
Effect of Cultivated Species and Planting Pattern on Plant Growth, Soil Properties, and Soil Metabolites in a Rain-Fed Orchard in Gansu, China


Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Determination of Plant Growth Characteristics
2.4. Analysis of Soil Chemical Properties and Metabolites
2.5. Data Analysis
3. Results
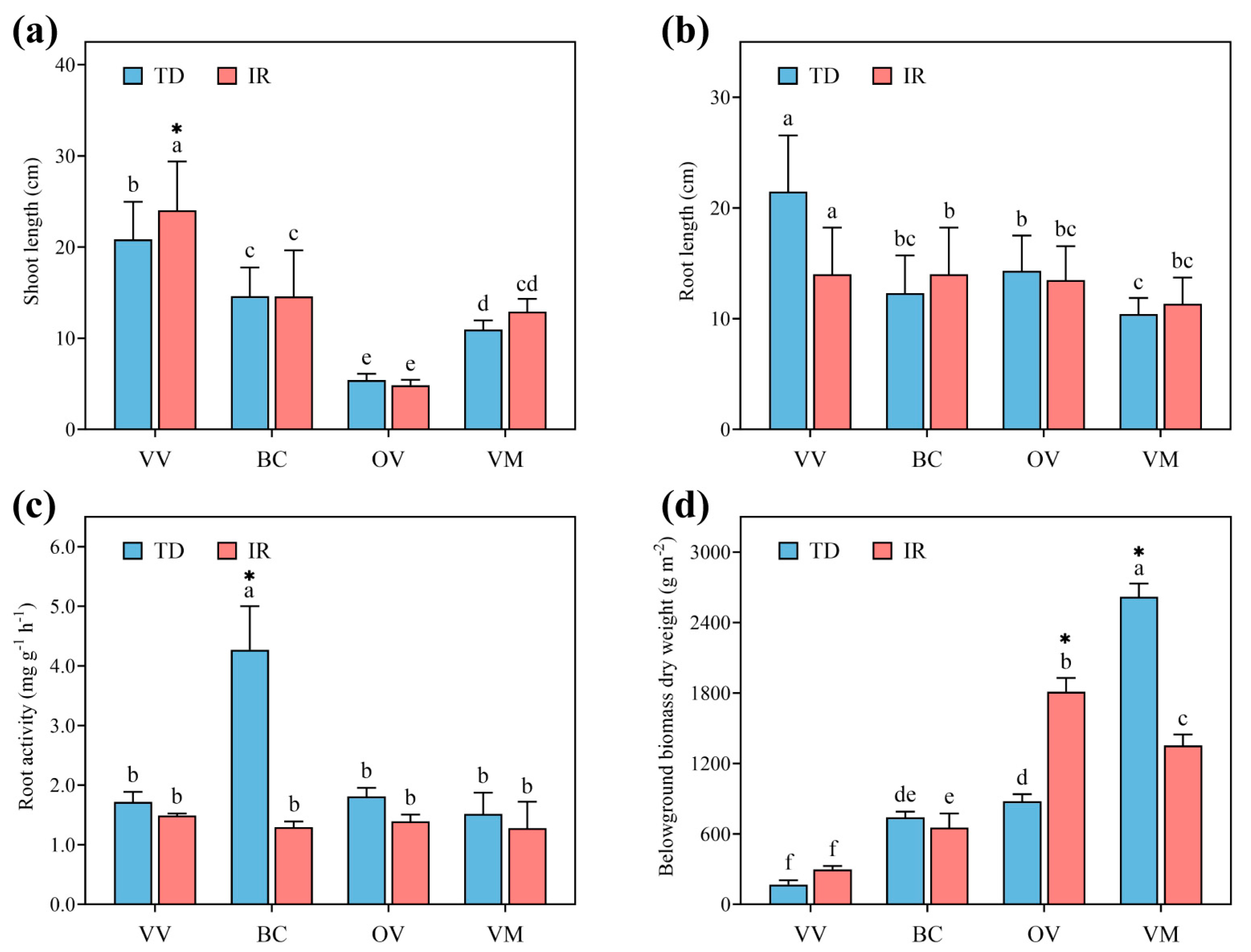
3.1. Orchard Cover Crops Growth
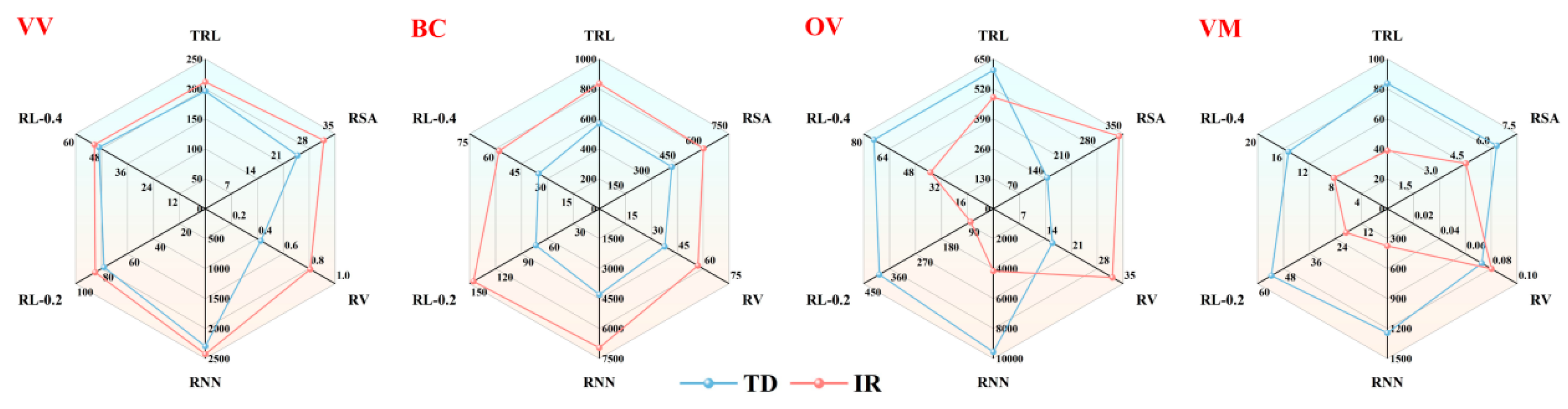
3.2. Root Morphology
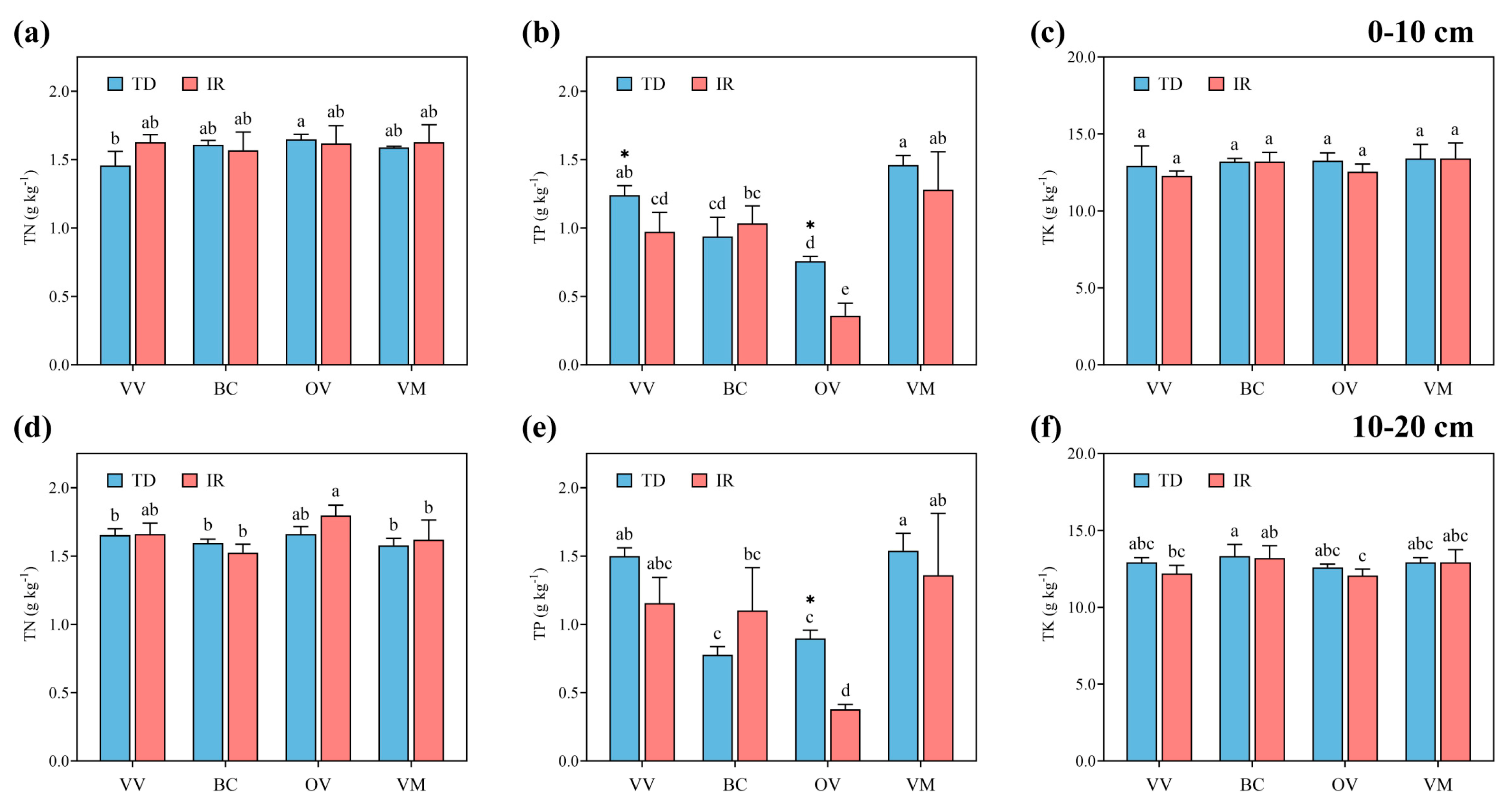
3.3. Soil Chemical Properties
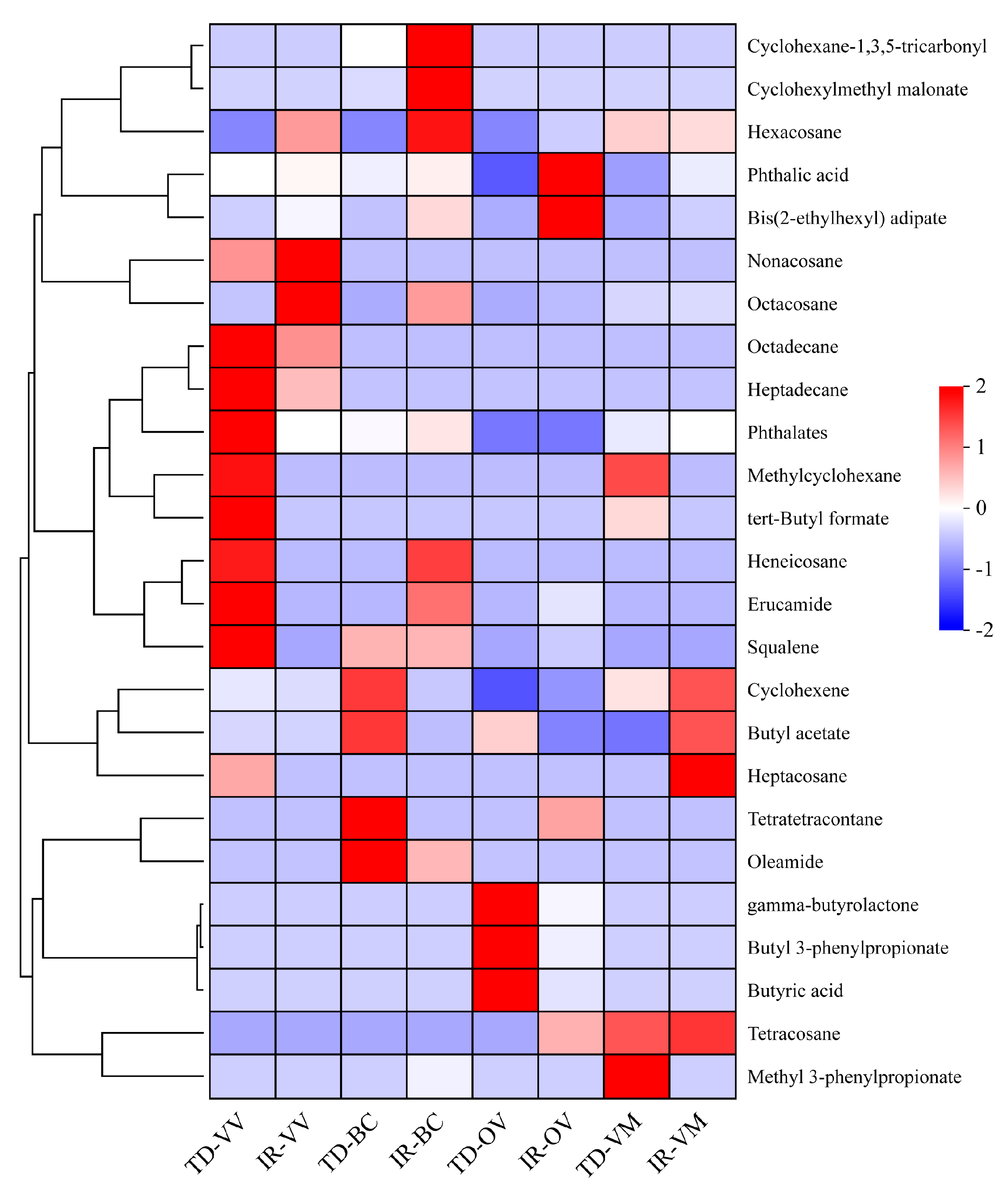
3.4. Rhizosphere Soil Metabolites
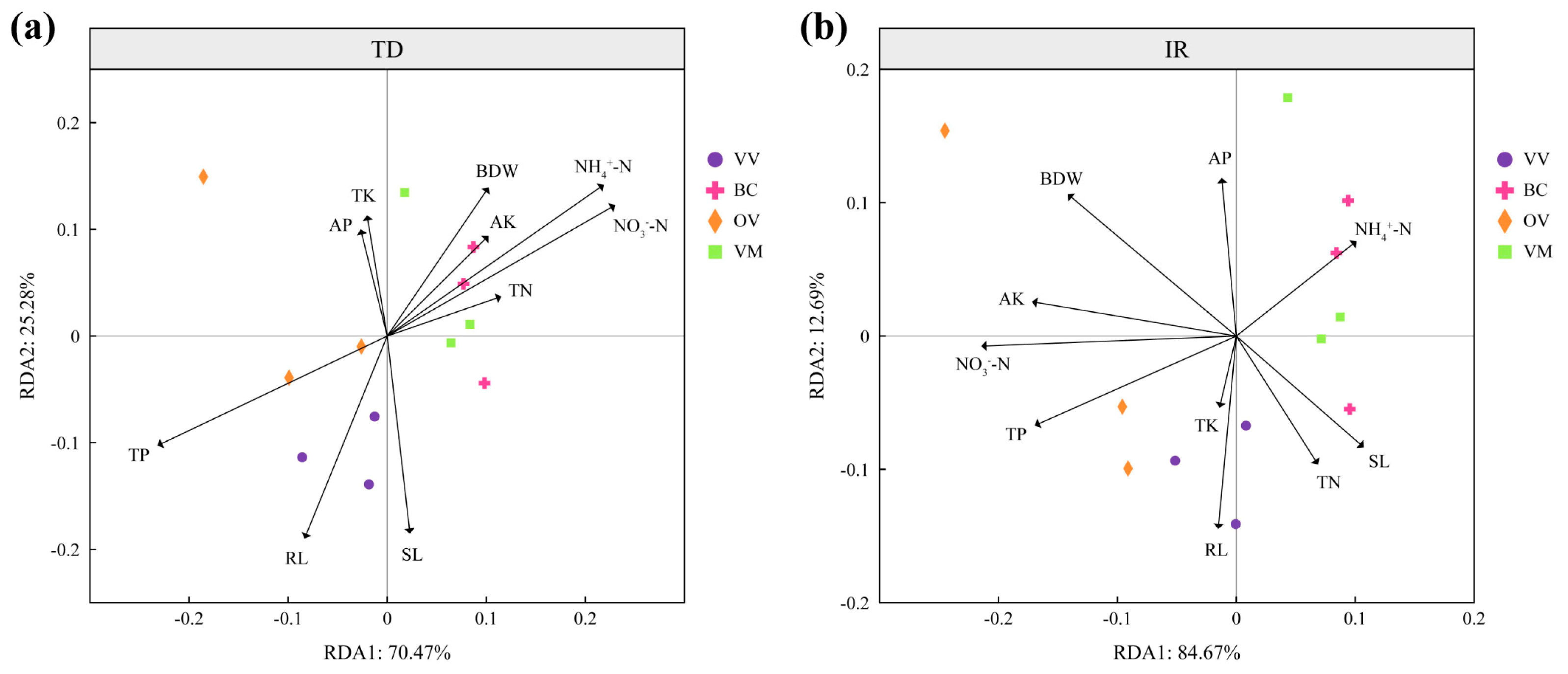
3.5. Redundancy Analysis
3.6. Correlation Analysis
3.7. Comprehensive Analysis of Membership Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| TD | Tree-disk | RL-0.4 | Root length at 0.4 mm diameters |
| IR | Inter-row | TN | Total nitrogen |
| VM | Vulpia myuros | TP | Total phosphorus |
| VV | Vicia villosa | TK | Total potassium |
| OV | Orychophragmus violaceus | AP | Available phosphorus |
| BC | Brassica campestris | AK | Available potassium |
| SL | Shoot length | RDA | Redundancy analysis |
| RL | Root length | TRL | Total root length |
| BDW | Belowground biomass dry weight | RSA | Root surface area |
| RA | Root activity | RV | Root volume |
| RL-0.2 | Root length at 0.2 mm diameters | RNN | Root node number |
References
- Liao, Y. The effects of conservation tillage practices on the soil water-holding capacity of a non-irrigated apple orchard in the Loess Plateau, China. Soil Tillage Res. 2013, 130, 7–12. [Google Scholar] [CrossRef]
