
Genome-Wide Identification of LACS Family Genes and Functional Characterization of CaLACS6/9 in Response to Cold Stress in Pepper (Capsicum annuum L.)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Databases
2.2. Dentification of CaLACS Genes Family Member in Pepper
2.3. Sequence Analysis and Evolutionary Tree Construction of CaLACS Members
2.4. Analysis of the Protein Conserved Motifs and Gene Structure
2.5. Analysis of CaLACSs Promoter Region
2.6. Analysis of Gene Collinearity and Selective Pressure
2.7. CaLACS Gene Expression Analysis of RNA-seq Data Under Different Conditions
2.8. VIGS of CaLACS6 and CaLACS9 Genes in Pepper
2.9. Plant Materials and Cold Stress Treatments
2.10. Measurement of Physiological Indicators
2.11. RNA Extraction and RT-qPCR
2.12. Statistical Analysis
3. Results
3.1. Identification of CaLACS Family Genes in Pepper
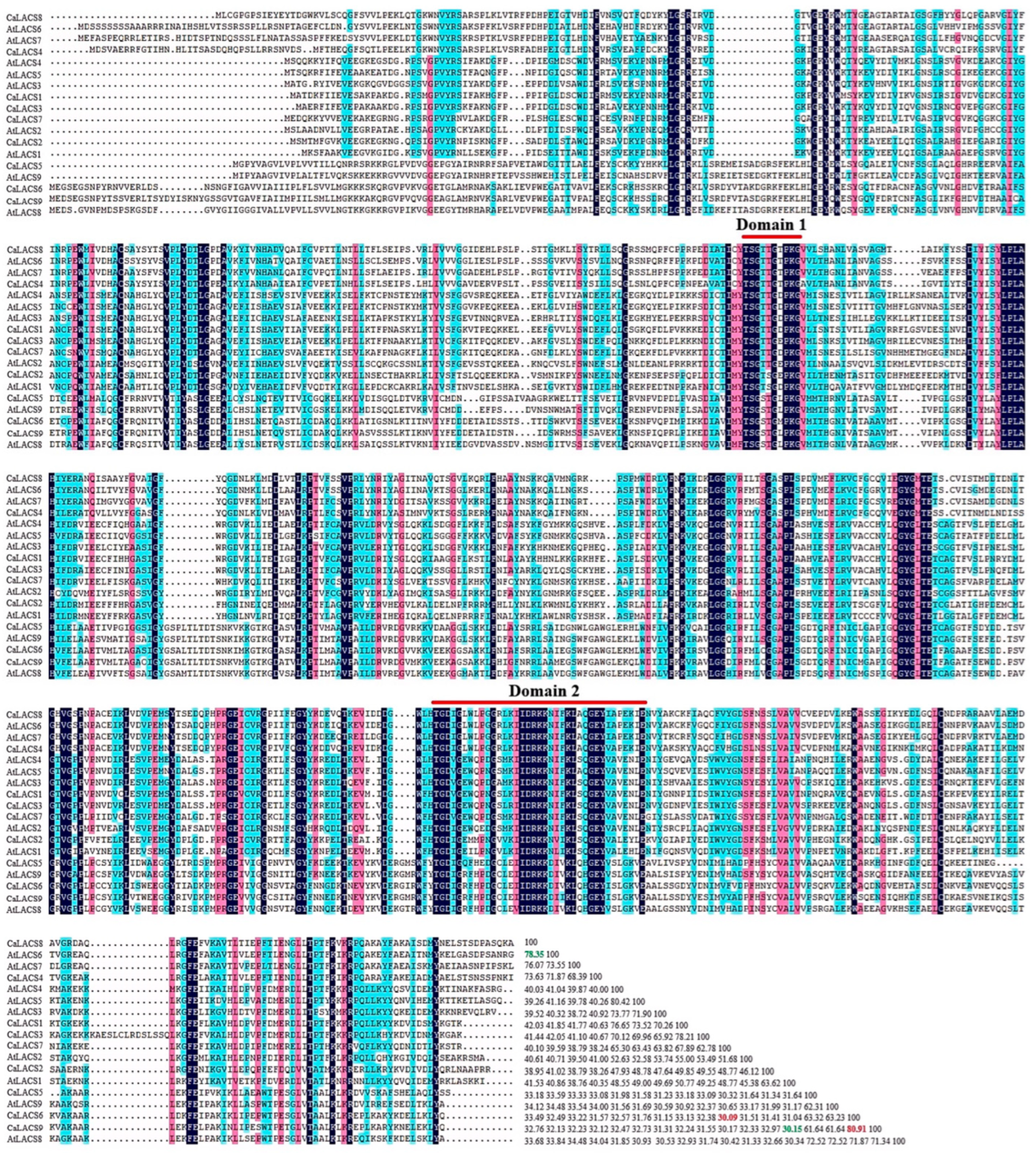
3.2. Sequence Alignment and Conserved Domains Analysis
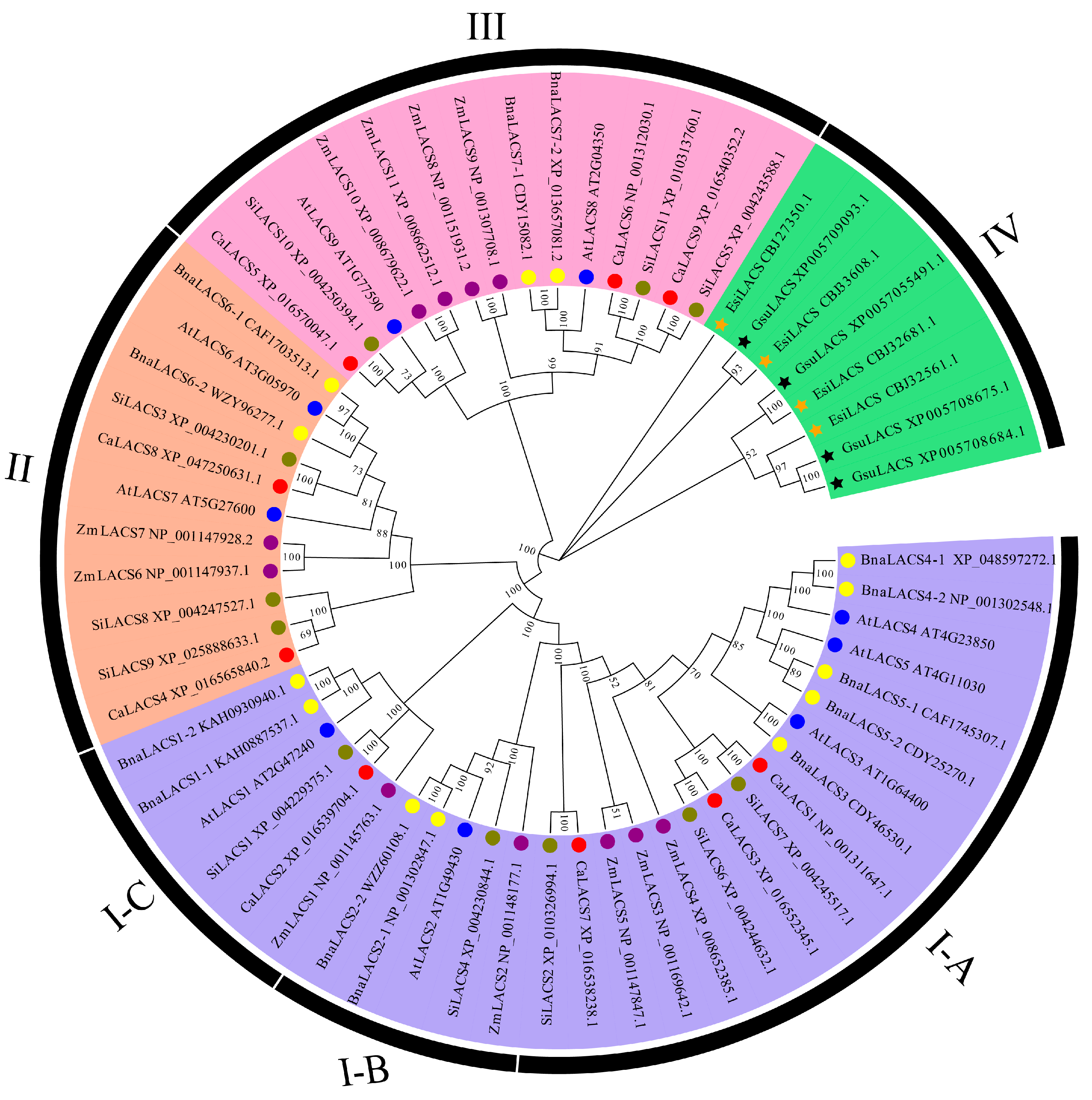
3.3. Phylogenetic Analysis of CaLACS Members in Pepper
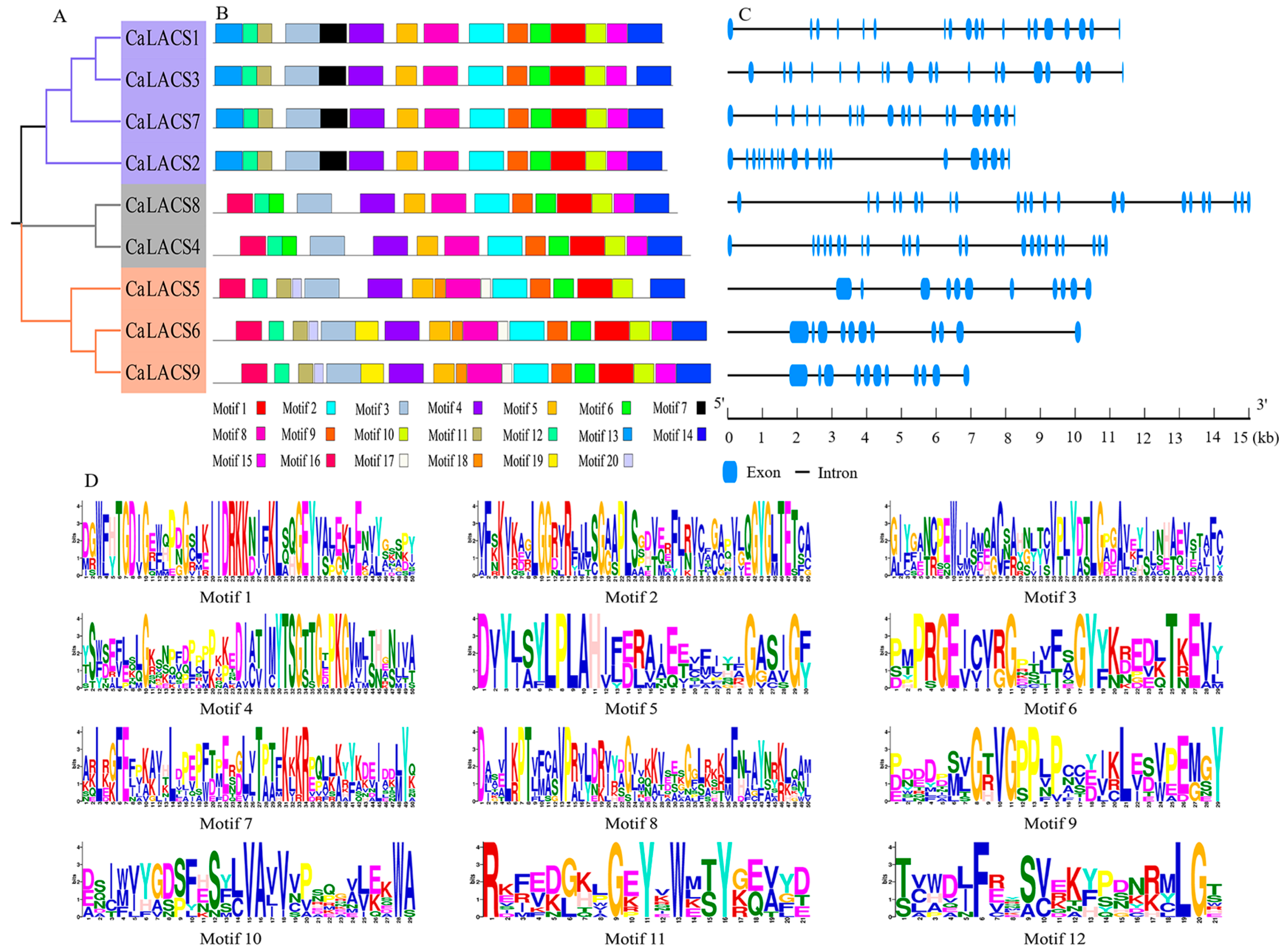
3.4. Motif and Structure of CaLACS Members in Pepper
3.5. Cis-Element Analysis of the CaLACS Promoter in Pepper
3.6. Collinearity Analysis and Selective Pressure of CaLACS Genes
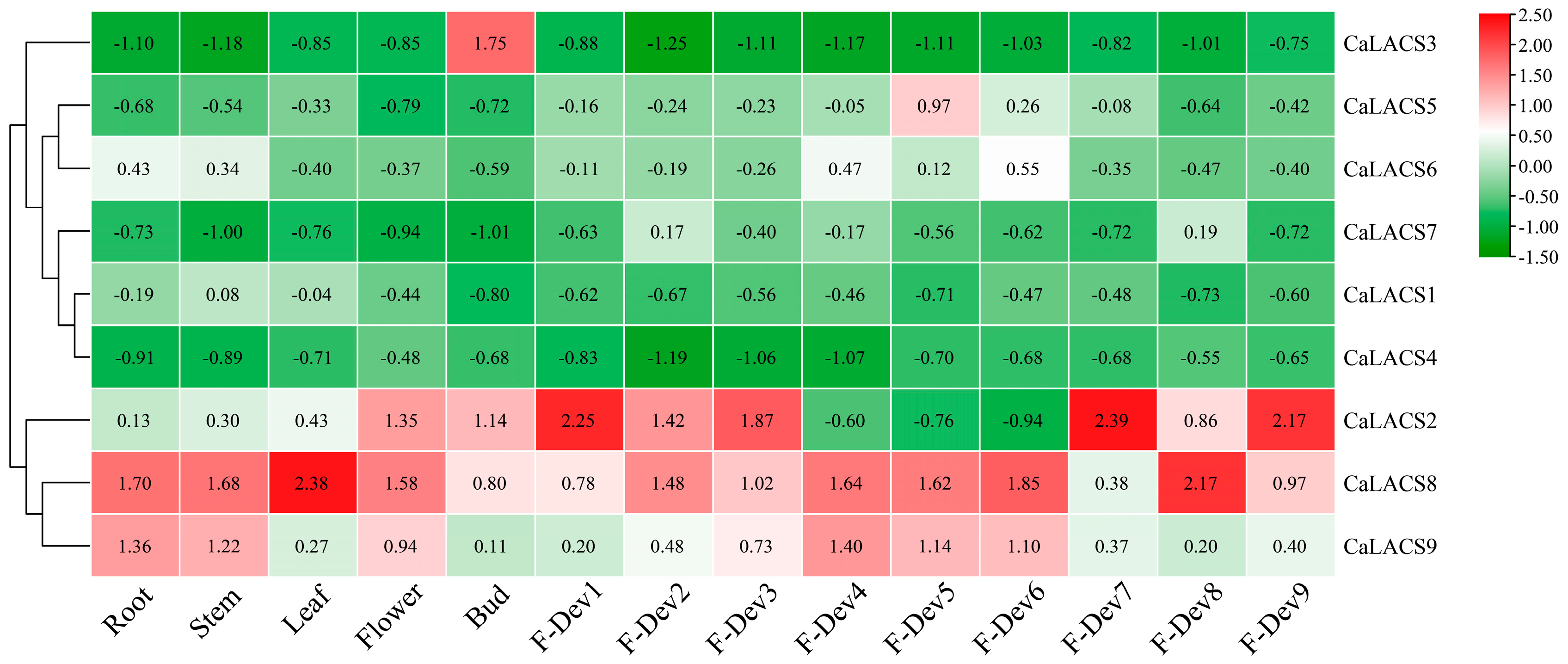
3.7. Expression of CaLACS Gene in Different Tissues
3.8. Expression Analysis of CaLACS Genes Under Different Phytohormone and Stresses Treatments
3.9. Influence of CaLACS6 and CaLACS9 Silenced Pepper Plants to Cold Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lü, S.Y.; Song, T.; Kosma, D.K.; Parsons, E.P.; Rowland, O.; Jenks, M.A. Arabidopsis CER8 encodes Long-Chain Acyl-Coa SynThetase 1 (LACS1) that has overlapping functions with LACS2 in plant wax and cutin synthesis. Plant J. 2009, 59, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Shockey, J.M.; Fulda, M.S.; Browse, J.A. Arabidopsis contains nine long-chain acyl-coenzyme a synthetase genes that participate in fatty acid and glycerolipid metabolism. Plant Physiol. 2002, 129, 1710–1722. [Google Scholar] [CrossRef] [PubMed]
