

Impacts of Cereal and Legume Cultivation on Soil Properties and Microbial Communities in the Mu Us Desert


Abstract
1. Introduction
2. Materials and Methods
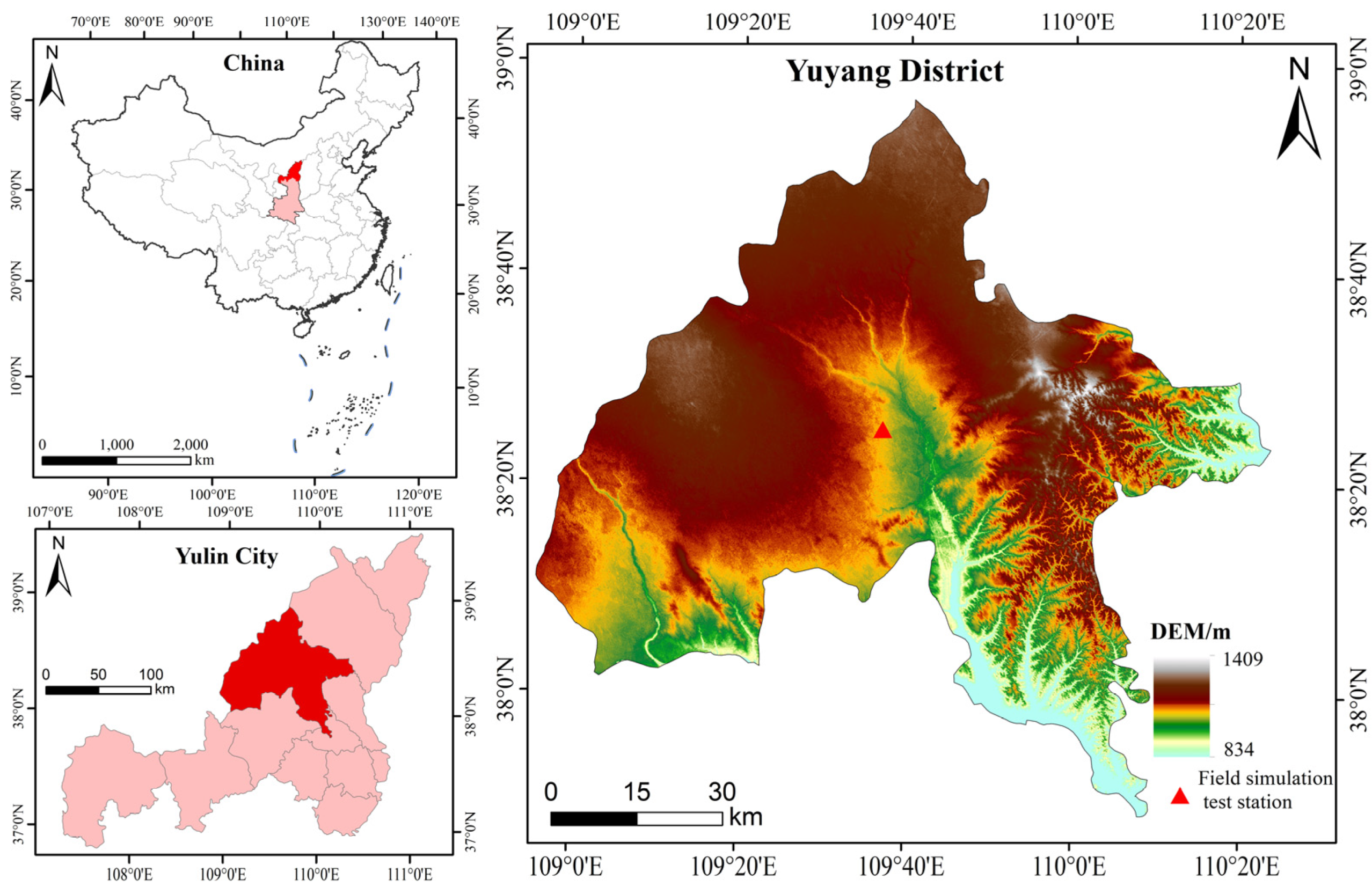
2.1. Design of the Experiment
2.2. Experimental Design and Sample Collection
2.3. Determination of Soil Chemical Properties
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Statistical Analysis of Data
3. Results
3.1. Inter-Root Soil Chemistry of Different Crops
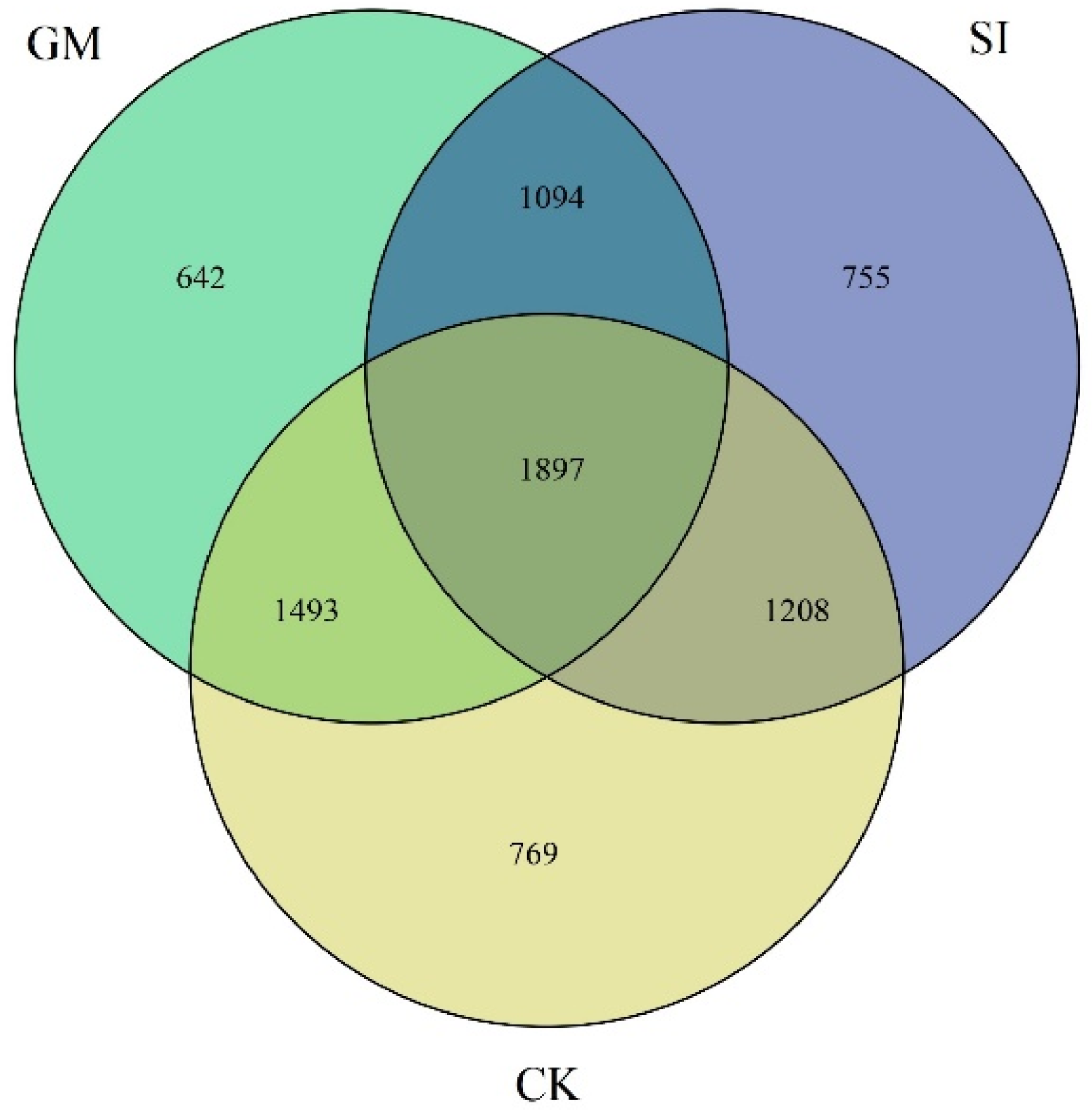
3.2. Venn Diagram Analysis of Inter-Root Soil Bacterial Composition
3.3. Bacterial Richness and Diversity in Inter-Root Soil
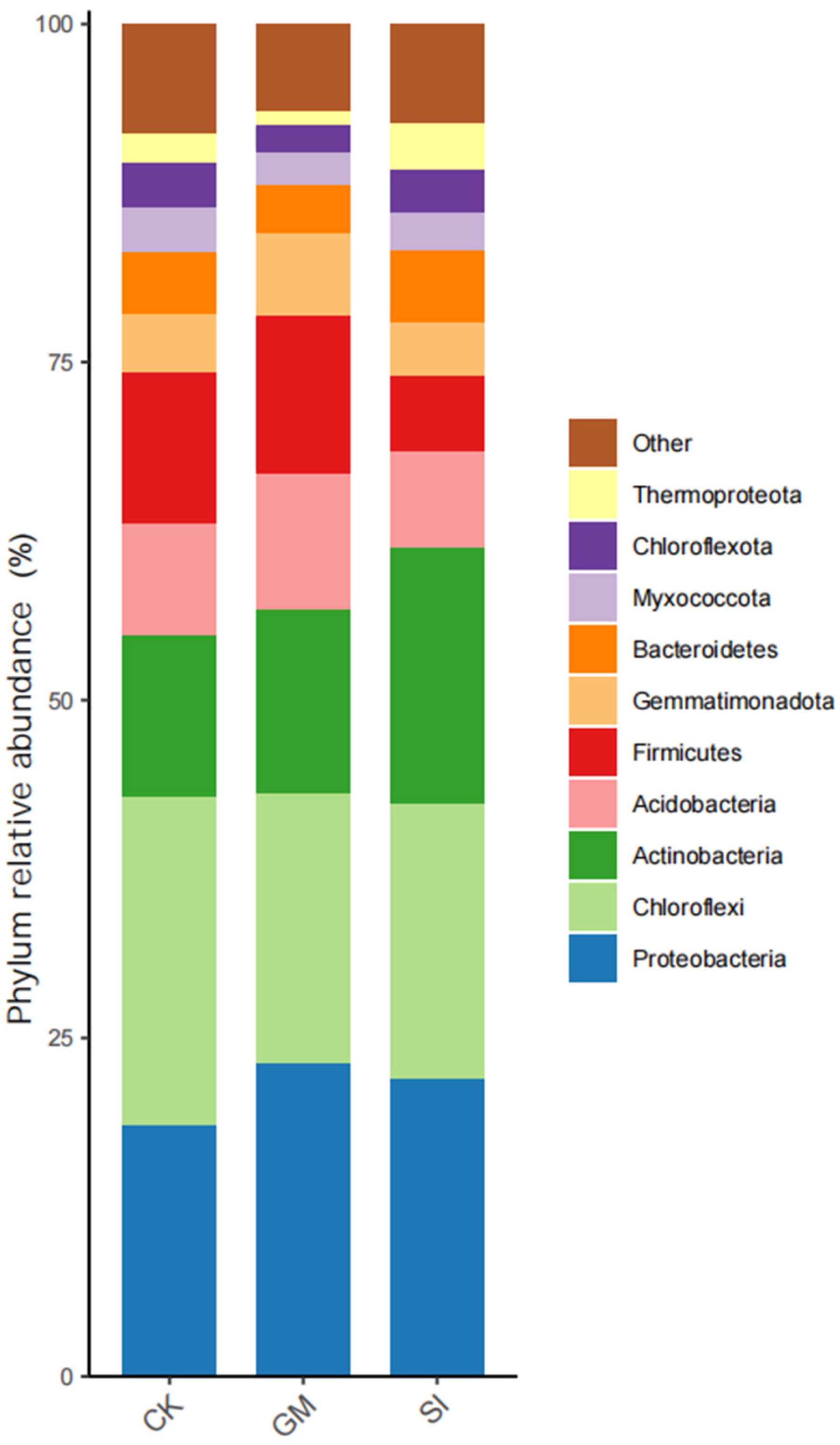
3.4. Differential Analysis of Inter-Root Soil Bacterial Community Composition
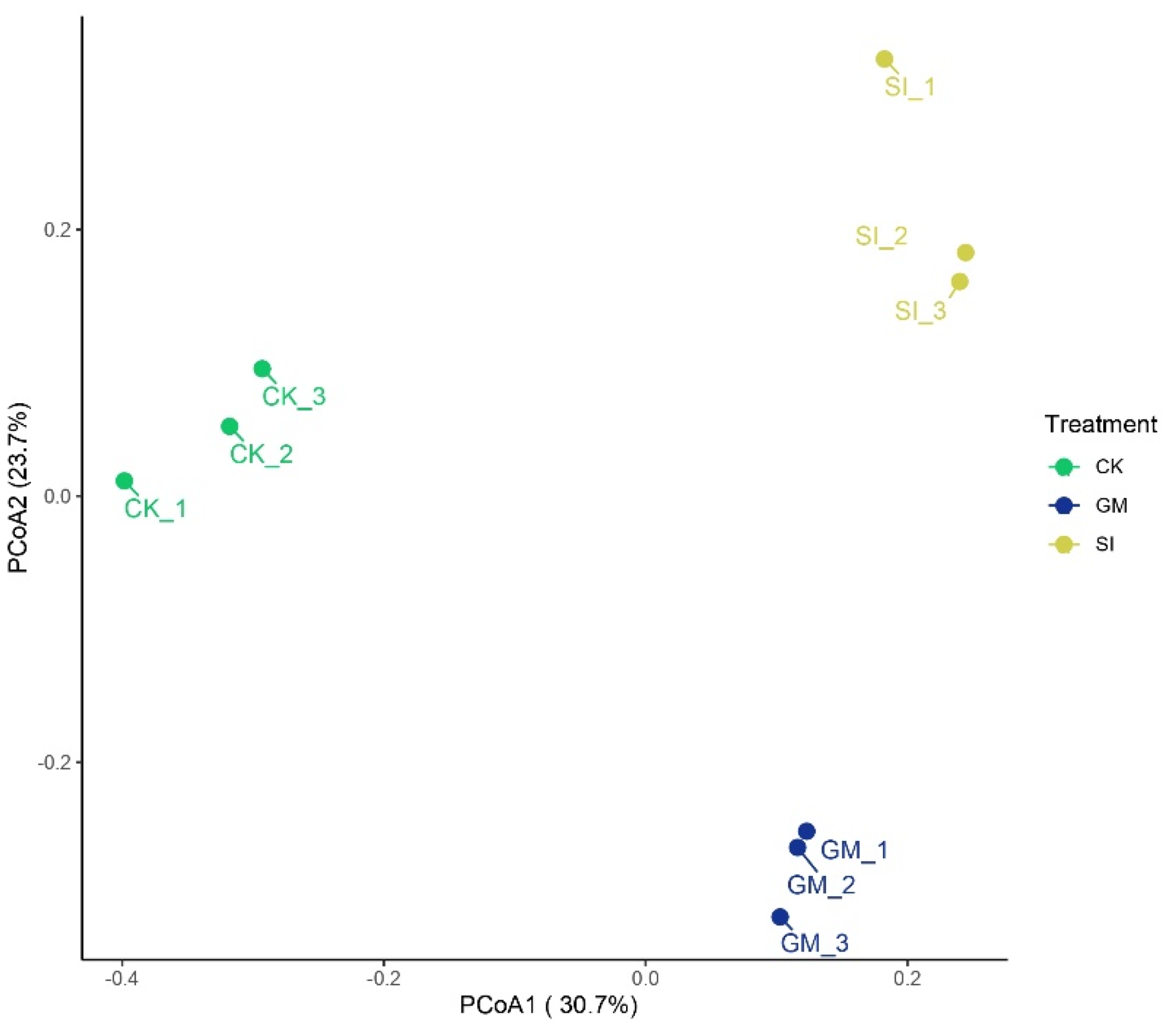
3.5. Analysis of Inter-Root Soil β-Diversity and Intergroup Variability
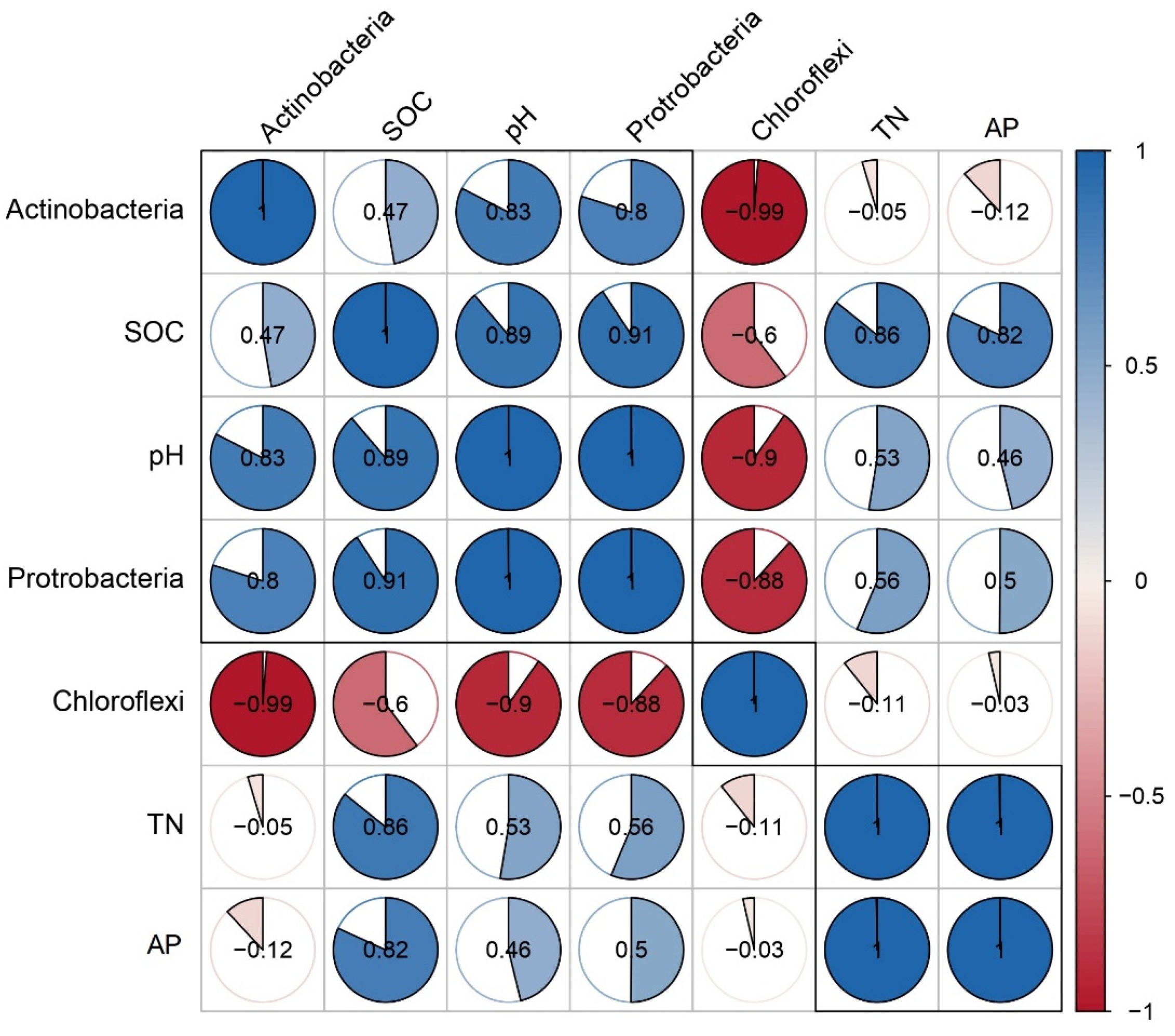
3.6. Correlation Between Soil Chemistry and Microbial Structure and Diversity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Runnström, M.C. Rangeland Development of the Mu Us Sandy Land in Semiarid China: An Analysis Using Landsat and NOAA Remote Sensing Data. Land Degrad. Dev. 2003, 14, 189–202. [Google Scholar] [CrossRef]
- Wang, X.; Chen, F.-H.; Dong, Z.; Xia, D. Evolution of the Southern Mu US Desert in North China over the Past 50 Years: An Analysis Using Proxies of Human Activity and Climate Parameters. Land Degrad. Dev. 2005, 16, 351–366. [Google Scholar] [CrossRef]
- Wang, N.; Xie, J.; Han, J. A Sand Control and Development Model in Sandy Land Based on Mixed Experiments of Arsenic Sandstone and Sand: A Case Study in Mu Us Sandy Land in China. Chin. Geogr. Sci. 2013, 23, 700–707. [Google Scholar] [CrossRef]
- Han, X.; Jia, G.; Yang, G.; Wang, N.; Liu, F.; Chen, H.; Guo, X.; Yang, W.; Liu, J. Spatiotemporal Dynamic Evolution and Driving Factors of Desertification in the Mu Us Sandy Land in 30 Years. Sci. Rep. 2020, 10, 21734. [Google Scholar] [CrossRef]
- Zhang, D.; Deng, H. Historical Human Activities Accelerated Climate-Driven Desertification in China’s Mu Us Desert. Sci. Total Environ. 2020, 708, 134771. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yan, C.; Wang, T.; Du, H. Monitoring Grassland Reclamation in the Mu Us Desert Using Remote Sensing from 2010 to 2015. Envorin. Earth Sci. 2019, 78, 311. [Google Scholar] [CrossRef]
