
Carob–Thyme Intercropping Systems Can Improve Yield Efficiency and Environmental Footprint Compared to Conservation Tillage



Abstract
1. Introduction
2. Materials and Methods
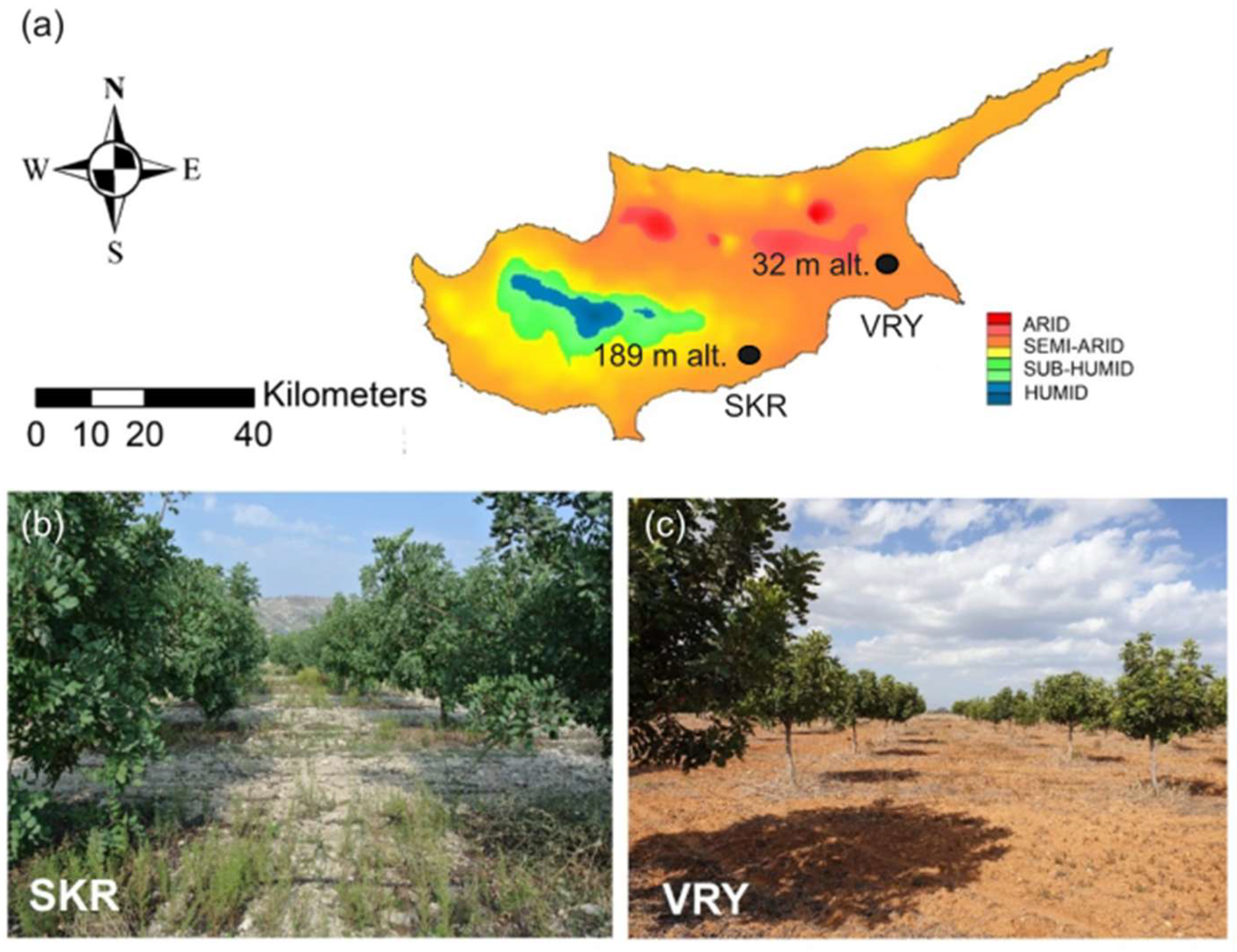
2.1. Experimental Sites and Tree Selection
2.2. Intercropping Experimental Design
2.2.1. Carob–Thyme Intercropping System (T-System)
2.2.2. Carob–Clover Intercropping System (C-System)
2.3. Control Plots
2.3.1. Conservation Tillage Control Plots Without Irrigation (TLGdry)
2.3.2. Irrigated Conservation Tillage Carob Trees (TLGirr)
2.4. Experimental Monitoring
2.4.1. Thyme Survival Rate
2.4.2. Thyme Biomass/Thyme Soil Cover/Total Soil Cover
2.4.3. Clover Biomass and Soil Cover
2.4.4. Water Potential
2.4.5. Wood Production
2.4.6. Carob Yield Efficiency
2.4.7. Soil Moisture Measurements
2.4.8. Surface Temperature Measurements
2.5. Climate Data
2.6. Environmental Indicators
2.7. Statistical Analysis
3. Results
3.1. Climatic Conditions
3.2. Establishment and Performance of T- and C-Systems
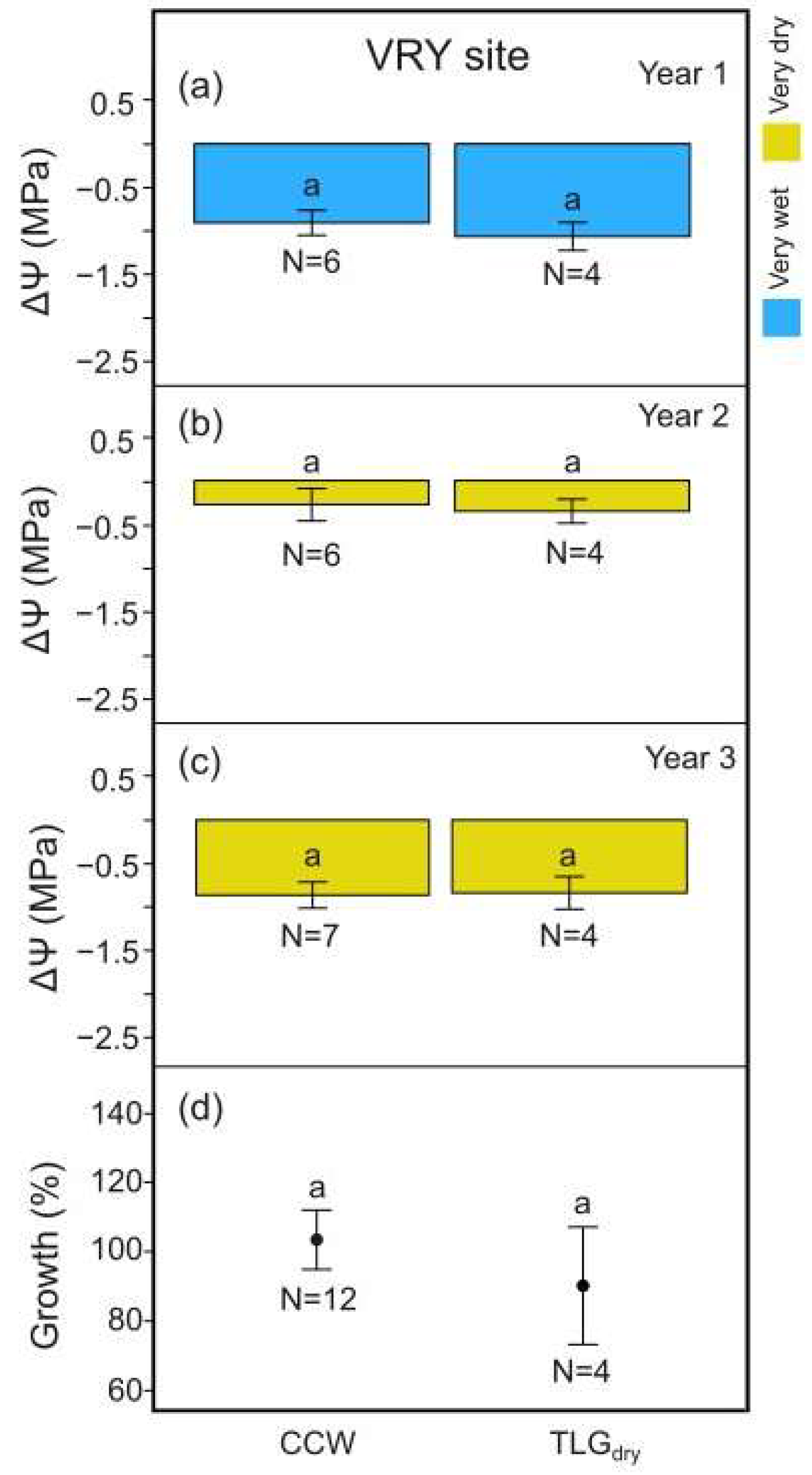
3.3. Effects on Yield Efficiency, Leaf Water Potential, and Tree Growth
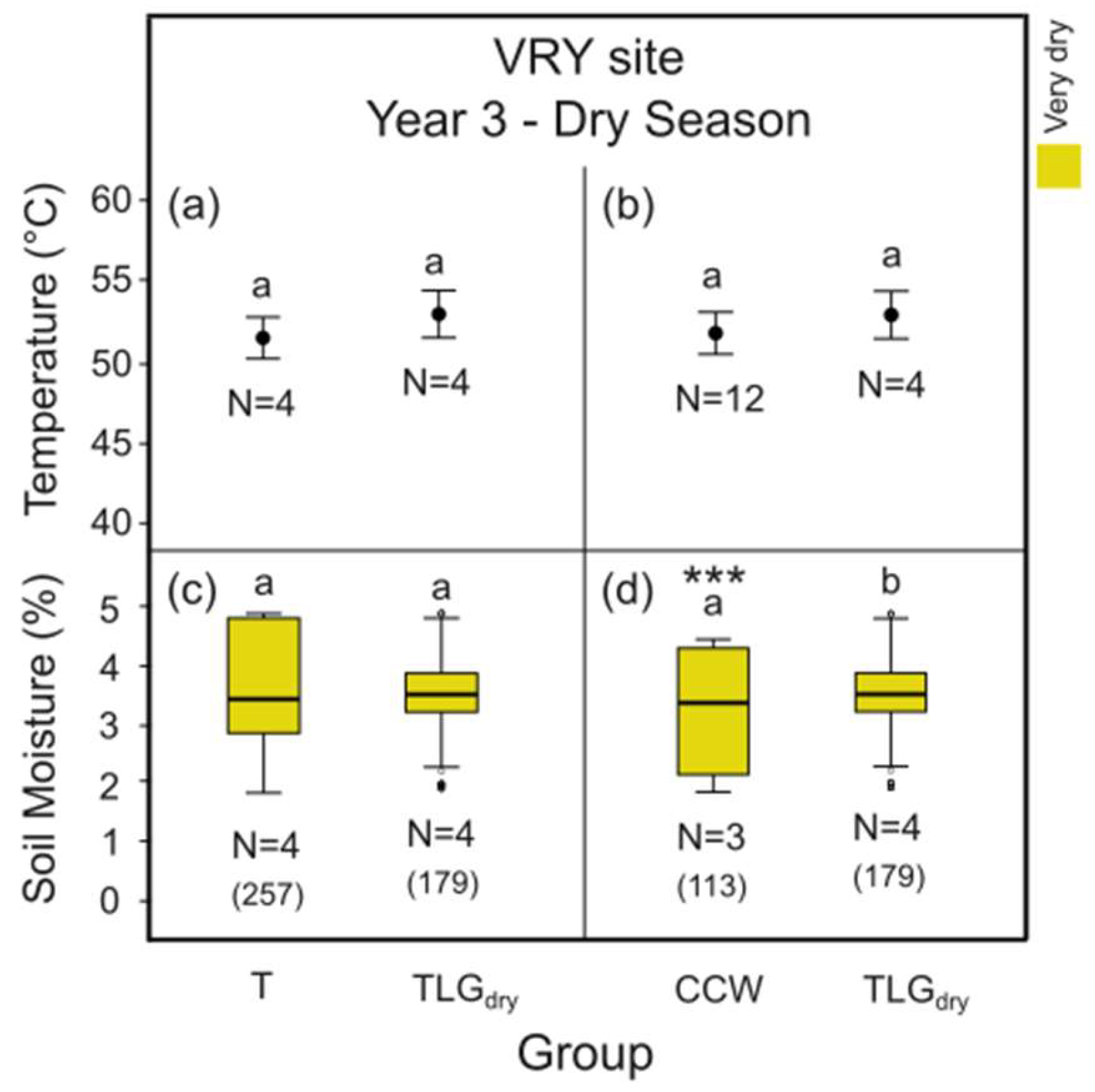
3.4. Thermal Stress on Soil and Soil Moisture
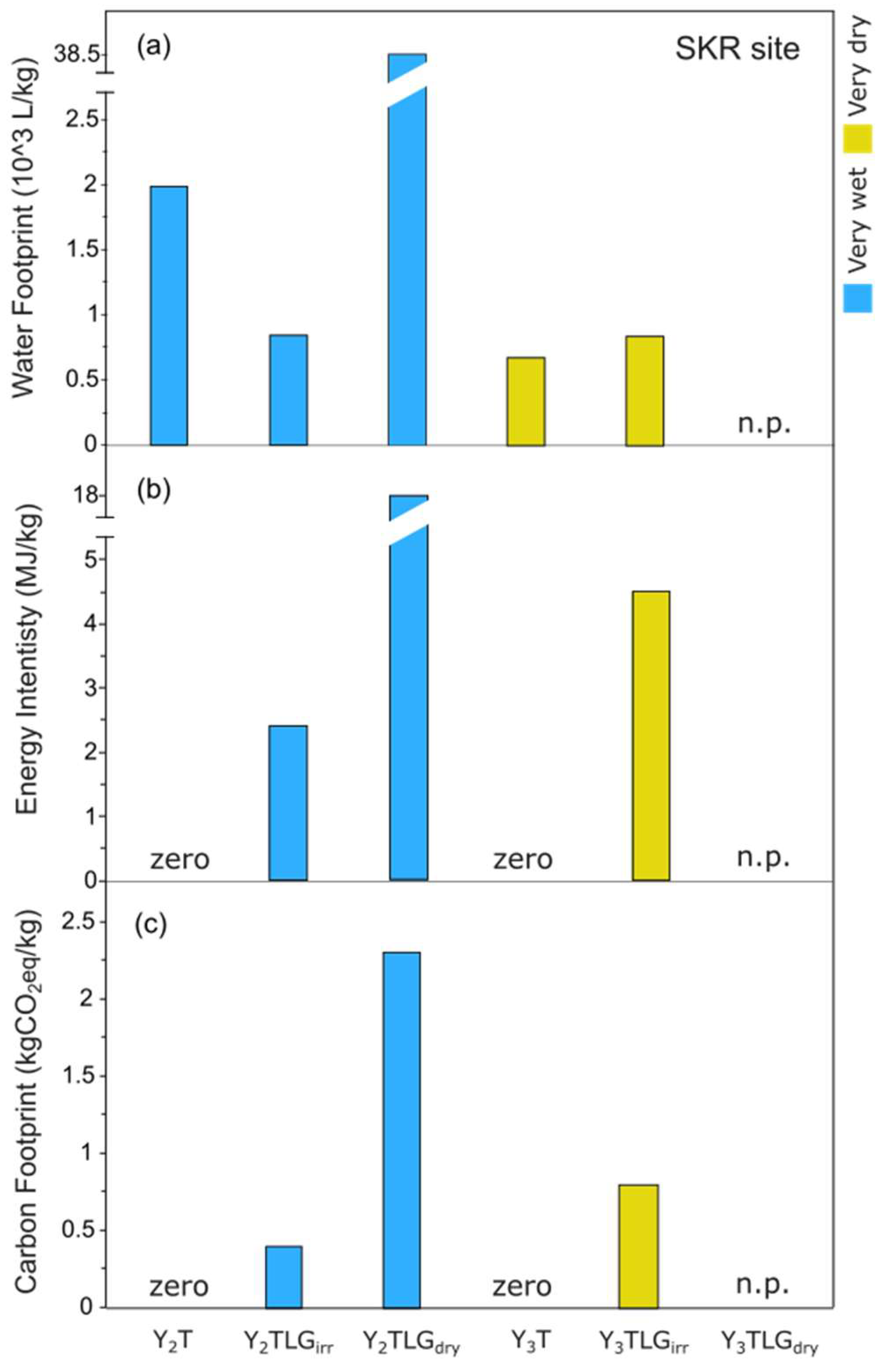
3.5. Environmental Performance Indicators of Carob Yield