- Tonon, T.; Qing, R.W.; Harvey, D.; Li, Y.; Larson, T.R.; Graham, I.A. Identification of a long-chain polyunsaturated fatty acid acyl-Coenzyme a synthetase from the Diatom Thalassiosira pseudonana. Plant Physiol. 2005, 138, 402–408. [Google Scholar] [CrossRef]
- Schnurr, J.; Shochey, J.; Browse, J. The Acyl-CoA synthetase encoded by LACS2 is essential for normal cuticle development in Arabidopsis. Plant Cell 2004, 16, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Fulda, M.; Schnurr, J.; Abbadi, A.; Heinz, E.; Browse, J. Peroxisomal acyl-CoA synthetase activity is essential for seedling development in Arabidopsis thaliana. Plant Cell 2004, 16, 394–405. [Google Scholar] [CrossRef]
- Kosma, D.K.; Bourdenx, B.; Bernard, A.; Parsons, E.P.; Lü, S.; Joubès, J.; Jenks, M.A. The impact of water deficiency on leaf cuticle lipids of Arabidopsis. Plant Physiol. 2009, 151, 1918–1929. [Google Scholar] [CrossRef]
- Liu, Y.J.; Liu, Z.H.; Zhang, H.S.; Yuan, S.H.; Li, Y.M.; Zhang, T.B.; Bai, J.F.; Zhang, L.P. Genome-wide identification and expression profiling analysis of the long-chain acyl-CoA synthetases reveal their potential roles in wheat male fertility. Int. J. Mol. Sci. 2022, 23, 11942. [Google Scholar] [CrossRef]
- Xiao, Z.C.; Li, N.N.; Wang, S.F.; Sun, J.J.; Zhang, L.Y.; Zhang, C.; Yang, H.; Zhao, H.Y.; Yang, B.; Wei, L.J.; et al. Genome-wide identification and comparative expression profile analysis of the long-chain acyl-CoA synthetase (LACS) gene family in two different oil content cultivars of brassica napus. Biochem. Genet. 2019, 57, 781–800. [Google Scholar] [CrossRef]
- Zhang, C.L.; Mao, K.; Zhou, L.J.; Wang, G.L.; Zhang, Y.L.; Li, Y.Y.; Hao, Y.J. Genome-wide identification and characterization of apple long-chain Acyl-CoA synthetases and expression analysis under different stresses. Plant Physiol. Biochem. 2018, 132, 320–332. [Google Scholar] [CrossRef]
- Wang, X.C.; Tian, X.; Zhang, H.R.; Li, H.T.; Zhang, S.D.; Li, H.; Zhu, J.T. Genome-wide analysis of the maize LACS gene family and functional characterization of the ZmLACS9 responses to heat stress. Plant Stress 2023, 10, 100271. [Google Scholar] [CrossRef]
- Xu, Y.C.; Fu, S.Y.; Huang, Y.W.; Zhou, D.Y.; Wu, Y.Z.; Peng, J.; Kuang, M. Genome-wide expression analysis of LACS gene family implies GhLACS25 functional responding to salt stress in cotton. BMC Plant Biol. 2024, 24, 392. [Google Scholar] [CrossRef]