- Horn, R.; Domżżał, H.; Słowińska-Jurkiewicz, A.; van Ouwerkerk, C. Soil Compaction Processes and Their Effects on the Structure of Arable Soils and the Environment. Soil Tillage Res. 1995, 35, 23–36. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, B.-Y.; Liu, S.-L.; Qi, J.-Y.; Wang, X.; Pu, C.; Li, S.-S.; Zhang, X.-Z.; Yang, X.-G.; Lal, R.; et al. Sustaining Crop Production in China’s Cropland by Crop Residue Retention: A Meta-Analysis. Land Degrad. Dev. 2020, 31, 694–709. [Google Scholar] [CrossRef]
- Bethany, J.; Giraldo-Silva, A.; Nelson, C.; Barger, N.N.; Garcia-Pichel, F. Optimizing the Production of Nursery-Based Biological Soil Crusts for Restoration of Arid Land Soils. Appl. Environ. Microbiol. 2019, 85, e00735-19. [Google Scholar] [CrossRef]
- Naorem, A.; Jayaraman, S.; Dang, Y.P.; Dalal, R.C.; Sinha, N.K.; Rao, C.S.; Patra, A.K. Soil Constraints in an Arid Environment—Challenges, Prospects, and Implications. Agronomy 2023, 13, 220. [Google Scholar] [CrossRef]
- Dasgupta, D.; Brahmaprakash, G.P. Soil Microbes Are Shaped by Soil Physico-Chemical Properties: A Brief Review of Existing Literature. Int. J. Plant Soil Sci. 2021, 33, 59–71. [Google Scholar] [CrossRef]
- Soong, J.L.; Fuchslueger, L.; Marañon-Jimenez, S.; Torn, M.S.; Janssens, I.A.; Penuelas, J.; Richter, A. Microbial Carbon Limitation: The Need for Integrating Microorganisms into Our Understanding of Ecosystem Carbon Cycling. Glob. Change Biol. 2020, 26, 1953–1961. [Google Scholar] [CrossRef]
- Yadav, A.N.; Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Dikilitas, M.; Abdel-Azeem, A.M.; Ahluwalia, A.S.; Saxena, A.K. Biodiversity, and Biotechnological Contribution of Beneficial Soil Microbiomes for Nutrient Cycling, Plant Growth Improvement and Nutrient Uptake. Biocatal. Agric. Biotechnol. 2021, 33, 102009. [Google Scholar] [CrossRef]
- Bhattacharyya, S.S.; Ros, G.H.; Furtak, K.; Iqbal, H.M.N.; Parra-Saldívar, R. Soil Carbon Sequestration—An Interplay between Soil Microbial Community and Soil Organic Matter Dynamics. Sci. Total Environ. 2022, 815, 152928. [Google Scholar] [CrossRef]
- Wang, J.L.; Liu, K.L.; Zhao, X.Q.; Zhang, H.Q.; Li, D.; Li, J.J.; Shen, R.F. Balanced Fertilization over Four Decades Has Sustained Soil Microbial Communities and Improved Soil Fertility and Rice Productivity in Red Paddy Soil. Sci. Total Environ. 2021, 793, 148664. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and Soil Microbial Community: A Review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Ek, L. Replenishment of Essential Natural Resources: An Assessment of the Ability of Freshwater Algae to Enhance Crop Quality and Soil Health. Ph.D. Thesis, University of South Carolina, Columbia, SC, USA, 2023. [Google Scholar]
