

Effect of Lighting Type on the Nitrates Concentration, Selective Bioactive Compounds and Yield of Sweet Basil (Ocimum basilicum L.) in Hydroponic Production

 , ,
, ,  ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
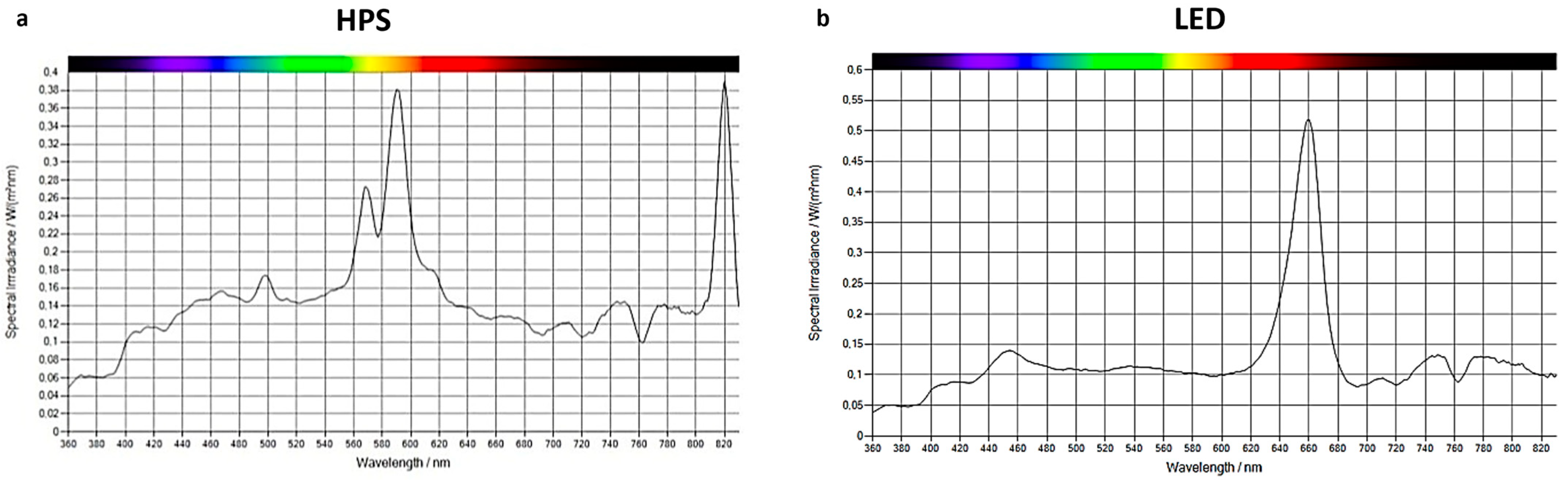
2.1. Experimental System and Plant Growing Conditions
2.2. Plant Growth
2.3. Yield and Dry Matter Content
2.4. Chemical Analyses
2.4.1. Nitrates, Macronutrients, Total Sugars (TS), Total Soluble Solids (TSS) and L-Ascorbic Acid (AA)
2.4.2. Chlorophylls, Carotenoids, SPAD Index
2.5. Statistical Analysis
3. Results
3.1. Plant Growth
3.2. Yield and Dry Matter Content
3.3. Chemical Analyses
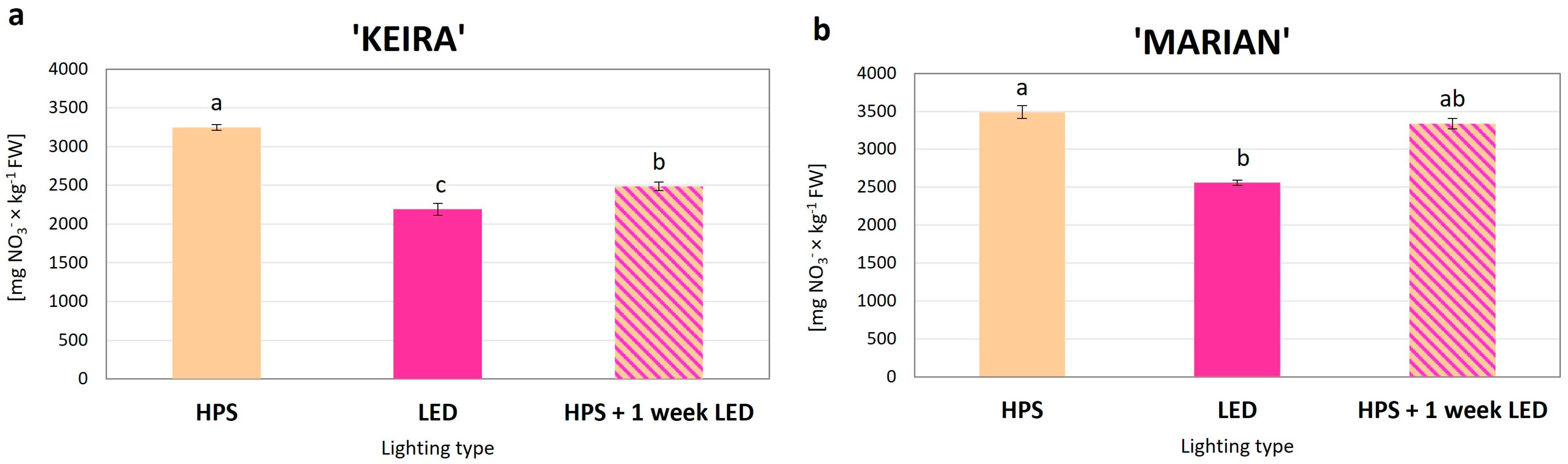
3.3.1. Nitrates
3.3.2. K, P, Ca, TS, TSS and AA
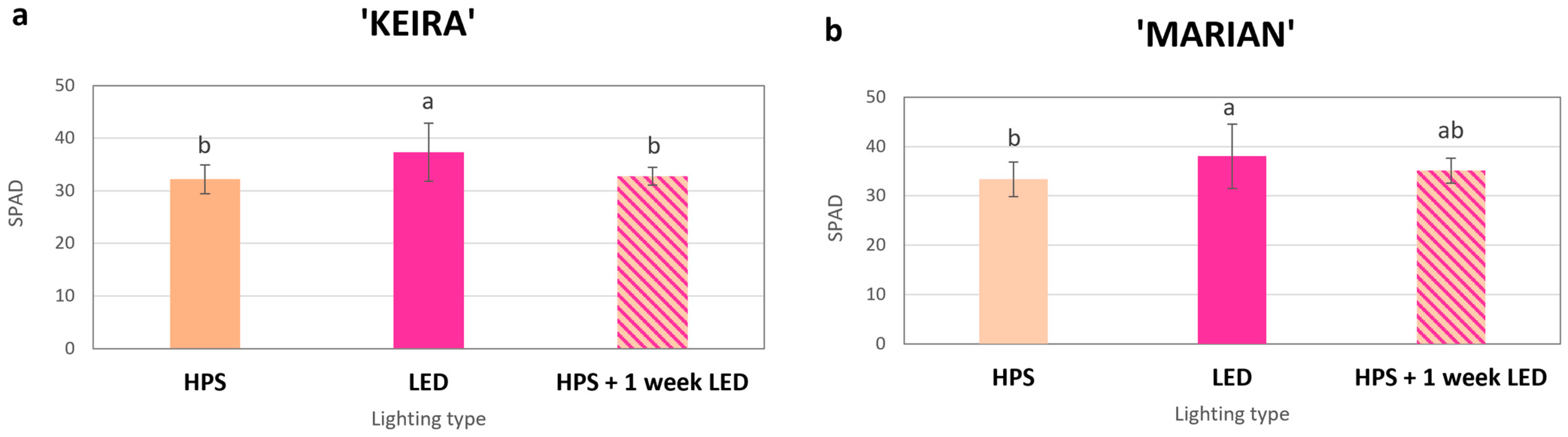
3.3.3. Chlorophylls and Carotenoids
4. Discussion
4.1. Basil Growth, Yield, and Dry Mass Content Under Different Types of Lighting
4.2. Effect of Lighting Type on Nitrate Content
4.3. Effect of Lighting Type on Macronutrients, TS, TSS and AA Content
4.4. Effect of Lighting Type on Chlorophylls and β-Carotene Content
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wolf, M.M.; Spittler, A.; Ahern, J. A profile of farmers’ market consumers and the perceived advantages of produce sold at farmers’ markets. J. Food Distrib. Res. 2005, 36, 192–201. [Google Scholar] [CrossRef]
- Giurgiu, R.M.; Morar, G.A.; Dumitras, A.; Boanca, P.; Duda, B.M.; Moldovan, C. Study regarding the suitability of cultivating medicinal plants in hydroponic systems in controlled environment. Res. J. Agric. Sci. 2014, 46, 84–93. [Google Scholar]
- Mattson, N.; Lieth, J.H. Liquid culture hydroponic system operation. In Soilless Culture (Theory and Practice), 2nd ed.; Raviv, M., Lieth, J.H., Bar-Tal, A., Eds.; Elsevier: Boston, MA, USA, 2019. [Google Scholar] [CrossRef]
- Cocetta, G.; Casciani, D.; Bulgari, R.; Musante, F.; Kołton, A.; Rossi, M.; Ferrante, A. Light use efficiency for vegetables production in protected and indoor environments. Eur. Phys. J. Plus 2017, 132, 43. [Google Scholar] [CrossRef]
