1. Introduction
Miscanthus (
Miscanthus × giganteus) and willow (
Salix spp.) are among the leading candidates for sustainable bioenergy production due to their high biomass yields and adaptability to diverse environmental conditions [
1]. This manuscript represents Part 2 of a two-part study. In Part 1 [
2], we assessed biomass yield potential, energy outputs, and input requirements across spatial and temporal scales under current and future climate scenarios using process-based modelling. Building on that foundation, Part 2 expands the analysis to include additional dimensions that are critical to assessing the broader sustainability of bioenergy systems: land use energy intensity (LUEI), the impacts of biomass transport on energy use efficiency (EUE), and soil carbon sequestration. These additional aspects were not fully explored in Part 1 due to scope limitations and data complexity; they require dedicated analysis, particularly for SOC, which depends on detailed biomass partitioning and turnover dynamics, and on EUE, which involves logistics and transport energy not addressed previously. By incorporating these new components, Part 2 provides a more holistic view of the environmental and energy implications of bioenergy crop deployment. For the purposes of this study, we consider the values relating to the biomass feedstock used as fuel at the farm gate or after transportation at the fuel use location gate; we do not consider thermal conversion efficiencies of the end use. While both crops are assessed for LUEI, analyses of biomass transport and carbon sequestration are conducted exclusively for
Miscanthus, as comparable data for willow were unavailable.
Miscanthus is widely recognized for its high biomass yield and substantial carbon sequestration potential, making it a promising option for sustainable bioenergy production and long-term carbon storage in agricultural systems [
3]. Its perennial growth cycle and extensive root system are valuable in promoting the accumulation of soil organic carbon (SOC), contributing to improved soil health and climate mitigation [
4,
5]. However, evaluating the true sustainability of
Miscanthus-based bioenergy systems requires more than just agronomic assessments. Energy use and emissions associated with biomass transport can significantly influence the overall EUE and carbon balance, potentially offsetting some of bioenergy benefits [
6]. To address this, the current study incorporates biomass transport considerations into the sustainability assessment of
Miscanthus, offering a more comprehensive evaluation that accounts for logistical constraints alongside agronomic potential. Through analysis of transportation-related energy consumption and greenhouse gas (GHG) emissions, the study aims to identify strategies that reduce fossil fuel dependence, improve EUE, and minimize climate impacts [
2]. This integrated approach provides critical insights for optimizing biomass supply chains, informing decisions about processing site locations, and guiding policy development to support low-carbon bioenergy systems. Although willow is not included in the carbon sequestration and transport analysis presented in this study, it remains an important species in bioenergy research. Its deep-rooted system contributes to soil stability and nutrient cycling, offering valuable ecosystem services beyond the scope of this study [
7]. However, other woody biomass species may be more suitable for use in tropical and subtropical climate zones, but are not considered here.
To conduct this analysis, we applied the MiscanFor and SalixFor models, which simulate biomass yields and energy requirements by integrating climatic conditions, soil properties, and management data [
6]. These process-based models are instrumental in evaluating the energy performance of bioenergy crops, particularly under projected climate change scenarios which may significantly influence both productivity and resource demands. Estimation of SOC turnover involves quantifying the incorporation of organic carbon from above- and below-ground biomass into the soil matrix. This is a key process in determining the crop’s long-term contribution to SOC accumulation, an essential factor for improving soil health, fertility, and carbon sequestration potential. Reliable assessment of these carbon inputs is vital for understanding the broader role of
Miscanthus systems in GHG mitigation and sustainable land use strategies [
8].
By integrating energy yield analysis, transport impacts, and SOC dynamics, this study provides a comprehensive evaluation of
Miscanthus and willow as sustainable bioenergy crops. It contributes to the ongoing discussion on their viability within the context of climate change mitigation and the global energy transition. Ultimately, the findings from this study underline the necessity of holistic approaches to bioenergy system design, approaches that account for productivity, environmental performance, and logistical considerations to maximize climate benefits [
4,
5,
6]. Therefore, the aim of this study is to provide an integrated sustainability assessment of
Miscanthus and willow as bioenergy crops by evaluating LUEI, the impacts of biomass transport on EUE, and soil carbon sequestration under current and projected future climate conditions.