- Zhong, Y.K.; Wang, Y.B.; Li, P.T.; Gong, W.K.; Wang, X.Y.; Yan, H.L.; Ge, Q.; Liu, A.Y.; Shi, Y.Z.; Shang, H.H.; et al. Genome-wide analysis and functional characterization of LACS gene family associated with lipid synthesis in cotton (Gossypium spp.). Int. J. Mol. Sci. 2023, 24, 8530. [Google Scholar] [CrossRef]
- Yan, Z.W.; Hou, J.; Leng, B.Y.; Yao, G.Q.; Ma, C.L.; Sun, Y.; Liu, Q.T.; Zhang, F.J.; Mu, C.H.; Liu, X. Genome-wide identification and characterization of maize Long-chain acyl-CoA snthetases and their expression profiles in different tissues and in response to multiple abiotic stresses. Genes 2024, 15, 983. [Google Scholar] [CrossRef] [PubMed]
- Weng, H.; Molina, I.; Shockey, J.; Browse, J. Organ fusion and defective cuticle function in a lacs1 lacs2 double mutant of Arabidopsis. Planta 2010, 231, 1089–1100. [Google Scholar] [CrossRef] [PubMed]
- Pulsifer, I.P.; Kluge, S.; Rowland, O. Arabidopsis long-chain acyl-CoA synthetase 1 (LACS1), LACS2, and LACS3 facilitate fatty acid uptake in yeast. Plant Physiol. Biochem. 2012, 51, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.F.; Haslam, T.M.; Sonntag, A.; Molina, I.; Kunst, L. Functional overlap of long-chain acyl-CoA synthetases in Arabidopsis. Plant Cell Physiol. 2019, 60, 1041–1054. [Google Scholar] [CrossRef]
- Zhao, L.F.; Katavic, V.; Li, F.L.; George, W.; Ljerka, H. Insertional mutant analysis reveals that long-chain acyl-CoA synthetase 1 (LACS1), but not LACS8, functionally overlaps with LACS9 in Arabidopsis seed oil biosynthesis. Plant J. 2010, 64, 1048–1058. [Google Scholar] [CrossRef]
- Aznar-Moreno, J.A.; Calerón, M.V.; Martínez-Force, E.; Garcés, R.; Mullen, R.; Gidda, S.K.; Salas, J.J. Sunflower (Helianthus annuus) long-chain acyl-coenzyme A synthetases expressed at high levels in developing seeds. Physiol. Plant. 2014, 150, 363–373. [Google Scholar] [CrossRef]
- Tan, X.L. Cloning and functional identification of a Long-chain acyl-CoA synthase gene (BnaLACS8A03) from Brassica napus L. Master’s Thesis, Jiangsu University, Zhenjiang, China, 2021; pp. 45–66. [Google Scholar] [CrossRef]
