
Identification and Analysis of Endoplasmic-Reticulum-Stress- and Salt-Stress-Related Genes in Solanum tuberosum Genome: StbZIP60 Undergoes Splicing in Response to Salt Stress and ER Stress

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Identification and Property Analysis of Genes Related to ER Stress
2.3. The Promoter Analysis of Candidate Genes
2.4. Tissue Expression and Expression Pattern of Candidate Genes Under Stress Conditions
2.5. Splicing and Subcellular Localization Analysis of StbZIP60
2.6. Statistical Analysis
3. Results
3.1. Identification of ER-Stress-Related Genes in Potato
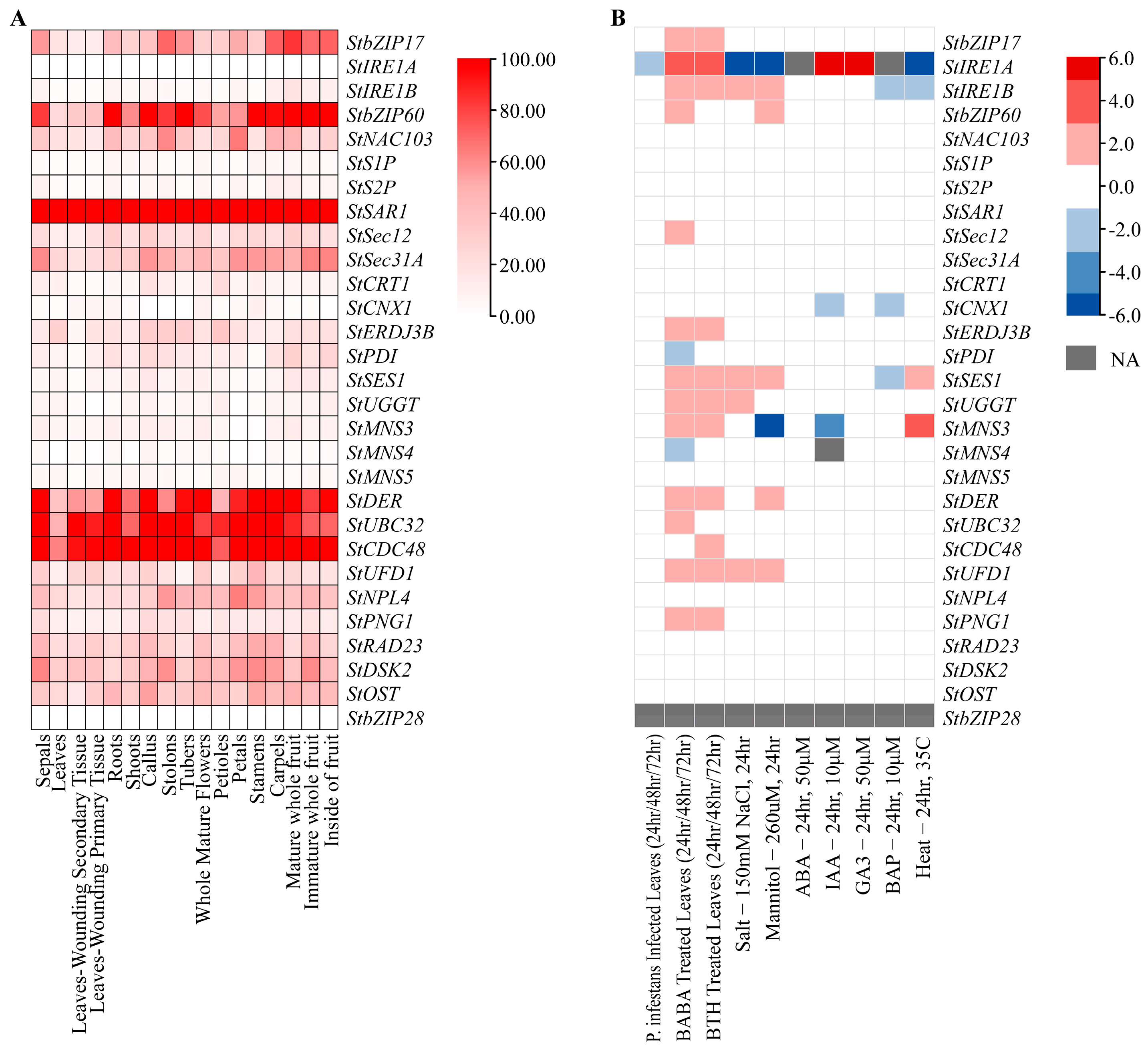
3.2. Expression Profiles of Genes Related to ER Stress in Potato
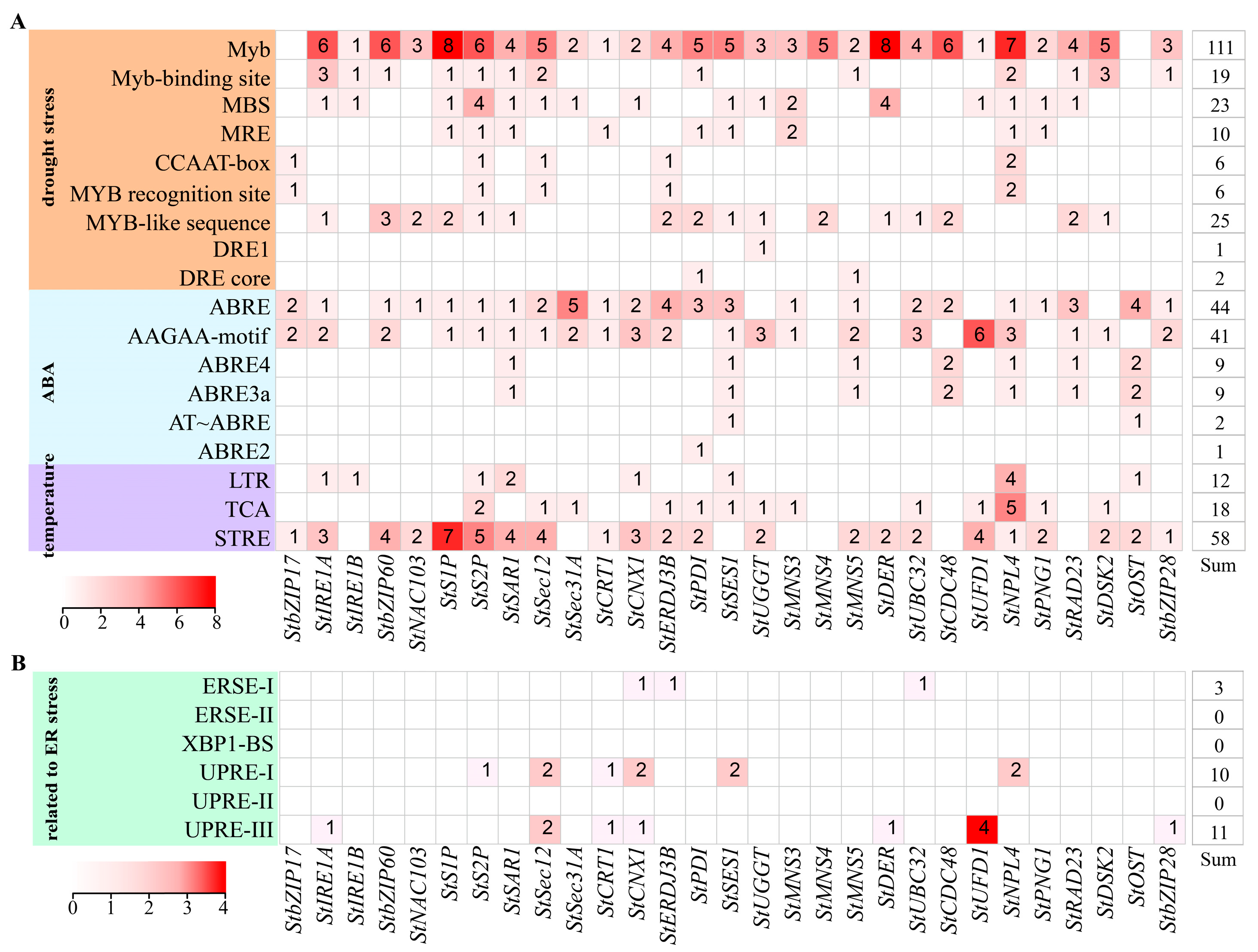
3.3. Promoter Analysis of Genes Related to ER Stress in Potato
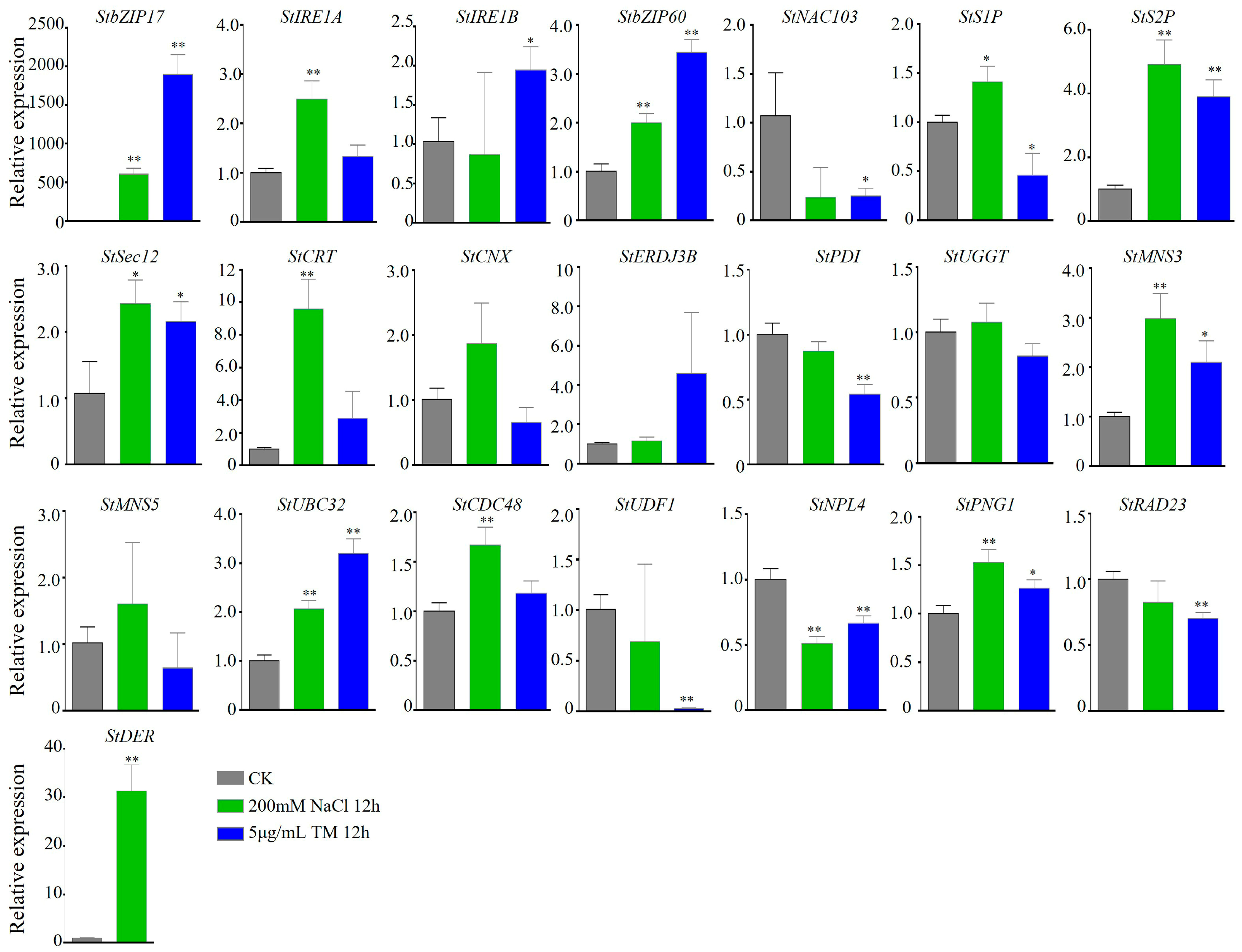
3.4. Detection of the Expression Levels of Genes Under Salt Stress and ER Stress
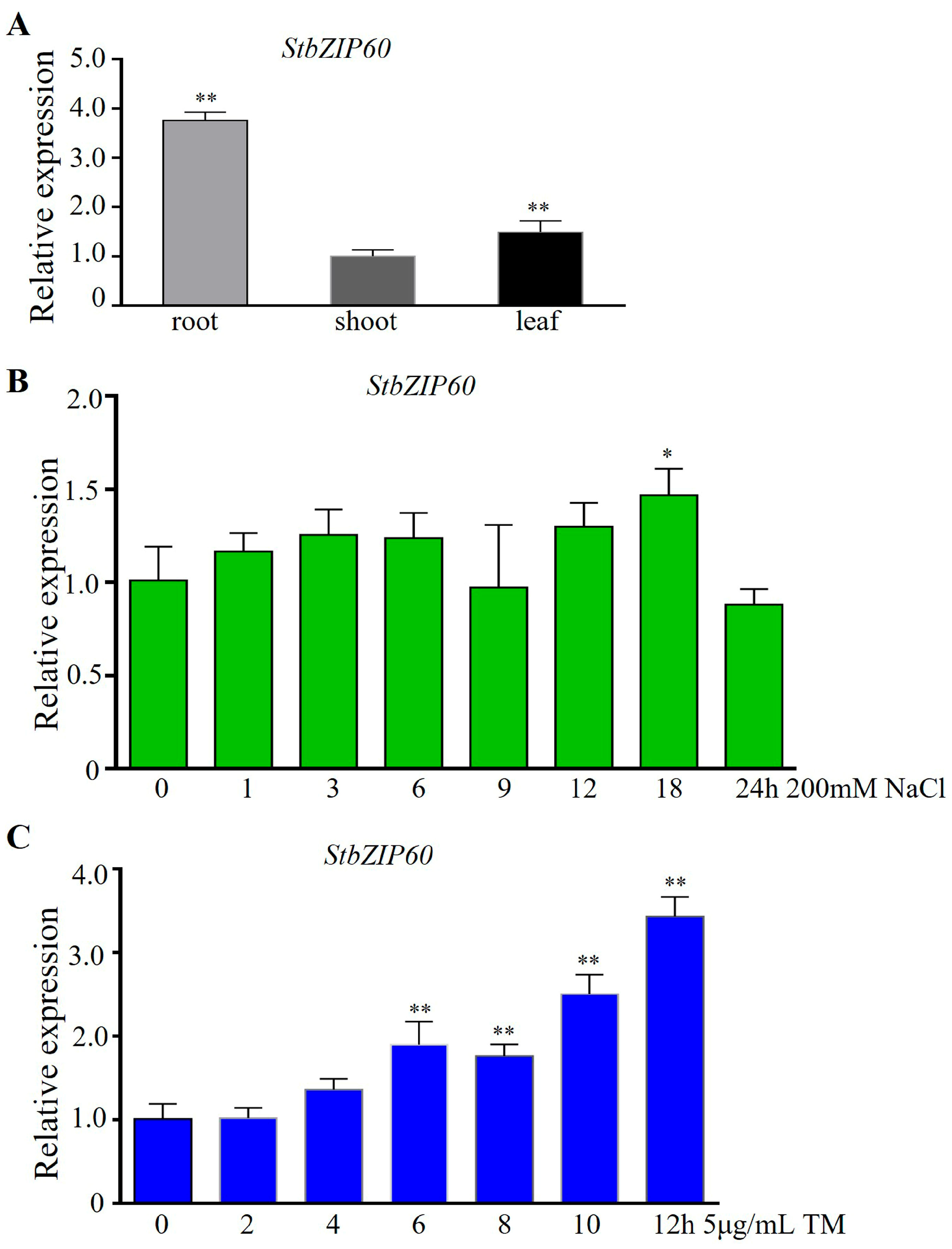
3.5. Analysis of Tissue Expression and Expression Pattern of StbZIP60 Under Salt Stress and ER Stress
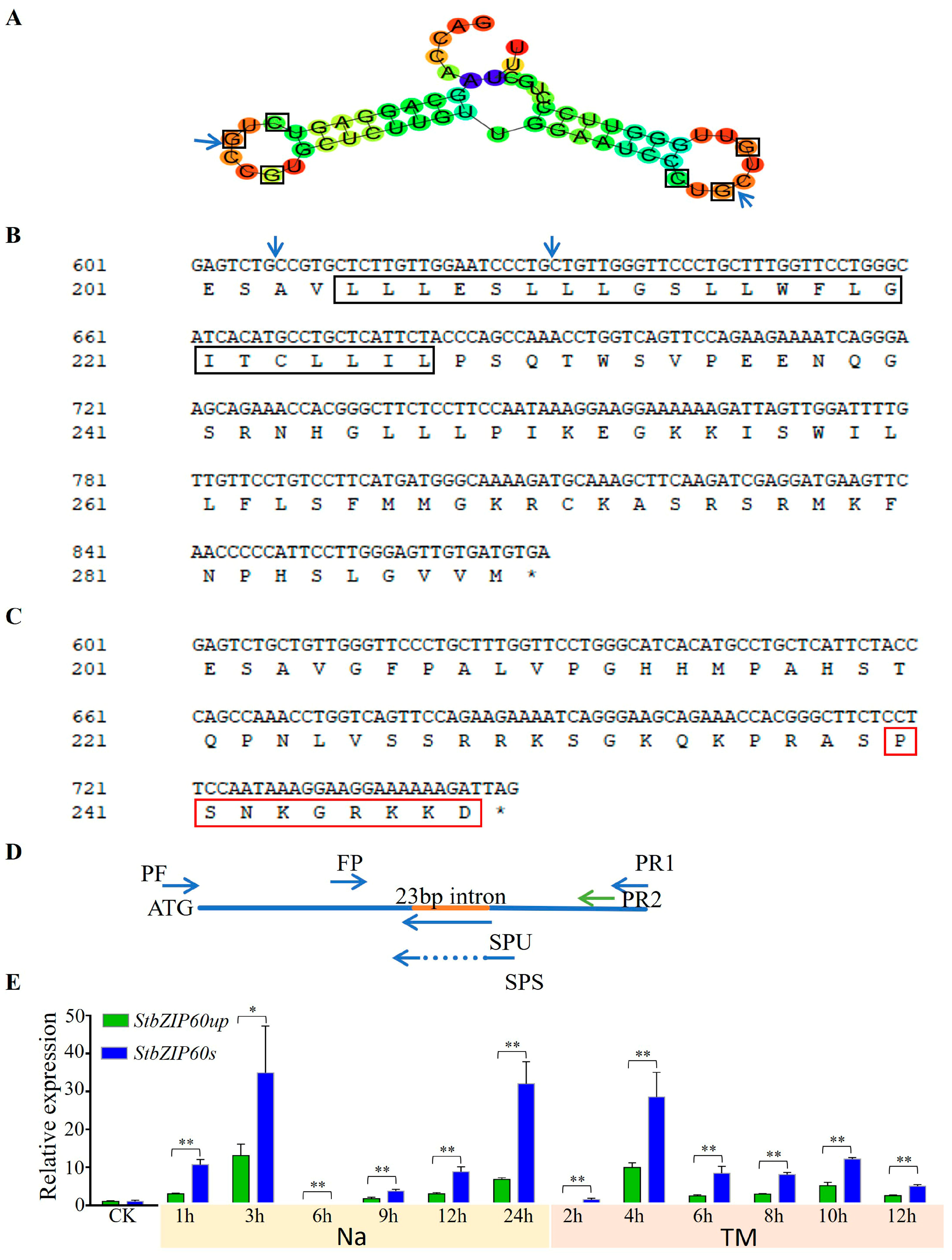
3.6. Unconventional Splicing of StbZIP60 Under Salt Stress and ER Stress in Potato
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ER | endoplasmic reticulum |
| ERQC | ER quality control |
| UPR | unfolded protein response |
| ERAD | endoplasmic-reticulum-associated degradation |
| TM | tunicamycin |
| ERSE | ER stress responsive element |
| UPRE | UPR element |
References
- Reyes-Impellizzeri, S.; Moreno, A.A. The Endoplasmic Reticulum Role in the Plant Response to Abiotic Stress. Front. Plant Sci. 2021, 12, 755447. [Google Scholar] [CrossRef] [PubMed]
- Strasser, R. Protein Quality Control in the Endoplasmic Reticulum of Plants. Annu. Rev. Plant Biol. 2018, 69, 147–172. [Google Scholar] [CrossRef] [PubMed]
- Angelos, E.; Ruberti, C.; Kim, S.J.; Brandizzi, F. Maintaining the factory: The roles of the unfolded protein response in cellular homeostasis in plants. Plant J. 2017, 90, 671–682. [Google Scholar] [CrossRef]
- Tintor, N.; Saijo, Y. ER-mediated control for abundance, quality, and signaling of transmembrane immune receptors in plants. Front. Plant Sci. 2014, 5, 65. [Google Scholar] [CrossRef]
- Ko, D.K.; Brandizzi, F. Dynamics of ER stress-induced gene regulation in plants. Nat. Rev. Genet. 2024, 25, 513–525. [Google Scholar] [CrossRef]
- Liu, Y.; Lv, Y.; Wei, A.; Guo, M.; Li, Y.; Wang, J.; Wang, X.; Bao, Y. Unfolded protein response in balancing plant growth and stress tolerance. Front. Plant Sci. 2022, 13, 1019414. [Google Scholar] [CrossRef] [PubMed]
- Christianson, J.C.; Jarosch, E.; Sommer, T. Mechanisms of substrate processing during ER-associated protein degradation. Nat. Rev. Mol. Cell Biol. 2023, 24, 777–796. [Google Scholar] [CrossRef]
- Chen, Q.; Yu, F.; Xie, Q. Insights into endoplasmic reticulum-associated degradation in plants. New Phytol. 2020, 226, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Nakatogawa, H. Autophagic degradation of the endoplasmic reticulum. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2020, 96, 1–9. [Google Scholar] [CrossRef]
- Walter, P.; Ron, D. The unfolded protein response: From stress pathway to homeostatic regulation. Science 2011, 334, 1081–1086. [Google Scholar] [CrossRef]