- Rehman, M.; Ullah, S.; Bao, Y.; Wang, B.; Peng, D.; Liu, L. Light-emitting diodes: Whether an efficient source of light for indoor plants? Environ. Sci. Pollut. Res. Int. 2017, 24, 24743–24752. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Xu, Z.G.; Jiao, X.L.; Liu, X.Y. Effects of different illumination times of red and blue LED lights on growth and quality of Lactuca sativa var. ramosa. J. Plant Resour. Environ. 2017, 26, 113–115. [Google Scholar]
- Rahman, M.M.; Vasiliev, M.; Alameh, K. LED illumination spectrum manipulation for increasing the yield of sweet basil (Ocimum basilicum L.). Plants 2021, 10, 344. [Google Scholar] [CrossRef]
- Larsen, D.H.; Woltering, E.J.; Nicole, C.C.S.; Marcelis, L.F.M. Response of basil growth and morphology to light intensity and spectrum in a vertical farm. Front. Plant Sci. 2020, 11, 597906. [Google Scholar] [CrossRef] [PubMed]
- Eghbal, E.; Aliniaeifard, S.; Mehrjerdi, M.Z.; Abdi, S.; Hassani, S.B.; Rassaie, T.; Gruda, N.S. Growth, phytochemical, and phytohormonal responses of basil to different light durations and intensities under constant daily light integral. BMC Plant Biol. 2024, 24, 935. [Google Scholar] [CrossRef]
- Pennisi, G.; Orsini, F.; Landolfo, M.; Pistillo, A.; Crepaldi, A.; Nicola, S.; Fernández, J.A.; Marcelis, L.F.M.; Gianquinto, G. Optimal photoperiod for indoor cultivation of leafy vegetables and herbs. Eur. J. Hortic. Sci. 2020, 85, 329–338. [Google Scholar] [CrossRef]
- Walters, K.J.; Tarr, S.; Lopez, R.G. Modeling purple basil, sage, spearmint, and sweet basil responses to daily light integral and mean daily temperature. PLoS ONE 2023, 18, e0294905. [Google Scholar] [CrossRef]
- Barickman, T.C.; Olorunwa, O.J.; Sehgal, A.; Walne, C.H.; Reddy, K.R.; Gao, W. Yield, Physiological performance, and phytochemistry of basil (Ocimum basilicum L.) under temperature stress and elevated CO2 concentrations. Plants 2021, 10, 1072. [Google Scholar] [CrossRef]
- Pan, T.; Wang, Y.; Wang, L.; Ding, J.; Cao, Y.; Qin, G.; Yan, L.; Xi, L.; Zang, J.; Zou, Z. Increased CO2 and light intensity regulate growth and leaf gas exchange in tomato. Physiol. Plant. 2020, 168, 694–708. [Google Scholar] [CrossRef]