- Ma, W.J.; Zhu, K.K.; Zhao, J.; Chen, M.Y.; Wei, L.; Qiao, Z.B.; Tan, P.P.; Peng, F.R. Genome-wide identification, characterization, and expression analysis of long-chain acyl-CoA synthetases in Carya illinoinensis under different treatments. Int. J. Mol. Sci. 2023, 24, 11558. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Zhang, S.; Li, M.; Wang, J.; Wu, T. The PoLACS4 gene may participate in drought stress resistance in tree peony (Paeonia ostia ‘Feng Dan Bai’). Genes 2022, 13, 1591. [Google Scholar] [CrossRef]
- Zhang, C.L.; Hu, X.; Zhang, Y.L.; Liu, Y.; Wang, G.L.; You, C.X.; Li, Y.Y.; Hao, Y.J. An apple long-chain acyl-CoA synthetase 2 gene enhances plant resistance to abiotic stress by regulating the accumulation of cuticular wax. Tree Physiol. 2020, 40, 1450–1465. [Google Scholar] [CrossRef]
- Ayaz, A.; Huang, H.D.; Zheng, M.L.; Zaman, W.; Li, D.H.; Saqib, S.; Zhao, H.Y.; Lü, S.Y. Molecular cloning and functional analysis of GmLACS2-3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Zhao, X.C.; Wei, Y.L.; Zhang, J.J.; Yang, L.; Liu, X.Y.; Zhang, H.Y.; Shao, W.J.; He, L.; Li, Z.T.; Zhang, Y.F.; et al. Membrane lipids’ metabolism and transcriptional regulation in maize roots under cold stress. Front. Plant Sci. 2021, 12, 639132. [Google Scholar] [CrossRef]
- Wei, H.; Dhanaraj, A.L.; Arora, R.; Rowland, L.J.; Fu, Y.; Sun, L. Identification of cold acclimation-responsive Rhododendron genes for lipid metabolism, membrane transport and lignin biosynthesis: Importance of moderately abundant ESTs in genomic studies. Plant Cell Environ. 2006, 29, 558–570. [Google Scholar] [CrossRef]
- Zou, Z.Y.; Zou, X.X. Geographical and Ecological Differences in Pepper Cultivation and Consumption in China. Front Nutr. 2021, 596, 718517. [Google Scholar] [CrossRef]
- Ou, L.J.; Wei, G.; Zhang, Z.Q.; Dai, X.Z.; Zou, X.X. Effects of low temperature and low irradiance on the physiological characteristics and related gene expression of different pepper species. Photosynthetica 2015, 53, 85–94. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Wang, H.J.; Wang, T.; Guo, D. Extreme spring cold spells in North China during 1961-2014 and the evolving processes. Atmos. Ocean. Sci. Lett. 2018, 11, 6. [Google Scholar] [CrossRef]