4. Discussion
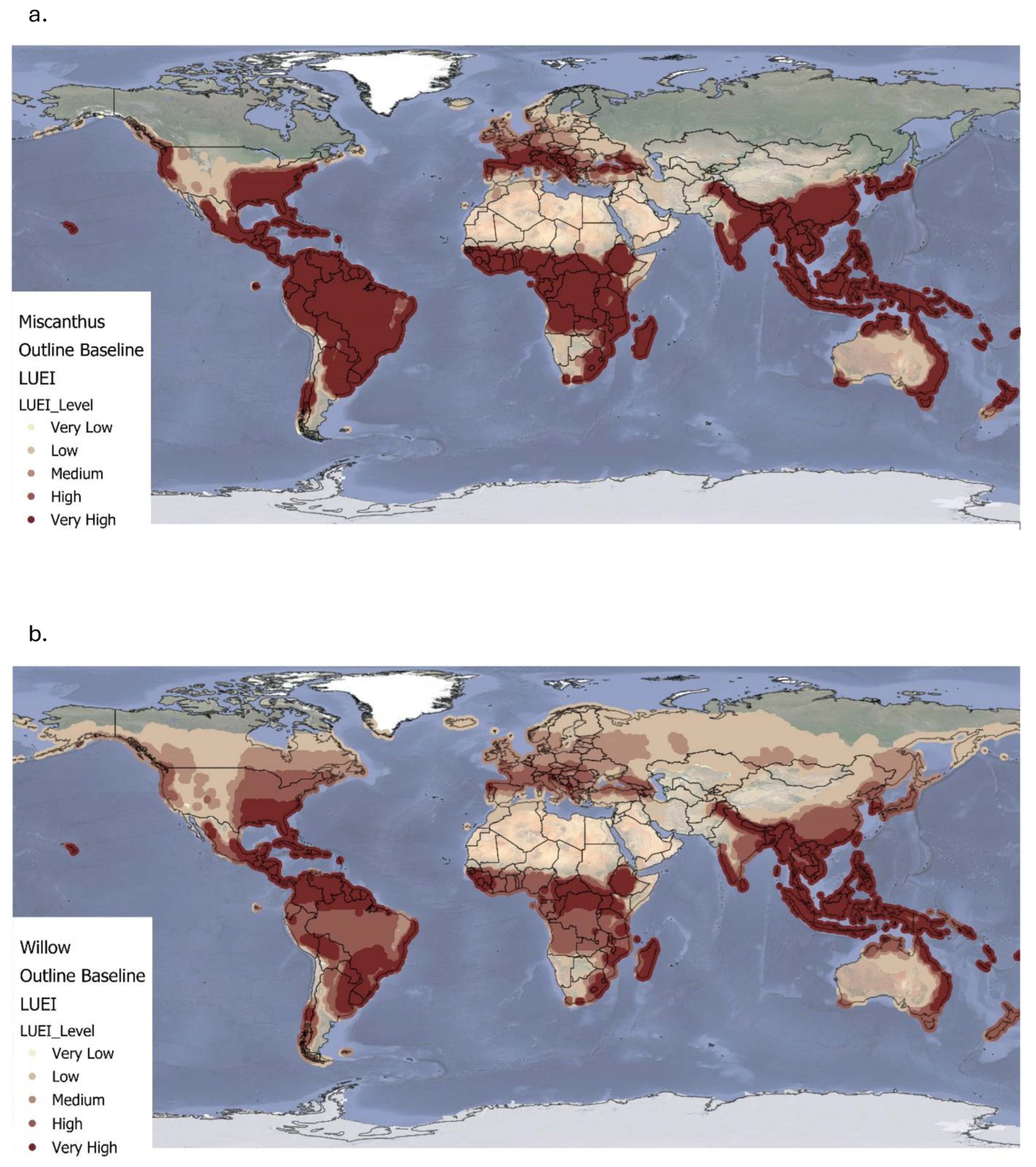
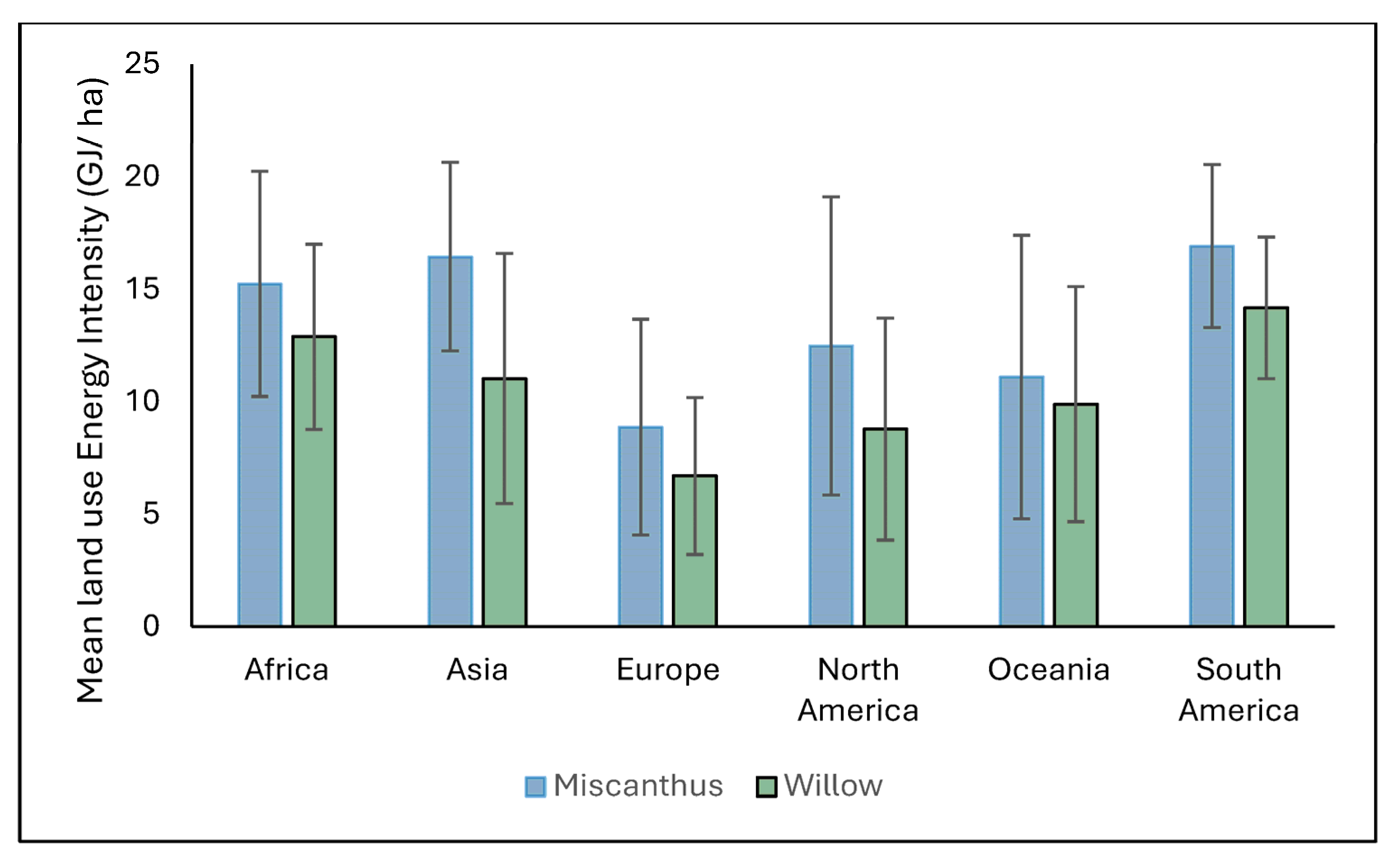
4.1. Land Use Energy Intensity for Miscanthus and Willow Under Current Climate Conditions
The spatial variation in LUEI for
Miscanthus highlights its strong dependence on climatic and biophysical conditions. On average,
Miscanthus delivers a high LUEI (321 ± 179.1 GJ ha
−1 y
−1), particularly in tropical and subtropical regions. These areas benefit from high solar radiation, long growing seasons, and sufficient rainfall, all of which promote high biomass yields and strong annual net energy returns [
1]. Our findings show LUEI values exceeding 700 GJ ha
−1 in tropical hotspots comparable to the success of sugarcane (a close relative of
Miscanthus) used in Brazil for ethanol and bagasse-based bioenergy production. In contrast, cooler temperate and boreal zones, such as Scandinavia, the Baltics, northern Russia, Canada, and parts of eastern Russia, exhibit much lower LUEI values, with some regions even experiencing negative energy balances. These results reflect biophysical limitations including shorter growing seasons, reduced heat accumulation, and lower winter survival, all of which constrain biomass production and reduce energy efficiency [
2].
