Response of Yield and Protein Content of Forage Mulberry to Irrigation in North China Plain


Abstract
1. Introduction
2. Materials and Methods
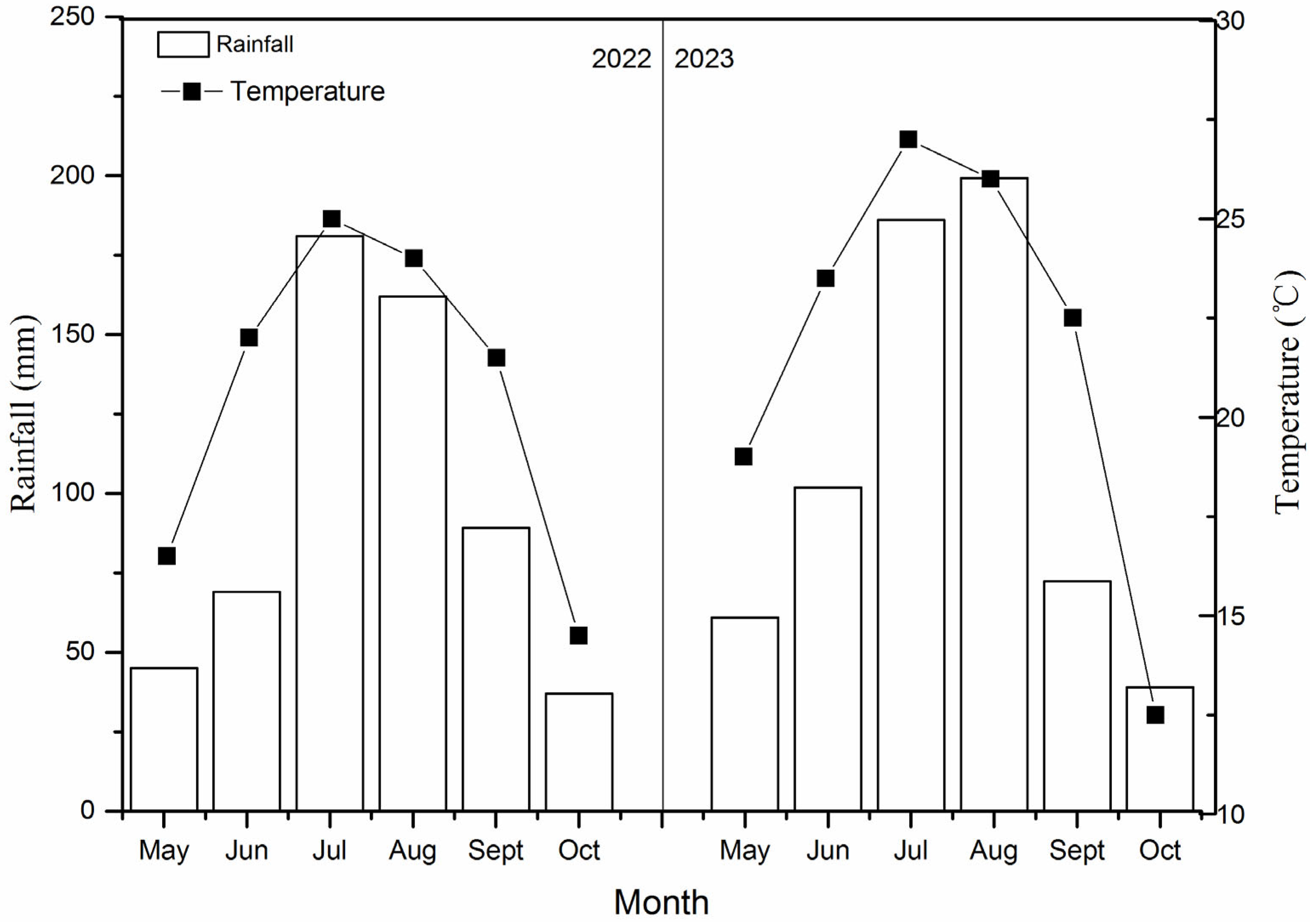
2.1. Experimental Site
2.2. Experimental Materials
2.3. Experimental Design
2.4. Measurements
2.4.1. Leaf Area
2.4.2. Fresh Leaf Weight and Leaf Dry Weight
2.4.3. Photosynthetic Rate and Leaf Water Use Efficiency (WUEL)
2.4.4. Protein Content Measurement
2.4.5. Yield Measurement
2.5. Statistical Analysis
3. Results
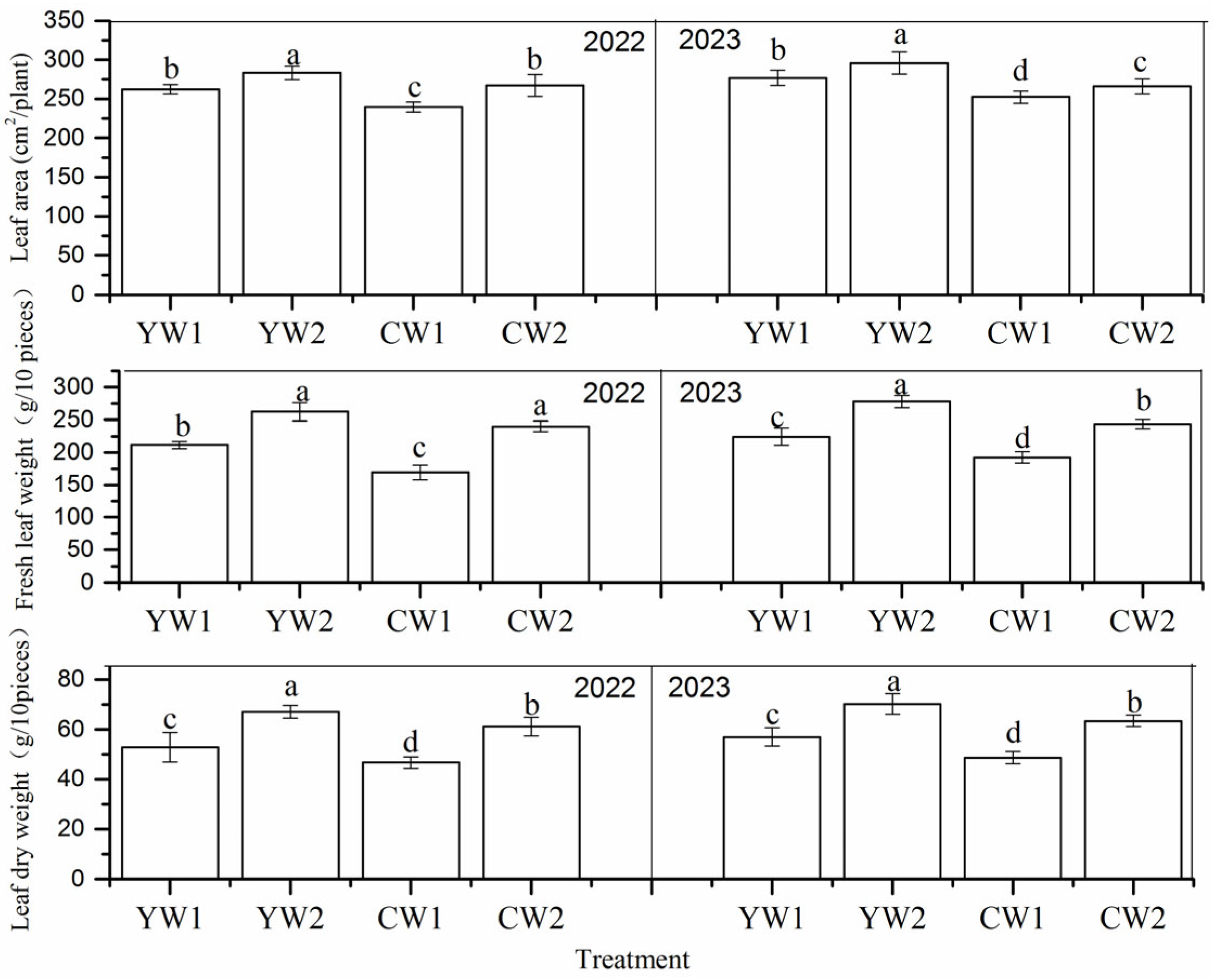
3.1. Leaf Area, Fresh Leaf Weight, and Leaf Dry Weight
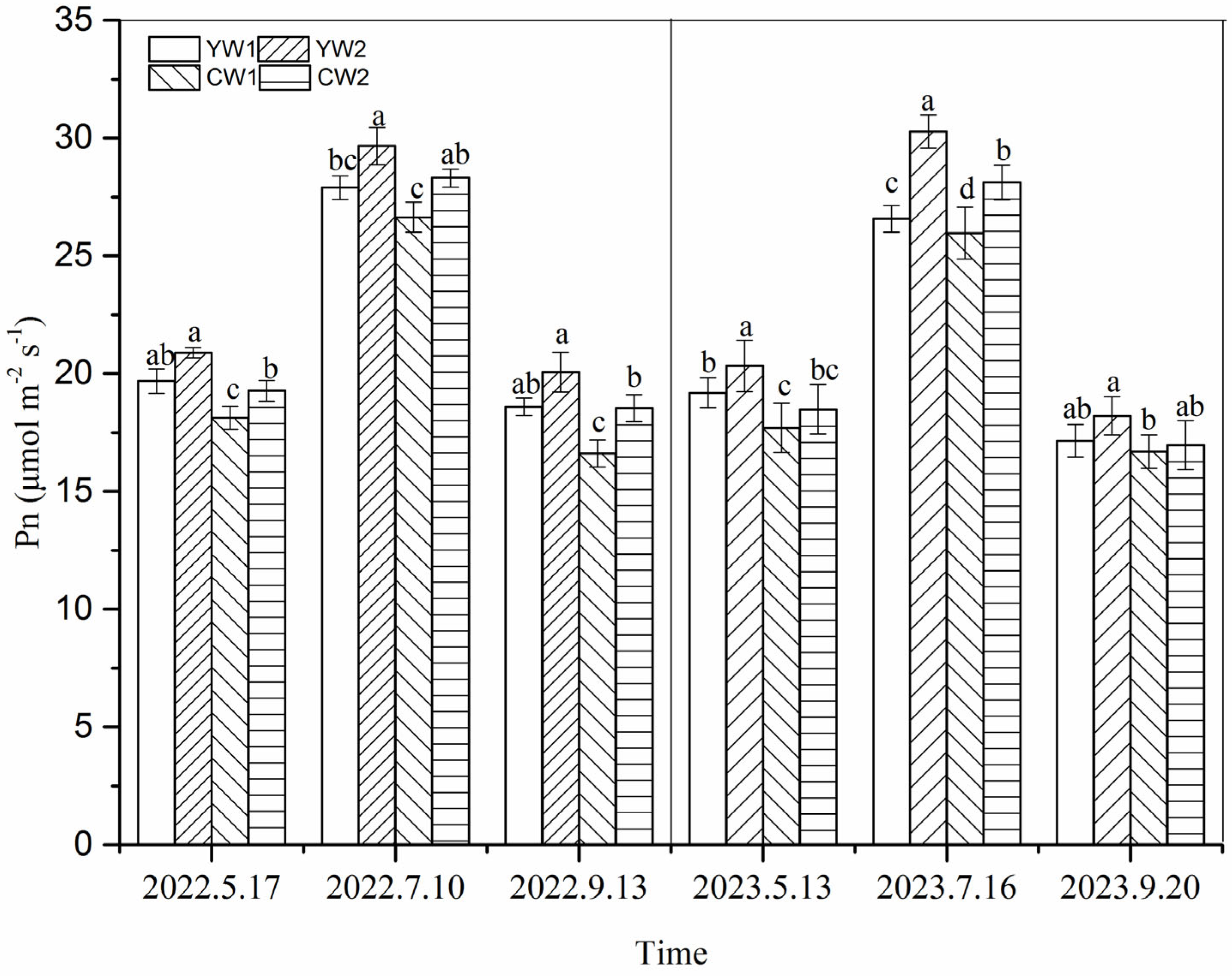
3.2. Photosynthetic Rate
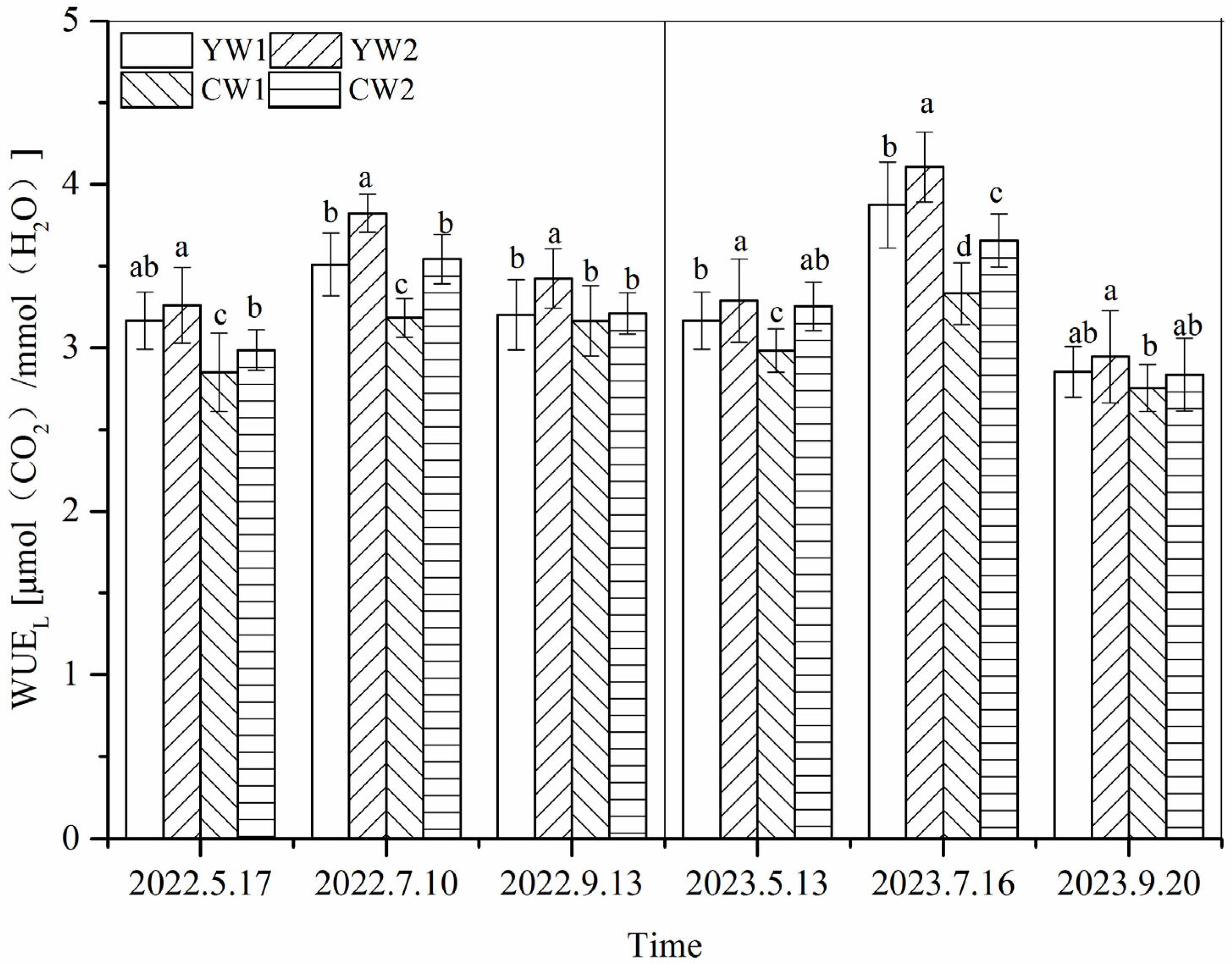
3.3. Leaf Water Use Efficiency (WUEL)
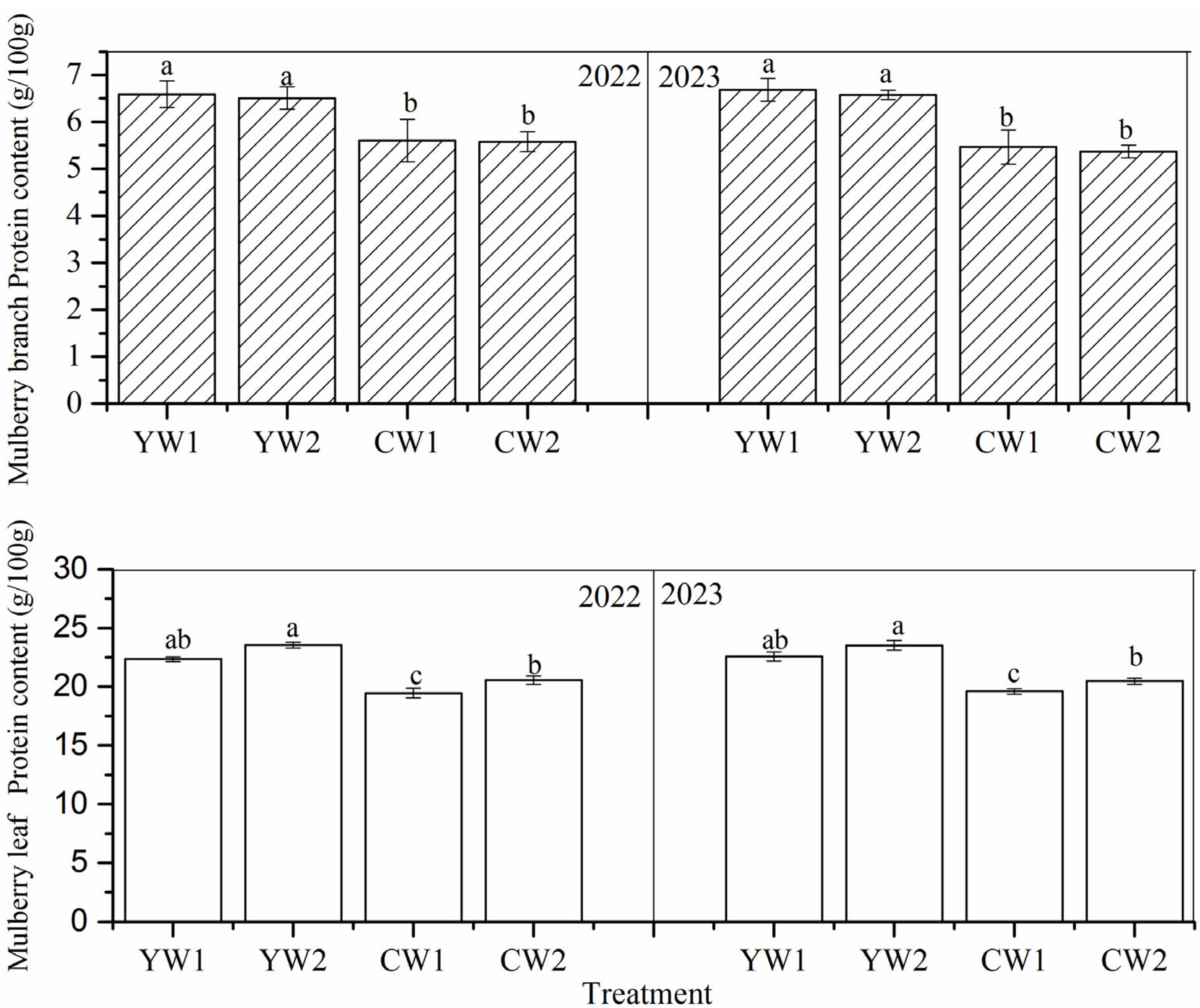
3.4. Protein Content
3.5. Yield
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shu, J.; Bai, Y.; Chen, Q.; Weng, C.; Zhang, F. Dynamic simulation of the water-land-food nexus for the sustainable agricultural development in the North China Plain. Sci. Total Environ. 2024, 912, 168771. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, J.; Jin, X.; Liu, S.; Tang, S.; Zhang, Z.; Long, S.; Piao, X. Effect of Dietary Supplemented with Mulberry Leaf Powder on Growth Performance, Serum Metabolites, Antioxidant Property and Intestinal Health of Weaned Piglets. Antioxidants 2023, 12, 307. [Google Scholar] [CrossRef]
- Wang, X.; Qian, Y.; Wei, M. Widely Targeted Metabolomics Analysis to Reveal Metabolite of Morus alba L. in Different Medicinal Parts. Molecules 2024, 29, 3981. [Google Scholar] [CrossRef]
- Fatima, M.; Dar, M.A.; Dhanavade, M.J.; Abbas, S.Z.; Bukhari, M.N.; Arsalan, A.; Liao, Y.; Wan, J.; Bukhari, J.S.; Zhen, O. Biosynthesis and Pharmacological Activities of the Bioactive Compounds of White Mulberry (Morus alba): Current Paradigms and Future Challenges. Biology 2024, 13, 506. [Google Scholar] [CrossRef]
- Garrett, N.R.; Pink, R.C.; Lawson, C. Contribution of Extracellular Particles Isolated from Morus sp. (Mulberry) Fruit to Their Reported Protective Health Benefits: An In Vitro Study. Int. J. Mol. Sci. 2024, 25, 6177. [Google Scholar] [CrossRef]