- Li, J.; Yang, P.; Fu, H.B.; Li, J.; Wang, Y.Z.; Zhu, K.Y.; Yu, J.H.; Li, J. Transcriptome analysis reveals key regulatory networks and genes involved in the acquisition of cold stress memory in pepper seedlings. BMC Plant Biol. 2024, 24, 959. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple sequence alignment using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2002, 2, 3. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.P.; Guo, A.Y.; Zhang, H.; Luo, J.C.; Gao, C. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative Toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.P.; Tang, H.B.; DeBarry, J.D.; Tan, X.; Li, J.P.; Wang, X.Y.; Lee, T.H.; Jin, H.Z.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence polymorphism analysis of large datasets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Kim, S.; Park, M.; Yeom, S.; Kim, Y.M.; Lee, J.M.; Lee, H.A.; Seo, E.; Choi, J.; Cheong, K.; Kim, K.T.; et al. Genome sequence of the hot pep-per provides insights into the evolution of pungency in Capsicum species. Nat. Genet. 2014, 46, 270–278. [Google Scholar] [CrossRef]
- Qin, C.; Yu, C.S.; Shen, Y.O.; Fang, X.D.; Chen, L.; Min, J.M.; Cheng, J.W.; Zhao, S.C.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of culti-vated and wild pepper provides insights into capsicum domesti-cation and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef]
- Lee, J.; Nam, J.Y.; Jang, H.; Kim, N.; Kim, Y.M.; Kang, W.H.; Yeom, S.I. Comprehensive transcriptome resource for response to phytohormone-induced signaling in Capsicum annuum L. BMC Res. Notes 2020, 13, 440. [Google Scholar] [CrossRef]
- Kang, W.H.; Sim, Y.M.; Koo, N.; Nam, J.Y.; Lee, J.; Kim, N.; Jang, H.; Kim, Y.M.; Yeom, S.I. Transcriptome profiling of abiotic responses to heat, cold, salt, and osmotic stress of Capsicum annuum L. Sci. Data 2020, 7, 17. [Google Scholar] [CrossRef]
- Fernandez-Pozo, N.; Rosli, H.G.; Martin, G.B.; Mueller, L.A. The SGN VIGS Tool: User-friendly software to design virus-induced gene silencing (VIGS) constructs for functional genomics. Mol. Plant 2014, 8, 486–488. [Google Scholar] [CrossRef]