Willow, used here as a representative of coppiced woody biomass, typically achieves a lower mean annual LUEI (164 ± 115.6 GJ ha
−1 y
−1) compared to
Miscanthus. This is primarily due to its C3 photosynthetic pathway and generally lower yield potential. However, willow exhibits relatively stable energy returns across a broader geographic range, particularly in temperate regions like the central and eastern United States and Central Europe [
19,
20]. Although it does not achieve the high yields of
Miscanthus, willow’s resilience makes it suitable for a wider range of latitudes and climatic conditions.
Regionally, both crops display high variability, but Miscanthus consistently delivers higher energy returns per hectare than willow. Even so, willow’s broader climatic suitability points to its complementary role in areas where Miscanthus performance is marginal. It is worth noting that MiscanFor’s LUEI calculations do not include energy costs associated with end-of-life crop removal, unless specifically modelled, which may affect full lifecycle assessments.
The concentration of high LUEI values in tropical and subtropical zones reinforces the strategic bioenergy potential of these areas. However, expansion in these regions must be carefully managed to avoid negative impacts on land availability, food security, and ecosystem conservation [
21,
22]. Thus, integrating environmental, economic, and social factors into land use planning is critical to aligning bioenergy deployment with broader sustainable development goals [
23]. While
Miscanthus delivers higher net energy returns under optimal conditions, willow’s broader climatic adaptability positions it as a valuable alternative. These findings support a dual-crop strategy that aligns crop selection with regional suitability and long-term sustainability goals.
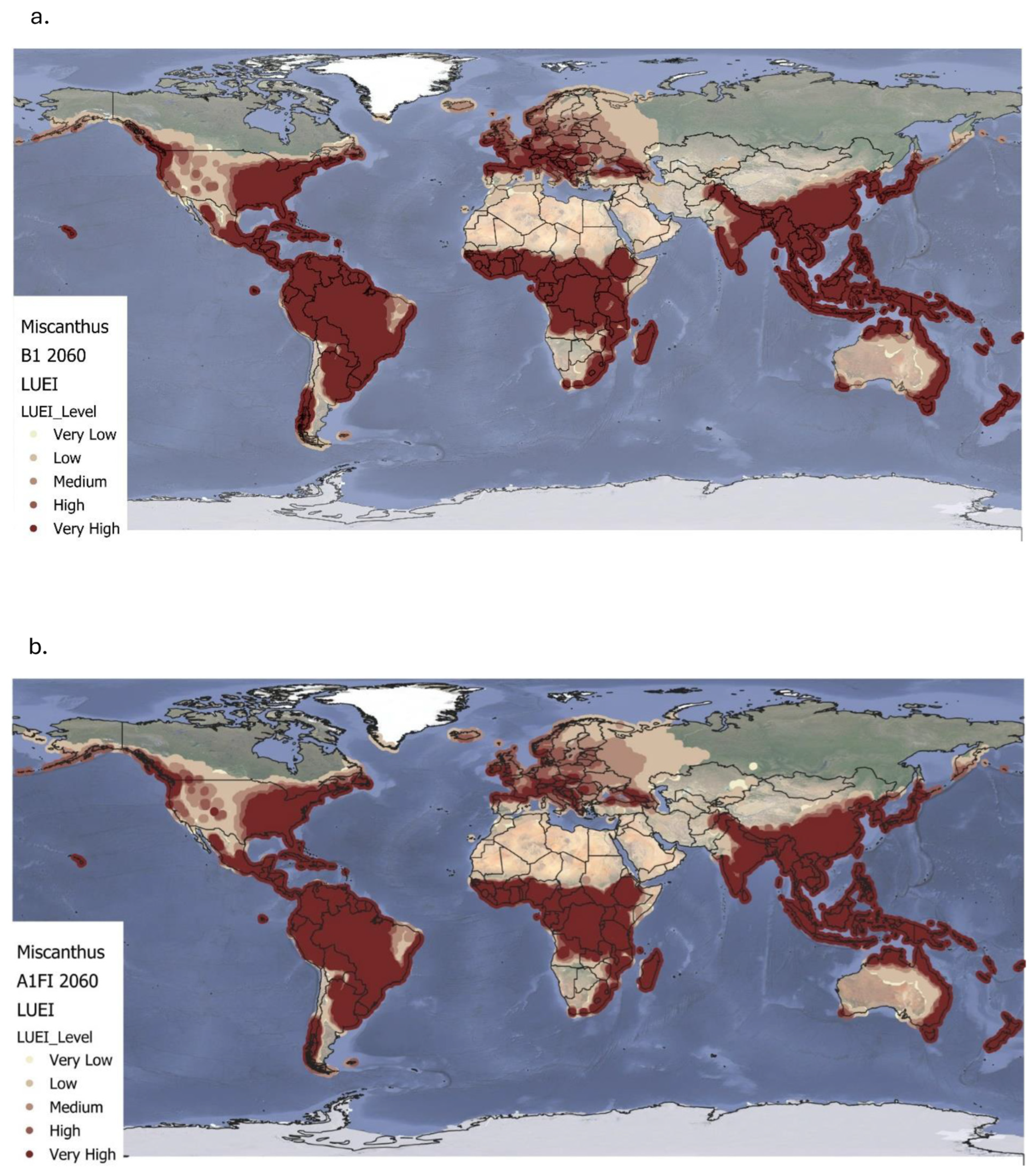
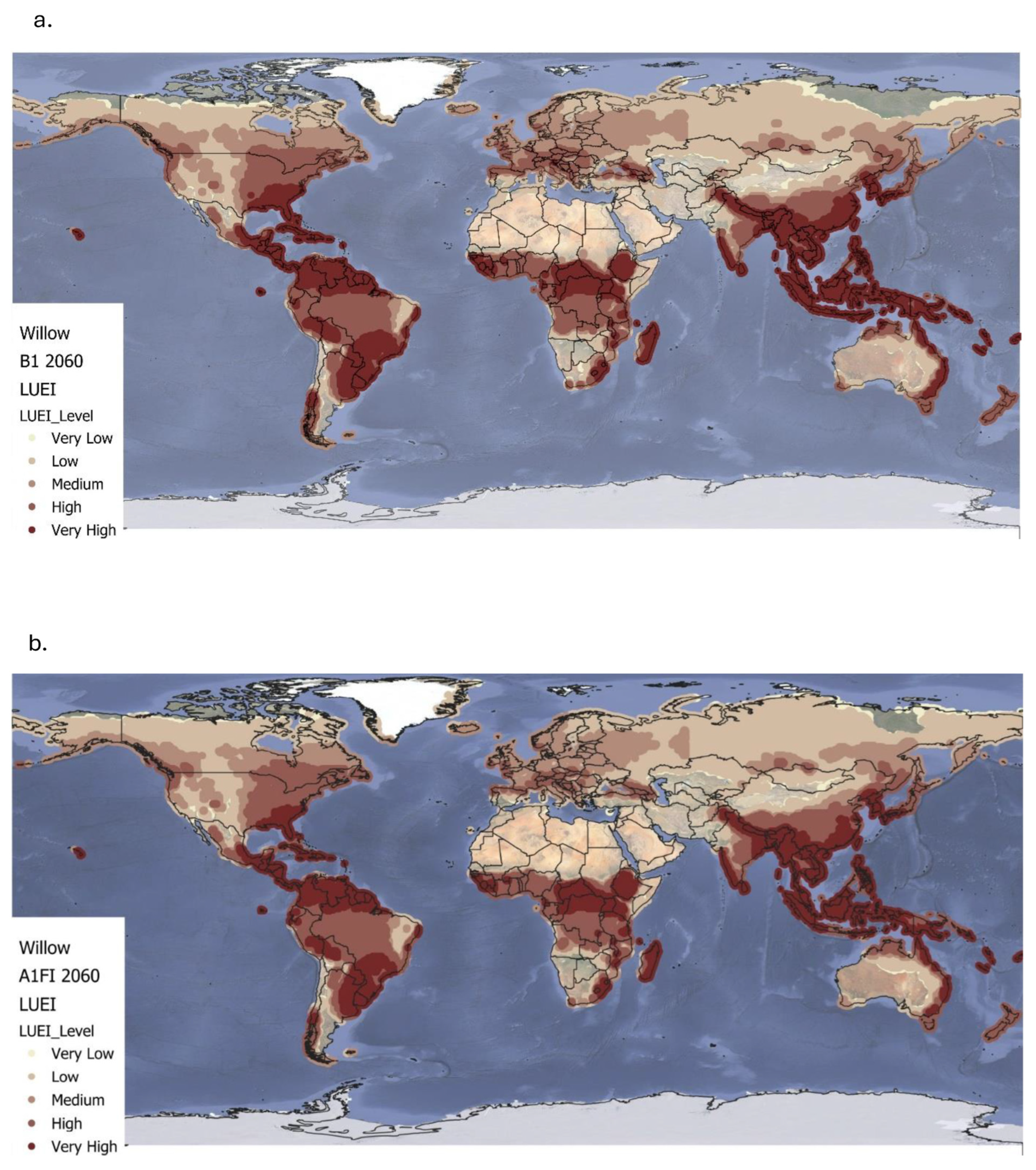
4.2. Land Use Energy Intensity for Miscanthus and Willow Under Future Climate Conditions
Under the B1 climate scenario up to 2060, both
Miscanthus and willow maintain high to very high LUEI across tropical and subtropical regions due to favourable conditions such as consistent solar radiation, adequate rainfall, and optimal temperatures [
24,
25,
26]. These results reinforce previous findings on the ecological adaptability of both crops to warm climates [
26,
27].
Beyond these stable regions, B1 also facilitates an expansion of medium-to-high LUEI into temperate areas for both species. Southern parts of the United States, southern and eastern Europe, and parts of Central Asia show increased suitability driven by extended growing seasons and milder winters [
6,
8,
28,
29]. These patterns suggest that moderate warming may improve biomass yields in areas previously seen as marginal. Despite this geographic expansion, global average LUEI declines modestly under B1 by 9% for
Miscanthus and 8% for willow likely due to increased climate variability, shifts in precipitation patterns, or emerging pest and disease pressures [
23,
29]. These findings highlight the trade-off between spatial expansion and localized productivity declines in core growing zones.
