Transcription Factors Involved in Plant Stress and Growth and Development: NAC

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. NAC Transcription Factor
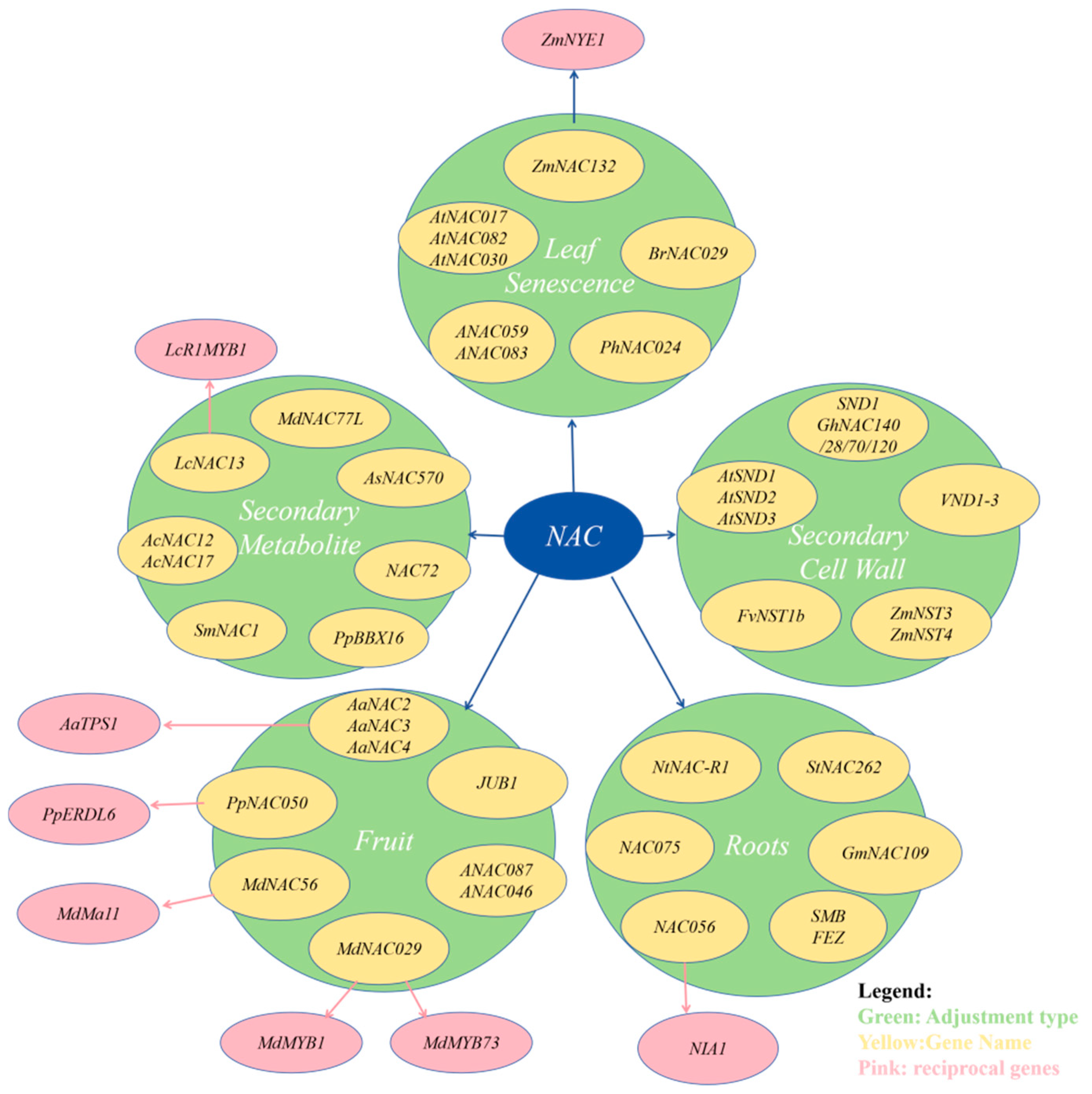
3. Studies on the Involvement of NAC Transcription Factors in Plant Growth and Development
3.1. Regulation of Plant Cell Secondary Wall Growth
3.2. Regulation of Root Development
3.3. Regulation of Leaf Senescence
3.4. Regulation of Fruit Growth
3.5. Summary
4. Studies on the Involvement of NAC Transcription Factors in Plant Stresses
4.1. Biological Stress
4.2. Abiotic Stresses
4.2.1. Cold Stress
4.2.2. Drought Stress
4.2.3. Salt Stress
4.2.4. Heavy Metal Stress
5. Summary and Outlook
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Souer, E.; van Houwelingen, A.; Kloos, D.; Jos, M.; Ronald, K. The no apical meristem gene of Petunia is required for pattern formation in embryosand flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef]
- Addie, N.; Heidi, E.; Leila, L.; Karen, S. NAC transcription factors:structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar]
- Ernst, H.A.; Olsen, A.N.; Larsen, S. Structure of theconserved domain of ANAC, a member of the NAC family oftranscription factors. EMBO Rep. 2004, 5, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.J.; Fan, H.; Chen, Y. Integrative omic and transgenic analyses reveal the positive effect of ultraviolet-B irradiation.on salvianolic acid biosynthesis through upregulation of SmNAC1. Plant J. 2020, 104, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Gao, Y.; Wan, W.; Shi, J.; Xv, C.; Ma, D.; Su, Y.; Yang, X. AsNAC570 Gene Cloning Analysis and Requlates Alin Biosynthesis Preliminary Exploration. Mol. Plant Breed 2022, 1–8. Available online: http://kns.cnki.net/kcms/detail/46.1068.S.20220715.1426.004.html (accessed on 10 October 2024).