4. Discussion
4.1. The Drivers for the Greater Carob Tree Productivity in T-System Plots
4.2. Overall Yield and Environmental Performance
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CB | Clover biomass |
| CCC | C-system with clover-based living mulch in 3 out of 3 years of monitoring |
| CCW | C-system with clover-based living mulch in 2 out of 3 years of monitoring |
| CF | Carbon Footprint |
| CR | Clover to clover plus weeds ratio |
| C-system | Clover (Trifolium squarossum)-based intercropping system with carob trees |
| CWB | Clover-and-weed biomass |
| CWSC | Soil cover by clover and wild vegetation |
| EI | Energy Intensity |
| SES | South (sun)-exposed soil side of carob trees |
| SKR | Skarinou site |
| TB | Thyme biomass |
| TLGdry | Conservation tillage carob trees without irrigation (control) |
| TLGirr | Conservation tillage carob trees with irrigation |
| TSC | Soil cover only by thyme |
| TSR | Thyme survival rate |
| T-System | Thyme (Thymbra capitata)-based intercropping system with carob trees |
| TWSC | Soil cover by thyme and wild vegetation |
| VRY | Vrysoulles site |
| WF | Water Footprint |
References
- Berdugo, M.; Delgado-Baquerizo, M.; Soliveres, S.; Hernández-Clemente, R.; Zhao, Y.; Gaitán, J.J.; Gross, N.; Saiz, H.; Maire, V.; Lehman, A.; et al. Global ecosystem thresholds driven by aridity. Science 2020, 367, 787–790. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Change 2015, 6, 166–171. [Google Scholar] [CrossRef]
- IUCN. Drylands and climate change. Issues Brief. International Union for Conservation of Nature and Natural Resources. Available online: https://www.iucn.org/resources/issues-brief/drylands-and-climate-change (accessed on 5 February 2025).
- Aguilera, E.; Díaz-Gaona, C.; García-Laureano, R.; Reyes-Palomo, C.; Guzmán, G.I.; Ortolani, L.; Sánchez-Rodríguez, M.; Rodríguez-Estévez, V. Agroecology for adaptation to climate change and resource depletion in the Mediterranean region. A review. Agric. Syst. 2020, 181, 102809. [Google Scholar] [CrossRef]
- Azzopardi, B.; Balzan, M.V.; Cherif, S.; Doblas-Miranda, E.; dos Santos, M.; Dobrinski, P.; Falder, M.; Hassoun, A.E.R.; Giupponi, C.; Koubi, V.V.; et al. Climate and Environmental Change in the Mediterranean Basin–Current Situation and Risks for the Future. First Mediterranean Assessment Report. 2020. Available online: https://www.medecc.org/medecc-reports/climate-and-environmental-change-in-the-mediterranean-basin-current-situation-and-risks-for-the-future-1st-mediterranean-assessment-report/ (accessed on 5 February 2025).