- Brennan, E.B.; Acosta-Martinez, V. Cover cropping frequency is the main driver of soil microbial changes during six years of organic vegetable production. Soil Biol. Biochem. 2017, 109, 188–204. [Google Scholar] [CrossRef]
- Verzeaux, J.; Alahmad, A.; Habbib, H.; Nivelle, E.; Roger, D. Cover crops prevent the deleterious effect of nitrogen fertilisation on bacterial diversity by maintaining the carbon content of ploughed soil. Geoderma 2016, 281, 49–57. [Google Scholar] [CrossRef]
- Steenwerth, K.; Belina, K.M. Cover crops enhance soil organic matter, carbon dynamics and microbiological function in a vineyard agroecosystem. Appl. Soil Ecol. 2008, 40, 359–369. [Google Scholar] [CrossRef]
- Xie, B.; Chen, Y.H.; Cheng, C.A.; Ma, R.P.; Zhao, D.Y.; Li, Z.; Li, Y.Q.; An, X.H.; Yang, X.Z. Long-term soil management practices influence the rhizosphere microbial community structure and bacterial function of hilly apple orchard soil. Appl. Soil Ecol. 2022, 180, 104627. [Google Scholar] [CrossRef]
- Ren, J.; Li, F.D.; Yin, C.B. Orchard grass safeguards sustainable development of fruit industry in China. J. Clean. Prod. 2023, 382, 135291. [Google Scholar] [CrossRef]
- Xiang, Y.; Li, Y.; Liu, Y.; Zhang, S.; Yue, X.; Yao, B.; Xue, J.; Lv, W.; Zhang, L.; Xu, X.; et al. Factors shaping soil organic carbon stoCTs in grass covered orchards across China: A meta-analysis. Sci. Total Environ. 2022, 807, 150632. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Wu, Q.S.; Mousavi, S.M.; Hota, D. Integrated soil fertility management in fruit crops: An overview. Int. J. Fruit Sci. 2021, 21, 413–439. [Google Scholar] [CrossRef]