- Jiang, T.; Zhang, Y.; Zuo, Z.; Luo, T.; Wang, H.; Zhang, R.; Luo, Z. Transcription factor PgNAC72 activates DAMMARENEDIOL SYNTHASE expression to promote ginseng saponin biosynthesis. Plant Physiol. 2024, kiae202. [Google Scholar] [CrossRef]
- Li, G.; Li, S.; Liu, W. Transcription factor NAC and its role in plant growth and development. Mol. Plant Breed. 2019, 17, 811–826. [Google Scholar] [CrossRef]
- Wang, F.; Sun, L.; Zhao, X.; Wang, J.; Song, X. Research progress of plant NAC transcription factors. Biotechnol. Bull. 2019, 35, 88–93. [Google Scholar] [CrossRef]
- Zhao, C.; Huang, X.-Q.; Yin, F.-Y. Progress of rice NAC transcription factor family. J. Plant Sci. 2020, 38, 278–287. [Google Scholar]
- Liang, J.; Liu, X.; Xu, L.; Mu, R.; Shen, N.; Li, S.; Cheng, C.; Ren, Y.; Ma, L.; Wang, B.; et al. A novel NAC transcription factor from Haloxylon ammodendron promotes reproductive growth in Arabidopsis thaliana under drought stress. Environ. Exp. Bot. 2024, 228 Pt A, 106043. [Google Scholar] [CrossRef]
- Park, S.R.; Jeong, Y.; Son, S. Functions of transcription factor superfamilies in rice immunity. Crop J. 2025, 13, 5–22. [Google Scholar] [CrossRef]
- Summat, P.; Tongmark, K.; Chakhonkaen, S.; Sangarwut, N.; Panyawut, N.; Pinsupa, S.; Thanananta, T.; Muangprom, A. Investigating cold tolerance mechanisms in rice seedlings: Alternative splicing, promoter analysis, and their applications for marker development. Plant Stress 2024, 13, 100530. [Google Scholar] [CrossRef]
- Chen, J.; Gong, Y.; Gao, Y.; Zhou, Y.; Chen, M.; Xu, Z.; Guo, C.; Ma, Y. TaNAC48 positively regulates drought tolerance and ABA responses in wheat (Triticum aestivum L.). Crop J. 2021, 9, 9. [Google Scholar] [CrossRef]
- Jia, D.; Jiang, Q.; van Nocker, S.; Ma, F. An apple (Malus domestica) NAC transcription factor enhances drought tolerance in transgenic apple plants. Plant Physiol. Biochem. 2019, 139, 504–512. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Gong, C. Progress in the study of biological functions of plant NAC transcription factors. Henan Sci. 2024, 1–13. [Google Scholar]
- Zhuo, M.; Wang, H. Regulatory role of NAC transcription factors in fruit ripening. J. Fruit Tree 2023, 40, 1455–1470. [Google Scholar] [CrossRef]
- Yan, S.; Wang, Y.; Yu, B.; Gan, Y.; Lei, J.; Chen, C.; Zhu, Z.; Qiu, Z.; Cao, B. A putative E3 ubiquitin ligase substrate receptor degrades transcription factor SmNAC to enhance bacterial wilt resistance in Eggplan. Hortic. Res. 2024, 11, uhad246. [Google Scholar] [CrossRef]
- Dastmalchi, K.; Chira, O.; Rodriguez, M.P.; Yoo, B.; Serra, O.; Figueras, M.; Stark, R. A chemical window into the impact of RNAi silencing of the StNAC103 gene in potato tuber periderms: Soluble metabolites, suberized cell walls, and antibacterial defense. Phytochemistry 2021, 190, 112885. [Google Scholar] [CrossRef]
- Ma, X.; Yin, Y.; Feng, J.X.; Chen, W.; Sun, L.; Xiao, Y. Progress in the study of plant NAC transcription factors. Plant Physiol. Lett. 2021, 57, 2225–2234. [Google Scholar] [CrossRef]
- Cui, X.; Cao, Y.; Zhang, H.; Zhang, L. A Picea wilsonii NAC transcription factor, PwNAC1, interacts with RNA-binding protein PwRBP1 and synergistically enhances drought and salt tolerance of transgenic Arabidopsis. Environ. Exp. Bot. 2023, 206, 105174. [Google Scholar] [CrossRef]
- Huang, Y.; Cui, L.; Chen, W.; Liu, Z.; Yuan, W.; Zhu, F.; Jiao, Z.; Zhang, X.; Deng, X.; Wang, L. Comprehensive analysis of NAC transcription factors and their expressions during taproot coloration in radish (Raphanus sativus L.). Sci. Hortic. 2022, 299, 111047. [Google Scholar] [CrossRef]
- Negi, S.; Tak, H.; Ganapathi, T. Native vascular related NAC transcription factors are efficient regulator of multiple classes of secondary wall associated genes in banana. Plant Sci. 2017, 265, 70–86. [Google Scholar] [CrossRef]
- Yang, X.; Kim, M.Y.; Ha, J.; Lee, S.H. Overexpression of the soybean NAC gene GmNAC109 increases lateral root formation and abiotic stress tolerance in transgenic Arabidopsis plants. Front. Plant Sci. 2019, 10, 1036. [Google Scholar] [CrossRef]
- Guo, F.; Siyuan, L.; Zhang, C.; Dong, T.; Meng, X.; Zhu, M. Genome-wide systematic survey and analysis of NAC transcription factor family and their response to abiotic stress in sweetpotato. Sci. Hortic. 2022, 299, 111048. [Google Scholar] [CrossRef]
- Qu, D.; Wu, F.; Yang, J.; Li, M.; Yang, L.; Xie, R.; Zhou, J.; Yang, J.; Wang, L.; Su, H.; et al. Transcription factor PtNAC101 negatively regulates the lignin synthesis and salt tolerance in Populus trichocarpa. Environ. Exp. Bot. 2023, 205, 105149. [Google Scholar] [CrossRef]