- Simoni, E.B.; Oliveira, C.C.; Fraga, O.T.; Reis, P.A.B.; Fontes, E.P.B. Cell Death Signaling from Endoplasmic Reticulum Stress: Plant-Specific and Conserved Features. Front. Plant Sci. 2022, 13, 835738. [Google Scholar] [CrossRef]
- Nagashima, Y.; Mishiba, K.; Suzuki, E.; Shimada, Y.; Iwata, Y.; Koizumi, N. Arabidopsis IRE1 catalyses unconventional splicing of bZIP60 mRNA to produce the active transcription factor. Sci. Rep. 2011, 1, 29. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Humbert, S.; Liu, J.X.; Srivastava, R.; Rothstein, S.J.; Howell, S.H. Heat induces the splicing by IRE1 of a mRNA encoding a transcription factor involved in the unfolded protein response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 7247–7252. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.P.; Dey, M.; Neculai, D.; Cao, C.; Dever, T.E.; Sicheri, F. Structure of the dual enzyme Ire1 reveals the basis for catalysis and regulation in nonconventional RNA splicing. Cell 2008, 132, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Iwata, Y.; Lee, M.H.; Koizumi, N. Analysis of a transcription factor using transient assay in Arabidopsis protoplasts. Methods Mol. Biol. 2011, 754, 107–117. [Google Scholar] [CrossRef]
- Li, Z.; Tang, J.; Srivastava, R.; Bassham, D.C.; Howell, S.H. The Transcription Factor bZIP60 Links the Unfolded Protein Response to the Heat Stress Response in Maize. Plant Cell 2020, 32, 3559–3575. [Google Scholar] [CrossRef]
- Iwata, Y.; Koizumi, N. An Arabidopsis transcription factor, AtbZIP60, regulates the endoplasmic reticulum stress response in a manner unique to plants. Proc. Natl. Acad. Sci. USA 2005, 102, 5280–5285. [Google Scholar] [CrossRef]
- Cho, Y. Arabidopsis AGB1 participates in salinity response through bZIP17-mediated unfolded protein response. BMC Plant Biol. 2024, 24, 586. [Google Scholar] [CrossRef]
- Liu, J.X.; Srivastava, R.; Che, P.; Howell, S.H. Salt stress responses in Arabidopsis utilize a signal transduction pathway related to endoplasmic reticulum stress signaling. Plant J. 2007, 51, 897–909. [Google Scholar] [CrossRef]
- Srivastava, R.; Deng, Y.; Howell, S.H. Stress sensing in plants by an ER stress sensor/transducer, bZIP28. Front. Plant Sci. 2014, 5, 59. [Google Scholar] [CrossRef]
- Che, P.; Bussell, J.D.; Zhou, W.; Estavillo, G.M.; Pogson, B.J.; Smith, S.M. Signaling from the endoplasmic reticulum activates brassinosteroid signaling and promotes acclimation to stress in Arabidopsis. Sci. Signal. 2010, 3, ra69. [Google Scholar] [CrossRef]
- Sun, L.; Zhang, S.S.; Lu, S.J.; Liu, J.X. Site-1 protease cleavage site is important for the ER stress-induced activation of membrane-associated transcription factor bZIP28 in Arabidopsis. Sci. China Life Sci. 2015, 58, 270–275. [Google Scholar] [CrossRef]
- Liu, J.X.; Howell, S.H. bZIP28 and NF-Y transcription factors are activated by ER stress and assemble into a transcriptional complex to regulate stress response genes in Arabidopsis. Plant Cell 2010, 22, 782–796. [Google Scholar] [CrossRef]