- Song, J.; Sun, P.P.; Kong, W.N.; Xie, Z.Z.; Li, C.L.; Liu, J.H. SnRK2.4-mediated phosphorylation of ABF2 regulates ARGININE DECARBOXYLASE expression and putrescine accumulation under drought stress. New Phytol. 2023, 238, 216–236. [Google Scholar] [CrossRef]
- Velásquez, A.; Chakravarthy, S.; Martin, G. Virus-induced gene silencing (VIGS) in Nicotiana benthamiana and tomato. J. Vis. Exp. 2009, 28, e1292. [Google Scholar] [CrossRef]
- Liu, J.; Lu, H.Y.; Wan, Q.; Qi, W.C.; Shao, H.B. Genome-wide analysis and expression profiling of respiratory burst oxidase homologue gene family in Glycine max. Environ. Exp. Bot. 2019, 161, 344–356. [Google Scholar] [CrossRef]
- Yin, Y.X.; Wang, S.B.; Xiao, H.J.; Zhang, H.X.; Zhang, Z.; Jing, H.; Zhang, Y.L.; Chen, R.G.; Gong, Z.H. Overexpression of the CaTIP1-1 pepper gene in tobacco enhances resistance to osmotic stresses. Int. J. Mol. Sci. 2014, 15, 20101–20116. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Ito, T.M.; Trevizan, C.B.; Santos, T.B.D.; Souza, S.G.H.D. Genome-wide identification and characterization of the Dof transcription factor gene family in Phaseolus vulgaris L. Am. J Plant Sci. 2017, 8, 3233–3257. [Google Scholar] [CrossRef]
- Ayaz, A.; Saqib, S.; Huang, H.D.; Zaman, W.; Lu, S.Y.; Zhao, H.Y. Genome-wide comparative analysis of long-chain acyl-CoA synthetases (LACSs) gene family: A focus on identification, evolution and expression profiling related to lipid synthesis. Plant Physiol. Biochem. 2021, 161, 1–11. [Google Scholar] [CrossRef]
- Dimitrios, T. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Rio, D.D.; Stewart, A.J.; Pellegrini, N. A review of recent studies on malondialdehyde as toxic molecule and biological marker of oxidative stress. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 316–328. [Google Scholar] [CrossRef]
- Kazemi-Shahandashti, S.S.; Maali-Amiri, R. Global insights of protein responses to cold stress in plants: Signaling, defence, and degradation. J. Plant Physiol. 2018, 226, 123–135. [Google Scholar] [CrossRef]
- Baxter, A.; Suzuki, N.; Mittler, R. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar] [CrossRef]