- Singh, H.; Poudel, M.R.; Dunn, B.L.; Fontanier, C.; Kakani, G. Effect of greenhouse CO2 supplementation on yield and mineral element concentrations of leafy greens grown using nutrient film technique. Agronomy 2020, 10, 323. [Google Scholar] [CrossRef]
- Solbach, J.A.; Fricke, A.; Stützel, H. Seasonal efficiency of supplemental LED lighting on growth and photomorphogenesis of sweet basil. Front. Plant Sci. 2021, 12, 609975. [Google Scholar] [CrossRef] [PubMed]
- Paucek, I.; Appolloni, E.; Pennisi, G.; Quaini, S.; Gianquinto, G.; Orsini, F. LED lighting systems for horticulture: Business growth and global distribution. Sustainability 2020, 12, 7516. [Google Scholar] [CrossRef]
- Nicole, C.C.S.; Charalambous, F.; Martinakos, S.; van de Voort, S.; Li, Z.; Verhoog, M.; Krijn, M. Lettuce growth and quality optimization in a plant factory. Acta Hortic. 2016, 1134, 231–238. [Google Scholar] [CrossRef]
- d’Aquino, L.; Cozzolino, R.; Nardone, G.; Borelli, G.; Gambale, E.; Sighicelli, M.; Menegoni, P.; Modarelli, G.C.; Rimauro, J.; Chianese, E.; et al. Effects of white and blue-red light on growth and metabolism of basil grown under microcosm conditions. Plants 2023, 12, 1450. [Google Scholar] [CrossRef] [PubMed]
- Sipos, L.; Balázs, L.; Székely, G.; Jung, A.; Sárosi, S.; Radácsi, P.; Csambalik, L. Optimization of basil (Ocimum basilicum L.) production in LED light environments—A review. Sci. Hortic. 2021, 289, 110486. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Aldarkazali, M.; Mohamed, S.J.; McMulkin, N.B.; Jbara, M.H.; Fuller, M.P. A novel new light recipe significantly increases the growth and yield of sweet basil (Ocimum basilicum) grown in a plant factory system. Agronomy 2020, 10, 934. [Google Scholar] [CrossRef]
- Appolloni, E.; Pennisi, G.; Zauli, I.; Carotti, L.; Paucek, I.; Quaini, S.; Orsini, F.; Gianquinto, G. Beyond vegetables: Effects of indoor LED light on specialized metabolite biosynthesis in medicinal and aromatic plants, edible flowers, and microgreens. J. Sci. Food Agric. 2022, 102, 472–487. [Google Scholar] [CrossRef]
- Bantis, F.; Ouzounis, T.; Radoglou, K. Artificial LED lighting enhances growth characteristics and total phenolic content of Ocimum basilicum, but variably affects transplant success. Sci. Hortic. 2016, 198, 277–283. [Google Scholar] [CrossRef]