- Manghwar, H.; Li, J. Endoplasmic Reticulum Stress and Unfolded Protein Response Signaling in Plants. Int. J. Mol. Sci. 2022, 23, 828. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Guo, C.; Ali, K.; Zheng, Q.; Wei, Q.; Zhu, Y.; Wang, L.; Li, G.; Li, W.; Zheng, B.; et al. A Non-redundant Function of MNS5: A Class I alpha-1, 2 Mannosidase, in the Regulation of Endoplasmic Reticulum-Associated Degradation of Misfolded Glycoproteins. Front. Plant Sci. 2022, 13, 873688. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Duan, M.; Shan, L.; Zheng, L.; Liu, J. HIP is involved in NaCl and endoplasmic reticulum stress resistance in Arabidopsis. Plant Physiol. Biochem. 2024, 217, 109226. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Wang, J.; Li, H.; Xie, C.; Zhang, S.; Wu, C.; Yang, G.; Yan, K.; Huang, J.; Zheng, C. SENSITIVE TO SALT1, An Endoplasmic Reticulum-Localized Chaperone, Positively Regulates Salt Resistance. Plant Physiol. 2018, 178, 1390–1405. [Google Scholar] [CrossRef]
- Jin, T.; Shan, Z.; Zhou, S.; Yang, Q.; Gai, J.; Li, Y. GmDNAJC7 from Soybean Is Involved in Plant Tolerance to Alkaline-Salt, Salt, and Drought Stresses. Agronomy 2022, 12, 1419. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 1 January 2022).
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef]
- Butcher, K.; Wick, A.F.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil Salinity: A Threat to Global Food Security. Agron. J. 2016, 108, 2189–2200. [Google Scholar] [CrossRef]
- Herath, V.; Verchot, J. Comprehensive Transcriptome Analysis Reveals Genome-Wide Changes Associated with Endoplasmic Reticulum (ER) Stress in Potato (Solanum tuberosum L.). Int. J. Mol. Sci. 2022, 23, 13795. [Google Scholar] [CrossRef] [PubMed]
- Chourasia, K.N.; Lal, M.K.; Tiwari, R.K.; Dev, D.; Kardile, H.B.; Patil, V.U.; Kumar, A.; Vanishree, G.; Kumar, D.; Bhardwaj, V.; et al. Salinity Stress in Potato: Understanding Physiological, Biochemical and Molecular Responses. Life 2021, 11, 545. [Google Scholar] [CrossRef]
- Han, X.; Yang, R.; Zhang, L.; Wei, Q.; Zhang, Y.; Wang, Y.; Shi, Y. A Review of Potato Salt Tolerance. Int. J. Mol. Sci. 2023, 24, 10726. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Demircan, N.; Ozgur, R.; Turkan, I.; Uzilday, B. Heavy metal toxicity leads to accumulation of insoluble proteins and induces endoplasmic reticulum stress-specific unfolded protein response in Arabidopsis thaliana. Environ. Sci. Pollut. Res. Int. 2024, 31, 53206–53218. [Google Scholar] [CrossRef]
- Gao, J.; Wang, M.J.; Wang, J.J.; Lu, H.P.; Liu, J.X. bZIP17 regulates heat stress tolerance at reproductive stage in Arabidopsis. aBIOTECH 2022, 3, 1–11. [Google Scholar] [CrossRef]
- Kim, J.S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. ER-Anchored Transcription Factors bZIP17 and bZIP28 Regulate Root Elongation. Plant Physiol. 2018, 176, 2221–2230. [Google Scholar] [CrossRef]
- Gao, H.; Brandizzi, F.; Benning, C.; Larkin, R.M. A membrane-tethered transcription factor defines a branch of the heat stress response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 16398–16403. [Google Scholar] [CrossRef]
- Hong, Z.H.; Qing, T.; Schubert, D.; Kleinmanns, J.A.; Liu, J.X. BLISTER-regulated vegetative growth is dependent on the protein kinase domain of ER stress modulator IRE1A in Arabidopsis thaliana. PLoS Genet. 2019, 15, e1008563. [Google Scholar] [CrossRef]
- Gaguancela, O.A.; Zuniga, L.P.; Arias, A.V.; Halterman, D.; Flores, F.J.; Johansen, I.E.; Wang, A.; Yamaji, Y.; Verchot, J. The IRE1/bZIP60 Pathway and Bax Inhibitor 1 Suppress Systemic Accumulation of Potyviruses and Potexviruses in Arabidopsis and Nicotiana benthamiana Plants. Mol. Plant Microbe Interact. 2016, 29, 750–766. [Google Scholar] [CrossRef]