- Adedayo, A.A.; Babalola, O.O. The Potential of Biostimulants on Soil Microbial Community: A Review. Front. Ind. Microbiol. 2023, 1, 1308641. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, Z.; Sun, C. Differences in Carbon Sequestration Ability of Diverse Tartary Buckwheat Genotypes in Barren Soil Caused by Microbial Action. Int. J. Environ. Res. Public Health 2023, 20, 959. [Google Scholar] [CrossRef]
- Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S.P.P. Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants 2023, 12, 3102. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Q.; Ju, M.; Yan, S.; Zhang, Q.; Gu, P. The Endophytic Fungi Diversity, Community Structure, and Ecological Function Prediction of Sophora Alopecuroides in Ningxia, China. Microorganisms 2022, 10, 2099. [Google Scholar] [CrossRef]
- Tang, K.; Liang, Y.; Yuan, B.; Meng, J.; Feng, F. Spatial Distribution and Core Community of Diazotrophs in Biological Soil Crusts and Subsoils in Temperate Semi-Arid and Arid Deserts of China. Front. Microbiol. 2023, 14, 1074855. [Google Scholar] [CrossRef]
- Islam, W.; Zeng, F.; Alotaibi, M.O.; Khan, K.A. Unlocking the Potential of Soil Microbes for Sustainable Desertification Management. Earth-Sci. Rev. 2024, 252, 104738. [Google Scholar] [CrossRef]
- Chen, Y.; Tian, W.; Shao, Y.; Li, Y.-J.; Lin, L.-A.; Zhang, Y.-J.; Han, H.; Chen, Z.-J. Miscanthus Cultivation Shapes Rhizosphere Microbial Community Structure and Function as Assessed by Illumina MiSeq Sequencing Combined with PICRUSt and FUNGUIld Analyses. Arch. Microbiol. 2020, 202, 1157–1171. [Google Scholar] [CrossRef]
- Kumar, V.; Singh, K.; Shah, M.P.; Singh, A.K.; Kumar, A.; Kumar, Y. Chapter 1—Application of Omics Technologies for Microbial Community Structure and Function Analysis in Contaminated Environment. In Wastewater Treatment; Shah, M.P., Sarkar, A., Mandal, S., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; pp. 1–40. ISBN 978-0-12-821881-5. [Google Scholar]
- Jiang, J.; Hu, X.; Ji, X.; Chen, H. High Throughput Sequencing Technology Facility Research of Genomic Modification Crop Cultivation Influencing Soil Microbe. Front. Plant Sci. 2023, 14, 1208111. [Google Scholar] [CrossRef]
- Yang, P.; Luo, Y.; Gao, Y.; Gao, X.; Gao, J.; Wang, P.; Feng, B. Soil Properties, Bacterial and Fungal Community Compositions and the Key Factors after 5-Year Continuous Monocropping of Three Minor Crops. PLoS ONE 2020, 15, e0237164. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Lupwayi, N.; Marc, S.-A.; Siddique, K.H.M.; Bainard, L.D. Anthropogenic Drivers of Soil Microbial Communities and Impacts on Soil Biological Functions in Agroecosystems. Glob. Ecol. Conserv. 2021, 27, e01521. [Google Scholar] [CrossRef]
