.66 | 2.43 | 2.43 | 2.01 |
| AN | 0.30 | 1.39 | 1.71 | 1.49 |
| AP | 3.65 | 5.43 | 2.12 | 8.27 |
| AK | 0.39 | 0.66 | 0.39 | 0.52 |
| Chao1 | Simpson | Shannon | Pielou | Observed Species | Faith pd | Goods Coverage | |
|---|---|---|---|---|---|---|---|
| ZM | 3386.95 ± 127.39 | 1.00 ± 0.00 | 9.92 ± 0.13 | 0.86 ± 0.01 | 3067.57 ± 123.07 | 212.46 ± 6.27 | 0.98 ± 0.00 |
| BX | 3594.65 ± 85.14 | 1.00 ± 0.00 | 10.05 ± 0.09 | 0.86 ± 0.01 | 3254.68 ± 64.19 | 231.85 ± 6.86 | 0.98 ± 0.00 |
| HSK | 3566.98 ± 173.18 | 1.00 ± 0.00 | 10.16 ± 0.14 | 0.87 ± 0.01 | 3272.92 ± 148.59 | 227.85 ± 8.48 | 0.98 ± 0.00 |
| DA | 3483.27 ± 145.6 | 1.00 ± 0.00 | 10.24 ± 0.11 | 0.88 ± 0.01 | 3135.93 ± 126.3 | 213.49 ± 10.54 | 0.98 ± 0.00 |
| Chao1 | Simpson | Shannon | Pielou | Observed Species | Faith pd | Goods Coverage | |
|---|---|---|---|---|---|---|---|
| ZM | 353.02 ± 62.95 | 0.66 ± 0.09 | 3.47 ± 0.61 | 0.41 ± 0.06 | 352.18 ± 62.84 | 1.00 ± 0.00 | 0.98 ± 0.00 |
| BX | 456.79 ± 49.27 | 0.83 ± 0.04 | 4.64 ± 0.35 | 0.53 ± 0.03 | 456.23 ± 49.34 | 1.00 ± 0.00 | 0.98 ± 0.00 |
| HSK | 394.58 ± 32.75 | 0.82 ± 0.03 | 4.31 ± 0.19 | 0.50 ± 0.02 | 393.58 ± 32.72 | 1.00 ± 0.00 | 0.98 ± 0.00 |
| DA | 523.55 ± 37.27 | 0.93 ± 0.01 | 5.53 ± 0.24 | 0.61 ± 0.02 | 522.9 ± 37.36 | 1.00 ± 0.00 | 0.98 ± 0.00 |
| Chao1 | Simpson | Shannon | Pielou | Observed Species | Goods Coverage | |
|---|---|---|---|---|---|---|
| pH | 0.13 | 0.00 | 0.06 | 0.04 | 0.13 | −0.06 |
| AN | 0.16 | 0.47 * | 0.37 | 0.41 * | 0.16 | −0.37 |
| AP | 0.38 | 0.13 | 0.33 | 0.30 | 0.38 | 0.06 |
| AK | 0.21 | 0.19 | 0.22 | 0.21 | 0.21 | −0.02 |
| SOM | 0.31 | 0.52 * | 0.43 * | 0.45 * | 0.31 | −0.09 |
| TN | 0.26 | 0.52 * | 0.41 * | 0.44 * | 0.26 | −0.03 |
| TP | 0.02 | 0.16 | 0.08 | 0.10 | 0.02 | 0.00 |
| TK | 0.08 | 0.34 | 0.21 | 0.24 | 0.08 | −0.55 * |
| Chao1 | Simpson | Shannon | Pielou | Observed Species | Goods Coverage | |
|---|---|---|---|---|---|---|
| Ph | 0.13 | 0.00 | 0.06 | 0.04 | 0.13 | −0.06 |
| AN | 0.16 | 0.47 * | 0.37 | 0.41 * | 0.16 | −0.37 |
| AP | 0.38 | 0.13 | 0.33 | 0.30 | 0.38 | 0.06 |
| AK | 0.21 | 0.19 | 0.22 | 0.21 | 0.21 | −0.02 |
| SOM | 0.31 | 0.52 * | 0.43 * | 0.45 * | 0.31 | −0.09 |
| TN | 0.26 | 0.52 * | 0.41 * | 0.44 * | 0.26 | −0.03 |
| TP | 0.02 | 0.16 | 0.08 | 0.10 | 0.02 | 0.00 |
| TK | 0.08 | 0.34 | 0.21 | 0.24 | 0.08 | −0.55 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Huang, W.; Lin, X.; Khan, W.; Zhao, H.; Sun, B.; Liu, S.; Zheng, P. Study on Soil Nutrients and Microbial Community Diversity in Ancient Tea Plantations of China. Agronomy 2025, 15, 1608. https://doi.org/10.3390/agronomy15071608
Li J, Huang W, Lin X, Khan W, Zhao H, Sun B, Liu S, Zheng P. Study on Soil Nutrients and Microbial Community Diversity in Ancient Tea Plantations of China. Agronomy. 2025; 15(7):1608. https://doi.org/10.3390/agronomy15071608
Chicago/Turabian StyleLi, Jiaxin, Wei Huang, Xinyuan Lin, Waqar Khan, Hongbo Zhao, Binmei Sun, Shaoqun Liu, and Peng Zheng. 2025. "Study on Soil Nutrients and Microbial Community Diversity in Ancient Tea Plantations of China" Agronomy 15, no. 7: 1608. https://doi.org/10.3390/agronomy15071608
APA StyleLi, J., Huang, W., Lin, X., Khan, W., Zhao, H., Sun, B., Liu, S., & Zheng, P. (2025). Study on Soil Nutrients and Microbial Community Diversity in Ancient Tea Plantations of China. Agronomy, 15(7), 1608. https://doi.org/10.3390/agronomy15071608