- Wang, H.; Qin, L.; Feng, C.; Wu, M.; Zhong, H.; Liu, J.; Wu, Q.; Que, Y. Pathogen resistance was negatively regulated by the NAC transcription factor ScATAF1 in sugarcane. Plant Physiol. Biochem. 2024, 213, 108828. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Chung, P.; Jeong, J.; Jang, G.; Bang, S.; Jung, H.; Kim, Y.; Ha, S.; Choi, Y.; Kim, J. The rice OsNAC6 transcription factor orchestrates multiple molecular mechanisms involving root structural adaptions and nicotianamine biosynthesis for drought tolerance. Plant Biotechnol. J. 2017, 15, 754–764. [Google Scholar] [CrossRef]
- Richa, S.; Lingaraj, S. Genome-wide analysis of cowpea NAC transcription factor family elucidating the genetic & molecular relationships that interface stress and growth regulatory signals. Plant Gene 2022, 31, 100363. [Google Scholar]
- Yuan, X.; Xu, J.; Yu, J.; Zhu, D.; Li, H.; Zhao, Q. The NAC transcription factor ZmNAC132 regulates leaf senescence and male fertility in maize. Plant Sci. 2023, 334, 111774. [Google Scholar] [CrossRef]
- Kang, C.; Guo, C.; Zhang, X.; Liu, Z. Genome-wide identification and analysis of the NAC gene family in walnut. J. Fruit Tree 2021, 38, 1444–1458. [Google Scholar]
- Nigarish, M.; Chen, Y.K.; Chen, X.H.; Muhammad, A.; Junaid, I.; Hafiz, M.; Shen, X.; Lin, Y.; Xu, X.; Lai, Z. Genome- wide identification and comprehensive analyses of NAC, transcription factor gene family and expression patterns during, somatic embryogenesis in Dimocarpus longan Lour. Plant Physiol. Biochem. 2020, 157, 169–184. [Google Scholar]
- Mohanta, T.K.; Yadav, D.; Khan, A. Genomics, molecular and evolutionary perspective of NAC transcription factors. PLoS ONE 2020, 15, e0231425. [Google Scholar] [CrossRef]
- Singh, S.; Koyama, H.; Bhati, K.K.; Alok, A. The biotechnological importance of the plant-specific NAC transcription factor family in crop improvement. J. Plant Res. 2021, 134, 475–495. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.G.; Jiang, S.H.; Zhang, T.L.; Xu, H.F.; Fang, H.C.; Zhang, J.; Su, M.Y.; Wang, Y.C.; Zhang, Z.Y.; Wang, N.; et al. Apple NAC transcription factor MdNAC52 regulates biosynthesis of anthocyanin and proanthocyanidin through MdMYB9 and MdMYB11. Plant Sci. 2019, 289, 110286. [Google Scholar] [CrossRef] [PubMed]
- Duan, Q.; Lee, J.; Liu, Y.; Chen, H.; Hu, H. Distribution of heavy metal pollution in surface soil samples in China: A graphical review. Bull. Environ. Contam. Toxicol. 2016, 97, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Wang, L.; Zhu, B.; Li, C. NAC transcription factors: Regulatory factors involved in many life processes in plants. Anhui Agric. Sci. 2024, 52, 22–29. [Google Scholar]
- Muskan, B.; Baljinder, S.; Avneesh, K.; Nandni, G.; Kashmir, S.; Ravindresh, C. Genome-wide identification of NAC transcription factors in Avena sativa under salinity stress. Plant Stress 2023, 10, 100276. [Google Scholar]
- Wang, J.; Zheng, C.; Shao, X.; Hu, Z.; Li, J.; Wang, P.; Wang, A.; Yu, J.; Shi, K. Transcriptomic and genetic approaches reveal an essential role of the NAC transcription factor SlNAP1 in the growth and defense response of tomato. Hortic. Res. 2020, 7, 209. [Google Scholar] [CrossRef]
- Zhao, X.; Xue, Z.; Liu, Y.; Huang, Z.; Sun, Y.; Wu, C.; Yan, S.; Kou, X. ChIP-seq revealed the role of tomato SNAC4 in response to biological and abiotic stresses and mediating auxin signaling response. Postharvest Biol. Technol. 2024, 209, 112696. [Google Scholar] [CrossRef]
- Xiang, C.; Gao, Q.; Yuan, C.; Zhang, X.; Liang, Y.; Zhao, Y. Identification and bioinformatics analysis of NAC transcription factor family in Calendula officinalis. Mol. Plant Breed. 2023, 12, 1518. [Google Scholar]
- Wang, Z.; Zhang, Z.; Wang, P.; Qin, C.; He, L.; Kong, L.; Ren, W.; Liu, X.; Ma, W. Genome-wide identification of the NAC transcription factors family and regulation of metabolites under salt stress in Isatis indigotica. Int. J. Biol. Macromol. 2023, 240, 124436. [Google Scholar] [CrossRef]
- Wang, Y.; Du, H.; Deng, P. Identification of NAC gene family in mulberry and analysis of expression pattern under rooting powder treatment. J. Fujian Agric. For. Univ. Nat. Sci. Ed. 2024, 53, 629–640. [Google Scholar] [CrossRef]
- Wan, F.; Gao, J.; Wang, G.; Niu, Y.; Wang, L.; Zhang, X.; Wang, Y.; Pan, Y. Genome-wide identification of NAC transcription factor family and expression analysis of ATAF subfamily members under abiotic stress in eggplant. Sci. Hortic. 2021, 289, 110424. [Google Scholar] [CrossRef]
- Liu, F.; Yang, C.; He, D.; Xing, B.; Li, M. Identification of pea NAC gene family and analysis of its response under drought stress. J. Agric. Biotechnol. 2024, 32, 1504–1517. [Google Scholar]