- Karlsson, M. Light quality affects growth and nutrient content of basil. Acta Hortic. 2022, 1337, 157–164. [Google Scholar] [CrossRef]
- Pennisi, G.; Blasioli, S.; Cellini, A.; Maia, L.; Crepaldi, A.; Braschi, I.; Spinelli, F.; Nicola, S.; Fernandez, J.A.; Stanghellini, C.; et al. Unraveling the role of red:blue LED lights on resource use efficiency and nutritional properties of indoor grown sweet basil. Front. Plant Sci. 2019, 10, 305. [Google Scholar] [CrossRef]
- Lobiuc, A.; Vasilache, V.; Pintilie, O.; Stoleru, T.; Burducea, M.; Oroian, M.; Zamfirache, M.-M. Blue and red LED illumination improves growth and bioactive compounds contents in acyanic and cyanic Ocimum basilicum L. microgreens. Molecules 2017, 22, 2111. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.D.; Schwieterman, M.L.; Abrahan, C.E.; Colquhoun, T.A.; Folta, K.M. Light quality dependent changes in morphology, antioxidant capacity, and volatile production in sweet basil (Ocimum basilicum). Front. Plant Sci. 2016, 7, 1328. [Google Scholar] [CrossRef] [PubMed]
- El Haddaji, H.; Akodad, M.; Skalli, A.; Moumen, A.; Bellahcen, S.; Elhani, S.; Urrestarazu, M.; Kolar, M.; Imperl, J.; Petrova, P.; et al. Effects of light-emitting diodes (LEDs) on growth, nitrates and osmoprotectant content in microgreens of aromatic and medicinal plants. Horticulturae 2023, 9, 494. [Google Scholar] [CrossRef]
- Larsen, D.H.; Marcelis, L.F.M.; van Kempen, D.P.S.I.M.; Kohlen, W.; Nicole, C.; Woltering, E.J. Far-red light during cultivation improves postharvest chilling tolerance in basil. Postharvest Biol. Technol. 2023, 198, 112232. [Google Scholar] [CrossRef]
- Lillo, C. Signalling cascades integrating light-enhanced nitrate metabolism. Biochem. J. 2008, 415, 11–19. [Google Scholar] [CrossRef]
- Olas, B. The cardioprotective role of nitrate-rich vegetables. Foods 2024, 13, 691. [Google Scholar] [CrossRef]
- Karwowska, M.; Kononiuk, A. Nitrates/nitrites in food-risk for nitrosative stress and benefits. Antioxidants 2020, 9, 241. [Google Scholar] [CrossRef]
