
Evaluation of Dual-Purpose Triticale: Grain and Forage Productivity and Quality Under Semi-Arid Conditions
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Description of Experimental Site and Growing Conditions
2.3. Experimental Setup
2.4. Forage and Grain Sample Collection and Preparation
2.5. Near-Infrared Reflectance Spectroscopy (NIRS) Analysis
2.6. Evaluation of Phenotypic Traits
2.7. Statistical Analysis
3. Results
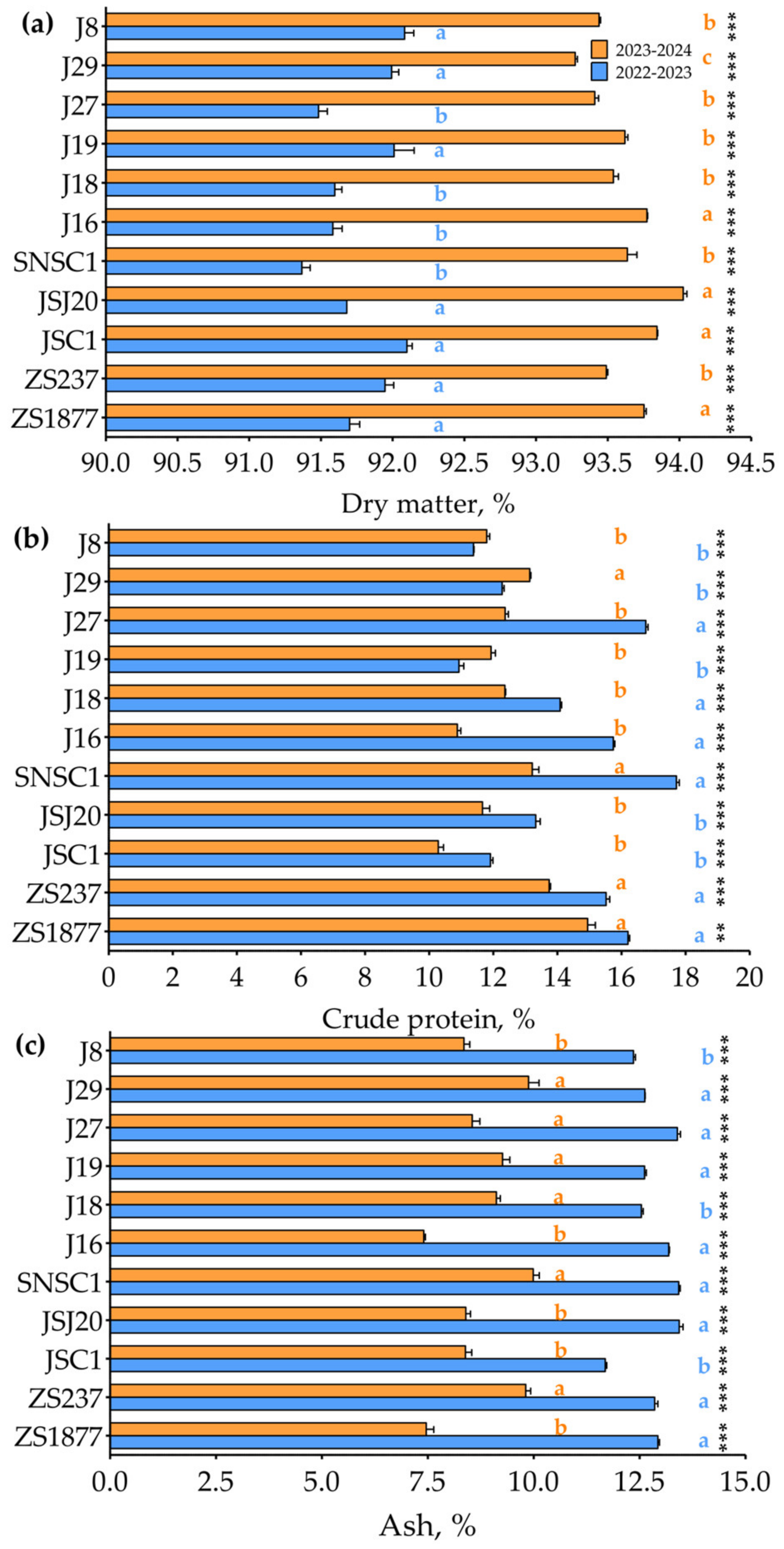
3.1. Assessment of Forage End-Use Quality of Triticale Genotypes Across Two Growing Seasons
3.1.1. Nutrient Composition
3.1.2. Mineral Content
3.1.3. Fiber and Carbohydrate Characteristics
3.1.4. Energy Characteristics
3.2. Assessment of Grain End-Use Quality of Triticale Genotypes Across Two Growing Seasons
3.2.1. Nutrient Composition
3.2.2. Carbohydrate and Fiber Characteristics
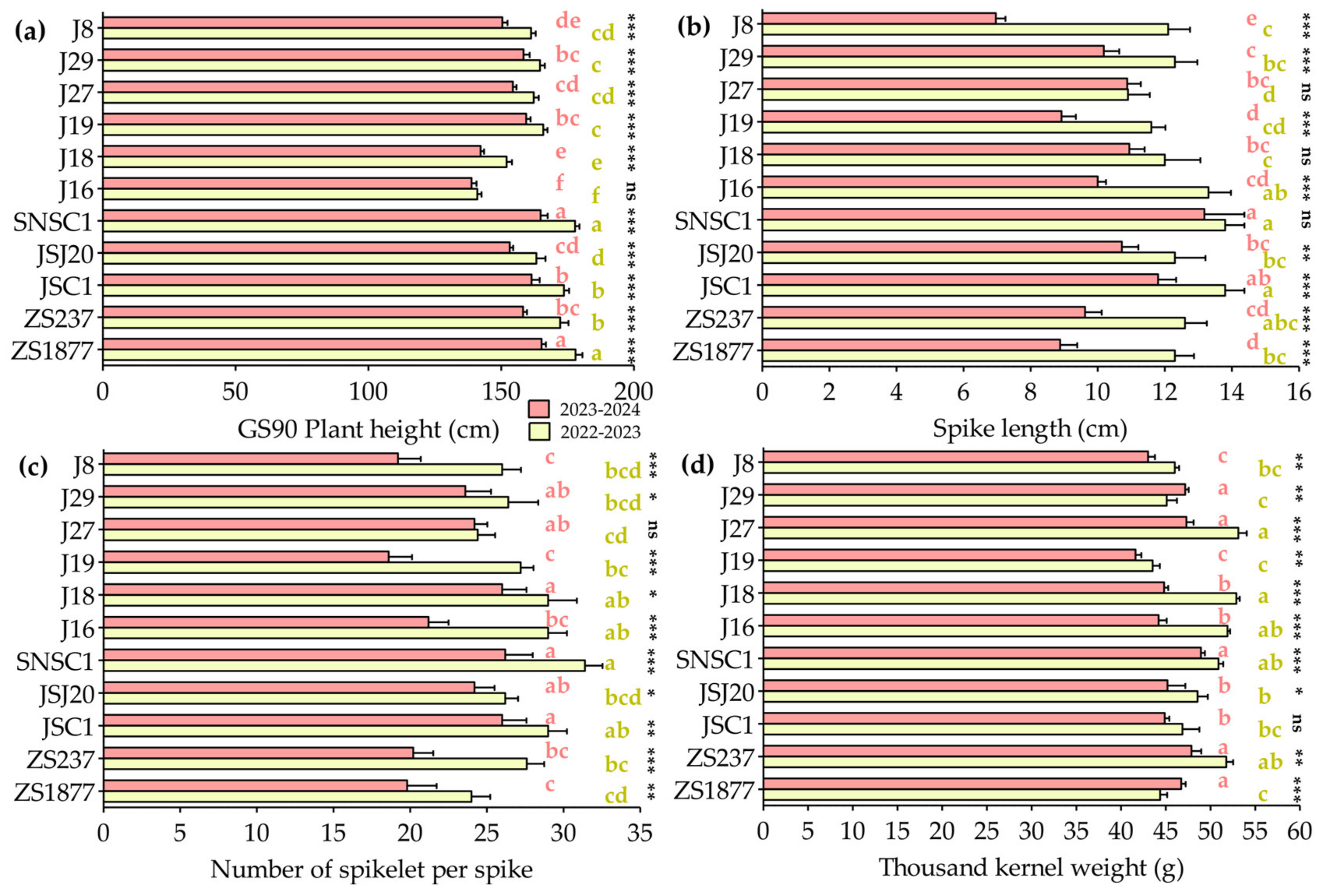
3.3. Evaluation of Agronomic Traits and Yield Performance of Triticale Genotypes Across Two Growing Seasons
3.4. Correlation Between Agronomic Traits, Forage, and Grain Quality in Triticale
4. Discussion
4.1. Grain and Forage Biomass Yield of Dual-Purpose Triticale Under Semi-Arid Conditions
4.2. Impact of Environmental Factors on Forage Quality
4.3. Trait Interactions and Their Implications for Dual-Purpose Triticale Breeding
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Poysa, V.W. Effect of forage harvest on grain yield and agronomic performance of winter triticale, wheat and rye. Can. J. Plant Sci. 1985, 65, 879–888. [Google Scholar] [CrossRef]
- Ammar, K.; Mergoum, M.; Rajaram, S. The history and evolution of triticale. In Triticale Improvement and Production; Mergoum, M., Gómez-Macpherson, H., Eds.; FAO: Rome, Italy, 2004; pp. 1–11. [Google Scholar]
- Mergoum, M.; Singh, P.K.; Peña, R.J.; Lozano-del Río, A.J.; Cooper, K.V.; Salmon, D.F.; Macpherson, H.G. Triticale: A “new” crop with old challenges. In Cereals; Carena, M.J., Ed.; Springer: New York, NY, USA, 2009; pp. 267–287. [Google Scholar] [CrossRef]
- Mergoum, M.; Sapkota, S.; ElDoliefy, A.E.A.; Naraghi, S.M.; Pirseyedi, S.; Alamri, M.S.; AbuHammad, W. Triticale (×Triticosecale Wittmack) Breeding. In Advances in Plant Breeding Strategies: Cereals; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland, 2019; pp. 405–451. [Google Scholar] [CrossRef]