- Xiong, Y.; Guo, C.; Wang, L.; Chen, F.; Dong, X.; Li, X.; Ni, K.; Yang, F. Effects of Paper Mulberry Silage on the Growth Performance, Rumen Microbiota and Muscle Fatty Acid Composition in Hu Lambs. Fermentation 2021, 7, 286. [Google Scholar] [CrossRef]
- Alpízar-Naranjo, A.; Arece-García, J.; Esperance, M.; López, Y.; Molina, M.; González-García, E. Partial or total replacement of commercial concentrate with on-farm-grown mulberry forage: Effects on lamb growth and feeding costs. Trop. Anim. Health Prod. 2017, 49, 537–546. [Google Scholar] [CrossRef] [PubMed]
- Rukmangada, M.S.; Sumathy, R.; Kruthika, H.S.; Naik, V.G. Mulberry (Morus spp.) growth analysis by morpho-physiological and biochemical components for crop productivity enhancement. Sci. Hortic. 2020, 259, 108819. [Google Scholar] [CrossRef]
- Sun, Z.; Yin, Y.; Zhu, W.; Zhou, Y. Morphological, physiological, and biochemical composition of Mulberry (Morus spp.) under drought stress. Forests 2023, 14, 949. [Google Scholar] [CrossRef]
- Shi, Y.; Jin, X.; Ackah, M.; Amoako-Frank, K.; Li, J.; Tsigbey-Victor, E.; Li, H.; Cui, Z.; Sun, L.; Zhao, C.; et al. Comparative Physio-Biochemical and Transcriptome Analyses Reveal Contrasting Responses to Magnesium Imbalances in Leaves of Mulberry (Morus alba L.) Plants. Antioxidants 2024, 13, 516. [Google Scholar] [CrossRef]
- Chen, L.; Chang, J.; Wang, Y.; Guo, A.; Liu, Y.; Wang, Q.; Zhu, Y.; Zhang, Y.; Xie, Z. Disclosing the future food security risk of China based on crop production and water scarcity under diverse socioeconomic and climate scenarios. Sci. Total Environ. 2021, 790, 148110. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Lu, J.; Xiang, Y.; Shi, H.; Sun, T.; Zhang, W.; Wang, H.; Zhang, X.; Li, Z.; Zhang, F. Farmland mulching and optimized irrigation increase water productivity and seed yield by regulating functional parameters of soybean (Glycine max L.) leaves. Agr. Water Manag. 2024, 298, 108875. [Google Scholar] [CrossRef]