- Wang, Y.J.; Liu, L.; Tian, Y.L.; Wu, X.P.; Yang, J.F.; Luo, Y.; Li, H.K.; Awasthi, M.K.; Zhao, Z.Y. Temporal and spatial variation of soil microorganisms and nutrient under white clover cover. Soil Tillage Res. 2020, 202, 104666. [Google Scholar] [CrossRef]
- Wang, J.W.; Qin, X.Y.; Tan, Y.Q.; Du, Y.Q.; Tdui, Y.; Yang, Y.H.; Ping, X.Y. Impact of grass cover on the soil physicochemical properties in China’s orchards: A meta-analysis. Agroforest. Syst. 2024, 98, 1745–1758. [Google Scholar] [CrossRef]
- Liu, F.T.; Zhang, L.S.; Li, X.W.; Li, B.Z.; Han, M.Y.; Gu, J.; Wang, X.L. Effects of inter-row planting grasses on soil organic carbon fractions and soil microbial community of apple orchard in Weibei dryland. J. Plant Nutr. Fertil. 2014, 20, 355–363. [Google Scholar]
- Cheraghi, M.; Motesharezadeh, B.; Alikhani, H.A.; Mousavi, S.M. Optimal management of plant nutrition in tomato (Lycopersicon esculent Mill) by using biologic, organic and inorganic fertilizers. J. Plant Nutr. 2023, 46, 1560–1579. [Google Scholar] [CrossRef]
- Kovacevic, M.S.; Juric-Kacunic, D.; Libric, L.; Ivos, G. Engineering soil classification according to EN ISO 14688-2:2018. Gradevinar 2018, 70, 873–879. [Google Scholar]
- Niu, G.; Zhong, B.; Wang, R.; Jin, Y.; Wang, Y.; Lü, X.; Jiang, Y.; Huang, J. Effects of nitrogen and water addition on soil carbon, nitrogen, phosphorus, sulfur, and their stoichiometry along soil profile in a semi-arid steppe. J. Soils Sediments 2023, 23, 3298–3309. [Google Scholar] [CrossRef]
- Luster, J.; Göttlein, A.; Nowack, B.; Sarret, G. Sampling, defining, characterising and modeling the rhizosphere-the soil science tool box. Plant Soil 2009, 321, 457–482. [Google Scholar] [CrossRef]