- Li, S.; Wang, C.; Li, X. Bioinformaticanalysis of the NAC transcription factor family in Punica granatum L. Mol. Plant Breed. 2021, 19, 88–99. [Google Scholar]
- Ribal, M.; Erzsébet, K. An integrative analysis of Vitis vinifera L. NAC Genes Response Botrytis Cinerea. Physiol. Mol. Plant Pathol. 2024, 131, 102247. [Google Scholar]
- Liu, H.; Jiang, B.; Zhang, J. Research progress of plant NAC transcription factors. J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2024, 1–12. [Google Scholar]
- Lin, S.; Longhui, L.; Jiang, Y.; Jing, Y. Advances in membrane-tethered NAC transcription factors in plants. Plant Sci. 2024, 342, 112034. [Google Scholar]
- Deng, Z.; Luo, L.; Yu, C.; Zhang, Q.; Sui, Y. Progress in the study of NAC transcription factors in ornamental plants. J. Plant Genet. Resour. 2024, 25, 737–750. [Google Scholar] [CrossRef]
- Zhong, R.; Lee, C.; Ye, Z.H. Functional characterization of poplar wood-associated.NAC domain transcription factors. Plant Physiol. 2010, 152, 104–1055. [Google Scholar] [CrossRef]
- Cosgrove, D.J. Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 2005, 6, 850–861. [Google Scholar] [CrossRef]
- Long, G.; Wu, P.; Fu, J.; Lu, H.; Zhang, R. Progress of peroxidase-regulated lignin synthesis. Mod. Agric. Sci. Technol. 2021, 805, 47–49+54. [Google Scholar]
- Dang, X.; Zhang, B.; Li, C.; Nagawa, S. FvNST1b NAC protein induces secondary cell wall formation in strawberry. Int. J. Mol. Sci. 2022, 23, 13212. [Google Scholar] [CrossRef]
- Fang, S.; Shang, X.; Yao, Y. NST- and SND-sub group NAC proteins coordinately act to regulate second ary cell wall formation in cotton. Plant Sci. 2020, 301, 110657. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Yang, Y.; Yu, J. ZmNST3 and ZmNST4 are master switches for secondary wall deposition in maize (Zea mays L.). Plant Sci. 2018, 266, 83–94. [Google Scholar] [CrossRef]
- Yu, Y.; Ran, G.; Pi, L. Plant nitrogen signal perception and root development. J. Plant Sci. 2024, 42, 825–832. [Google Scholar]
- Meng, J.; Liu, H.; Wu, Z.; Zuo, W.; Wang, L. Effect of PsWOX11 gene on growth and development of lateral roots in Populus tremula. For. Sci. Res. 2024, 37, 54–61. [Google Scholar]
- Fu, Y.; Guo, H.; Cheng, Z.; Wang, R.; Li, G.; Huo, G.; Liu, W. NtNAC-R1, a novel NAC transcription factor gene in tobacco roots, responds to mechanical damage of shoot meristem. Plant Physiol. Biochem. 2013, 69, 74–81. [Google Scholar] [CrossRef]
- Zhang, L.; Yao, L.; Zhang, N.; Yang, J.; Zhu, X.; Tang, X.; Calderon-Urrea, A.; Si, H. Lateral root development in potato is mediated by Stu-mi164 regulation of NAC transcription factor. Front. Plant Sci. 2018, 9, 383. [Google Scholar] [CrossRef]
- Viola, W.; Marion, B.; Tom, B.; Ana, C.; Harald, W.; Jian, X.; Jim, H.; Ben, S. The NAC Domain Transcription Factors FEZ and SOMBRERO Control the Orientation of Cell Division Plane in Arabidopsis Root Stem Cells. Dev. Cell 2008, 15, 913–922. [Google Scholar] [CrossRef]
- Xu, P.; Ma, W.; Hu, J.; Cai, W. The nitrate-inducible NAC transcription factor NAC056 controls nitrate assimilation and promotes lateral root growth in Arabidopsis thaliana. PLoS Genet. 2022, 18, e1010090. [Google Scholar] [CrossRef]
- Chenbo, D. Functional Analysis of MdNAC56 in the regulation of ACIDITY in Apple Fruit; Northwest Agriculture and Forestry University: Xianyang, China, 2024. [Google Scholar] [CrossRef]
- Xiao, H.; Hu, Y.; Wang, Y.; Cheng, J.; Wang, J.; Chen, G.; Li, Q.; Wang, S.; Wang, Y.; Wang, S. Nitrate availability controls translocation of the transcription factor NAC075 for cell-type-specific reprogramming of root growth. Dev. Cell 2022, 57, 2638–2651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.M.; Guo, P.R.; Xia, X.L.; Guo, H.W.; Li, Z.H. Multiple layers of regulation on leaf senescence: New advances and perspectives. Front. Plant Sci. 2021, 12, 788996. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Guo, Y. Research progress of leaf senescence related genes in tobacco. J. Plant Sci. 2018, 6, 87. [Google Scholar] [CrossRef]
- Wang, H.L.; Zhang, Y.; Wang, T.; Yang, Q.; Yang, Y.L.; Li, Z.; Li, B.S.; Wen, X.; Li, W.Y.; Yin, W.L.; et al. An alternative splicing variant of PtRD26 delays leaf senescence by regulating multiple NAC transcription factors in Populus. Plant Cell 2021, 33, 1594–1614. [Google Scholar] [CrossRef]
- Li, F.; Shan, Y.; Wang, H.; Jiang, G.; Ding, X.; Liang, H.; Wang, C.; Kong, X.; Xie, L.; Jiang, Y. A NAC transcriptional factor BrNAC029 is involved in cytokinin-delayed leaf senescence in postharvest Chinese flowering cabbage. Food Chem. 2023, 404 Pt B, 134657. [Google Scholar] [CrossRef] [PubMed]