- Chang, W.H.; Chen, P.H.; Herianto, S.; Chen, H.L.; Lee, C.C. Aggregating exposures and toxicity equivalence approach into an integrated probabilistic dietary risk assessment for perchlorate, nitrate, and thiocyanate: Results from the National Food Monitoring Study and National Food Consumption Database. Environ. Res. 2022, 21, 112989. [Google Scholar] [CrossRef] [PubMed]
- Iammarino, M.; Di Taranto, A.; Cristino, M. Monitoring of nitrites and nitrates levels in leafy vegetables (spinach and lettuce): A contribution to risk assessment. J. Sci. Food Agric. 2014, 94, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Brkić, D.; Bošnir, J.; Bevardi, M.; Bošković, A.G.; Miloš, S.; Lasić, D.; Krivohlavek, A.; Racz, A.; Ćuić, A.M.; Trstenjak, N.U. Nitrate in leafy green vegetables and estimated intake. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 31–41. [Google Scholar] [CrossRef]
- Telesiński, A.; Grzeszczuk, M.; Jadczak, D.; Wysocka, G.; Onyszko, M. Assessment of changes in content of nitrates (V) in selected spice herbs depending on their preservation method and storage time. Food Sci. Technol. Qual. 2013, 20, 168–176. [Google Scholar] [CrossRef]
- Shen, W.; Zhang, W.; Li, J.; Huang, Z.; Tao, Y.; Hong, J.; Zhang, L.; Zhou, Y. Pre-harvest short-term continuous LED lighting improves the nutritional quality and flavor of hydroponic purple-leaf lettuce. Sci. Hortic. 2024, 334, 113304. [Google Scholar] [CrossRef]
- Viršilė, A.; Brazaitytė, A.; Jankauskienė, J.; Miliauskienė, J.; Vaštakaitė-Kairienė, V.; Odminytė, I.; Novickovas, A.; Samuoliene, G. Pre-harvest LED lighting strategies for reduced nitrate contents in leafy vegetables. Zemdirb.-Agric. 2018, 105, 249–256. [Google Scholar] [CrossRef]
- Ciriello, M.; Carillo, P.; Lentini, M.; Rouphael, Y. Influence of pre-harvest factors on the storage of fresh basil (Ocimum basilicum L.): A review. Horticulturae 2025, 11, 326. [Google Scholar] [CrossRef]
- Sale, A.I.; Uthairatanakij, A.; Laohakunjit, N.; Jitareerat, P.; Kaisangsri, N. Pre-harvest supplemental LED treatments led to improved postharvest quality of sweet basil leaves. J. Photochem. Photobiol. B Biol. 2023, 248, 112788. [Google Scholar] [CrossRef]
- Webkiosk Enza Zaden. Available online: https://webkiosk.enzazaden.com/warzywa-szklarniowe-i-tunelowe-2023/68445686/51 (accessed on 4 April 2025).