- Daryanto, S.; Fu, B.; Wang, L.; Jacinthe, P.A.; Zhao, W. Quantitative synthesis on the ecosystem services of cover crops. Earth-Sci. Rev. 2018, 185, 357–373. [Google Scholar] [CrossRef]
- Halli, H.M.; Govindasamy, P.; Choudhary, M.; Srinivasan, R.; Prasad, M.; Wasnik, V.K.; Yadav, V.K.; Singh, A.K.; Kumar, S.; Vijay, D.; et al. Range grasses to improve soil properties, carbon sustainability, and fodder security in degraded lands of semi-arid regions. Sci. Total Environ. 2022, 851, 158211. [Google Scholar] [CrossRef]
- Yao, S.; Merwin, I.A.; Bird, G.W.; Abawi, G.S.; Thies, J.E. Orchard floor management practices that maintain vegetative or biomass groundcover stimulate soil microbial activity and alter soil microbial community composition. Plant Soil 2005, 271, 377–389. [Google Scholar] [CrossRef]
- Zhang, W.; Gong, J.; Zhang, S.; Lambers, H.; Dong, X.D.; Hu, Y.X.; Yang, G.S.; Yan, C.Y. Soil phosphorus availability affects niche characteristics of dominant C3 perennial and sub-dominant C4 annual species in a typical temperate grassland of northern China. Plant Soil 2024, 504, 737–761. [Google Scholar] [CrossRef]
- Li, T.; Wang, Y.; Kamran, M.; Chen, X.; Tan, H.; Long, M. Effects of grass inter-planting on soil nutrients, enzyme activity, and bacterial community diversity in an apple orchard. Front. Plant Sci. 2022, 13, 901143. [Google Scholar] [CrossRef]