- Balazadeh, S.; Kwasniewski, M.; Caldana, C.; Mehrnia, M.; Zanor, M.I.; Xue, G.P.; Mueller-Roeber, B. ORS1, an H(2)O(2)-responsive NAC transcription factor, controls senescence in Arabidopsis thaliana. Mol. Plant 2011, 4, 346–360. [Google Scholar] [CrossRef]
- Yang, S.D.; Seo, P.J.; Yoon, H.K.; Park, C.M. The Arabidopsis NAC transcription factor VNI2 integrates abscisic acid signals into leaf senescence via the COR/RD genes. Plant Cell 2011, 23, 2155–2168. [Google Scholar] [CrossRef]
- Trupkin, S.A.; Astigueta, F.H.; Baigorria, A.H.; García, M.N.; Delfosse, V.C.; González, S.A.; de la Torre, M.C.P.; Moschen, S.; Lía, V.V.; Fernández, P.; et al. Identification and expression analysis of NAC transcription factors potentially involved in leaf and petal senescence in Petunia hybrida. Plant Sci. 2019, 287, 110195. [Google Scholar] [CrossRef]
- Chang, W.J. Analysis of the Mechanism of MdNAC029 Regulating the Acidity of Apple Fruit; Northwest Agriculture and Forestry University: Xianyang, China, 2024. [Google Scholar] [CrossRef]
- Liu, J.; Jing, Y.; Liu, Y.; Xu, Y.; Yu, Y.; Ge, X.; Xie, H. Identification of peach NAC gene family and promotion of fruit fructose accumulation by PpNAC050. J. Hortic. 2024, 51, 1983–1996. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Chen, X.Y.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural variation in monoterpene synthesis in kiwifruit: Transcriptional regulation of terpene synthases by NAC and ETHYLENE-INSENSITIVE3-like transcription factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef]
- Wu, A.H.; Allu, A.D.; Garapati, P.; Siddiqui, H.; Dortay, H.; Zanor, M.; Asensi-Fabado, M.; Munné-Bosch, S.; Antonio, C.; Tohge, T.; et al. JUNGBRUNNEN1, a reactive oxygen species-responsive NAC transcription factor, regulates longevity in Arabidopsis. Plant Cell 2012, 2, 482–506. [Google Scholar] [CrossRef]
- Huysmans, M.; Buono, R.A.; Skorzinski, N.; Radio, M.; De, W.; Parizot, B.; Mertens, J.; Karimi, M.; Fendrych, M.; Nowack, M. NAC transcription factors ANAC087 and ANAC046 control distinct aspects of programmed cell death in the Arabidopsis columella and lateral root cap. Plant Cell 2018, 30, 2197–2213. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.; Yang, H.; Hu, R.; Wei, D.; Tang, Q.; Wang, Z. Role of NAC transcription factors in plant flower development. J. Biol. Eng. 2022, 38, 2687–2699. [Google Scholar]
- Cao, Y.; Hwarari, D.; Radani, Y.; Guan, Y.; Yang, L. Molecular Mechanism Underlying Plant Response to Cold Stress. Phyton-Int. J. Exp. Bot. 2023, 92, 2665–2682. [Google Scholar] [CrossRef]
- Li, S.; Ma, H.; Li, C. Progress in the study of bHLH transcription factors in response to low temperature stress in plants. Jiangsu Agric. Sci. 2024, 52, 11–19. [Google Scholar] [CrossRef]
- An, J.; Li, R.; Qu, F.; You, C.; Wang, X.; Hao, Y. An apple NAC transcription factor negatively regulates cold tolerance via CBF-dependent pathway. J. Plant Physiol. 2018, 221, 74–80. [Google Scholar] [CrossRef]
- Shan, W. Mechanistic Analysis of NAC-Like Transcription Factors Regulating Banana fruit INDUCED Cold Tolerance and Ripening; South China Agricultural University: Guangzhou, China, 2016. [Google Scholar]
- Song, C. In-depth genome-wide characterization of MaNAC25 and MaNAC28 cold-responsive transcription factor binding sites in banana via DAP-Seq. Curr. Plant Biol. 2024, 40, 100389. [Google Scholar] [CrossRef]
- Hou, X.; Zhang, H.; Liu, S.; Wang, X.; Zhang, Y.; Meng, Y.; Luo, D.; Chen, R. The NAC transcription factor CaNAC064 is a regulator of cold stress tolerance in peppers. Plant Sci. 2020, 291, 110346. [Google Scholar] [CrossRef]
- Liang, Z.; Shi, Y.; Huang, Y.; Lu, J.; Zhang, M.; Cao, X.; Hu, R.; Li, D.; Chen, W.; Zhu, C.; et al. YLEM NAC DOMAIN 1 (EjXND1) relieves cold-induced lignification by negatively regulating the EjHB1-EjPRX12 module in loquat fruit. J. Adv. Res. 2024, in press. [Google Scholar] [CrossRef]
- Ge, H. Transcription Factors NAC and MADS are Involved in the Regulation of Postharvest Loquat Fruit Lignification by Cold Damage; Zhejiang University: Hangzhou, China, 2018. [Google Scholar]
- Fábio, T.; Paulo, S.; Sandra, R.; Jorge, H.; Vicente, E.; Patrícia, P.; Paulo, A. SsNAC23, a member of the NAC domain protein family, is associated with cold, herbivory and water stress in sugarcane. Plant Sci. 2005, 169, 93–106. [Google Scholar]
- Zhaoqiang, Z.; Ping, W.; Linqi, L.; Qiang, F.; Yibo, D.; Peng, C.; Ping, X.; Tian, W.; Haiyun, S. Recent development on drought propagation: A comprehensive review. J. Hydrol. 2024, 645 Pt B, 132196. [Google Scholar]