- Khmelevtsova, L.E.; Sazykin, I.S.; Azhogina, T.N.; Sazykina, M.A. Influence of Agricultural Practices on Bacterial Community of Cultivated Soils. Agriculture 2022, 12, 371. [Google Scholar] [CrossRef]
- Chamberlain, L.A.; Bolton, M.L.; Cox, M.S.; Suen, G.; Conley, S.P.; Ané, J.-M. Crop Rotation, but Not Cover Crops, Influenced Soil Bacterial Community Composition in a Corn-Soybean System in Southern Wisconsin. Appl. Soil Ecol. 2020, 154, 103603. [Google Scholar] [CrossRef]
- Town, J.R.; Gregorich, E.G.; Drury, C.F.; Lemke, R.; Phillips, L.A.; Helgason, B.L. Diverse Crop Rotations Influence the Bacterial and Fungal Communities in Root, Rhizosphere and Soil and Impact Soil Microbial Processes. Appl. Soil Ecol. 2022, 169, 104241. [Google Scholar] [CrossRef]
- Zhao, Y.; Yao, Y.; Xu, H.; Xie, Z.; Guo, J.; Qi, Z.; Jiang, H. Soil Metabolomics and Bacterial Functional Traits Revealed the Responses of Rhizosphere Soil Bacterial Community to Long-Term Continuous Cropping of Tibetan Barley. PeerJ 2022, 10, e13254. [Google Scholar] [CrossRef]
- Gong, X.; Feng, Y.; Dang, K.; Jiang, Y.; Qi, H.; Feng, B. Linkages of Microbial Community Structure and Root Exudates: Evidence from Microbial Nitrogen Limitation in Soils of Crop Families. Sci. Total Environ. 2023, 881, 163536. [Google Scholar] [CrossRef]
- Ji, L.; Si, H.; He, J.; Fan, L.; Li, L. The Shifts of Maize Soil Microbial Community and Networks Are Related to Soil Properties under Different Organic Fertilizers. Rhizosphere 2021, 19, 100388. [Google Scholar] [CrossRef]
- Xu, C.; Li, Y.; Hu, X.; Zang, Q.; Zhuang, H.; Huang, L. The Influence of Organic and Conventional Cultivation Patterns on Physicochemical Property, Enzyme Activity and Microbial Community Characteristics of Paddy Soil. Agriculture 2022, 12, 121. [Google Scholar] [CrossRef]
- Wongkiew, S.; Chaikaew, P.; Takrattanasaran, N.; Khamkajorn, T. Evaluation of Nutrient Characteristics and Bacterial Community in Agricultural Soil Groups for Sustainable Land Management. Sci. Rep. 2022, 12, 7368. [Google Scholar] [CrossRef] [PubMed]
- Zhouchang, Y.; Wei, Z.; Liyun, Z.; Shihai, M.; Lin, S.; Yushu, L.; Ying, Z.; AHejiang, S.; Jiayi, S.; Ling, C.; et al. Efficient Vegetation Restoration in Mu Us Desert Reduces Microbial Diversity Due to the Transformation of Nutrient Requirements. Ecol. Indic. 2023, 154, 110758. [Google Scholar] [CrossRef]
- Wang, L.; Li, X. Soil Microbial Community and Their Relationship with Soil Properties across Various Landscapes in the Mu Us Desert. Forests 2023, 14, 2152. [Google Scholar] [CrossRef]