- Arseniuk, E. Triticale abiotic stresses-an overview. Commun. Agric. Appl. Biol. Sci. 2015, 79, 82–100. [Google Scholar] [CrossRef]
- Oettler, G. The fortune of a botanical curiosity–triticale: Past, present and future. J. Agric. Sci. 2005, 143, 329–346. [Google Scholar] [CrossRef]
- Blum, A. The abiotic stress response and adaptation of triticale-a review. Cereal Res. Commun. 2014, 42, 359–375. [Google Scholar] [CrossRef]
- Garcia del Moral, L.F.; Boujenna, A.; Yanez, J.A.; Ramos, J.M. Forage production, grain yield, and protein content in dual-purpose triticale grown for both grain and forage. Agron. J. 1995, 87, 902–908. [Google Scholar] [CrossRef]
- Grabovets, A.I.; Dremucheva, G.F.; Karchevskaya, O.E. Breeding of triticale for baking purposes. Russ. Agric. Sci. 2013, 39, 197–202. [Google Scholar] [CrossRef]
- Baron, V.S.; Juskiw, P.E.; Aljarrah, M. Triticale as a forage. In Triticale; Eudes, F., Ed.; Springer: Cham, Switzerland, 2015; pp. 189–212. [Google Scholar] [CrossRef]
- Royo, C.; Insa, J.A.; Boujenna, A.; Ramos, J.M.; Montesinos, E. Yield and quality of spring triticale used for forage and grain as influenced by sowing date and cutting stage. Field Crops Res. 1994, 37, 161–168. [Google Scholar] [CrossRef]
- Wang, Z.; Sun, Y.; Ammar, K.; Bonjean, A.P.; Chen, X.; Sun, F. Development of triticale in China. In Cereals in China; He, Z., Bonjean, A.P.A., Eds.; CIMMYT: El Batán, Mexico, 2010; pp. 79–86. [Google Scholar]
- Lelley, T. Triticale: A low-input cereal with untapped potential. In Genetic Resources Chromosome Engineering and Crop Improvement; Singh, R.J., Jauhar, P.P., Eds.; CRC Press: Boca Raton, FL, USA, 2006; Volume 2, pp. 395–430. [Google Scholar]
- Cazzato, E.; Laudadio, V.; Tufarelli, V. Effects of harvest period, nitrogen fertilization and mycorrhizal fungus inoculation on triticale (×Triticosecale Wittmack) forage yield and quality. Renew. Agr. Food Syst. 2012, 27, 278–286. [Google Scholar] [CrossRef]
- McGoverin, C.M.; Snyders, F.; Muller, N.; Botes, W.; Fox, G.; Manley, M. A review of triticale uses and the effect of growth environment on grain quality. J. Sci. Food Agric. 2011, 91, 1155–1165. [Google Scholar] [CrossRef]
- Dumbravă, M.; Ion, V.; Epure, L.I.; Băşa, A.G.; Ion, N.; Duşa, E.M. Grain yield and yield components at triticale under different technological conditions. Agric. Agric. Sci. Procedia 2016, 10, 94–103. [Google Scholar] [CrossRef]