- Liu, W.C.; Song, R.F.; Qiu, Y.M.; Zheng, S.Q.; Li, T.T.; Wu, Y.; Song, C.P.; Lu, Y.T.; Yuan, H.M. Sulfenylation of ENOLASE2 facilitates H2O2-conferred freezing tolerance in Arabidopsis. Dev. Cell 2022, 57, 1883–1898. [Google Scholar] [CrossRef]
- Guo, Q.; Han, J.; Li, C.; Hou, X.; Zhao, C.; Wang, Q.; Wu, J.; Mur, L.A.J. Defining key metabolic roles in osmotic adjustment and ROS homeostasis in the recretohalophyte Karelinia caspia under salt stress. Physiol. Plant. 2022, 174, e13663. [Google Scholar] [CrossRef] [PubMed]
- Huchzermeyer, B.; Menghani, E.; Khardia, P.; Shilu, A. Metabolic Pathway of Natural Antioxidants, Antioxidant Enzymes and ROS Providence. Antioxidants 2022, 11, 761. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.H.; Chong, P.F.; Liu, Z.H.; Bao, X.G.; Tan, B.B. Exogenous hydrogen sulfide improves salt stress tolerance of Reaumuria soongorica seedlings by regulating active oxygen metabolism. PeerJ 2023, 24, e15881. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.T.; Ding, Y.L.; Yang, S.H. Molecular regulation of CBF signaling in cold acclimation. Trends Plant Sci. 2018, 23, 623–637. [Google Scholar] [CrossRef]
- Tang, B.Q.; Xie, L.L.; Yang, H.P.; Li, X.M.; Chen, Y.; Zou, X.X.; Liu, F.; Dai, X.G. Analysis of the expression and function of key genes in pepper under low-temperature stress. Front Plant Sci. 2022, 13, 852511. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Protein ID | ORF (bp) | Protein Length | Molecular Mass (KDa) | PI | Instability Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| CaLACS1 | LOC107855307 | NP_001311647.1 | 1977 | 658 | 73.56 | 6.90 | 33.10 | Cytoplasmic |
| CaLACS2 | LOC107840375 | XP_016539704.1 | 1992 | 663 | 75.11 | 6.50 | 38.29 | Cytoplasmic |
| CaLACS3 | LOC107851776 | XP_016552345.1 | 2016 | 671 | 74.88 | 7.76 | 39.70 | Cytoplasmic |
| CaLACS4 | LOC107864092 | XP_016565840.2 | 2094 | 697 | 77.19 | 8.44 | 37.90 | Mitochondrial |
| CaLACS5 | LOC107868016 | XP_016570047.1 | 2070 | 689 | 75.43 | 7.15 | 30.91 | Cytoplasmic |
| CaLACS6 | LOC107878096 | NP_001312030.1 | 2181 | 726 | 78.37 | 8.54 | 25.06 | Cytoplasmic |
| CaLACS7 | LOC107839318 | XP_016538238.1 | 1980 | 659 | 73.70 | 5.87 | 24.88 | Cytoplasmic |
| CaLACS8 | LOC107854884 | XP_047250631.1 | 2037 | 678 | 75.03 | 6.48 | 32.99 | Plasma Membrane |