Under the more extreme A1FI scenario, the LUEI trends for
Miscanthus and willow diverge more dramatically. For both crops, temperate regions including the United States, Midwest and large parts of Europe experience significant LUEI gains due to warmer temperatures and longer frost-free periods [
30,
31]. These shifts suggest potential benefits in utilizing both crops for bioenergy production as part of climate adaptation strategies in temperate zones. However, in tropical and subtropical regions, A1FI triggers yield declines for both species. Rising heat stress, drought, and disrupted nutrient cycles in these areas appear to surpass the physiological thresholds of both crops, reducing their net energy returns [
32]. For
Miscanthus, this results in a 15% drop in global LUEI, while willow shows a 13% reduction compared to baseline levels. These findings mirror broader concerns about the limits of ecological resilience under high-emissions scenarios [
23,
33].
In both cases, future viability will depend on region-specific climate conditions, particularly temperature extremes and water availability. While moderate climate change may expand bioenergy opportunities, high-emissions futures pose risks to productivity stability.
To maximize the potential of
Miscanthus and willow as sustainable bioenergy crops, adaptive land use planning and targeted breeding programmes are essential. These should focus on enhancing stress tolerance, especially in marginal environments, and managing the transition into new suitable areas [
34]. Furthermore, although willow is used here as a proxy for short rotation coppice biomass due to widespread trials, other woody crops such as poplar and eucalyptus may prove better suited to some regions, particularly outside temperate climates.
Ultimately, integrated policy frameworks must address where these crops can be grown, how to manage them sustainably, and how to align their use with broader goals for climate resilience, food security, and biodiversity protection [
35].
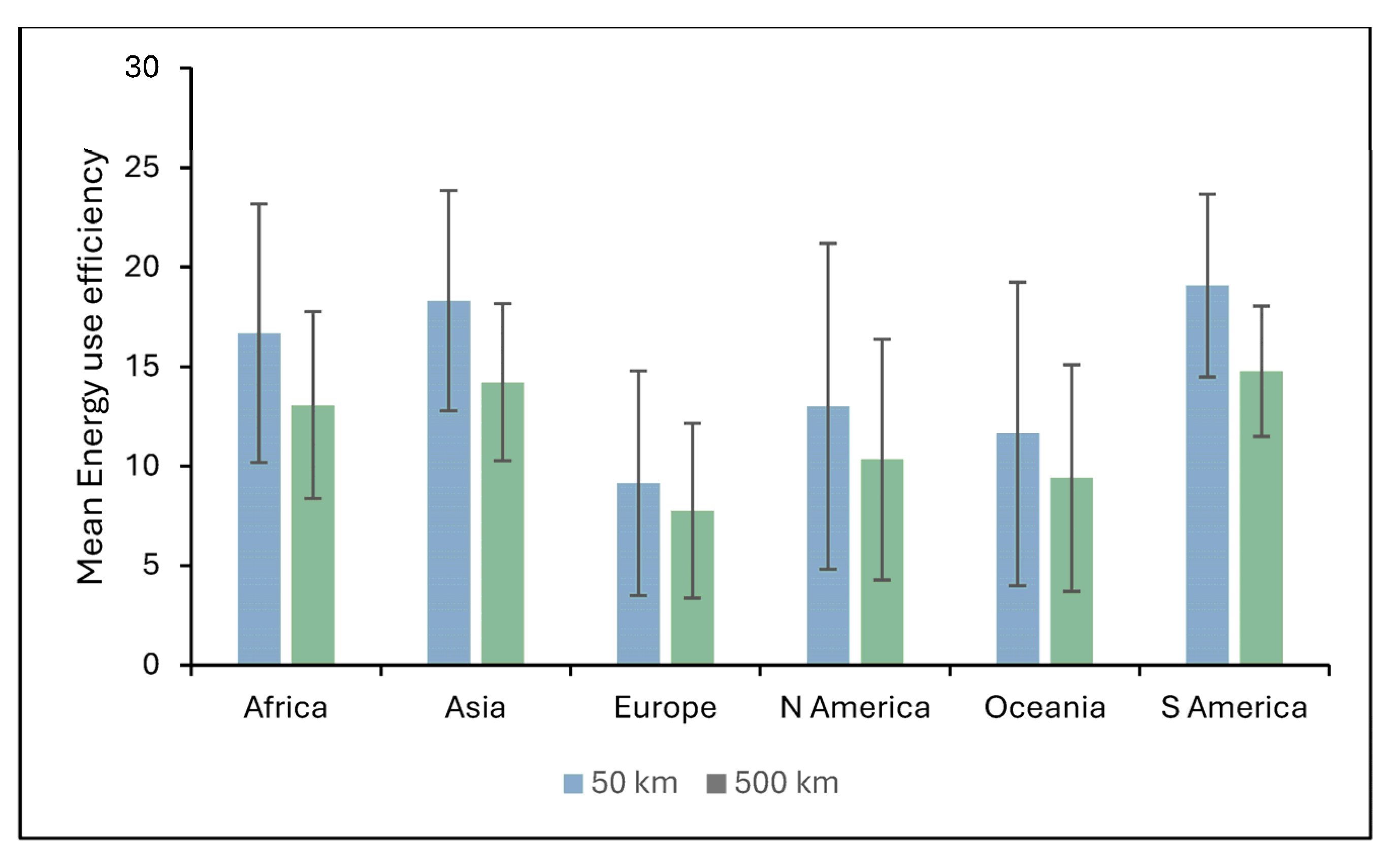
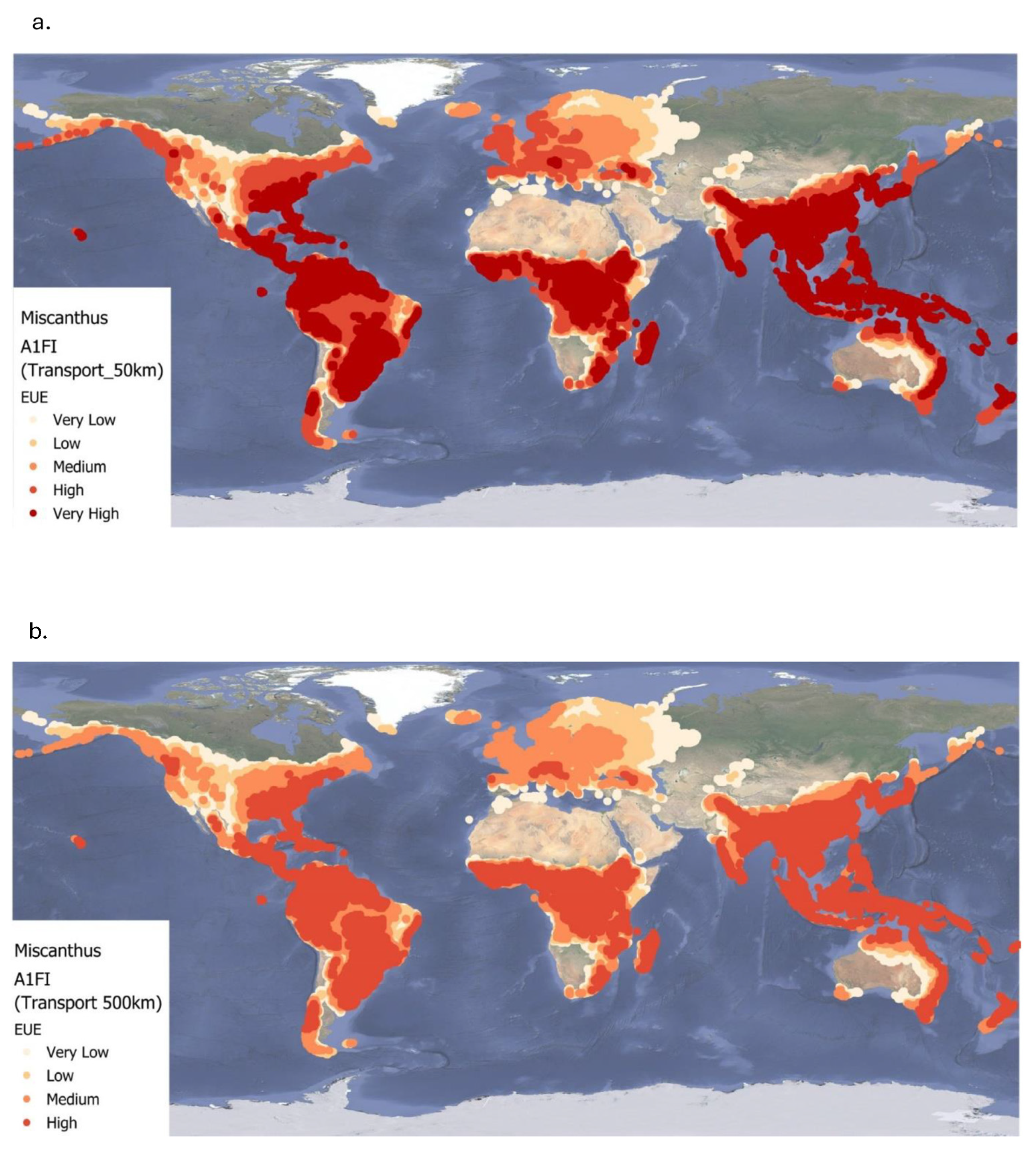
4.3. Impacts of Biomass Transport on the Energy Use Efficiency of Miscanthus
This study provides a detailed assessment of how transport distance affects the EUE of
Miscanthus-based bioenergy systems under current and future climate scenarios. While
Miscanthus is used as the focus crop due to robust existing data [
5], there is a pressing need for similar analyses on other perennials like willow to enable multi-crop comparisons in supply chain planning. Across all climate scenarios, the highest EUE values are observed in tropical and subtropical regions. These areas offer optimal conditions for