- Merino, A.; Doni, S.; Evelpidou, N.; Ferreira, T.; García-Arias, A.I.; Masciandaro, G.; Rodríguez-González, P.M. Best Practices in Evaluation and Restoration of Degraded Mediterranean Environments; London Library: London, UK, 2019; p. 327. [Google Scholar]
- Palm, C.; Blanco-Canqui, H.; DeClerck, F.; Gatere, L.; Grace, P. Conservation agriculture and ecosystem services: An overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef]
- Wittwer, R.A.; Dorn, B.; Jossi, W.; Van Der Heijden, M.G.A. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar] [CrossRef]
- Iseman, T.; Miralles-Wilhelm, F. Nature-Based Solutions in Agriculture: The Case and Pathway for Adoption; Food & Agriculture Organization: Rome, Italy, 2021. [Google Scholar]
- Almagro, M.; de Vente, J.; Boix-Fayos, C.; García-Franco, N.; Melgares de Aguilar, J.; González, D.; Solé-Benet, A.; Martínez-Mena, M. Sustainable land management practices as providers of several ecosystem services under rainfed Mediterranean agroecosystems. Mitig. Adapt. Strateg. Glob. Change 2016, 21, 1029–1043. [Google Scholar] [CrossRef]
- Lee, H.; Lautenbach, S.; Nieto, A.P.G.; Bondeau, A.; Cramer, W.; Geijzendorffer, I.R. The impact of conservation farming practices on Mediterranean agro-ecosystem services provisioning—A meta-analysis. Reg. Environ. Change 2019, 19, 2187–2202. [Google Scholar] [CrossRef]
- McLennon, E.; Dari, B.; Jha, G.; Sihi, D.; Kankarla, V. Regenerative agriculture and integrative permaculture for sustainable and technology driven global food production and security. Agron. J. 2021, 113, 4541–4559. [Google Scholar] [CrossRef]
- Rehman, S.; Ijaz, S.S.; Din, A.M.U.; Al-Dosary, M.A.; Ansar, M.; Fatima, S.; Siddiqa, A.; Ashraf, M.N.; Haider, I.; Junaid, M.B.; et al. Combined Effects of Reduced Tillage and Strip Intercropping on Soil Carbon Sequestration in Semi-Arid Environment. J. Soil Sci. Plant Nutr. 2025, 25, 3124–3139. [Google Scholar] [CrossRef]
- Mrunalini, K.; Behera, B.; Jayaraman, S.; Abhilash, P.C.; Dubey, P.K.; Swamy, G.N.; Prasad, J.V.N.S.; Rao, K.V.; Krishnan, P.; Pratibha, G.; et al. Nature-based solutions in soil restoration for improving agricultural productivity. Land Degrad. Dev. 2022, 33, 1269–1289. [Google Scholar] [CrossRef]
- Conti, J.; Cummins, N.; Gentry, B. Strategies for Operationalizing Nature-Based Solutions in the Private Sector. The Nature Conservancy Business Council-Natural Infrastructure Working Group 2019. (Whitepaper). Available online: https://www.preventionweb.net/publication/strategies-operationalizing-nature-based-solutions-private-sector (accessed on 5 February 2025).
- Lv, W.; Zhao, X.; Wu, P.; Lv, J.; He, H. A scientometric analysis of worldwide intercropping research based on web of science database between 1992 and 2020. Sustainability 2021, 13, 2430. [Google Scholar] [CrossRef]
- Maitra, S.; Hossain, A.; Brestic, M.; Skalicky, M.; Ondrisik, P.; Gitari, H.; Brahmachari, K.; Shankar, T.; Bhadra, P.; Palai, J.B.; et al. Intercropping—A low input agricultural strategy for food and environmental security. Agronomy 2021, 11, 343. [Google Scholar] [CrossRef]
- Gu, C.; Bastiaans, L.; Anten, N.P.R.; Makowski, D.; van der Werf, W. Annual intercropping suppresses weeds: A meta-analysis. Agric. Ecosyst. Environ. 2021, 322, 107658. [Google Scholar] [CrossRef]
- Marotti, I.; Whittaker, A.; Bağdat, R.B.; Akin, P.A.; Ergün, N.; Dinelli, G. Intercropping Perennial Fruit Trees and Annual Field Crops with Aromatic and Medicinal Plants (MAPs) in the Mediterranean Basin. Sustainability 2023, 15, 12054. [Google Scholar] [CrossRef]
- Restuccia, A.; Scavo, A.; Lombardo, S.; Pandino, G.; Fontanazza, S.; Anastasi, U.; Abbate, C.; Mauromicale, G. Long-term effect of cover crops on species abundance and diversity of weed flora. Plants 2020, 9, 1506. [Google Scholar] [CrossRef]
- Tziolas, E.; Ispikoudis, S.; Mantzanas, K.; Koutsoulis, D.; Pantera, A. Economic and Environmental Assessment of Olive Agroforestry Practices in Northern Greece. Agriculture 2022, 12, 851. [Google Scholar] [CrossRef]
- Casas, G.L.; Ciaccia, C.; Iovino, V.; Ferlito, F.; Torrisi, B.; Lodolini, E.M.; Giuffrida, A.; Catania, R.; Nicolosi, E.; Bella, S. Effects of Different Inter-Row Soil Management and Intra-Row Living Mulch on Spontaneous Flora, Beneficial Insects, and Growth of Young Olive Trees in Southern Italy. Plants 2022, 11, 545. [Google Scholar] [CrossRef]
- Li, C.; Stomph, T.J.; Makowski, D.; Li, H.; Zhang, C.; Zhang, F.; Van der Werf, W. The productive performance of intercropping. Proc. Natl. Acad. Sci. USA 2023, 120, e2201886120. [Google Scholar] [CrossRef]
- Li, C.; Hoffland, E.; Kuyper, T.W.; Yu, Y.; Zhang, C.; Li, H.; Zhang, F.; van der Werf, W. Syndromes of production in intercropping impact yield gains. Nat. Plants. 2020, 6, 653–660. [Google Scholar] [CrossRef]
- Tamburini, G.; Bommarco, R.; Wanger, T.C.; Kremen, C.; van der Heijden, M.G.A.; Liebman, M.; Hallin, S. Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv. 2020, 6, 45. [Google Scholar] [CrossRef]