- Nowosielski, O. Methods for Determining Fertilization Needs of Horticultural Plants; PWRiL: Warsaw, Poland, 1988; p. 310. [Google Scholar]
- Zhou, L.-X. Determination of phosphorus in vanadium-phosphorus-iron ore by phosphorus vanadium molybdenum yellow spectrophotometry. Yejin Fenxi/Metall. Anal. 2018, 38, 75–80. [Google Scholar] [CrossRef]
- Kaban, J.; Reveny, J.; Tarigan, J.; Zebua, N. Modificated extraction and purity test of Arenga pinnata gum. Asian J. Pharm. Clin. Res. 2018, 11, 148–150. [Google Scholar] [CrossRef]
- Aćamović-Djoković, G.; Pavlović, R.; Mladenović, J. Vitamin C content of different types of lettuce. Acta Agric. Serb. 2011, 16, 83–89. [Google Scholar]
- Yang, Z.; Wang, T.; Wang, H.; Huang, X.; Qin, Y.; Hu, G. Patterns of enzyme activities and gene expressions in sucrose metabolism in relation to sugar accumulation and composition in the aril of Litchi chinensis Sonn. J. Plant Physiol. 2013, 170, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A review of environment effects on nitrate accumulation in leafy vegetables grown in controlled environments. Foods 2020, 9, 732. [Google Scholar] [CrossRef]
- Kim, D.; Son, J.E. Adding far-red to red, blue supplemental light-emitting diode interlighting improved sweet pepper yield but attenuated carotenoid content. Front. Plant Sci. 2022, 13, 938199. [Google Scholar] [CrossRef]
- Morrow, R.C. LED lighting in horticulture. HortScience 2008, 49, 1947–1950. [Google Scholar] [CrossRef]
- Nelson, J.A.; Bugbee, B. Analysis of environmental effects on leaf temperature under sunlight, High Pressure Sodium and Light Emitting Diodes. PLoS ONE 2015, 10, e138930. [Google Scholar] [CrossRef]
- Dörr, O.S.; Brezina, S.; Rauhut, D.; Mibus, H. Plant architecture and phytochemical composition of basil (Ocimum basilicum L.) under the influence of light from microwave plasma and high-pressure sodium lamps. J. Photochem. Photobiol. B Biol. 2020, 202, 111678. [Google Scholar] [CrossRef]
- Jeong, S.J.; Niu, G.; Zhen, S. Far-red light and temperature interactively regulate plant growth and morphology of lettuce and basil. Environ. Exp. Bot. 2024, 218, 105589. [Google Scholar] [CrossRef]
- Oehler, M.A.; Kelly, N.; Fonseca, J.M.; Evensen, E.; Park, E.; Gu, G.; Teng, Z.; Luo, Y. Influence of supplementary blue and far-red light on the morphology and texture of Ocimum basilicum L. grown in controlled environments. Horticulturae 2025, 11, 287. [Google Scholar] [CrossRef]
- Luetic, S.; Knezovic, Z.; Jurcic, K.; Perasovic, M.L.; Sutlovic, D. Nitrates and nitrites in leafy vegetables: The influence of culinary processing on concentration levels and possible impact on health. Int. J. Mol. Sci. 2025, 26, 3018. [Google Scholar] [CrossRef]
- Signore, A.; Bell, L.; Santamaria, P.; Wagstaff, C.; Van Labeke, M.C. Red light is effective in reducing nitrate concentration in rocket by increasing nitrate reductase activity, and contributes to increased total glucosinolates content. Front. Plant Sci. 2020, 11, 604. [Google Scholar] [CrossRef] [PubMed]