- Song, D.; Hu, Z.; Zeng, J.; Sun, H. Influence of Mining on Vegetation in Semi-Arid Areas of Western China Based on the Coupling of above Ground and below Ground—A Case Study of Daliuta Coalfield. Ecol. Indic. 2024, 161, 111964. [Google Scholar] [CrossRef]
- Xiao, N.; Wang, Y.; Guo, Z.; Shao, T.; Dong, Z.; Xing, B. Tire Plastic and Road-Wear Particles on Yujing Expressway in the Restoration Area of Mu Us Sandy Land: Occurrence Characteristics and Ecological Risk Screening. J. Hazard. Mater. 2024, 468, 133860. [Google Scholar] [CrossRef]
- Rossini, F.; Virga, G.; Loreti, P.; Iacuzzi, N.; Ruggeri, R.; Provenzano, M.E. Hops (Humulus lupulus L.) as a Novel Multipurpose Crop for the Mediterranean Region of Europe: Challenges and Opportunities of Their Cultivation. Agriculture 2021, 11, 484. [Google Scholar] [CrossRef]
- Gebretsadikan, T.; Munro, P.; Forge, T.A.; Jones, M.D.; Nelson, L.M. Mulching Improved Soil Fertility, Plant Growth and Productivity, and Postharvest Deficit Irrigation Reduced Water Use in Sweet Cherry Orchards in a Semi-Arid Region. Arch. Agron. Soil Sci. 2023, 69, 1419–1436. [Google Scholar] [CrossRef]
- Jing, X.; Chen, X.; Fang, J.; Ji, C.; Shen, H.; Zheng, C.; Zhu, B. Soil Microbial Carbon and Nutrient Constraints Are Driven More by Climate and Soil Physicochemical Properties than by Nutrient Addition in Forest Ecosystems. Soil Biol. Biochem. 2020, 141, 107657. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.; Feng, Y. The Effects of Biochar Addition on Soil Physicochemical Properties: A Review. Catena 2021, 202, 105284. [Google Scholar] [CrossRef]
- Matteoli, F.P.; Silva, A.M.M.; Feiler, H.P.; de Araújo, V.L.V.P.; Cardoso, E.J.B.N. Predicting soil farming system and attributes based on soil bacterial community. Appl. Soil Ecol. 2022, 171, 104335. [Google Scholar] [CrossRef]
- Ding, Z.; Lu, R.; Wang, L.; Yu, L.; Liu, X.; Liu, Y.; Liu, L.; Bai, M.; Wang, S. Early-Mid Holocene Climatic Changes Inferred from Colors of Eolian Deposits in the Mu Us Desert. Geoderma 2021, 401, 115172. [Google Scholar] [CrossRef]
- Guo, Z.; Li, J.; Zhang, Y.; Wang, H.; Li, W. The Soft Rock Can Promote the Improvement of Aeolian Sandy Soil in Mu Us Sandy Land, China. Sci. Rep. 2023, 13, 11813. [Google Scholar] [CrossRef]
- Zhang, J.; Cheng, T.; Shi, L.; Wang, W.; Niu, Z.; Guo, W.; Ma, X. Combining Spectral and Texture Features of UAV Hyperspectral Images for Leaf Nitrogen Content Monitoring in Winter Wheat. Int. J. Remote Sens. 2022, 43, 2335–2356. [Google Scholar] [CrossRef]
- Xu, Y.; Pu, L.; Zhang, R.; Zhu, M.; Zhang, M.; Bu, X.; Xie, X.; Wang, Y. Effects of Agricultural Reclamation on Soil Physicochemical Properties in the Mid-Eastern Coastal Area of China. Land 2021, 10, 142. [Google Scholar] [CrossRef]