- Wang, Y.J.; Huang, Q.Q.; Gao, H.; Zhang, R.Q.; Yang, L.; Guo, Y.; Li, H.K.; Awasthi, M.K.; Li, G.C. Long-term cover crops improved soil phosphorus availability in a rain-fed apple orchard. Chemosphere 2021, 275, 130093. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, R.; Fu, L.; Tao, S.; Bao, J. Effects of orchard grass on soil fertility and nutritional status of fruit trees in Korla fragrant pear orchard. Horticulturae 2023, 9, 903. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Ncube, B.; Mulidzi, R.; Lewu, F.B. Management impact and benefit of cover crops on soil quality: A review. Soil Tillage Res. 2020, 204, 104717. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Chomel, M.; Segura, N.A.; de Castro, F.; Goodall, T.; Magilton, M.; Rhymes, J.M.; Delgado-Baquerizo, M.; Griffiths, R.I.; Baggs, E.M.; et al. Land management shapes drought responses of dominant soil microbial taxa across grasslands. Nat. Commun. 2024, 15, 29. [Google Scholar] [CrossRef]
- Scavo, A.; Restuccia, A.; Abbate, C.; Lombardo, S.; Fontanazza, S.; Pandino, G.; Anastasi, U.; Mauromicale, G. Trifolium subterraneum cover cropping enhances soil fertility and weed seedbank dynamics in a Mediterranean apricot orchard. Agron. Sustain. Dev. 2021, 41, 70. [Google Scholar] [CrossRef]
- Dung, T.V.; Ngoc, N.P.; Dang, L.V.; Hung, N.N. Impact of cover crop and mulching on soil physical properties and soil nutrients in a citrus orchard. PeerJ 2022, 10, e14170. [Google Scholar] [CrossRef]
- Samain, E.; Duclercq, J.; Barka, E.A.; Eickermann, M.; Ernenwein, C.; Mazoyon, C.; Sarazin, V.; Dubois, F.; Aussenac, T.; Selim, S. PGPR-soil microbial communities’ interactions and their influence on wheat growth promotion and resistance induction against Mycosphaerella graminicola. Biology 2023, 12, 1416. [Google Scholar] [CrossRef]