Miscanthus growth due to favourable temperatures, sufficient rainfall, and long growing seasons [
2]. However, increasing the biomass transport distance from 50 km to 500 km results in a consistent decline in EUE across all scenarios. This decline is most pronounced in high-yielding regions, reflecting the significant energy cost of moving bulky biomass over long distances [
36,
37].
Under the B1 and A1FI scenario, some high-latitude temperate regions show modest EUE gains due to warmer growing conditions. However, these improvements are often offset by reduced yields in already hot regions due to increased heat stress. Consequently, the global average EUE still declines under A1FI. This underscores the complex interaction between regional climate shifts and crop performance, highlighting the need for site-specific strategies [
2,
28]. Transport represents a major bottleneck to fully realizing the sustainability potential of Miscanthus. Regional EUE losses are especially evident where poor infrastructure and environmental constraints magnify transport inefficiencies, particularly in parts of Africa, Asia, and South America. A key strategy to mitigate these losses is to develop localized biomass supply chains, where production, processing, and utilization occur in proximity. Brazil offers a model example: co-locating sugarcane farms with ethanol and bagasse power plants reduces transport-related energy use and emissions [
38]. Another option is to preprocess biomass into energy-dense forms such as pellets, briquettes, or bio-oils before long-distance transport. While this can improve transport efficiency, the pre-processing itself consumes energy and may reduce overall system efficiency [
5,
37]. Additionally, decentralization of biomass systems brings trade-offs. It may reduce transport distances but require more machinery, labour, and logistics, which could offset some energy gains. Balancing these factors is crucial to designing sustainable systems. Our results suggest that
Miscanthus is most sustainable when cultivated and used locally, especially in tropical and subtropical zones. Long transport distances significantly reduce energy returns, a challenge that persists across all future climate scenarios. Therefore, we suggest that bioenergy policies must prioritize:
Geographically optimized supply chains.
Investment in low-emissions transport infrastructure (e.g., rail and waterways).
Climate-resilient planning that considers yield variability and regional logistics.
Safeguards to avoid unintended land use impacts, particularly the disturbance of high-carbon soils like peatlands, which could release large quantities of CO2 and compromise climate goals.