- van der Heyde, M.; Bunce, M.; Dixon, K.; Wardell-Johnson, G.; White, N.E.; Nevill, P. Changes in Soil Microbial Communities in Post Mine Ecological Restoration: Implications for Monitoring Using High Throughput DNA Sequencing. Sci. Total Environ. 2020, 749, 142262. [Google Scholar] [CrossRef]
- Yin, D.; Li, H.; Wang, H.; Guo, X.; Wang, Z.; Lv, Y.; Ding, G.; Jin, L.; Lan, Y. Impact of Different Biochars on Microbial Community Structure in the Rhizospheric Soil of Rice Grown in Albic Soil. Molecules 2021, 26, 4783. [Google Scholar] [CrossRef]
- Iturbe-Espinoza, P.; Brandt, B.W.; Braster, M.; Bonte, M.; Brown, D.M.; van Spanning, R.J.M. Effects of DNA Preservation Solution and DNA Extraction Methods on Microbial Community Profiling of Soil. Folia Microbiol. 2021, 66, 597–606. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing Taxonomic Classification of Marker-Gene Amplicon Sequences with QIIME 2’s Q2-Feature-Classifier Plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Salehi, F.; Inanloodoghouz, M.; Ghazvineh, S. Influence of Microwave Pretreatment on the Total Phenolics, Antioxidant Activity, Moisture Diffusivity, and Rehydration Rate of Dried Sweet Cherry. Food Sci. Nutr. 2023, 11, 7870–7876. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Zhou, Y. Effects of Land Use Patterns on the Bacterial Community Structure and Diversity of Wetland Soils in the Sanjiang Plain. J. Soil Sci. Plant Nutr. 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Wang, Z.; Bai, Y.; Hou, J.; Li, F.; Li, X.; Cao, R.; Deng, Y.; Wang, H.; Jiang, Y.; Yang, W. The Changes in Soil Microbial Communities across a Subalpine Forest Successional Series. Forests 2022, 13, 289. [Google Scholar] [CrossRef]
- Jiang, R.; Wang, M.; Chen, W.; Li, X.; Balseiro-Romero, M. Changes in the Integrated Functional Stability of Microbial Community under Chemical Stresses and the Impacting Factors in Field Soils. Ecol. Indic. 2020, 110, 105919. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, L.; Zhang, Y.-G.; Tu, T.-Y.; Wang, S.-T.; Shen, C.-H.; Yuan, H.-W.; Zhong, X.-Z. A Comparison of Microbial Communities and Volatile Compounds in Wheat Qu from Different Geographic Locations. LWT 2021, 148, 111752. [Google Scholar] [CrossRef]
- Siddiqui, A.U.; Jain, M.K.; Masto, R.E. Pollution Evaluation, Spatial Distribution, and Source Apportionment of Trace Metals around Coal Mines Soil: The Case Study of Eastern India. Environ. Sci. Pollut. Res. 2020, 27, 10822–10834. [Google Scholar] [CrossRef]