- Zappa, L.; Dari, J.; Modanesi, S.; Raphael, Q.; Luca, B.; Gabrielle-De, L.; Christian, M.; Quintana-Seguí, P.; Barella-Ortiz, A.; Dorigo, W. Benefits and pitfalls of irrigation timing and water amounts derived from satellite soil moisture. Agr. Water Manag. 2024, 295, 108773. [Google Scholar] [CrossRef]
- Ma, G.; Chai, X.; Hou, G.; Zhao, F.; Meng, Q. Phytochemistry, bioactivities and future prospects of mulberry leaves: A review. Food Chem. 2022, 372, 131335. [Google Scholar] [CrossRef]
- Cui, X.; Xie, W. Adapting Agriculture to Climate Change through Growing Season Adjustments: Evidence from Corn in China. Am. J. Agr. Econ. 2021, 104, 249–272. [Google Scholar] [CrossRef]
- Kayad, A.; Sozzi, M.; Gatto, S.; Whelan, B.; Sartori, L.; Marinello, F. Ten years of corn yield dynamics at field scale under digital agriculture solutions: A case study from North Italy. Comput. Electron. Agr. 2021, 185, 106126. [Google Scholar] [CrossRef]
- Banger, K.; Wagner-Riddle, C.; Grant, B.B.; Grant-Ward, N.; Smith-Craig, D.; Yang, J. Modifying fertilizer rate and application method reduces environmental nitrogen losses and increases corn yield in Ontario. Sci. Total Environ. 2020, 722, 137851. [Google Scholar] [CrossRef]
- Adeyemi, O.; Keshavarz-Afshar, R.; Jahanzad, E.; Battaglia, M.L.; Luo, Y.; Sadeghpour, A. Effect of Wheat Cover Crop and Split Nitrogen Application on Corn Yield and Nitrogen Use Efficiency. Agr. J. 2020, 10, 1081. [Google Scholar] [CrossRef]
- Mei, J.; Ji, K.; Su, L.; Wu, M.; Zhou, X.; Duan, E. Effects of FeSO4 dosage on nitrogen loss and humification during the composting of cow dung and corn straw. Bioresour. Technol. 2021, 341, 125867. [Google Scholar] [CrossRef]
- Li, J.; Han, T.; Liu, K.; Shen, Z.; Nano-Alemu, D. Optimizing potassium and nitrogen fertilizer strategies to mitigate greenhouse gas emissions in global agroecosystems4. Sci. Total Environ. 2024, 916, 170270. [Google Scholar] [CrossRef]
- Dai, F.; Zhuo, X.; Luo, G.; Wang, Z.; Xu, Y.; Wang, D.; Zhong, J.; Lin, S.; Chen, L.; Li, Z.; et al. Genomic resequencing unravels the genetic basis of domestication, expansion, and trait improvement in Morus atropurpurea. Adv. Sci. 2023, 10, 2300039. [Google Scholar] [CrossRef] [PubMed]