- Kebede, E. Contribution, Utilization, and Improvement of Legumes-Driven Biological Nitrogen Fixation in Agricultural Systems. Front. Sustain. Food Syst. 2021, 5, 767998. [Google Scholar] [CrossRef]
- Ranaldo, M.; Carlesi, S.; Costanzo, A.; Bàrberi, P. Functional diversity of cover crop mixtures enhances biomass yield and weed suppression in a Mediterranean agroecosystem. Weed Res. 2020, 60, 96–108. [Google Scholar] [CrossRef]
- Chehade, L.; Antichi, D.; Frasconi, C.; Sbrana, M.; Tramacere, L.G.; Mazzoncini, M.; Peruzzi, A. Legume Cover Crop Alleviates the Negative Impact of No-Till on Tomato Productivity in a Mediterranean Organic Cropping System. Agronomy 2023, 13, 2027. [Google Scholar] [CrossRef]
- Morugán-Coronado, A.; Linares, C.; Gómez-López, M.D.; Faz, Á.; Zornoza, R. The impact of intercropping, tillage and fertilizer type on soil and crop yield in fruit orchards under Mediterranean conditions: A meta-analysis of field studies. Agric. Syst. 2020, 178, 102736. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, M.; Song, M.; Tian, J.; Song, B.; Hu, Y.; Zhang, J.; Yao, Y. Intercropping With Aromatic Plants Increased the Soil Organic Matter Content and Changed the Microbial Community in a Pear Orchard. Front. Microbiol. 2021, 12, 616932. [Google Scholar] [CrossRef]
- Sánchez-Navarro, V.; Shahrokh, V.; Martínez-Martínez, S.; Acosta, J.A.; Almagro, M.; Martínez-Mena, M.; Boix-Fayos, C.; Díaz-Pereira, E.; Zornoza, R. Perennial alley cropping contributes to decrease soil CO2 and N2O emissions and increase soil carbon sequestration in a Mediterranean almond orchard. Sci. Total Environ. 2022, 845, 157225. [Google Scholar] [CrossRef]
- Rao, S.G.R.; Radhika, R.M. Prospects for sustainable cultivation of medicinal and aromatic plants in India. In Sustainable Uses and Prospects of Medicinal Plants; CRC Press: Boca Raton, FL, USA, 2023; pp. 227–243. [Google Scholar]
- Rao, M.R.; Palada, M.C.; Becker, B.N. Medicinal and aromatic plants in agroforestry systems. In New Vistas in Agroforestry. Advances in Agroforestry: A Compendium for 1st World Congress of Agroforestry; Springer: Dordrecht, The Netherlands, 2004; Volume 1, pp. 107–122. [Google Scholar]
- Steenwerth, K.; Guerra, B. Influence of floor management technique on grapevine growth, disease pressure, and juice and wine composition: A review. Am. J. Enol. Vitic. 2012, 63, 149–164. [Google Scholar] [CrossRef]
- Montemurro, F.; Persiani, A.; Diacono, M. Cover crop as living mulch: Effects on energy flows in mediterranean organic cropping systems. Agronomy 2020, 10, 667. [Google Scholar] [CrossRef]
- Markou, M.; Moraiti, C.A.; Stylianou, A. Addressing Climate Change Impacts on Agriculture: Adaptation Measures For Six Crops in Cyprus. Atmosphere 2020, 11, 483. [Google Scholar] [CrossRef]
- Angon, P.B.; Anjum, N.; Akter, M.M.; Kc, S.; Suma, R.P.; Jannat, S. An Overview of the Impact of Tillage and Cropping Systems on Soil Health in Agricultural Practices. Adv. Agric. 2023, 2023, 861216. [Google Scholar] [CrossRef]
- Rusu, T. Energy efficiency and soil conservation in conventional, minimum tillage and no-tillage. Int. Soil Water Conserv. Res. 2014, 2, 42–49. [Google Scholar] [CrossRef]
- Reicosky, D.C. Conservation tillage is not conservation agriculture. J. Soil Water Conserv. 2015, 70, 103–108. [Google Scholar] [CrossRef]
- Chauhan, B.S.; Singh, R.G.; Mahajan, G. Ecology and management of weeds under conservation agriculture: A review. Crop Prot. 2012, 38, 57–65. [Google Scholar] [CrossRef]
- Battle, I.; Tous, J. Carob tree Ceratonia siliqua L. In Promoting the Conservation and Use of Underutilized and Neglected Crops; Leibniz Institute of Plant Genetics and Crop Plant Research: Gatersleben, Germany, 1997. [Google Scholar]
- Palaiogianni, A.; Stylianou, M.; Sarris, D.; Agapiou, A. Carob-agro-industrial waste and potential uses in the circular economy. In Mediterranean Fruits Bio-Wastes: Chemistry, Functionality and Technological Applications; Springer International Publishing: Berlin/Heidelberg, Germany, 2022; pp. 765–797. [Google Scholar]
- Goulas, V.; Stylos, E.; Chatziathanasiadou, M.V.; Mavromoustakos, T.; Tzakos, A.G. Functional components of carob fruit: Linking the chemical and biological space. Int. J. Mol. Sci. 2016, 17, 1875. [Google Scholar] [CrossRef]
- Stavrou, I.J.; Christou, A.; Kapnissi-Christodoulou, C.P. Polyphenols in carobs: A review on their composition, antioxidant capacity and cytotoxic effects, and health impact. Food Chem. 2018, 269, 355–374. [Google Scholar] [CrossRef]
- Flintan, F. Participatory rangeland management-an enabling process for improving silvopastoral management and governance. In Grazing with Trees; Haddad, F.F., Herrera, P.M., Besbes, B., Eds.; FAO: Rome, Italy, 2022. [Google Scholar]
- Valdecantos, A.; Vallejo, V.R.; Baeza, J.; Bautista, S.; Boeschoten, M.; Christoforou, M.; Daliakopoulos, I.N.; González-Pelayo, O.; Guixot, L.; Keizer, J.J.; et al. Report on the Restoration Potential for Preventing and Reversing Regime Shifts. 2016. Available online: https://research.wur.nl/en/publications/report-on-the-restoration-potential-for-preventing-and-reversing- (accessed on 5 February 2025).