Thus, realizing the full potential of Miscanthus-based bioenergy systems will require an integrated approach linking agronomic performance, transport efficiency, and sustainable land management under changing climatic and socio-environmental conditions.
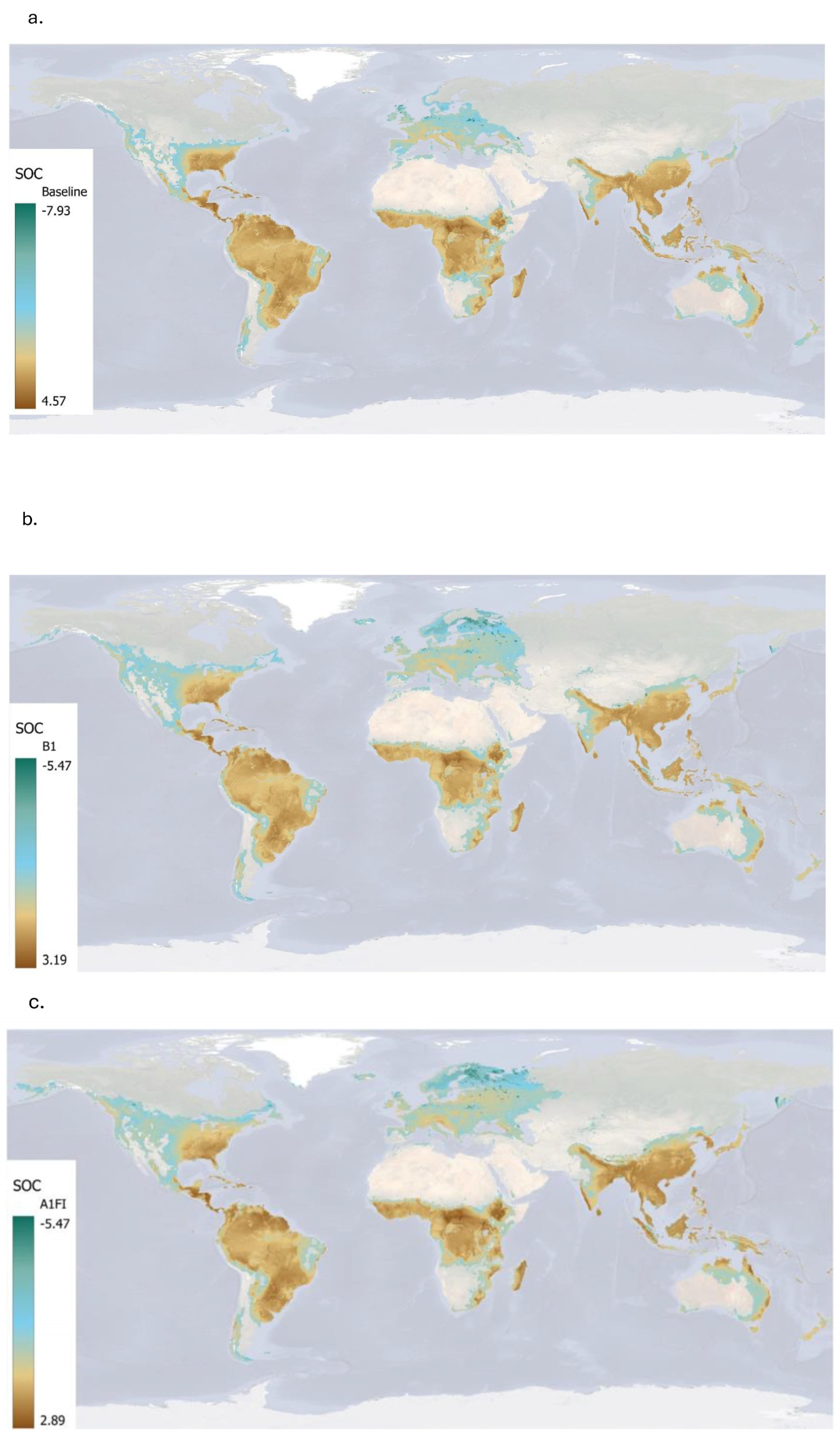
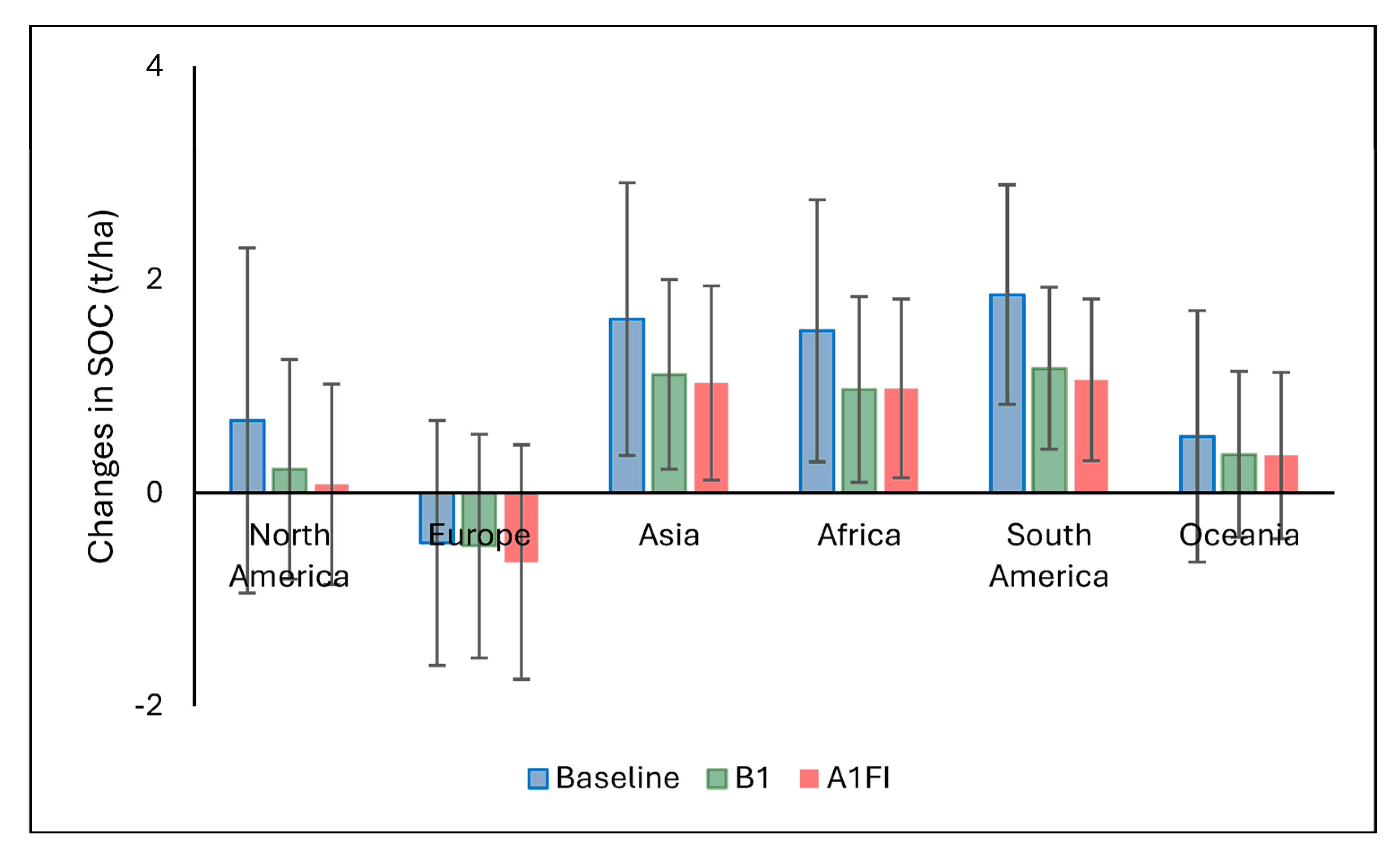
4.4. Estimated Soil Organic Carbon Sequestered by Miscanthus
This study demonstrates that
Miscanthus cultivation has significant potential to enhance SOC sequestration globally, particularly under current climatic conditions. The highest SOC accumulation was observed under the baseline climate (1961–1990), with a global mean annual ∆SOC of 1.20 ± 1.46 t ha
−1, and hotspots reaching up to 4.57 t ha
−1. These results align with previous findings that perennial bioenergy crops can contribute meaningfully to long-term soil carbon storage [
16,
39]. Tropical and subtropical regions in Africa, Southeast Asia, and Latin America emerged as the most promising areas. These regions benefit from warm temperatures and adequate moisture, enhancing plant productivity and litter input, both of which promote SOC accumulation [
40]. However, the analysis also reveals trade-offs. In regions with high initial SOC, especially carbon-rich soils such as histosols or peatlands and/or those with low crop productivity,
Miscanthus cultivation can lead to net SOC losses. This is largely due to disturbance during crop establishment (e.g., ploughing or drainage), which accelerates the decomposition of existing soil carbon. Importantly, high plant productivity does not always guarantee a net SOC gain if losses from disturbed soils are greater than the new inputs from crop biomass.
Under future climate scenarios, both the spatial extent and magnitude of SOC gains decline. The mean ∆SOC drops to 0.62 ± 1.11 t ha
−1 under the B1 scenario and 0.49 ± 1.13 t ha
−1 under A1FI. Warmer temperatures and increased drought risk likely reduce plant productivity and slow carbon accumulation, especially in already vulnerable regions. These results echo earlier studies that warn of diminished mitigation benefits from bioenergy crops under warming scenarios [
41]. Regional patterns further underscore the importance of biophysical conditions. Tropical and subtropical regions maintain some SOC gains due to higher net primary productivity [