- Rouphael, Y.; Kyriacou, M.C.; Petropoulos, S.A.; De Pascale, S.; Colla, G. Improving vegetable quality in controlled environments. Sci. Hortic. 2018, 234, 275–289. [Google Scholar] [CrossRef]
- Ohashi-Kaneko, K.; Takase, M.; Kon, N.; Fujiwara, K.; Kurata, K. Effect of light quality on growth and vegetable quality in leaf lettuce, spinach and komatsuna. Environ. Control Biol. 2007, 45, 189–198. [Google Scholar] [CrossRef]
- Viršilė, A.; Brazaitytė, A.; Vaštakaitė-Kairienė, V.; Miliauskienė, J.; Jankauskienė, J.; Novičkovas, A.; Laužikė, K.; Samuolienė, G. The distinct impact of multi-color LED light on nitrate, amino acid, soluble sugar and organic acid contents in red and green leaf lettuce cultivated in controlled environment. Food Chem. 2020, 310, 125799. [Google Scholar] [CrossRef]
- Chen, X.-L.; Wang, L.-C.; Li, T.; Yang, Q.-C.; Guo, W.-Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 6926. [Google Scholar] [CrossRef] [PubMed]
- Min, Q.; Marcelis, L.F.; Nicole, C.C.; Woltering, E.J. High light intensity applied shortly before harvest improves lettuce nutritional quality and extends the shelf life. Front. Plant Sci. 2021, 12, 615355. [Google Scholar] [CrossRef]
- Gao, M.; He, R.; Shi, R.; Zhang, Y.; Song, S.; Su, W.; Liu, H. Differential effects of low light intensity on broccoli microgreens growth and phytochemicals. Agronomy 2021, 11, 537. [Google Scholar] [CrossRef]
- Ferrón-Carrillo, F.; Guil-Guerrero, J.L.; González-Fernández, M.J.; Lyashenko, S.; Battafarano, F.; da Cunha-Chiamolera, T.P.L.; Urrestarazu, M. LED enhances plant performance and both carotenoids and nitrates profiles in lettuce. Plant Foods Hum. Nutr. 2021, 76, 210–218. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Viršilė, A.; Jankauskienė, J.; Sakalauskienė, S.; Duchovskis, P. Red light-dose or wavelength-dependent photoresponse of antioxidants in herb microgreens. PLoS ONE 2016, 11, e0163405. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lighting Type | Lighting Period (Week After Sowing) | |||
|---|---|---|---|---|
| 3 | 4 | 5 | 6 | |
| HPS | ||||
| LED | ||||
| HPS + 1 week LED | ||||
| LOQ (µg × L−1) | LOD (µg × L−1) | Linear Range (µg × mL−1) | R (r2) | Calibration Equation | Precision Inter-Day (CV) | Precision Intra-Day (CV) | tR | Compound |
|---|---|---|---|---|---|---|---|---|
| 1.86 | 0.56 | 0.41–408.60 | 0.999 | y = 10553.4x + 324.5 | 0.87 | 0.49 | 0.76 | Lutein |
| 0.50 | 0.15 | 0.38–338.80 | 0.999 | y = 5245.2x + 654.3 | 0.89 | 0.50 | 1.29 | Chlorophyll b |
| 3.16 | 0.94 | 0.39–387.60 | 0.999 | y = 3143.4x + 432.2 | 0.78 | 0.46 | 1.82 | Chlorophyll a |
| 3.26 | 0.97 | 0.38–383.80 | 0.999 | y = 4324.2x − 324.5 | 0.67 | 0.32 | 3.77 | β-carotene |
| Lighting Type | ‘Keira’ | ‘Marian’ |
|---|---|---|
| HPS | 8.58 ± 0.36 b | 8.34 ± 0.59 b |
| LED | 9.33 ± 0.41 a | 8.84 ± 0.38 a |
| HPS + 1 week LED | 8.67 ± 0.38 ab | 8.74 ± 0.40 ab |
| ‘KEIRA’ | ||||||
|---|---|---|---|---|---|---|
| Lighting Type | K (mg × 100 g−1 FW) | P (mg × 100 g−1 FW) | Ca (mg × 100 g−1 FW) | TS (g × 100 g−1 FW) | TSS (°Brix) | AA (mg × 100 g−1 FW) |