- Fan, X.; Ge, A.H.; Qi, S.; Guan, Y.; Wang, R.; Yu, N.; Wang, E. Root exudates and microbial metabolites: Signals and nutrients in plant-microbe interactions. Sci. China Life Sci. 2025. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.N.; Schachtman, D.P. Root exudates impact plant performance under abiotic stress. Trends Plant Sci. 2022, 27, 80–91. [Google Scholar] [CrossRef]
- Guo, X.R.; Song, T.; Deng, L.J.; Zhang, W.F.; Jiao, X.Q. Effects of grass growing on soil fertility and productivity of orchards in China: A meta analysis. Chin. J. Appl. Ecol. 2021, 32, 4021–4028. [Google Scholar]
- Kušliene, G.; Rasmussen, J.; Kuzyakov, Y.; Eriksen, J. Medium-term response of microbial community to rhizodeposits of white clover and ryegrass and tracing of active processes induced by 13C and 15N labelled exudates. Soil Biol. Biochem. 2014, 76, 22–33. [Google Scholar] [CrossRef]
- Chen, N.; Wei, R.; Cao, X.; Duan, X.; Li, H.; Wang, H. Evaluation of inter-row cover crops effects on the microbial diversity during Cabernet Sauvignon (Vitis vinifera L.) maturation. Food Res. Int. 2022, 162, 112113. [Google Scholar] [CrossRef]
- Kelly, C.; Schipanski, M.E.; Tucker, A.; Trujillo, W.; Holman, J.D.; Obour, A.K.; Johnson, S.K.; Brummer, J.E.; Haag, L.; Fonte, S.J. Dryland cover crop soil health benefits are maintained with grazing in the US High and Central Plains. Agric. Ecosyst. Environ. 2021, 313, 107358. [Google Scholar] [CrossRef]
- Seitz, V.A.; Mc Givern, B.B.; Borton, M.A.; Chaparro, J.M.; Schipanski, M.E.; Prenni, J.E.; Wrighton, K.C. Cover crop root exudates impact soil microbiome functional trajectories in agricultural soils. Microbiome 2024, 12, 183. [Google Scholar] [CrossRef]