42], whereas Europe exhibits consistent SOC losses, possibly due to cooler climates, lower yields, higher initial SOC levels, and large histosol coverage [
43]. These findings affirm that site-specific assessments are critical when evaluating
Miscanthus as a climate mitigation option [
24]. While the crop offers strong potential to sequester carbon, especially under current conditions, its effectiveness diminishes under climate change, highlighting the importance of adaptive land management, such as:
Using drought-tolerant or regionally adapted Miscanthus cultivars.
Applying soil amendments (e.g., compost, biochar) to improve soil structure and moisture retention.
Avoiding cultivation on high-carbon-stock soils like undisturbed peatlands.
Based on our findings, land use strategies should encourage region-specific zoning for energy crops, prioritize the use of marginal or underutilized land, and incorporate sustainability safeguards (e.g., protecting high-carbon or high-biodiversity areas). Policy frameworks should support localized biomass supply chains, promote adaptive land management, and ensure bioenergy development aligns with broader goals for climate mitigation, biodiversity conservation, and food security.
5. Conclusions
This study provides a spatially explicit global assessment of the sustainability of Miscanthus and willow as bioenergy crops under current and future climate scenarios. Miscanthus generally outperforms willow in terms of energy productivity, particularly in tropical and subtropical regions, and shows resilience under moderate climate change. However, extreme climate scenarios significantly reduce its productivity in some regions, underscoring the importance of climate-resilient deployment strategies.
Biomass transport distance strongly influences energy use efficiency, with longer distances reducing returns. This highlights the importance of localized processing and strategic supply chain planning.
Miscanthus also contributes to climate mitigation through SOC sequestration, especially in tropical regions, although future climate change may reduce this benefit in some areas. These findings emphasize the need for region-specific management and adaptation strategies.
Overall, the study demonstrates the potential of Miscanthus as a key component in sustainable bioenergy systems, provided deployment is supported by informed policy, site-specific assessments, and adaptation to future climate risks.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}