- Gill, K.S.; Omokanye, A.T. Spring triticale varieties forage yield, nutrients composition and suitability for beef cattle production. J. Agric. Sci. 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Bilgili, U.; Cifci, E.A.; Hanoglu, H.; Yagdi, K.; Acikgoz, E. Yield and quality of triticale forage. J. Food Agric. Environ. 2009, 7, 556–560. [Google Scholar]
- van Soest, P.J. Nutritional Ecology of the Ruminant; Cornell University Press: New York, NY, USA, 1994. [Google Scholar]
- Motzo, R.; Giunta, F.; Deidda, M. Factors affecting the genotype × environment interaction in spring triticale grown in a Mediterranean environment. Euphytica. 2001, 121, 317–324. [Google Scholar] [CrossRef]
- Milgate, A.; Ovenden, B.; Adorada, D.; Lisle, C.; Lacy, J.; Coombes, N. Genetic improvement of triticale for irrigated systems in south-eastern Australia: A study of genotype and genotype× environment interactions. Crop Pasture Sci. 2015, 66, 782–792. [Google Scholar] [CrossRef]
- Shanxi Meteorological Bureau. Available online: http://sx.cma.gov.cn/ (accessed on 24 August 2024).
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Sadreddine, B.E.J.I. Yield and quality of dual-purpose barley and triticale in a semi-arid environment in Tunisia. Afr. J. Agric. Res. 2016, 11, 2730–2735. [Google Scholar] [CrossRef]
- Shanazari, M.; Golkar, P.; Maibody, A.M.M. Effects of drought stress on some agronomic and bio-physiological traits of Trititicum aestivum, Triticale, and Tritipyrum genotypes. Arch. Agron. Soil Sci. 2018, 64, 2005–2018. [Google Scholar] [CrossRef]
- Mergoum, M.; Gómez-Macpherson, H. Triticale Improvement and Production; FAO: Rome, Italy, 2004; p. 179. [Google Scholar]
- Ayalew, H.; Kumssa, T.T.; Butler, T.J.; Ma, X.F. Triticale improvement for forage and cover crop uses in the southern great plains of the United States. Front. Plant Sci. 2018, 9, 1130. [Google Scholar] [CrossRef]
- Giunta, F.; Pruneddu, G.; Motzo, R. Grain yield and grain protein of old and modern durum wheat cultivars grown under different cropping systems. Field Crops Res. 2019, 230, 107–120. [Google Scholar] [CrossRef]
- Raffrenato, E.; Fievisohn, R.; Cotanch, K.W.; Grant, R.J.; Chase, L.E.; Van Amburgh, M.E. Effect of lignin linkages with other plant cell wall components on in vitro and in vivo neutral detergent fiber digestibility and rate of digestion of grass forages. J. Dairy Sci. 2017, 100, 8119–8131. [Google Scholar] [CrossRef]