- Lu, X.P.; Jiang, L.L.; Li, Z.H.; Wu, H.; Ma, Z.Q. Analysis of endogenous hormones in different organs reveals the critical role of phthalanilic acid in the yield and quality of pepper (Capsicum annuum) fruits. Sci. Hortic. 2023, 319, 112148. [Google Scholar] [CrossRef]
- Xue, S.Q.; Li, X.Y.; Fu, Y.R.; Zhu, P.C.; Liu, J.Y.; Kou, W.; Huang, D.L.; Gao, Y.J.; Wang, X.D. New insights into organic carbon mineralization: Combining soil organic carbon fractions, soil bacterial composition, microbial metabolic potential, and soil metabolites. Soil Tillage Res. 2024, 244, 106243. [Google Scholar] [CrossRef]
- Alotaibi, F.; St-Arnaud, M.; Hijri, M. In-depth characterization of plant growth promotion potentials of selected alkanes-degrading plant growth-promoting bacterial isolates. Front. Microbiol. 2022, 13, 863702. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, I.; Wang, J.; Sainju, U.M.; Zhang, S.H.; Zhao, F.Z.; Khan, A. Cover cropping enhances soil microbial biomass and affects microbial community structure: A meta-analysis. Geoderma 2021, 381, 114696. [Google Scholar] [CrossRef]
- Negin, B.; Hen-Avivi, S.; Almekias-Siegl, E.; Shachar, L.; Jander, G.; Aharoni, A. Tree tobacco (Nicotiana glauca) cuticular wax composition is essential for leaf retention during drought, facilitating a speedy recovery following rewatering. New Phytol. 2023, 237, 1574–1589. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patterns | Cultivated Species | TN | TK | TP | AK | AP | NH4+-N | NO3−-N | Score | Ranking |
|---|---|---|---|---|---|---|---|---|---|---|