| HPS | 453.8 ± 9.5 ns | 38.32 ± 2.9 ns | 39.58 ± 4.4 ns | 0.50 ± 0.10 b | 3.57 ± 0.18 b | 14.2 ± 2.4 b |
| LED | 454.7 ± 5.0 ns | 39.47 ± 0.5 ns | 39.19 ± 2.1 ns | 0.58 ± 0.09 a | 4.55 ± 0.05 a | 16.6 ± 1.1 a |
| HPS + LED | 451.3 ± 7.8 ns | 39.22 ± 1.2 ns | 40.84 ± 1.0 ns | 0.49 ± 0.07 b | 3.22 ± 0.08 b | 14.9 ± 3.2 b |
| ‘MARIAN’ | ||||||
|---|---|---|---|---|---|---|
| Lighting Type | K (mg × 100 g−1 FW) | P (mg × 100 g−1 FW) | Ca (mg × 100 g−1 FW) | TS (g × 100 g−1 FW) | TSS (°Brix) | AA (mg × 100 g−1 FW) |
| HPS | 406.9 ± 12.0 ns | 44.84 ± 2.2 ns | 63.85 ± 3.5 ns | 0.43 ± 0.02 b | 3.32 ± 0.10 b | 12.5 ± 0.50 b |
| LED | 394.2 ± 10.5 ns | 45.58 ± 0.6 ns | 63.80 ± 7.2 ns | 0.50 ± 0.05 a | 3.90 ± 0.00 a | 16.1 ± 0.25 a |
| HPS + LED | 411.4 ± 13.0 ns | 45.08 ± 2.9 ns | 63.94 ± 3.5 ns | 0.46 ± 0.00 b | 4.03 ± 0.15 a | 13.2 ± 3.33 b |
| ‘KEIRA’ | ||||
|---|---|---|---|---|
| Lighting Type | Chlorophyll a | Chlorophyll b | Lutein | β-Carotene |
| HPS | 107.51 ± 0.94 b | 29.92 ± 0.33 ns | 10.17 ± 0.09 ns | 16.51 ± 0.26 b |
| LED | 119.25 ± 2.87 a | 31.37 ± 0.60 ns | 10.69 ± 0.12 ns | 17.76 ± 0.38 a |
| HPS + LED | 107.60 ± 1.75 b | 29.25 ± 0.47 ns | 9.96 ± 0.35 ns | 16.44 ± 0.38 b |
| ‘MARIAN’ | ||||
|---|---|---|---|---|
| Lighting Type | Chlorophyll a | Chlorophyll b | Lutein | β-Carotene |
| HPS | 108.99 ± 3.53 b | 30.00 ± 1.01 ns | 10.07 ± 0.15 ns | 16.64 ± 0.19 b |
| LED | 112.55 ± 3.25 a | 30.36 ± 0.37 ns | 10.18 ± 0.06 ns | 17.90 ± 0.52 a |
| HPS + LED | 110.74 ± 3.70 b | 29.76 ± 1.05 ns | 10.19 ± 0.19 ns | 16.79 ± 0.12 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mirgos, M.; Geszprych, A.; Przybył, J.L.; Niedzińska, M.; Sujkowska-Rybkowska, M.; Gajc-Wolska, J.; Kowalczyk, K. Effect of Lighting Type on the Nitrates Concentration, Selective Bioactive Compounds and Yield of Sweet Basil (Ocimum basilicum L.) in Hydroponic Production. Agronomy 2025, 15, 966. https://doi.org/10.3390/agronomy15040966
Mirgos M, Geszprych A, Przybył JL, Niedzińska M, Sujkowska-Rybkowska M, Gajc-Wolska J, Kowalczyk K. Effect of Lighting Type on the Nitrates Concentration, Selective Bioactive Compounds and Yield of Sweet Basil (Ocimum basilicum L.) in Hydroponic Production. Agronomy. 2025; 15(4):966. https://doi.org/10.3390/agronomy15040966
Chicago/Turabian StyleMirgos, Małgorzata, Anna Geszprych, Jarosław L. Przybył, Monika Niedzińska, Marzena Sujkowska-Rybkowska, Janina Gajc-Wolska, and Katarzyna Kowalczyk. 2025. "Effect of Lighting Type on the Nitrates Concentration, Selective Bioactive Compounds and Yield of Sweet Basil (Ocimum basilicum L.) in Hydroponic Production" Agronomy 15, no. 4: 966. https://doi.org/10.3390/agronomy15040966
APA StyleMirgos, M., Geszprych, A., Przybył, J. L., Niedzińska, M., Sujkowska-Rybkowska, M., Gajc-Wolska, J., & Kowalczyk, K. (2025). Effect of Lighting Type on the Nitrates Concentration, Selective Bioactive Compounds and Yield of Sweet Basil (Ocimum basilicum L.) in Hydroponic Production. Agronomy, 15(4), 966. https://doi.org/10.3390/agronomy15040966