- Moreno, A.A.; Mukhtar, M.S.; Blanco, F.; Boatwright, J.L.; Moreno, I.; Jordan, M.R.; Chen, Y.; Brandizzi, F.; Dong, X.; Orellana, A.; et al. IRE1/bZIP60-mediated unfolded protein response plays distinct roles in plant immunity and abiotic stress responses. PLoS ONE 2012, 7, e31944. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Du, H.; Zhang, Z.; Xu, W.; Deng, X. BhbZIP60 from Resurrection Plant Boea hygrometrica is an mRNA Splicing-Activated Endoplasmic Reticulum Stress Regulator Involved in Drought Tolerance. Front. Plant Sci. 2017, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Henriquez-Valencia, C.; Moreno, A.A.; Sandoval-Ibanez, O.; Mitina, I.; Blanco-Herrera, F.; Cifuentes-Esquivel, N.; Orellana, A. bZIP17 and bZIP60 Regulate the Expression of BiP3 and Other Salt Stress Responsive Genes in an UPR-Independent Manner in Arabidopsis thaliana. J. Cell Biochem. 2015, 116, 1638–1645. [Google Scholar] [CrossRef]
- Fujita, M.; Mizukado, S.; Fujita, Y.; Ichikawa, T.; Nakazawa, M.; Seki, M.; Matsui, M.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Identification of stress-tolerance-related transcription-factor genes via mini-scale Full-length cDNA Over-eXpressor (FOX) gene hunting system. Biochem. Biophys. Res. Commun. 2007, 364, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, L.P.; Wang, Y.Z.; Yang, L.; Wang, M.J.; Liu, J.X. NAC103, a NAC family transcription factor, regulates ABA response during seed germination and seedling growth in Arabidopsis. Planta 2020, 252, 95. [Google Scholar] [CrossRef]
- Sun, L.; Yang, Z.T.; Song, Z.T.; Wang, M.J.; Sun, L.; Lu, S.J.; Liu, J.X. The plant-specific transcription factor gene NAC103 is induced by bZIP60 through a new cis-regulatory element to modulate the unfolded protein response in Arabidopsis. Plant J. 2013, 76, 274–286. [Google Scholar] [CrossRef]
- Iwata, Y.; Ashida, M.; Hasegawa, C.; Tabara, K.; Mishiba, K.I.; Koizumi, N. Activation of the Arabidopsis membrane-bound transcription factor bZIP28 is mediated by site-2 protease, but not site-1 protease. Plant J. 2017, 91, 408–415. [Google Scholar] [CrossRef]
- Srivastava, R.; Chen, Y.; Deng, Y.; Brandizzi, F.; Howell, S.H. Elements proximal to and within the transmembrane domain mediate the organelle-to-organelle movement of bZIP28 under ER stress conditions. Plant J. 2012, 70, 1033–1042. [Google Scholar] [CrossRef]
- Pastor-Cantizano, N.; Bernat-Silvestre, C.; Marcote, M.J.; Aniento, F. Loss of Arabidopsis p24 function affects ERD2 trafficking and Golgi structure, and activates the unfolded protein response. J. Cell Sci. 2018, 131, jcs203802. [Google Scholar] [CrossRef]
- Gimeno-Ferrer, F.; Pastor-Cantizano, N.; Bernat-Silvestre, C.; Selvi-Martinez, P.; Vera-Sirera, F.; Gao, C.; Perez-Amador, M.A.; Jiang, L.; Aniento, F.; Marcote, M.J. alpha2-COP is involved in early secretory traffic in Arabidopsis and is required for plant growth. J. Exp. Bot. 2017, 68, 391–401. [Google Scholar] [CrossRef]
- Joshi, R.; Paul, M.; Kumar, A.; Pandey, D. Role of calreticulin in biotic and abiotic stress signalling and tolerance mechanisms in plants. Gene 2019, 714, 144004. [Google Scholar] [CrossRef]