- Xu, H.; Croot, P.; Zhang, C. Exploration of the Spatially Varying Relationships between Lead and Aluminium Concentrations in the Topsoil of Northern Half of Ireland Using Geographically Weighted Pearson Correlation Coefficient. Geoderma 2022, 409, 115640. [Google Scholar] [CrossRef]
- Thepbandit, W.; Athinuwat, D. Rhizosphere Microorganisms Supply Availability of Soil Nutrients and Induce Plant Defense. Microorganisms 2024, 12, 558. [Google Scholar] [CrossRef]
- Liang, Y.; Al-Kaisi, M.; Yuan, J.; Liu, J.; Zhang, H.; Wang, L.; Cai, H.; Ren, J. Effect of Chemical Fertilizer and Straw-Derived Organic Amendments on Continuous Maize Yield, Soil Carbon Sequestration and Soil Quality in a Chinese Mollisol. Agric. Ecosyst. Environ. 2021, 314, 107403. [Google Scholar] [CrossRef]
- Tang, H.; Cheng, K.; Shi, L.; Li, C.; Wen, L.; Li, W.; Sun, M.; Sun, G.; Long, Z. Effects of Long-Term Organic Matter Application on Soil Carbon Accumulation and Nitrogen Use Efficiency in a Double-Cropping Rice Field. Environ. Res. 2022, 213, 113700. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.; Pan, Y.; Wang, P.; Ran, L.; Qin, G.; Li, Q.; Kang, P. Response of Phyllosphere and Rhizosphere Microbial Communities to Salt Stress of Tamarix Chinensis. Plants 2024, 13, 1091. [Google Scholar] [CrossRef]
- Kuang, W.; Chen, W.; Lei, C.; Dai, Y.; Tian, X.; Tang, S.; Qian, Q.; Zhang, C.; Fu, L.; Zhou, G.; et al. Diversity of Endophytic Bacterial Community in Rice Roots and Their Roles in Phosphate Solubilization and Plant Growth. Rhizosphere 2024, 30, 100877. [Google Scholar] [CrossRef]
- Dellagi, A.; Quillere, I.; Hirel, B. Beneficial Soil-Borne Bacteria and Fungi: A Promising Way to Improve Plant Nitrogen Acquisition. J. Exp. Bot. 2020, 71, 4469–4479. [Google Scholar] [CrossRef]
- Scavo, A.; Fontanazza, S.; Restuccia, A.; Pesce, G.R.; Abbate, C.; Mauromicale, G. The Role of Cover Crops in Improving Soil Fertility and Plant Nutritional Status in Temperate Climates. A Review. Agron. Sustain. Dev. 2022, 42, 1–25. [Google Scholar] [CrossRef]
- Zimmermann, B.; Claß-Mahler, I.; von Cossel, M.; Lewandowski, I.; Weik, J.; Spiller, A.; Nitzko, S.; Lippert, C.; Krimly, T.; Pergner, I.; et al. Mineral-Ecological Cropping Systems—A New Approach to Improve Ecosystem Services by Farming without Chemical Synthetic Plant Protection. Agronomy 2021, 11, 1710. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, G.; Ding, H.; Ci, D.; Dai, L.; Zhang, Z. Influence of Salt Stress on the Rhizosphere Soil Bacterial Community Structure and Growth Performance of Groundnut (Arachis hypogaea L.). Int. Microbiol. 2020, 23, 453–465. [Google Scholar] [CrossRef]