- Hassan, F.-U.; Arshad, M.A.; Li, M.; Rehman, M.S.; Loor-Juan, J.; Huang, J. Potential of Mulberry Leaf Biomass and Its Flavonoids to Improve Production and Health in Ruminants: Mechanistic Insights and Prospects. Animal 2020, 10, 2076. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Zhang, G.-Z.; Rita-Cindy, S.A.-A. Quantification of Water, Protein and Soluble Sugar in Mulberry Leaves Using a Handheld Near-Infrared Spectrometer and Multivariate Analysis. Molecules 2019, 24, 4439. [Google Scholar] [CrossRef]
- Zhao, L.; Ouyang, D.; Cheng, X.; Zhou, a.; Lin, L.; Wang, J.; Wu, Q.; Jia, J. Multi-frequency ultrasound-assisted cellulase extraction of protein from mulberry leaf: Kinetic, thermodynamic, and structural properties. Ultrason. Sonochem. 2023, 99, 554. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Duan, H.; Ren, X.; Guo, X.; Zhang, Y.; Li, J.; Zhang, F.; Chen, J.; Yang, X. Optimization of ultrasound-assisted cellulase degradation method on the extraction of mulberry leaf protein and its effect on the functional characteristics. Ultrason. Sonochem. 2023, 99, 561. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, W.; Cheng, J.; Guo, W.; Li, Y.; Li, C. Physiological and Proteomic Analysis of Seed Germination under Salt Stress in Mulberry. Front. Biosci. 2023, 28, 49. [Google Scholar] [CrossRef]
- Yang, S.; Fan, L.; Tan, P.; Lei, W.; Liang, J.; Gao, Z. Effects of Eurotium cristatum on chemical constituents and α-glucosidase activity of mulberry leaf tea. Food Biosci. 2023, 53, 102557. [Google Scholar] [CrossRef]
- Rao, L.; Li, S.; Cui, X. Leaf morphology and chlorophyll fluorescence characteristics of mulberry seedlings under waterlogging stress. Sci. Rep. 2021, 11, 13379. [Google Scholar] [CrossRef]
- Wang, K.; Kang, S.; Li, F.; Wang, X.; Xiao, Y.; Wang, J.; Xu, H. Relationship between fruit density and physicochemical properties and bioactive composition of mulberry at harvest. J. Food Compos. Anal. 2022, 106, 104322. [Google Scholar] [CrossRef]
- González-García, E.; Martín, G.M. Biomass yield and nutrient content of a tropical mulberry forage bank: Effects of season, harvest frequency and fertilization rate. Grass Forage Sci. 2017, 72, 248–260. [Google Scholar] [CrossRef]