- Kim, J.H.; Nguyen, N.H.; Nguyen, N.T.; Hong, S.W.; Lee, H. Loss of all three calreticulins, CRT1, CRT2 and CRT3, causes enhanced sensitivity to water stress in Arabidopsis. Plant Cell Rep. 2013, 32, 1843–1853. [Google Scholar] [CrossRef] [PubMed]
- Montpetit, J.; Clua, J.; Hsieh, Y.F.; Vogiatzaki, E.; Muller, J.; Abel, S.; Strasser, R.; Poirier, Y. Endoplasmic reticulum calnexins participate in the primary root growth response to phosphate deficiency. Plant Physiol. 2023, 191, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Qu, S.; Chowdhury, S.; Noxon, I.C.; Schonhoft, J.D.; Plate, L.; Powers, E.T.; Kelly, J.W.; Lander, G.C.; Wiseman, R.L. The endoplasmic reticulum HSP40 co-chaperone ERdj3/DNAJB11 assembles and functions as a tetramer. EMBO J. 2017, 36, 2296–2309. [Google Scholar] [CrossRef]
- Lu, D.P.; Christopher, D.A. Endoplasmic reticulum stress activates the expression of a sub-group of protein disulfide isomerase genes and AtbZIP60 modulates the response in Arabidopsis thaliana. Mol. Genet. Genom. 2008, 280, 199–210. [Google Scholar] [CrossRef]
- Guan, P.; Xie, C.; Zhao, D.; Wang, L.; Zheng, C. SES1 is vital for seedling establishment and post-germination growth under high-potassium stress conditions in Arabidopsis thaliana. PeerJ 2022, 10, e14282. [Google Scholar] [CrossRef]
- Guan, P.; Wang, J.; Xie, C.; Wu, C.; Yang, G.; Yan, K.; Zhang, S.; Zheng, C.; Huang, J. SES1 positively regulates heat stress resistance in Arabidopsis. Biochem. Biophys. Res. Commun. 2019, 513, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Herrera, F.; Moreno, A.A.; Tapia, R.; Reyes, F.; Araya, M.; D’Alessio, C.; Parodi, A.; Orellana, A. The UDP-glucose: Glycoprotein glucosyltransferase (UGGT), a key enzyme in ER quality control, plays a significant role in plant growth as well as biotic and abiotic stress in Arabidopsis thaliana. BMC Plant Biol. 2015, 15, 127. [Google Scholar] [CrossRef]
- Farid, A.; Malinovsky, F.G.; Veit, C.; Schoberer, J.; Zipfel, C.; Strasser, R. Specialized roles of the conserved subunit OST3/6 of the oligosaccharyltransferase complex in innate immunity and tolerance to abiotic stresses. Plant Physiol. 2013, 162, 24–38. [Google Scholar] [CrossRef]
- Xia, T.; Zhan, Y.; Mu, Y.; Zhang, J.; Xu, W. MNSs-mediated N-glycan processing is essential for auxin homeostasis in Arabidopsis roots during alkaline response. iScience 2022, 25, 104298. [Google Scholar] [CrossRef]
- Huttner, S.; Veit, C.; Vavra, U.; Schoberer, J.; Liebminger, E.; Maresch, D.; Grass, J.; Altmann, F.; Mach, L.; Strasser, R. Arabidopsis Class I alpha-Mannosidases MNS4 and MNS5 Are Involved in Endoplasmic Reticulum-Associated Degradation of Misfolded Glycoproteins. Plant Cell 2014, 26, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Kamauchi, S.; Nakatani, H.; Nakano, C.; Urade, R. Gene expression in response to endoplasmic reticulum stress in Arabidopsis thaliana. FEBS J. 2005, 272, 3461–3476. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, R.; Wu, Y.; Wei, S.; Wang, Q.; Zheng, Y.; Xia, R.; Shang, X.; Yu, F.; Yang, X.; et al. ERAD-related E2 and E3 enzymes modulate the drought response by regulating the stability of PIP2 aquaporins. Plant Cell 2021, 33, 2883–2898. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Liu, L.; Zhao, Q.; Zhang, Z.; Li, Q.; Lin, B.; Wu, Y.; Tang, S.; Xie, Q. Arabidopsis ubiquitin conjugase UBC32 is an ERAD component that functions in brassinosteroid-mediated salt stress tolerance. Plant Cell 2012, 24, 233–244. [Google Scholar] [CrossRef]