- Li, Y.; Chi, J.; Ao, J.; Gao, X.; Liu, X.; Sun, Y.; Zhu, W. Effects of Different Continuous Cropping Years on Bacterial Community and Diversity of Cucumber Rhizosphere Soil in Solar-Greenhouse. Curr. Microbiol. 2021, 78, 2380–2390. [Google Scholar] [CrossRef]
- Guo, J.; Wu, Y.; Wu, X.; Ren, Z.; Wang, G. Soil Bacterial Community Composition and Diversity Response to Land Conversion Is Depth-Dependent. Glob. Ecol. Conserv. 2021, 32, e01923. [Google Scholar] [CrossRef]
- Cui, J.; Zhu, R.; Wang, X.; Xu, X.; Ai, C.; He, P.; Liang, G.; Zhou, W.; Zhu, P. Effect of High Soil C/N Ratio and Nitrogen Limitation Caused by the Long-Term Combined Organic-Inorganic Fertilization on the Soil Microbial Community Structure and Its Dominated SOC Decomposition. J. Environ. Manag. 2022, 303, 114155. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | pH | Soil Organic Carbon (g·kg−1) | Total Nitrogen (g·kg−1) | Effective Phosphorus (g·kg−1) |
|---|---|---|---|---|
| CK | 8.02 ± 0.02 a | 13.58 ± 0.05 a | 0.28 ± 0.02 c | 247.4 ± 11.05 b |
| SI | 7.89 ± 0.02 b | 11.35 ± 0.04 b | 0.44 ± 0.02 b | 326.9 ± 13.05 a |
| GM | 7.95 ± 0.01 a | 12.67 ± 0.04 b | 0.55 ± 0.03 a | 253.2 ± 11.17 b |
| Treatment |
Observed Features |
Goods Coverage | Chao1 | Shannon | Simpson | ACE |
|---|---|---|---|---|---|---|
| CK | 2198.67 ± 66 a | 0.9982 ± 0.001 a | 2508.7 ± 162.13 b | 9.76 ± 0.15 b | 0.9983 ± 0.0018 a | 2583.2 ± 187.14 c |
| SI | 2164.67 ± 51 a | 0.9984 ± 0.002 a | 2693.8 ± 124.57 a | 10.07 ± 0.05 a | 0.9979 ± 0.0005 b | 3068.3 ± 108.87 a |
| GM | 2274.00 ± 37 a | 0.9980 ± 0.002 a | 2539.4 ± 245.18 a | 9.95 ± 0.21 a | 0.9961 ± 0.0016 b | 2723.5 ± 318.42 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, L.; Shi, L.; Wu, Y.; Wang, G.; Liu, G. Impacts of Cereal and Legume Cultivation on Soil Properties and Microbial Communities in the Mu Us Desert. Agronomy 2025, 15, 968. https://doi.org/10.3390/agronomy15040968
He L, Shi L, Wu Y, Wang G, Liu G. Impacts of Cereal and Legume Cultivation on Soil Properties and Microbial Communities in the Mu Us Desert. Agronomy. 2025; 15(4):968. https://doi.org/10.3390/agronomy15040968
Chicago/Turabian StyleHe, Lirong, Lei Shi, Yang Wu, Guoliang Wang, and Guobin Liu. 2025. "Impacts of Cereal and Legume Cultivation on Soil Properties and Microbial Communities in the Mu Us Desert" Agronomy 15, no. 4: 968. https://doi.org/10.3390/agronomy15040968
APA StyleHe, L., Shi, L., Wu, Y., Wang, G., & Liu, G. (2025). Impacts of Cereal and Legume Cultivation on Soil Properties and Microbial Communities in the Mu Us Desert. Agronomy, 15(4), 968. https://doi.org/10.3390/agronomy15040968