- Jung, H.G.; Casler, M.D. Maize stem tissues: Impact of development on cell wall degradability. Crop Sci. 2006, 46, 1801–1809. [Google Scholar] [CrossRef]
- Hu, Y.; Schmidhalter, U. Drought and salinity: A comparison of their effects on mineral nutrition of plants. J. Plant Nutri. Soil Sci. 2005, 168, 541–549. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Prueger, J.H. Temperature extremes: Effect on plant growth and development. Weather Clim. Extrem. 2015, 10, 4–10. [Google Scholar] [CrossRef]
- Sánchez-Díaz, M.F.; Kramer, P.J. Behavior of corn and sorghum under water stress and during recovery. Plant Physiol. 1971, 48, 613–616. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.S.M.A.; Fujita, D.B.S.M.A.; Basra, S.M. Plant drought stress: Effects, mechanisms and management. Sustain. Agric. 2009, 29, 153–188. [Google Scholar] [CrossRef]
- Reynolds, M.; Foulkes, M.J.; Slafer, G.A.; Berry, P.; Parry, M.A.; Snape, J.W.; Angus, W.J. Raising yield potential in wheat. J. Exp. Bot. 2009, 60, 1899–1918. [Google Scholar] [CrossRef]
- Marcos-Barbero, E.L.; Pérez, P.; Martínez-Carrasco, R.; Arellano, J.B.; Morcuende, R. Screening for higher grain yield and biomass among sixty bread wheat genotypes grown under elevated CO2 and high-temperature conditions. Plants 2021, 10, 1596. [Google Scholar] [CrossRef]
- Simmonds, N.W. The relation between yield and protein in cereal grain. J. Sci. Food Agric. 1995, 67, 309–315. [Google Scholar] [CrossRef]
- Martre, P.; He, J.; Le Gouis, J.; Semenov, M.A. In silico system analysis of physiological traits determining grain yield and protein concentration for wheat as influenced by climate and crop management. J. Exp. Bot. 2015, 66, 3581–3598. [Google Scholar] [CrossRef]
- Shewry, P.R.; Pellny, T.K.; Lovegrove, A. Is modern wheat bad for health? Nat. Plants 2016, 2, 16097. [Google Scholar] [CrossRef]
- Poudel, M.R.; Poudel, P.B.; Puri, R.R.; Paudel, H.K. Variability, correlation and path coefficient analysis for agro-morphological traits in wheat genotypes (Triticum aestivum L.) under normal and heat stress conditions. Int. J. Appl. Sci. Biotechnol. 2021, 9, 65–74. [Google Scholar] [CrossRef]
- Tanin, M.J.; Sharma, A.; Saini, D.K.; Singh, S.; Kashyap, L.; Srivastava, P.; Mavi, G.S.; Kaur, S.; Kumar, V.; Kumar, V.; et al. Ascertaining yield and grain protein content stability in wheat genotypes having the Gpc-B1 gene using univariate, multivariate, and correlation analysis. Front. Genet. 2022, 13, 1001904. [Google Scholar] [CrossRef]