- Ines, D.; Courty, P.E.; Wendehenne, D.; Rosnoblet, C. CDC48 in plants and its emerging function in plant immunity. Trends Plant Sci. 2024, 29, 786–798. [Google Scholar] [CrossRef]
- Wei, L.; Cheng, J.; Xiang, J.; Wu, J. Genome-wide identification and characterization of grapevine UFD1 genes during berry development and salt stress response. J. Plant Biochem. Biotechnol. 2022, 31, 592–601. [Google Scholar] [CrossRef]
- Bays, N.W.; Wilhovsky, S.K.; Goradia, A.; Hodgkiss-Harlow, K.; Hampton, R.Y. HRD4/NPL4 is required for the proteasomal processing of ubiquitinated ER proteins. Mol. Biol. Cell 2001, 12, 4114–4128. [Google Scholar] [CrossRef]
- Masahara-Negishi, Y.; Hosomi, A.; Della Mea, M.; Serafini-Fracassini, D.; Suzuki, T. A plant peptide: N-glycanase orthologue facilitates glycoprotein ER-associated degradation in yeast. Biochim. Biophys. Acta 2012, 1820, 1457–1462. [Google Scholar] [CrossRef]
- Raasi, S.; Wolf, D.H. Ubiquitin receptors and ERAD: A network of pathways to the proteasome. Semin. Cell Dev. Biol. 2007, 18, 780–791. [Google Scholar] [CrossRef]
- Kaur, N.; Kaitheri Kandoth, P. Tomato bZIP60 mRNA undergoes splicing in endoplasmic reticulum stress and in response to environmental stresses. Plant Physiol. Biochem. 2021, 160, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Humbert, S.; Howell, S.H. ZmbZIP60 mRNA is spliced in maize in response to ER stress. BMC Res. Notes 2012, 5, 144. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.A.; Silva, J.C.; Caetano, H.D.; Machado, J.P.; Mendes, G.C.; Reis, P.A.; Brustolini, O.J.; Dal-Bianco, M.; Fontes, E.P. Comprehensive analysis of the endoplasmic reticulum stress response in the soybean genome: Conserved and plant-specific features. BMC Genom. 2015, 16, 783. [Google Scholar] [CrossRef]
- Bi, W.; Chen, Y.; Song, Y.; Liu, J.; Zhao, T.; Sun, C.; Qin, J.; Tu, Z.; Li, Y.; Wang, X.; et al. Potato DMP2 positively regulates plant immunity by modulating endoplasmic reticulum homeostasis. J. Integr. Plant Biol. 2025. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Liu, Y.; Cai, H.; Guo, M.; Chai, M.; She, Z.; Ye, L.; Cheng, Y.; Wang, B.; Qin, Y. The bZIP Transcription Factor GmbZIP15 Negatively Regulates Salt- and Drought-Stress Responses in Soybean. Int. J. Mol. Sci. 2020, 21, 7778. [Google Scholar] [CrossRef]
- Hooks, K.B.; Griffiths-Jones, S. Conserved RNA structures in the non-canonical Hac1/Xbp1 intron. RNA Biol. 2011, 8, 552–556. [Google Scholar] [CrossRef]
- Howell, S.H. Evolution of the unfolded protein response in plants. Plant Cell Environ. 2021, 44, 2625–2635. [Google Scholar] [CrossRef]
- Alhabsi, A.; Ling, Y.; Crespi, M.; Reddy, A.S.N.; Mahfouz, M. Alternative Splicing Dynamics in Plant Adaptive Responses to Stress. Annu. Rev. Plant Biol. 2025. Online ahead of print. [Google Scholar] [CrossRef]
- Li, Z.; Srivastava, R.; Tang, J.; Zheng, Z.; Howell, S.H. Cis-Effects Condition the Induction of a Major Unfolded Protein Response Factor, ZmbZIP60, in Response to Heat Stress in Maize. Front. Plant Sci. 2018, 9, 833. [Google Scholar] [CrossRef]