| TD | VV | 0.25 | 0.49 | 0.75 | 0.05 | 0.11 | 0.02 | 0.04 | 0.24 | 7 |
| IR | VV | 0.50 | 0.19 | 0.53 | 0.10 | 0.05 | 0.03 | 0.02 | 0.20 | 8 |
| TD | BC | 0.53 | 0.49 | 0.35 | 0.10 | 0.03 | 0.63 | 0.26 | 0.34 | 5 |
| IR | BC | 0.68 | 0.22 | 0.03 | 0.13 | 0.05 | 0.06 | 0.97 | 0.31 | 6 |
| TD | OV | 0.33 | 0.59 | 0.84 | 0.14 | 0.52 | 0.07 | 0.01 | 0.36 | 4 |
| IR | OV | 0.44 | 0.59 | 0.71 | 0.69 | 0.45 | 0.79 | 0.24 | 0.56 | 1 |
| TD | VM | 0.38 | 0.64 | 0.38 | 0.58 | 0.91 | 0.52 | 0.31 | 0.53 | 2 |
| IR | VM | 0.22 | 0.61 | 0.53 | 0.37 | 0.84 | 0.05 | 0.05 | 0.38 | 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, Y.; Li, Q.; Shen, Y. Effect of Cultivated Species and Planting Pattern on Plant Growth, Soil Properties, and Soil Metabolites in a Rain-Fed Orchard in Gansu, China. Agronomy 2025, 15, 1385. https://doi.org/10.3390/agronomy15061385
Zou Y, Li Q, Shen Y. Effect of Cultivated Species and Planting Pattern on Plant Growth, Soil Properties, and Soil Metabolites in a Rain-Fed Orchard in Gansu, China. Agronomy. 2025; 15(6):1385. https://doi.org/10.3390/agronomy15061385
Chicago/Turabian StyleZou, Yali, Qi Li, and Yuying Shen. 2025. "Effect of Cultivated Species and Planting Pattern on Plant Growth, Soil Properties, and Soil Metabolites in a Rain-Fed Orchard in Gansu, China" Agronomy 15, no. 6: 1385. https://doi.org/10.3390/agronomy15061385
APA StyleZou, Y., Li, Q., & Shen, Y. (2025). Effect of Cultivated Species and Planting Pattern on Plant Growth, Soil Properties, and Soil Metabolites in a Rain-Fed Orchard in Gansu, China. Agronomy, 15(6), 1385. https://doi.org/10.3390/agronomy15061385