| CaLACS9 | LOC107840916 | XP_016540352.2 | 2163 | 720 | 79.84 | 7.49 | 28.59 | Chloroplast |
| Species | Homologous Gene Pairs | Ka | Ks | Ka/Ks | Divergent/ Duplication Time (Mya) |
|---|---|---|---|---|---|
| Pepper/ Arabidopsis | CaLACS1/AtLACS2 | 0.21 | 2.78 | 0.08 | 177.07 a |
| CaLACS1/AtLACS4 | 0.17 | 2.14 | 0.08 | 136.31 a | |
| CaLACS1/AtLACS5 | 0.18 | 1.88 | 0.10 | 119.75 a | |
| CaLACS2/AtLACS1 | 0.27 | - | - | - | |
| CaLACS3/AtLACS2 | 0.23 | 2.11 | 0.11 | 134.39 a | |
| CaLACS3/AtLACS4 | 0.19 | 3.08 | 0.06 | 196.18 a | |
| CaLACS3/AtLACS5 | 0.19 | 2.11 | 0.09 | 134.39 a | |
| CaLACS4/AtLACS1 | - | - | - | - | |
| CaLACS7/AtLACS2 | - | - | - | - | |
| CaLACS7/AtLACS4 | - | - | - | - | |
| CaLACS7/AtLACS5 | 0.26 | 2.58 | 0.10 | 164.33 a | |
| CaLACS8/AtLACS8 | 1.81 | - | - | - | |
| Pepper/Pepper | CaLACS1/CaLACS3 | 0.11 | 0.70 | 0.16 | 44.59 b |
| CaLACS1/CaLACS7 | 0.23 | 2.21 | 0.10 | 140.76 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Chen, Y.; He, J.; Wang, D.; Jiang, Y.; Chen, X.; Yang, Q.; Li, H. Genome-Wide Identification of LACS Family Genes and Functional Characterization of CaLACS6/9 in Response to Cold Stress in Pepper (Capsicum annuum L.). Agronomy 2025, 15, 970. https://doi.org/10.3390/agronomy15040970
Zhang J, Chen Y, He J, Wang D, Jiang Y, Chen X, Yang Q, Li H. Genome-Wide Identification of LACS Family Genes and Functional Characterization of CaLACS6/9 in Response to Cold Stress in Pepper (Capsicum annuum L.). Agronomy. 2025; 15(4):970. https://doi.org/10.3390/agronomy15040970
Chicago/Turabian StyleZhang, Jianwei, Yue Chen, Jing He, Dong Wang, Yao Jiang, Xianjun Chen, Qin Yang, and Huanxiu Li. 2025. "Genome-Wide Identification of LACS Family Genes and Functional Characterization of CaLACS6/9 in Response to Cold Stress in Pepper (Capsicum annuum L.)" Agronomy 15, no. 4: 970. https://doi.org/10.3390/agronomy15040970
APA StyleZhang, J., Chen, Y., He, J., Wang, D., Jiang, Y., Chen, X., Yang, Q., & Li, H. (2025). Genome-Wide Identification of LACS Family Genes and Functional Characterization of CaLACS6/9 in Response to Cold Stress in Pepper (Capsicum annuum L.). Agronomy, 15(4), 970. https://doi.org/10.3390/agronomy15040970