- Wu, A.; Brider, J.; Busch, F.A.; Chen, M.; Karine, C.; Victoria-Clarke, C.; Brian, C.; Ermakova, M.; John-Evans, R.; Graham-Farquhar, D.; et al. A cross-scale analysis to understand and quantify the effects of photosynthetic enhancement on crop growth and yield across environments. Plant Cell Environ. 2023, 46, 23–44. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Matthews, M.L. Seasonal climate conditions impact the effectiveness of improving photosynthesis to increase soybean yield. Field Crop. Res. 2023, 296, 108907. [Google Scholar] [CrossRef]
- Hoover, D.L.; Abendroth, L.J.; Browning, D.M.; Saha, A.; Snyder, K.; Wagle, P.; Witthaus, L.; Baffaut, C.; Biederman, J.A.; Bosch, D.D.; et al. Indicators of water use efficiency across diverse agroecosystems and spatiotemporal scales. Sci. Total Environ. 2023, 864, 160992. [Google Scholar] [CrossRef]
- Pu, X.; Lyu, L. Disentangling the impact of photosynthesis and stomatal conductance on rising water-use efficiency at different altitudes on the Tibetan plateau. Agr. Forest Meteorol. 2023, 341, 109659. [Google Scholar] [CrossRef]
- Haworth, M.; Marino, G.; Materassi, A.; Antonio, R.; Charles, P.S.; Mauro, C. The functional significance of the stomatal size to density relationship: Interaction with atmospheric [CO2] and role in plant physiological behaviour. Sci. Total Environ. 2023, 863, 160908. [Google Scholar] [CrossRef]
- Hu, J.; Zhao, X.; Gu, L.; Liu, P.; Zhao, B.; Zhang, J.; Ren, B. The effects of high temperature, drought, and their combined stresses on the photosynthesis and senescence of summer maize. Agr. Water Manag. 2023, 289, 108525. [Google Scholar] [CrossRef]
- Berbel, J.; Cuadrado-Alarcón, B.; Martínez-Dalmau, J.; Delgado-Ramos, F. Hydro-economic basin impacts of extensive adoption of deficit irrigation by farmers: Are we overestimating water resources? J. Hydrol. 2024, 634, 131075. [Google Scholar] [CrossRef]
- Tolimir, M.; Gajić, B.; Kresović, B.; Životić, L.; Gajić, K. Impact of deficit irrigation and planting density on grain yield and water productivity of maize grown under temperate continental climatic conditions. Agr. Water Manag. 2024, 302, 109009. [Google Scholar] [CrossRef]