- Du, L.; Huang, X.; Ding, L.; Wang, Z.; Tang, D.; Chen, B.; Ao, L.; Liu, Y.; Kang, Z.; Mao, H. TaERF87 and TaAKS1 synergistically regulate TaP5CS1/TaP5CR1-mediated proline biosynthesis to enhance drought tolerance in wheat. New Phytol. 2023, 237, 232–250. [Google Scholar] [CrossRef]
- Wang, J.; Li, C.; Hu, G.; Zhang, Y.; Reynolds, M.; Zhang, X.; Jia, J.; Mao, X.; Jing, R. DIW1 encoding a clade I PP2C phosphatase negatively regulates drought tolerance by de-phosphorylating TaSnRK1.1 in wheat. J. Integr. Plant Biol. 2023, 65, 1918–1936. [Google Scholar] [CrossRef]
- Mao, H.; Li, S.; Chen, B.; Jian, C.; Mei, F.; Zhang, Y.; Li, F.; Chen, N.; Li, T.; Du, L.; et al. Variation in cis-regulation of a NAC transcription factor contributes to drought tolerance in wheat. Mol. Plant 2022, 15, 276–292. [Google Scholar] [CrossRef]
- Gong, F.; Zhang, F.; Lu, Y.; Velu, G.; Liu, R.; Liu, J.; Wang, X.; Liu, D.; Zheng, Y.; Huang, L.; et al. Overexpression of TdNACB improves the drought resistance of rice. Plant Physiol. Biochem. 2024, 216, 109157. [Google Scholar] [CrossRef]
- Xia, L.; Sun, S.; Han, B.; Yang, X. NAC domain transcription factor gene GhNAC3 confers drought tolerance in plants. Plant Physiol. Biochem. 2023, 195, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Wu, Y.; Mao, Z.; Yin, K.; He, Y.; Pan, X.; Zhu, X.; Liao, C.; Cui, L.; Jia, Q.; et al. A novel NAC transcription factor ZmNAC55 negatively regulates drought stress in Zea mays. Plant Physiol. Biochem. 2024, 214, 108938. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Zheng, Y.; Yu, T.; Cao, H.; Chen, Y.; Cui, Q.; Xu, C.; Li, Z. SlNAC6, A NAC transcription factor, is involved in drought stress response and reproductive process in tomato. J. Plant Physiol. 2021, 264, 153483. [Google Scholar] [CrossRef]
- Zhao, J.; Wu, Q.; Wu, H.; Wang, A.; Wang, X.; Li, C.; Zhao, H.; Wu, Q. FtNAC31, a Tartary buckwheat NAC transcription factor, enhances salt and drought tolerance in transgenic Arabidopsis. Plant Physiol. Biochem. 2022, 191, 20–33. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, S.; Guan, C.; Kong, X.; Wang, Y.; Cui, Y.; Liu, B.; Zho, Y.; Zhang, Y. Overexpressing the NAC transcription factor LpNAC13 from Lilium pumilum in tobacco negatively regulates the drought response and positively regulates the salt response. Plant Physiol. Biochem. 2020, 149, 96–110. [Google Scholar] [CrossRef]
- Rakesh, S.; Bhavesh, L.; Abdul, A.; Bal, K.; Prafullachandra, V.; Aniruddha, P. Differential regulation of the banana stress NAC family by individual and combined stresses of drought and heat in susceptible and resistant genotypes. Plant Physiol. Biochem. 2019, 145, 184–194. [Google Scholar]
- Liu, R.; Meng, J.; Zuo, W.; Zuo, W.; Wang, L.; Sun, T. The phloem protein 2 (PP2) is positively regulated by ThNAC13 that enhances salt tolerance of Tamarix. Environ. Exp. Bot. 2024, 224, 105784. [Google Scholar] [CrossRef]
- Yang, H.; Yi, Z.; Shanwu, L.; Liu, Y.; Jian, S.; Deng, S. MpNAC1, a transcription factor from the mangrove associate Millettia pinnata, confers salt and drought stress tolerance in transgenic Arabidopsis and rice. Plant Physiol. Biochem. 2024, 211, 108721. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Han, T.; Xiang, Y.; Wang, C.; Zhang, A. The transcription factor ZmNAC84 increases maize salt tolerance by regulating ZmCAT1 expression. Crop J. 2024, 12, 1344–1356. [Google Scholar] [CrossRef]
- Gong, C.; Zheng, Y.; Pan, V. Salt tolerance analysis of Capsicum annuum CaNAC36. Northwest J. Botany 2024, 44, 593–601. [Google Scholar]
- Han, D.; Du, M.; Zhou, Z.; Wang, S.; Li, T.; Han, J.; Xu, T.; Yang, G. An NAC transcription factor gene from Malus baccata, MbNAC29, increases cold and high salinity tolerance in Arabidopsis. Vitr. Cell. Dev. Biol. Plant 2020, 56, 588–599. [Google Scholar] [CrossRef]
- Li, P.; Peng, Z.; Xu, P.; Tang, G.; Ma, C.; Zhu, J.; Shan, L.; Wan, S. Genome-wide identification of NAC transcription factors and their functional prediction of abiotic stress response in peanut. Front. Genet. 2021, 12, 630292. [Google Scholar] [CrossRef]
- Harshita, J.; Klaus, H.; Leander, R.; Shashank, K.; Puneet, S. Elucidation of PGPR-responsive OsNAM2 regulates salt tolerance in Arabidopsis by AFP2 and SUS protein interaction. Microbiol. Res. 2024, 289, 127890. [Google Scholar] [CrossRef]
- Trishla, V.S.; Kirti, P.B. KirtiStructure-function relationship of Gossypium hirsutum NAC transcription factor, GhNAC4 with regard to ABA and abiotic stress responses. Plant Sci. 2021, 302, 110718. [Google Scholar] [CrossRef]
- Abu, S.; Li, X.; Li, H.; Huang, T.; Gao, C.; Guo, M.; Cheng, W.; Zhao, G.; Liao, Y. A rice stress-responsive NAC gene enhances tolerance of transgenic wheat to drought and salt stresses. Plant Sci. 2013, 203–204, 33–40. [Google Scholar]