- Diamond, N.K. An Agroforestry System for Semi-Arid Mediterranean Areas and Its Potential for Technology Transfer. Ph.D. Thesis, California Polytechnic State University, San Luis Obispo, CA, USA, 1988. [Google Scholar]
- Martins-Loução, M.A.; Correia, P.J.; Romano, A. Carob: A Mediterranean Resource for the Future. Plants 2024, 13, 1188. [Google Scholar] [CrossRef]
- Qin, J.; Duan, W.; Zou, S.; Chen, Y.; Huang, W.; Rosa, L. Global energy use and carbon emissions from irrigated agriculture. Nat. Commun. 2024, 15, 3084. [Google Scholar] [CrossRef]
- Karimi, F.; Ghahderijani, M.; Bakhoda, H. Optimizing cropping patterns and resource allocation for sustainable agricultural development: A case study of Ilam province. Iran. Environ. Sustain. Indic. 2024, 23, 100464. [Google Scholar] [CrossRef]
- Litskas, V.D.; Chrysargyris, A.; Tzortzakis, N.; Stavrinides, M.C.; Petropoulos, S.A. Can the commercial cultivation of wild edible species contribute to sustainable food production? A case study of golden thistle (Scolymus hispanicus L.). Int. J. Life Cycle Assess. 2025, 30, 446–461. [Google Scholar] [CrossRef]
- Moradi, P.; Ford-Lloyd, B.; Pritchard, J. Plant-water responses of different medicinal plant thyme (Thymus spp.) species to drought stress condition. Aust. J. Crop Sci. 2014, 8, 666–673. [Google Scholar]
- Cordovilla, M.P.; Bueno, M.; Aparicio, C.; Urrestarazu, M. Effects of Salinity and the Interaction Between Thymus Vulgaris and Lavandula Angustifolia on Growth, Ethylene Production and Essential Oil Contents. J. Plant Nutr. 2014, 37, 875–888. [Google Scholar] [CrossRef]
- Martínez-Mena, M.; Boix-Fayos, C.; Carrillo-López, E.; Díaz-Pereira, E.; Zornoza, R.; Sánchez-Navarro, V.; Acosta, J.A.; Martínez-Martínez, S.; Almagro, M. Short-term impact of crop diversification on soil carbon fluxes and balance in rainfed and irrigated woody cropping systems under semiarid Mediterranean conditions. Plant Soil 2021, 467, 499–514. [Google Scholar] [CrossRef]
- Almagro, M.; Díaz-Pereira, E.; Boix-Fayos, C.; Zornoza, R.; Sánchez-Navarro, V.; Re, P.; Fernández, C.; Martínez-Mena, M. The combination of crop diversification and no tillage enhances key soil quality parameters related to soil functioning without compromising crop yields in a low-input rainfed almond orchard under semiarid Mediterranean conditions. Agric. Ecosyst. Environ. 2023, 345, 108320. [Google Scholar] [CrossRef]
- Durán Zuazo, V.H.; Pleguezuelo, C.R.R.; Francia Martínez, J.R.; Martínez Raya, A.; Arroyo Panadero, L.; Càrceles Rodríguez, B.; Navarro Moll, M.C. Benefits of plant strips for sustainable mountain agriculture. Agron. Sustain. Dev. 2008, 28, 497–505. [Google Scholar] [CrossRef]
- IACO Ltd. National Action Plan to Combat Desertification in Cyprus—Contract No. 3/2007—Environment Service Executive Summary; I.A.CO Environmental & Water Consultants Ltd.: Nicosia, Cyprus, 2007. [Google Scholar]
- Matsi, S.; Sarris, D.; Konstantinou, M. Effects of Climate and Weeds on the Establishment of Thyme-Based Living Mulch Systems in Drylands of Cyprus. Int. J. Environ. Clim. Change 2024, 14, 791–803. [Google Scholar] [CrossRef]
- Wilson, J.B. Cover plus: Ways of measuring plant canopies and the terms used for them. J. Veg. Sci. 2011, 22, 197–206. [Google Scholar] [CrossRef]
- Davies, F.S.; Lakso, A.N. Diurnal and Seasonal Changes in Leaf Water Potential Components and Elastic Properties in Response to Water Stress in Apple Trees. Physiol. Plant. 1979, 46, 109–114. [Google Scholar] [CrossRef]
- Oyebade, B.A.; Anaba, J.C. Individual tree basal area equation for a young Tectona Grandis (Teak) plantation in Choba, Port Harcourt, Rivers State, Nigeria. World News Nat. Sci. 2018, 16, 130–140. [Google Scholar]
- Tous, J.; Romero, A.; Hermoso, J.F.; Ninot, A.; Plana, J.; Batlle, I. Agronomic and commercial performance of four Spanish carob cultivars. Horttechnology 2009, 19, 465–470. [Google Scholar] [CrossRef]
- Pawar, M.A.D.; Patil, M.S. Orchard Tree Canopy Detection. Int. Res. J. Eng. Technol. 2019, 6, 7642–7644. [Google Scholar]
- Liang, X.; Rehman, S.U.; Zhiqi, W.; Raza, M.A.; Haider, I.; Khalid, M.H.B.; Saeed, A.; Iqbal, Z.; Fatima, S.; Siddiqa, A.; et al. Impacts of Conservation Tillage on Agricultural Land Development: A Review. J. Soil Sci. Plant Nutr. 2024, 25, 428–449. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Eldridge, D.J.; Berdugo, M.; Trivedi, P.; Sokoya, B.; Cano-Díaz, C.; Abades, S.; Alfaro, F.; Bamigboye, A.R.; Bastida, F.; et al. Unearthing the soil-borne microbiome of land plants. Glob. Change Biol. 2024, 30, e17295. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.P.; Li, X.X.; Xiao, Z.H.; Lambers, H.; Li, L. Phosphorus facilitation and covariation of root traits in steppe species. New Phytol. 2020, 226, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Lithourgidis, A.S.; Dordas, C.A.; Damalas, C.A.; Vlachostergios, D.N. Annual intercrops: An alternative pathway for sustainable agriculture. Aust. J. Crop Sci. 2011, 5, 396–410. [Google Scholar]
- Constantinou, E.; Sarris, D.; Vogiatzakis, I.N. The possible role of Ziziphus lotus as an ecosystem engineer in semiarid landscapes. J. Arid Environ. 2021, 195, 104614. [Google Scholar] [CrossRef]