- Liu, L.; Guo, Z.; Kang, X.; Li, S.; Huang, S.; Zheng, L.; Fu, R.; Keermula, Y.; Chao, N. Comparative Transcriptome Analysis of Different Mulberry Varieties to Reveal Candidate Genes and Small Secreted Peptides Involved in the Sclerotiniose Response. Forests 2024, 15, 1126. [Google Scholar] [CrossRef]
- Wu, S.; Deng, L.; Guo, L.; Wu, Y. Wheat leaf area index prediction using data fusion based on high-resolution unmanned aerial vehicle imagery. Plant Methods 2022, 18, 68. [Google Scholar] [CrossRef]
- Guha, A.; Sengupta, D.; Reddy, A.R. Physiological optimality, allocation trade-offs and antioxidant protection linked to better leaf yield performance in drought exposed mulberry. J. Sci. Food Agr. 2010, 90, 2649–2659. [Google Scholar] [CrossRef] [PubMed]
- Zahra, N.; Hafeez, M.B.; Kausar, A.; Zeidi, M.A.; Asekova, S.; Siddique, H.M.; Farooq, M. Plant photosynthetic responses under drought stress: Effects and management. J. Agron. Crop. Sci. 2023, 209, 651–672. [Google Scholar] [CrossRef]
- Zhang, Y.; Gentine, P.; Luo, X.; Lian, X.; Liu, Y.; Zhou, S.; Keenan, T.F. Increasing sensitivity of dryland vegetation greenness to precipitation due to rising atmospheric CO2. Nat. Commun. 2022, 13, 4875. [Google Scholar] [CrossRef] [PubMed]
- Konings, A.G.; Saatchi, S.S.; Frankenberg, C.; Keller, M.; Leshyk, V.; Anderegg, W.L.; Pieter, A.Z. Detecting Forest Response to Droughts with Global Observations of Vegetation Water Content. Glob. Change Biol. 2021, 27, 6005–6024. [Google Scholar] [CrossRef]
- Cooley, S.S.; Fisher, J.B.; Goldsmith, G.R. Convergence in water use efficiency within plant functional types across contrasting climates. Nat. Plants 2022, 8, 341–345. [Google Scholar] [CrossRef]
- Deans, R.M.; Brodribb, T.J.; Busch, F.A.; Farquhar, G.D. Optimization can provide the fundamental link between leaf photosynthesis, gas exchange and water relations. Nat. Plants 2020, 6, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, J.N.; Schmuker, P.; Dmitrieva, A.; Quach, T.; Zhang, T.; Ge, Z.; Nersesian, N.; Sato, S.J.; Clemente, T.E.; Leakey, A.D.B. Reducing stomatal density by expression of a synthetic Epidermal Patterning Factor increases leaf intrinsic water use efficiency and reduces plant water use in a C4 crop. J. Exp. Bot. 2024, 21, 6823–6836. [Google Scholar] [CrossRef]