- Wang, D.; Lliu, Z.; Lu, X.; Gao, Y.; Sun, S.; Guo, H.; Tian, W.; Wang, L.; Li, Z.; Li, L.; et al. Progress and prospect of salt tolerance mechanism in plants. North China J. Agric. 2024, 39, 80–92. [Google Scholar]
- Hu, S.; Shinwari, K.I.; Song, Y.; Xia, J.; Xu, H.; Du, B.; Luo, L.; Zheng, L. OsNAC300 positively regulates cadmium stress responses and tolerance in rice roots. Agronomy 2021, 11, 95. [Google Scholar] [CrossRef]
- Zhu, B.; Huo, D.A.; Hong, X.X.; Guo, J.; Peng, T.; Liu, J.; Huang, X.; Yan, H.; Weng, Q.; Zhang, X.; et al. The Salvia miltiorrhiza NAC transcription factor SmNAC1 enhances zinc content in transgenic Arabidopsis. Gene 2019, 688, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; He, F.; Zhu, B.; Ren, M.; Tang, H. NAC transcription factors from Aegilops markgrafii reduce cadmium concentration in transgenic wheat. Plant Soil 2020, 449, 39–50. [Google Scholar] [CrossRef]
- Meng, Y.; Zhang, X.; Wu, Q.; Fang, S.; Fang, Z. Transcription factor ANAC004 enhances Cd tolerance in Arabidopsis thaliana by regulating cell wall fixation, translocation and vacuolar detoxification of Cd, ABA accumulation and antioxidant capacity. J. Hazard. Mater. 2022, 436, 129121. [Google Scholar] [CrossRef]
- Xin, Y.; Huang, R.; Xu, M.; Xu, L. Transcriptome-Wide Identification and Response Pattern Analysis of the Salix integra NAC Transcription Factor in Response to Pb Stress. Int. J. Mol. Sci. 2023, 24, 11334. [Google Scholar] [CrossRef]
- Tao, Y.; Wan, J.X.; Liu, J.X.; Zheng, Y.; Fang, S.; Fang, Z. The NAC transcription factor ANAC017 regulates aluminum tolerance by regulating the cell wall-modifying genes. Plant Physiol. 2022, 189, 2517–2534. [Google Scholar] [CrossRef]
- Zheng, Y.; Lu, Y.; Feng, H.; Feng, J.; Chen, C.; Xu, C.; Li, H. Progress on the response mechanism of mustard to salt stress. Vegetables 2024, 17–23. [Google Scholar]
- Ou, X.; Sun, L.; Chen, Y.; Zhao, Z.; Jian, W. Characteristics of NAC transcription factors in Solanaceae crops and their roles in responding to abiotic and biotic stresses. Biochem. Biophys. Res. Commun. 2024, 709, 149840. [Google Scholar] [CrossRef]
- Wei, Y.; Tian, H. Progress of research on environmental heavy metal pollution. Mod. Agric. Sci. Technol. 2024, 104–106+120. [Google Scholar]
- Wang, Y.F. Research progress on the effects of heavy metal stress on alfalfa. Contemp. Anim. Husb. 2024, 45–47. [Google Scholar]
- Xu, S. Hazards of soil heavy metal pollution on agricultural cultivation and management strategy. Heilongjiang Grain 2024, 119–121. [Google Scholar]
- Angon, P.B.; Islam, M.S.; Das, A.; Anjum, N.; Poudel, A.; Suchi, S.A. Sources, effects and present perspectives of heavy metals contamination: Soil, plants and human food chain. Heliyon 2024, 10, e28357. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Tao, C.; Cao, L.; Liu, L.; Zheng, J.; Zhu, W.; Xiang, M.; Wang, L.; Liu, Y.; Zhang, C.; et al. AcNRZ1, an endoplasmic reticulum-localized NAC transcription factor in kiwifruit, enhances zinc stress tolerance in Arabidopsis. Plant Stress 2024, 13, 100559. [Google Scholar] [CrossRef]
- Wang, B.; Zhong, Z.; Zhang, H.; Wang, X.; Liu, B.; Yang, L.; Han, X.; Yu, D.; Zheng, X.; Wang, C. Targeted Mutagenesis of NAC Transcription Factor Gene, OsNAC041, Leading to Salt Sensitivity in Rice. Rice Sci. 2019, 26, 98–108. [Google Scholar]
- Sun, L.; Zhang, P.; Li, W. The bifunctional transcription factor NAC32 modulates nickel toxicity responses through repression of root-nickel compartmentalization and activation of auxin biosynthesis. J. Hazard. Mater. 2024, 480, 135925. [Google Scholar] [CrossRef]
- Takasaki, H.; Maruyama, K.; Kidokoro, S.; Ito, Y.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K.; Nakashima, K. The abiotic stress-responsive NAC-type transcription factor OsNAC5 regulates stress-inducible genes and stress tolerance in rice. Mol. Genet. Genom. 2010, 284, 173–183. [Google Scholar] [CrossRef]
- Nakashima, K.; Takasaki, H.; Mizoi, J. NAC transcription factors in plant abiotic stress responses. Biochim. Biophys. Acta (BBA) Gene Regul. Mech. 2012, 1819, 97–103. [Google Scholar] [CrossRef]
- Zhang, X.; Long, Y.; Chen, X.; Zhang, B.; Xin, Y.; Li, L.; Cao, S.; Liu, F.; Wang, Z.; Huang, H. A NAC Transcription Factor OsNAC3 Positively Regulates ABA Response and Salt Tolerance in Rice. BMC Plant Biol. 2021, 21, 546. [Google Scholar] [CrossRef]
- Chen, Y.; Xia, P. NAC transcription factors as biological macromolecules responded to abiotic stress: A comprehensive review. Int. J. Biol. Macromol. 2025, 308 Pt 1, 142400. [Google Scholar] [CrossRef]