- Geng, X.; Zang, X.; Li, H.; Liu, Z.; Zhao, A.; Liu, J.; Peng, H.; Yao, Y.; Hu, Z.; Ni, Z.; et al. Unconventional splicing of wheat TabZIP60 confers heat tolerance in transgenic Arabidopsis. Plant Sci. 2018, 274, 252–260. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Reported Functions | Ortholog Gene in Potato |
|---|---|---|
| Activators | ||
| bZIP17 (At2g40950) | Involved in the perception and signaling of ER stress, salt stress, heat stress, and heavy metal toxicity [19,37,38]; can also regulate root elongation [39]. | StbZIP17 (Soltu.DM.10G029910) |
| bZIP28 (AT3g10800) | An ER stress sensor/transducer found to be involved in heat stress and plant development [39,40]. | StbZIP28 (Soltu.DM.04G021020) |
| IRE1A (At2g17520) | A subunit of ER stress sensor kinase IRE1, which was found to be involved in plant response to pathogens, abiotic stress, and plant growth and development [41,42,43]. | StIRE1A (Soltu.DM.02G022440) |
| IRE1B (At5g24360) | A subunit of ER stress sensor kinase IRE1, which can catalyze splicing of bZIP60 mRNA [13]. | StIRE1B (Soltu.DM.04G037840) |
| Immediate downstream components | ||
| bZIP60 (At1g42990) | A proteolysis-activated transcription factor involved in the ER stress response; plays important roles in plant immunity and abiotic stress [13,42,43,44,45,46]. | StbZIP60 (Soltu.DM.04G038150) |
| NAC103 (At5g64060) | A transcription factor that can be induced by bZIP60; it also regulates ABA response during seed germination and seedling growth [47,48]. | StNAC103 (Soltu.DM.03G011580) |
| S1P (At5g19660) | Protease is important for ER-stress-induced activation of bZIP28 and bZIP17 [19,22]. | StS1P (Soltu.DM.09G006520) |
| S2P (At4g20310) | A protease which can activate the bZIP28 transcription factor [49]. | StS2P (Soltu.DM.01G015980) |
| SAR1 (At3g62560) | A component of the COPII machinery; can interact with bZIP28 under ER stress [50]. | StSAR1 (Soltu.DM.01G040110) |
| Sec12 (At3g52190) | A component of the COPII machinery; can interact with bZIP28 under ER stress [50]. | StSec12 (Soltu.DM.07G016000) |
| Sec31A (AT1g18830) | A subunit of the COPII coat; can be upregulated in the UPR [51,52]. | StSec31A (Soltu.DM.01G026610) |
| Downstream components | ||
| (1) Molecular chaperones/foldases | ||
| CRT1 (At5g19380) | An ER-localized Ca2+-binding protein; related to biotic and abiotic stress signaling and tolerance [53,54]. | StCRT1 (Soltu.DM.12G001140) |
| CNX1 (At5g61790) | A critical component of ERQC machinery; it plays roles in growth, development, and response to stress [55]. | StCNX1 (Soltu.DM.06G024590) |
| ERDJ3B (At3g62600) | A co-chaperone of HSP40; it functions in coordinating ER and extracellular proteostasis [56]. | StERDJ3B (Soltu.DM.01G024680) |
| PDI (AT1g52260) | An ER foldase; its expression was induced under ER stress [57]. | StPDI (Soltu.DM.03G035420) |
| SES1 (AT4g29520) | It is an ER-localized chaperone which is related to plant response abiotic stress [27,58,59]. | StSES1 (Soltu.DM.07G010710) |
| (2) Folding of glycoproteins | ||
| UGGT (At1g71220) | A key player in the ERQC; plays roles in plant vegetative development and responses to abiotic and biotic stressors [60]. | StUGGT (Soltu.DM.01G042970) |
| OST (AT1g61790) | An oligosaccharyl-transferase; deficiency of it results in activation of the UPR and hypersensitivity to abiotic stress [61]. | StOST (Soltu.DM.03G002900) |