- Casler, M.D.; Jung, H.J.G. Relationships of fibre, lignin, and phenolics to in vitro fibre digestibility in three perennial grasses. Anim Feed Sci Technol. 2006, 125, 151–161. [Google Scholar] [CrossRef]
- Reuter, D.; Robinson, J.B. Plant Analysis: An Interpretation Manual; CSIRO Publishing: Collingwood, VIC, Australia, 1997. [Google Scholar]
- Perelman, A.; Imas, P.; Bansal, S.K. Role of potassium for improving nutrient use efficiency in agriculture. In Input Use Efficiency for Food and Environmental Security; Bhatt, R., Meena, R.S., Hossain, A., Eds.; Springer Nature: Singapore, 2021; pp. 397–420. [Google Scholar] [CrossRef]
- Jaiswal, D.K.; Verma, J.P.; Prakash, S.; Meena, V.S.; Meena, R.S. Potassium as an important plant nutrient in sustainable agriculture: A state of the art. In Potassium Solubilizing Microorganisms for Sustainable Agriculture; Meena, V., Maurya, B., Verma, J., Meena, R., Eds.; Springer: New Delhi, India, 2016; pp. 21–29. [Google Scholar] [CrossRef]
- Jung, H.G.; Allen, M.S. Characteristics of plant cell walls affecting intake and digestibility of forages by ruminants. J. Anim. Sci. 1995, 73, 2774–2790. [Google Scholar] [CrossRef] [PubMed]
- Katoch, R. Forage nutritional quality management. In Techniques in Forage Quality Analysis; Springer: Singapore, 2023; pp. 211–221. [Google Scholar] [CrossRef]
- Buxton, D.R.; Casler, M.D. Environmental and genetic effects on cell wall composition and digestibility. In Forage Cell Wall Structure and Digestibility; Jung, H.G., Buxton, D.R., Hatfield, R.D., Ralph, J., Eds.; American Society of Agronomy, Crop Science Society of America: Madison, WI, USA, 1993; pp. 685–714. [Google Scholar] [CrossRef]
- Hoffman, P.C.; Shaver, R.D.; Combs, D.K.; Undersander, D.J.; Bauman, L.M.; Seeger, T.K. Understanding NDF digestibility of forages. Focus Forage 2001, 3, 1–3. [Google Scholar]
- DuPont, F.M.; Altenbach, S.B. Molecular and biochemical impacts of environmental factors on wheat grain development and protein synthesis. J. Cereal Sci. 2003, 38, 133–146. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Year | Genotype | Year × Genotype |
|---|---|---|---|
| Degree of freedom | 1 | 10 | 9 |
| Grain yield (kg ha⁻1) | 13,370,679.61 *** | 2,754,475.72 *** | 70,776.24 ns |