- Kurowska, M.; Daszkowska-Golec, A. Molecular mechanisms of SNAC1 (Stress-responsive NAC1) in conferring the abiotic stress tolerance. Plant Sci. 2023, 337, 111894. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Name of the Species | Total No. of NACTFs | Identified NAC Transcription Factors | Function | References |
|---|---|---|---|---|
| Brassica napus | 410 | [15] | ||
| Malus domestica | 253 | MdNAC029/MdNAC1/MdNAC56 | Change fruit/drought tolerance | [16,17,18] |
| Nicotiana benthamiana | 227 | [19] | ||
| Brassica rapa | 204 | [20] | ||
| Musa paradisiaca | 181 | VND1-3 | Regulate lignin | [21,22] |
| Glycine max | 180 | GmNAC109 | lateral root development | [20,23] |
| Ipomoea batatas | 175 | [24] | ||
| Populus trichocarpa | 169 | PtNAC101 | Salt tolerance | [15,25] |
| Nicotiana tabacum | 154 | ScATAF1 | Disease resistance | [19,26] |
| Oryza sativa | 151 | OsNAC6/OsNAC300 | Drought tolerance, salt tolerance/cadmium tolerance | [21,27] |
| Vigna unguiculata | 130 | [28] | ||
| Zea mays | 130 | ZmNAC132/ZmNAC55 | Regulation of senescence/drought resistance | [20,29] |
| Fragaria vesca | 127 | [16] | ||
| Phyllostachys edulis | 125 | [19] | ||
| Juglans regia | 121 | [30] | ||
| Osmanthus fragrans | 119 | [19] | ||
| Dimocarpus longan | 114 | [31] | ||
| Arabidopsis thaliana | 113 | AtSND1/AtNAC056/AtNAC090 | Growth and development | [32,33,34] |
| Apium graveolens | 111 | [35] | ||
| Camellia sinensis | 104 | [36] | ||
| Avena sativa L. | 101 | [37] | ||
| Lycopersicon esculentum | 101 | SlNAP1/SNAC4 | Disease resistance | [15,38,39] |
| Erigeron breviscapus | 98 | [40] | ||
| Isatidis Radix | 96 | [41] | ||
| Morus alba | 92 | [42] | ||
| Solanum melongena L. | 90 | [43] | ||
| Pisum sativum | 84 | [44] | ||
| Cucumis sativus | 83 | [15] | ||
| Cucumis melo | 82 | [36] | ||
| Fagopyrum tataricum | 80 | [19] | ||
| Punica granatum | 73 | [45] | ||
| Vitis vinifera | 70 | VvNAC08/VvNAC30 | Disease resistance | [36,46] |
| Camellia oleifera | 67 | [36] | ||
| Petunia hybrida | 41 | [31] |
| Name of the Species | Gene Name | Biocompression | Adjustment Method | References |
|---|---|---|---|---|
| Lycopersicon esculentum | SlNAP1 | Leaf spot disease bacterial blight | SA | [38] |
| SNAC4 | Gray mold | Change-related genes | [39] | |
| Vitis vinifera L. | VvNAC08, VvNAC30, VvNAC36, VvNAC39, VvNAC44 | [46] | ||
| Saharum officinarum | ScATAF1 | Smut | Reduction of ROS | [26] |
| Solanum melongena L. | SmNAC | Blue blight | SA | [17] |
| Solanum tuberosum L. | StNAC103 | Soft rot | SCW | [18] |
| Genes | Stress Type | Function/Mechanism | References |
|---|---|---|---|
| MdNAC029, EjNAC3 | Cold Stress | Binding Related Proteins | [71,84] |
| MaNAC25, MaNAC28 | Transcription Factor Interaction | [81] | |
| MaNAC1, CaNAC064, EjXND1 | Hormone Regulation | [80,82,83] | |
| SsNAC23 | Others | [85] | |
| PtNAC101, ThNAC13 | Salt Stress | Antioxidant Enzyme Activity | [25,97] |
| OsNAM2 | Ion Transport | [103] | |
| MpNAC1, SNAC1 | Hormone Regulation | [98,103] | |
| MpNAC1, ZmNAC84, MbNAC25, PgNAC2, CaNAC46, OsNAM2, GhNAC4 | Binding Related Proteins | [98,99,100,102] | |
| OsNAC041 | Others | [120] | |
| PwNAC1 | Drought Stress | Binding Related Proteins | [20] |
| GhNAC3, SlNAC6, FtNAC31, LpNAC13 | Hormone Regulation | [91,93,94,95] | |
| SlNAC6, LpNAC13, MdNAC1 | Antioxidant Enzyme Activity | [14,93,95] | |
| ZmNAC55, MaNAC100, MaNAC136 | Others | [92,96] | |
| AcNRZ1 | Heavy Metal Stress | Binding Related Proteins | [119] |
| NAC32 | Ion Transport | [121] | |
| SmNAC1, AemNAC2, AemNAC3 | Change of Metal Content | [109] | |
| OsNAC300, SiNAC004, SiNAC120, ANAC017 | Others | [107,110,112] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zheng, C.; Yang, Q.; Wang, X.; Chen, Y.; He, R.; Li, X.; Pan, H.; Zhuo, R.; Qu, T.; Qiu, W. Transcription Factors Involved in Plant Stress and Growth and Development: NAC. Agronomy 2025, 15, 949. https://doi.org/10.3390/agronomy15040949
Zheng C, Yang Q, Wang X, Chen Y, He R, Li X, Pan H, Zhuo R, Qu T, Qiu W. Transcription Factors Involved in Plant Stress and Growth and Development: NAC. Agronomy. 2025; 15(4):949. https://doi.org/10.3390/agronomy15040949
Chicago/Turabian StyleZheng, Chenjia, Qin Yang, Xin Wang, Yu Chen, Ruoyu He, Xinmeng Li, Huanhuan Pan, Renying Zhuo, Tongbao Qu, and Wenmin Qiu. 2025. "Transcription Factors Involved in Plant Stress and Growth and Development: NAC" Agronomy 15, no. 4: 949. https://doi.org/10.3390/agronomy15040949
APA StyleZheng, C., Yang, Q., Wang, X., Chen, Y., He, R., Li, X., Pan, H., Zhuo, R., Qu, T., & Qiu, W. (2025). Transcription Factors Involved in Plant Stress and Growth and Development: NAC. Agronomy, 15(4), 949. https://doi.org/10.3390/agronomy15040949