| (3) ERAD | ||
| MNS3 (At1g30000) | Class I α-mannosidase; plays important roles in N-glycan processing, root development, and cell wall biosynthesis [62]. | StMNS3 (Soltu.DM.02G013570) |
| MNS4 (At5g43710) | Class I α-mannosidase; involved in the degradation of misfolded glycoproteins [63]. | StMNS4 (Soltu.DM.06G014870) |
| MNS5 (At1g27520) | Class I α-mannosidase; involved in the degradation of misfolded glycoproteins [63]. | StMNS5 (Soltu.DM.03G027800) |
| DER (AT4g21810) | Can be upregulated by ER stress and is involved in the ERAD pathway [64]. | StDER (Soltu.DM.02G019990) |
| UBC32 (At3g17000) | A ubiquitin conjugation enzyme (E2) localized in the ER membrane; plays a role in salt and drought stress response [65,66]. | StUBC32 (Soltu.DM.12G001640) |
| CDC48 (At5g03340) | A hexametric AAA-ATPase complex required for ERAD; can maintain protein homeostasis [67]. | StCDC48 (Soltu.DM.06G030350) |
| UFD1 (At2g21270) | Ubiquitin fusion degradation protein that contributes to the regulation of stress responses through the ERAD pathway [68]. | StUFD1 (Soltu.DM.01G050260) |
| NPL4 (At2g47970) | An allele of HRD4 and participates in the ERAD pathway by forming the Cdc48p-Ufd1p-Npl4p complex [69]. | StNPL4 (Soltu.DM.06G034950) |
| PNG1 (At5g49570) | Has PNGase activity in vivo and facilitates ERAD through its de-glycosylation activity [70]. | StPNG1 (Soltu.DM.01G036610) |
| RAD23 (At1g16190) | It is a substrate-delivery co-factor; can deliver substrates to the proteasome for degradation [71]. | StRAD23 (Soltu.DM.03G032000) |
| DSK2 (At2g17200) | A substrate-delivery co-factor; can deliver substrates to the proteasome for degradation [71]. | StDSK2 (Soltu.DM.02G022800) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guan, P.; Zhao, D.; Zhang, C.; Qiu, Z.; Chen, Q.; Solyanikova, I.P.; Sun, P.; Cui, P.; Yu, R.; Zhang, X.; et al. Identification and Analysis of Endoplasmic-Reticulum-Stress- and Salt-Stress-Related Genes in Solanum tuberosum Genome: StbZIP60 Undergoes Splicing in Response to Salt Stress and ER Stress. Agronomy 2025, 15, 1224. https://doi.org/10.3390/agronomy15051224
Guan P, Zhao D, Zhang C, Qiu Z, Chen Q, Solyanikova IP, Sun P, Cui P, Yu R, Zhang X, et al. Identification and Analysis of Endoplasmic-Reticulum-Stress- and Salt-Stress-Related Genes in Solanum tuberosum Genome: StbZIP60 Undergoes Splicing in Response to Salt Stress and ER Stress. Agronomy. 2025; 15(5):1224. https://doi.org/10.3390/agronomy15051224
Chicago/Turabian StyleGuan, Peiyan, Dongbo Zhao, Chenxi Zhang, Zhennan Qiu, Qingshuai Chen, Inna P. Solyanikova, Peinan Sun, Peipei Cui, Ru Yu, Xia Zhang, and et al. 2025. "Identification and Analysis of Endoplasmic-Reticulum-Stress- and Salt-Stress-Related Genes in Solanum tuberosum Genome: StbZIP60 Undergoes Splicing in Response to Salt Stress and ER Stress" Agronomy 15, no. 5: 1224. https://doi.org/10.3390/agronomy15051224
APA StyleGuan, P., Zhao, D., Zhang, C., Qiu, Z., Chen, Q., Solyanikova, I. P., Sun, P., Cui, P., Yu, R., Zhang, X., Li, Y., & Hu, L. (2025). Identification and Analysis of Endoplasmic-Reticulum-Stress- and Salt-Stress-Related Genes in Solanum tuberosum Genome: StbZIP60 Undergoes Splicing in Response to Salt Stress and ER Stress. Agronomy, 15(5), 1224. https://doi.org/10.3390/agronomy15051224