- Constantinou, E.; Sarris, D.; Psichoudaki, M.; Cabello, J.; Vogiatzakis, I.N. How can ecosystem engineer plants boost productivity in east Mediterranean drylands. Ecol. Process. 2023, 12, 30. [Google Scholar] [CrossRef]
- Constantinou, E.; Montesinos-Navarro, A.; Sarris, D.; Vogiatzakis, I.N. Facilitation network in phryganic plant communities: Evidence from a Mediterranean island. Plant Biosyst. 2025, 159, 596–605. [Google Scholar] [CrossRef]
- Ahmed, Z.; Gui, D.; Abd-Elmabod, S.K.; Murtaza, G.; Ali, S. An overview of global desertification control efforts: Key challenges and overarching solutions. Soil Use Manag. 2024, 40, e13154. [Google Scholar] [CrossRef]
- Gerowitt, B.; Bàrberi, P.; Darmency, H.; Petit, S.; Storkey, J.; Westerman, P. Weeds and biodiversity. In Weed Research: Expanding Horizons; John Wiley & Sons Ltd.: West Sussex, UK, 2017. [Google Scholar]
- Valdes, Y.B. The role of weeds as a component of biodiversity in agroecosystems. Cultiv. Trop. 2016, 37, 34–56. [Google Scholar] [CrossRef]
- Correia, P.J.; Cota, T.; Pestana, M. Evaluation of Carob Tree Productivity during a 30-Year Period, in Relation to Precipitation and Air Temperature. Environ. Process. 2020, 7, 1221–1233. [Google Scholar] [CrossRef]
- Correia, P.J.; Anastácio, I.; Da Fé Candeias, M.; Martins-Loução, M.A. Nutritional diagnosis in carob-tree: Relationships between yield and leaf mineral concentration. Crop Sci. 2002, 42, 1577–1583. [Google Scholar] [CrossRef]
- Bonanomi, G.; Stinca, A.; Chirico, G.B.; Ciaschetti, G.; Saracino, A.; Incerti, G. Cushion plant morphology controls biogenic capability and facilitation effects of Silene acaulis along an elevation gradient. Funct. Ecol. 2016, 30, 1216–1226. [Google Scholar] [CrossRef]
- Bonanomi, G.; Idbella, M.; Stinca, A.; Maisto, G.; De Marco, A.; Del Galdo, G.P.G.; Guarino, R.; Zotti, M. Nitrogen-fixing cushion Astragalus siculus modulates soil fertility, microclimate, plant facilitation, bacterial and fungal microbiota along an elevation gradient. J. Veg. Sci. 2023, 34, e13193. [Google Scholar] [CrossRef]
- Momberg, M.; Roux, P.C. Testing for consistency in ecosystem engineering: Do cushion plants always turn up the heat? Acta Oecologica 2020, 104, 103532. [Google Scholar] [CrossRef]
- Pugnaire, F.I.; Losapio, G.; Schöb, C. Species interactions involving cushion plants in high-elevation environments under a changing climate. Ecosistemas 2021, 30, 2186. [Google Scholar] [CrossRef]
- Tátrai, Z.A.; Sanoubar, R.; Pluhár, Z.; Mancarella, S.; Orsini, F.; Gianquinto, G. Morphological and Physiological Plant Responses to Drought Stress in Thymus citriodorus. Int. J. Agron. 2016, 2016, 4165750. [Google Scholar] [CrossRef]
- Nyawade, S.O.; Karanja, N.N.; Gachene, C.K.K.; Gitari, H.I.; Schulte-Geldermann, E.; Parker, M.L. Intercropping Optimizes Soil Temperature and Increases Crop Water Productivity and Radiation Use Efficiency of Rainfed Potato. Am. J. Potato Res. 2019, 96, 457–471. [Google Scholar] [CrossRef]
- Safari, N.; Kazemi, F.; Tehrani, A. Examining temperature and soil moisture contents of mulches in the urban landscaping of an arid region. Desert 2021, 26, 139–156. [Google Scholar] [CrossRef]
- Correia, P.J.; Martins-Loução, M.A. The use of macronutrients and water in marginal Mediterranean areas: The case of carob-tree. Field Crops Res. 2005, 91, 1–6. [Google Scholar] [CrossRef]
- Kolyva, F.; Stratakis, E.; Rhizopoulou, S.; Chimona, C.; Fotakis, C. Leaf surface characteristics and wetting in Ceratonia siliqua L. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 551–556. [Google Scholar] [CrossRef]
- Sarris, D.; Mazza, G. Mediterranean Pine Root Systems Under Drought. In Pines and Their Mixed Forest Ecosystems in the Mediterranean Basin; Ne’eman, G., Osem, Y., Eds.; Managing Forest Ecosystems; Springer: Cha, Switzerland, 2021; Volume 38, pp. 129–140. [Google Scholar] [CrossRef]
- Sarris, D.; Christodoulakis, D. Topographic and climatic effects on Pinus halepensis s.l. growth at its drought tolerance margins under climatic change. J. For. Res. 2024, 35, 102. [Google Scholar] [CrossRef]
- Markou, L. Effects of Intercropping Systems on Carob Productivity Under Rainfed Conditions, Master in Ecology and Biodiversity. Master’s Thesis, Department of Biological Sciences, University of Cyprus, Nicosia, Cyprus, 2021. [Google Scholar]
- Dorman, M.; Perevolotsky, A.; Sarris, D.; Svoray, T. Amount vs. temporal pattern: On the importance of intra-annual climatic conditions on tree growth in a dry environment. J. Arid. Environ. 2015, 118, 65–68. [Google Scholar] [CrossRef]
- Mazza, G.; Sarris, D. Identifying the full spectrum of climatic signals controlling a tree species’ growth and adaptation to climate change. Ecol. Indic. 2021, 130, 108109. [Google Scholar] [CrossRef]
- Mazza, G.; Markou, L.; Sarris, D. Species-specific growth dynamics and vulnerability to drought at the single tree level in a Mediterranean reforestation. Trees Struct. Funct. 2021, 35, 1697–1710. [Google Scholar] [CrossRef]
- Mazza, G.; Manetti, M.C.; Kraushaar, G.; Pezzi, G.; Krebs, P.; Coşgun, S.; Tinner, W.; Conedera, M. Climate sensitivity of Abies alba Mill. in marginal Mediterranean low-elevation stands reveals new insights into the ecological potential of the species. For. Ecol. Manag. 2025, 585, 122670. [Google Scholar] [CrossRef]