- Yan, C.; Chen, F.; Yang, Y.; Shen, L.; Xun, X.; Zhang, Z.; Zhan, Y.; You, S.; Wang, J. Biochemical and protein nutritional potential of mulberry (Morus alba L.) leaf: Partial substitution improves the nutrition of conventional protein. J. Sci. Food Agr. 2023, 4, 2204–2214. [Google Scholar] [CrossRef]
- Wu, Z.; Liang, C.; Huang, R.; Ouyang, J.; Zhao, L.; Bu, D. Replacing alfalfa hay with paper mulberry (Broussonetia papyrifera L.) silage in diets do not affect the production performance of the low lactating dairy cows. Anim. Feed Sci. Technol. 2022, 294, 115477. [Google Scholar] [CrossRef]
- Geng, B.; Gao, J.; Cheng, H.; Guo, G.; Wang, Z. Effects of dietary mulberry leaves on growth, production performance, gut microbiota, and immunological parameters in poultry and livestock: A systematic review and meta-analysis. Anim. Biosci. 2024, 37, 1065. [Google Scholar] [CrossRef]
- Cui, Y.; Liu, Z.; Deng, D.; Tian, Z.; Song, M. Influence of Fermented Mulberry Leaves as an Alternative Animal Feed Source on Product Performance and Gut Microbiome in Pigs. Fermentation 2024, 10, 215. [Google Scholar] [CrossRef]
- Ackah, M.; Shi, Y.; Wu, M.; Wang, L.; Guo, P.; Guo, L.; Jin, X.; Li, S.; Zhang, Q.; Qiu, C.; et al. Metabolomics Response to Drought Stress in Morus alba L. Variety Yu-711. Plants 2021, 10, 1636. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, T.; Krishnan, R.; Doss, S.G.; Sarkar, T.B. Evaluation of Mulberry Genotypes Under Moisture Stress for the Identification of Physio-biochemical Markers of Stress Tolerance. J. Plant Growth Regul. 2024, 43, 1999–2013. [Google Scholar] [CrossRef]
- Guo, Y.; Kang, X.; Huang, Y.; Guo, Z.; Wang, Y.; Ma, S.; Li, H.; Chao, N.; Liu, L. Functional characterization of MaEXPA11 and its roles in response to biotic and abiotic stresses in mulberry. Plant Physiol. Bioch. 2024, 206, 108289. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Main Plot Genotypes | Subplot Irrigation Water |