| Forage biomass (kg ha⁻1) | 92,844,254.93 *** | 66,968,931.47 *** | 1,120,807.55 ns |
| GS59 Plant height (cm) | 531.04 *** | 390.99 *** | 6.69 ns |
| GS90 Plant height (cm) | 1520.07 *** | 569.47 *** | 28.69 *** |
| Spike length (cm) | 79.58 *** | 8.38 *** | 4.36 *** |
| Number of spikelets per spike | 380.02 *** | 37.11 *** | 12.65 *** |
| Thousand kernel weight (g) | 200.21 *** | 40.54 *** | 14.63 *** |
| Factor | Grain Yield (kg/ha) | Forage Biomass (kg/ha) | GS59 Plant Height (cm) | ||||
|---|---|---|---|---|---|---|---|
| Genotype | ZS1877 | 5745 | b | 26,398.60 | b | 145.5 | b |
| ZS237 | 6449.9 | cd | 28,134.30 | c | 140.4 | b | |
| JSC1 | 6722 | d | 28,971.00 | d | 142.3 | b | |
| JSJ20 | 6816.9 | de | 28,929.40 | d | 137.8 | b | |
| SNSC1 | 6417 | c | 29,660.30 | e | 144.8 | b | |
| J16 | 4830.2 | a | 20,059.10 | a | 123.6 | a | |
| J18 | 5429.7 | b | 21,262.10 | a | 124.3 | a | |
| J19 | 6924.3 | e | 28,925.20 | d | 136.9 | b | |
| J27 | 6882.2 | e | 28,059.40 | c | 149 | c | |
| J29 | 6432.9 | cd | 28,367.40 | c | 143.5 | b | |
| J8 | 6670.4 | d | 29,774.70 | e | 138.6 | b | |
| Year | 2022–2023 | 6752 | b | 28,326.20 | b | 141.6 | b |
| 2023–2024 | 5851.8 | a | 25,957.60 | a | 135.9 | a | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, L.; Xu, L.; Wang, H.; Fan, X.; Yan, C.; Zhang, Y.; Jiang, C.; Zhou, T.; Guo, Q.; Sun, Y.; et al. Evaluation of Dual-Purpose Triticale: Grain and Forage Productivity and Quality Under Semi-Arid Conditions. Agronomy 2025, 15, 881. https://doi.org/10.3390/agronomy15040881
Cui L, Xu L, Wang H, Fan X, Yan C, Zhang Y, Jiang C, Zhou T, Guo Q, Sun Y, et al. Evaluation of Dual-Purpose Triticale: Grain and Forage Productivity and Quality Under Semi-Arid Conditions. Agronomy. 2025; 15(4):881. https://doi.org/10.3390/agronomy15040881
Chicago/Turabian StyleCui, Lei, Linyuan Xu, Huihui Wang, Xiangtian Fan, Chahong Yan, Yanming Zhang, Changtong Jiang, Tong Zhou, Qing Guo, Yu Sun, and et al. 2025. "Evaluation of Dual-Purpose Triticale: Grain and Forage Productivity and Quality Under Semi-Arid Conditions" Agronomy 15, no. 4: 881. https://doi.org/10.3390/agronomy15040881
APA StyleCui, L., Xu, L., Wang, H., Fan, X., Yan, C., Zhang, Y., Jiang, C., Zhou, T., Guo, Q., Sun, Y., Yang, F., & Li, H. (2025). Evaluation of Dual-Purpose Triticale: Grain and Forage Productivity and Quality Under Semi-Arid Conditions. Agronomy, 15(4), 881. https://doi.org/10.3390/agronomy15040881