- von Hasalberg, C.D. Vegetative Growth and Flower and Fruit Development in Carob Tree (Ceratonia siliqua L.) with Special Emphasis on Environmental Conditions at Marginal Production Sites in South Portugal. Ph.D. Thesis, Humboldt-Universitat Zu Berlin, Berlin, Germany, 2000. [Google Scholar]
- Tous, J.; Romero, A.; Batlle, I. The Carob Tree: Botany, Horticulture and Genetic Resources. In Horticultural Reviews; John Wiley & Sons Inc.: Hobokeny, NJ, USA, 2013; Volume 41, pp. 385–456. [Google Scholar]
- Von Haselberg, C. Factors influencing flower and fruit development in carob (Ceratonia siliqua L.). In Proceedings of the Third International Carob Symposium, Tavira, Portugal, 9–11 September 1996. [Google Scholar]
- Moore, E.A.; Norton, U. Improving semi-arid agroecosystem services with cover crop mixes. PLoS ONE 2024, 19, e0306567. [Google Scholar] [CrossRef]
- Opoku, A.; Ogunleye, A.M.; Solomon, J.K.Q.; Payne, W.A. Cover crop systems impact on biomass production, carbon-to-nitrogen ratio, forage quality, and soil health in a semi-arid environment. Heliyon 2024, 10, e39600. [Google Scholar] [CrossRef]
- Everwand, G.; Cass, S.; Dauber, J.; Williams, M.; Stout, J. Legume crops and biodiversity. In Legumes in Cropping Systems; CABI: Wallingford, UK, 2017; pp. 55–59. [Google Scholar]
- Simoes, M.P.; Belo, A.F.; Pinto-Cruz, C.; Pinheiro, A.C. Natural vegetation management to conserve biodiversity and soil water in olive orchards. Span. J. Agric. Res. 2014, 12, 633–643. [Google Scholar] [CrossRef]
- Zhu, S.G.; Zhu, H.; Zhou, R.; Zhang, W.; Wang, W.; Zhou, Y.N.; Wang, B.Z.; Yang, Y.M.; Wang, J.; Tao, H.Y.; et al. Intercrop overyielding weakened by high inputs: Global meta-analysis with experimental validation. Agric. Ecosyst. Environ. 2023, 342, 108239. [Google Scholar] [CrossRef]
- Brooker, R.; Kikvidze, Z.; Pugnaire, F.I.; Callaway, R.M.; Choler, P.; Lortie, C.J.; Michalet, R. The importance of importance. Oikos 2005, 109, 63–70. [Google Scholar] [CrossRef]
- Echarte, L.; Della Maggiora, A.; Cerrudo, D.; Gonzalez, V.H.; Abbate, P.; Cerrudo, A.; Sadras, V.O.; Calvino, P. Yield response to plant density of maize and sunflower intercropped with soybean. Field Crops Res. 2011, 121, 423–429. [Google Scholar] [CrossRef]
- Willey, R.W. Resource use in intercropping systems. Agric. Water Manag. 1990, 1–3, 215–231. [Google Scholar] [CrossRef]
- Rossi, L.; Regni, L.; Rinaldi, S.; Sdringola, P.; Calisti, R.; Brunori, A.; Dini, F.; Proietti, P. Long-term water footprint assessment in a rainfed olive tree grove in the Umbria region, Italy. Agriculture 2020, 10, 8. [Google Scholar] [CrossRef]
- Litskas, V.; Mandoulaki, A.; Vogiatzakis, I.N.; Tzortzakis, N.; Stavrinides, M. Sustainable viticulture: First determination of the environmental footprint of grapes. Sustainability 2020, 12, 8812. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exp.: Hydrological Year | r (mm) | % Change from Previous Year | % of Normal r | Ta (°C) | DMAI | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| SKR | VRY | SKR | VRY | SKR | VRY | SKR | VRY | SKR | VRY | |
| Y0: 2018–2019 | 678 | 469 | 160 | 142 | 20.4 | 20.5 | 22.28 | 15.35 | ||
| Y1: 2019–2020 | 655 | 513 | −<1% | +<1% | 154 | 155 | 20.8 | 20.4 | 21.29 | 16.88 |
| Y2: 2020–2021 | 311 | 289 | −53% | −44% | 73 | 87 | 21.2 | 20.8 | 9.96 | 9.40 |
| Y3: 2021–2022 | 496 | 283 | 59% | −2% | 117 | 85 | 20 | 20.3 | 15.77 | 9.33 |
| SKR Site | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T-System | C-System: CCC | ||||||||||||
| Exp. Year | Climate Year | Weed Comp. | TB (g/m2) Mean | TSC (%) Mean | TWSC (%) Mean | TSR (%) Median | CB (g/m2) Mean | TB (g/m2) Mean | TSC (%) Mean | CWB (g/m2) Mean | CR (%) Mean | CWSC (%) Mean | TSR (%) Median |
| Y1 | Very wet | Low | 43 | 15 | >40 | 97 | 157 | 1 | <0.3 | 215 | 70 | >90 | 2 |
| Y2 | Very dry | Low | 335 | 35 | 55 | 97 | 329 * | 97 * | |||||
| Y3 | Normal | Low | 420 | 83 | 85 | 94 | >180 | >320 | 55 | 100 | |||
| VRY Site | |||||||||||||
| T-System | C-System: CCW | ||||||||||||
| Y1 | Very wet | High | 28 | 11 | >55 | 72 | 171 | 0 | 0 | 305 | 50 | 97 | 0 |
| Y2 | Very dry | High | 39 | 9 | >80 | 40 | >400 * | >95 * | |||||
| Y3 | Very dry | High | 45 | 25 | >97 | 40 | 22 | 316 | 7 | 100 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsi, S.; Sarris, D.; Litskas, V. Carob–Thyme Intercropping Systems Can Improve Yield Efficiency and Environmental Footprint Compared to Conservation Tillage. Agronomy 2025, 15, 1560. https://doi.org/10.3390/agronomy15071560
Matsi S, Sarris D, Litskas V. Carob–Thyme Intercropping Systems Can Improve Yield Efficiency and Environmental Footprint Compared to Conservation Tillage. Agronomy. 2025; 15(7):1560. https://doi.org/10.3390/agronomy15071560
Chicago/Turabian StyleMatsi, Sofia, Dimitrios Sarris, and Vassilis Litskas. 2025. "Carob–Thyme Intercropping Systems Can Improve Yield Efficiency and Environmental Footprint Compared to Conservation Tillage" Agronomy 15, no. 7: 1560. https://doi.org/10.3390/agronomy15071560
APA StyleMatsi, S., Sarris, D., & Litskas, V. (2025). Carob–Thyme Intercropping Systems Can Improve Yield Efficiency and Environmental Footprint Compared to Conservation Tillage. Agronomy, 15(7), 1560. https://doi.org/10.3390/agronomy15071560