|---|---|---|
| YW1 | Feng Yuan No. 1 (Y) | 15 mm (W1) |
| YW2 | Feng Yuan No. 1 (Y) | 30 mm (W2) |
| CW1 | Feng Chi (C) | 15 mm (W1) |
| CW2 | Feng Chi (C) | 30 mm (W2) |
| Time | Leaf Number | Branch Weight | Branch Number | Yield |
|---|---|---|---|---|
| (Piece/Plant) | (g/Plant) | (Branch/Plant) | (t/ha) | |
| 2022 | ||||
| Interaction | ||||
| Y | 109.29 a | 587.10 a | 7.67 a | 9.38 a |
| C | 97.82 b | 532.30 b | 6.29 b | 8.39 b |
| W1 | 96.65 b | 521.20 b | 6.61 b | 8.45 b |
| W2 | 110.45 a | 599.09 a | 7.35 a | 9.22 a |
| Coupling | ||||
| YW1 | 100.38 b | 536.06 b | 7.37 c | 8.98 b |
| YW2 | 118.19 a | 639.10 a | 7.79 a | 9.79 a |
| CW1 | 92.93 c | 507.03 c | 5.86 d | 7.91 c |
| CW2 | 102.71 b | 558.02 b | 6.73 b | 8.86 b |
| 2023 | ||||
| Interaction | ||||
| Y | 112.72 a | 618.20 a | 8.11 a | 10.83 a |
| C | 100.78 b | 552.60 b | 6.51 b | 9.71 b |
| W1 | 99.410 b | 546.05 b | 6.77 b | 9.73 b |
| W2 | 114.10 a | 624.03 a | 7.86 a | 10.86 a |
| Coupling | ||||
| YW1 | 100.38 b | 577.07 b | 7.54 b | 10.47 b |
| YW2 | 121.56 a | 659.60 a | 8.68 a | 11.19 a |
| CW1 | 94.94 c | 515.80 c | 5.99 c | 8.97 c |
| CW2 | 106.62 b | 588.01 b | 7.03 b | 10.46 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, Y.; Guo, G.; Wang, Z.; Zhu, L.; Geng, B. Response of Yield and Protein Content of Forage Mulberry to Irrigation in North China Plain. Agronomy 2025, 15, 1016. https://doi.org/10.3390/agronomy15051016
Ren Y, Guo G, Wang Z, Zhu L, Geng B. Response of Yield and Protein Content of Forage Mulberry to Irrigation in North China Plain. Agronomy. 2025; 15(5):1016. https://doi.org/10.3390/agronomy15051016
Chicago/Turabian StyleRen, Yujie, Guang Guo, Zhaohong Wang, Lin Zhu, and Bing Geng. 2025. "Response of Yield and Protein Content of Forage Mulberry to Irrigation in North China Plain" Agronomy 15, no. 5: 1016. https://doi.org/10.3390/agronomy15051016
APA StyleRen, Y., Guo, G., Wang, Z., Zhu, L., & Geng, B. (2025). Response of Yield and Protein Content of Forage Mulberry to Irrigation in North China Plain. Agronomy, 15(5), 1016. https://doi.org/10.3390/agronomy15051016