

Soil Carbon Sequestration in Nothofagus obliqua Forests with Different Canopy Cover Levels Under Silvopastoral Management
 ,
,  , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Study Area
2.2. Conditioning of Experimental Treatments
2.3. Soil Sampling
2.4. Soil Analysis
2.4.1. Evaluation of Physical Parameters
2.4.2. Evaluation of Chemical Parameters
2.4.3. Evaluation of Biological Parameters
2.4.4. Soil Physical Fractionation
2.4.5. Total Carbon, Oxidizable Carbon, Non-Oxidizable Carbon, and Carbon Stocks
2.5. Statistical Analysis
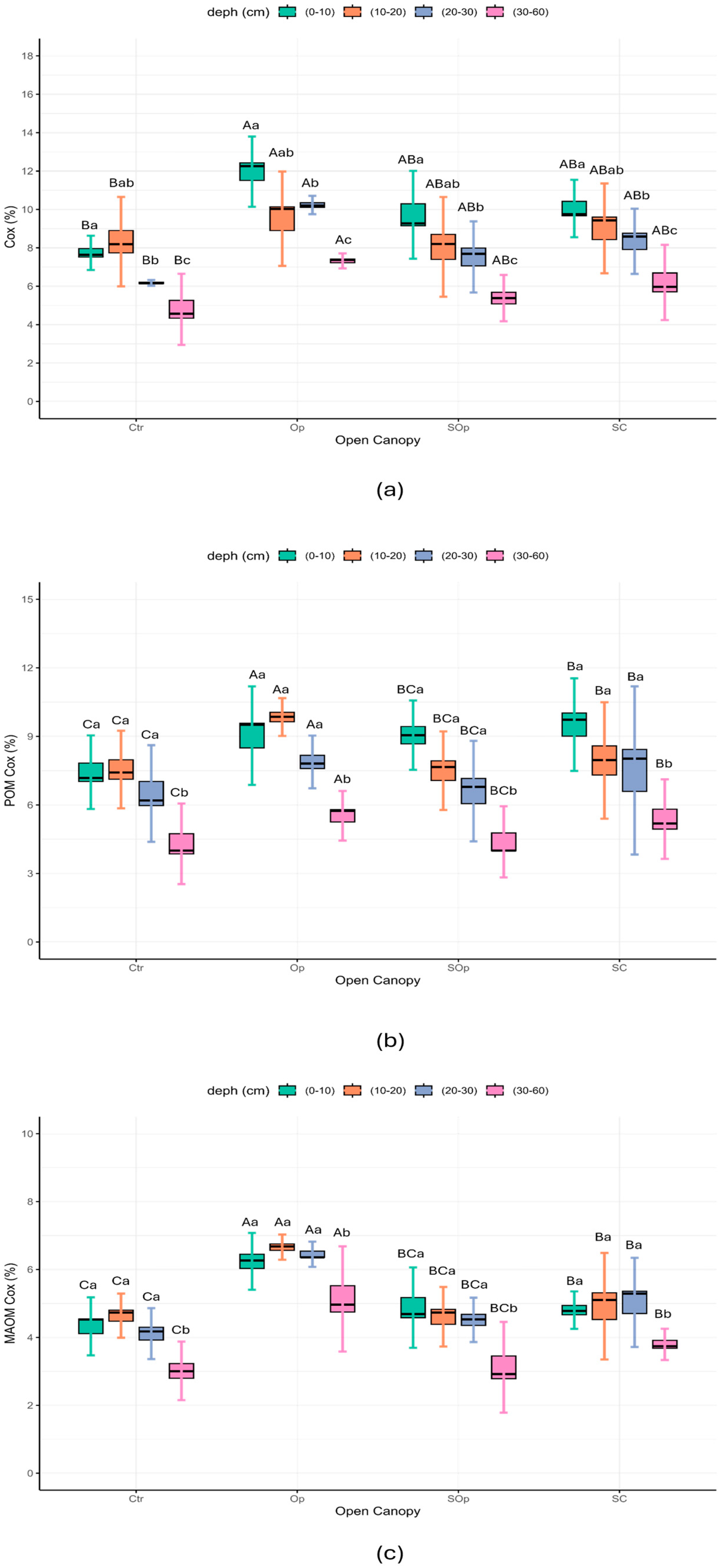
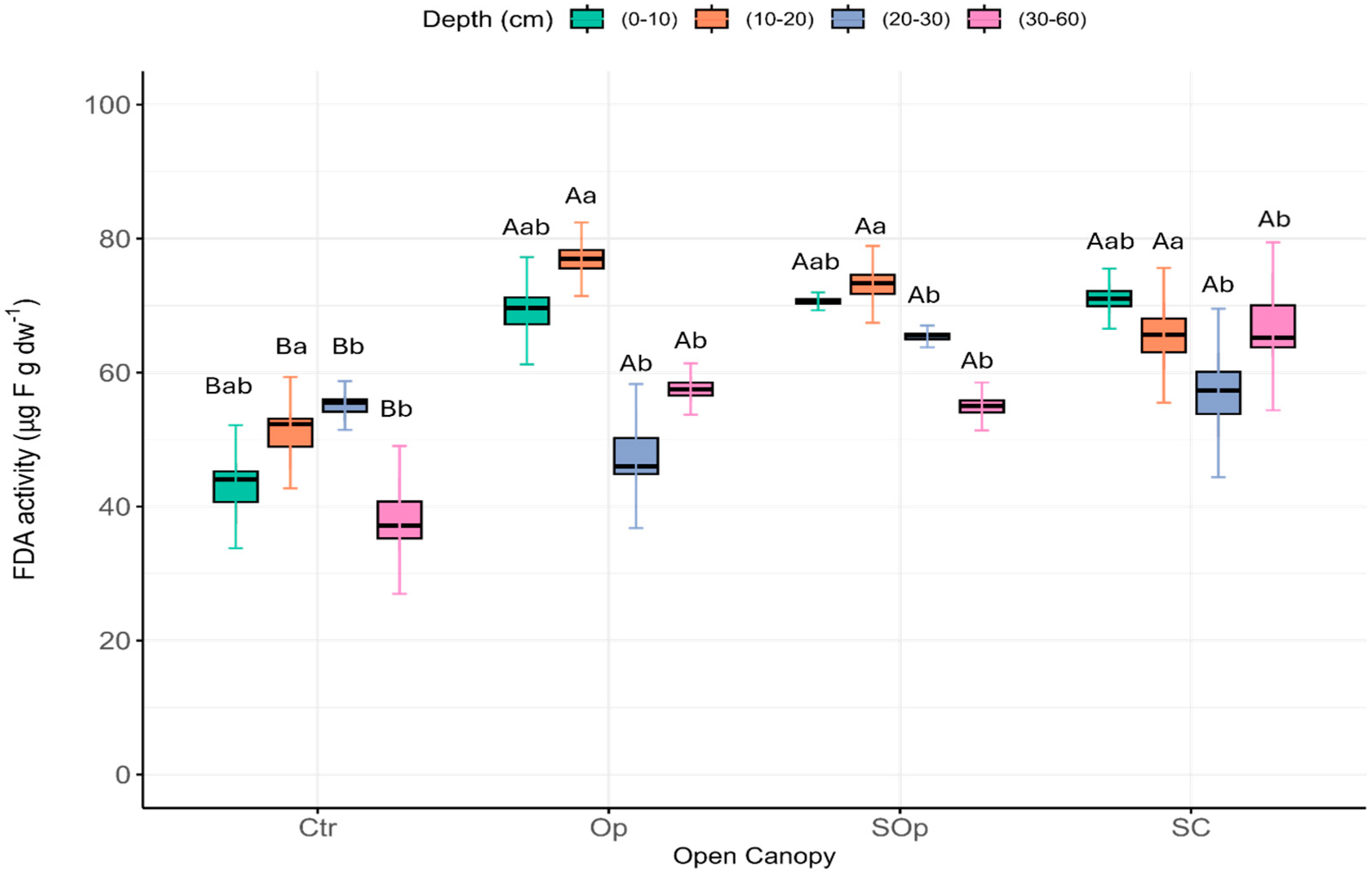
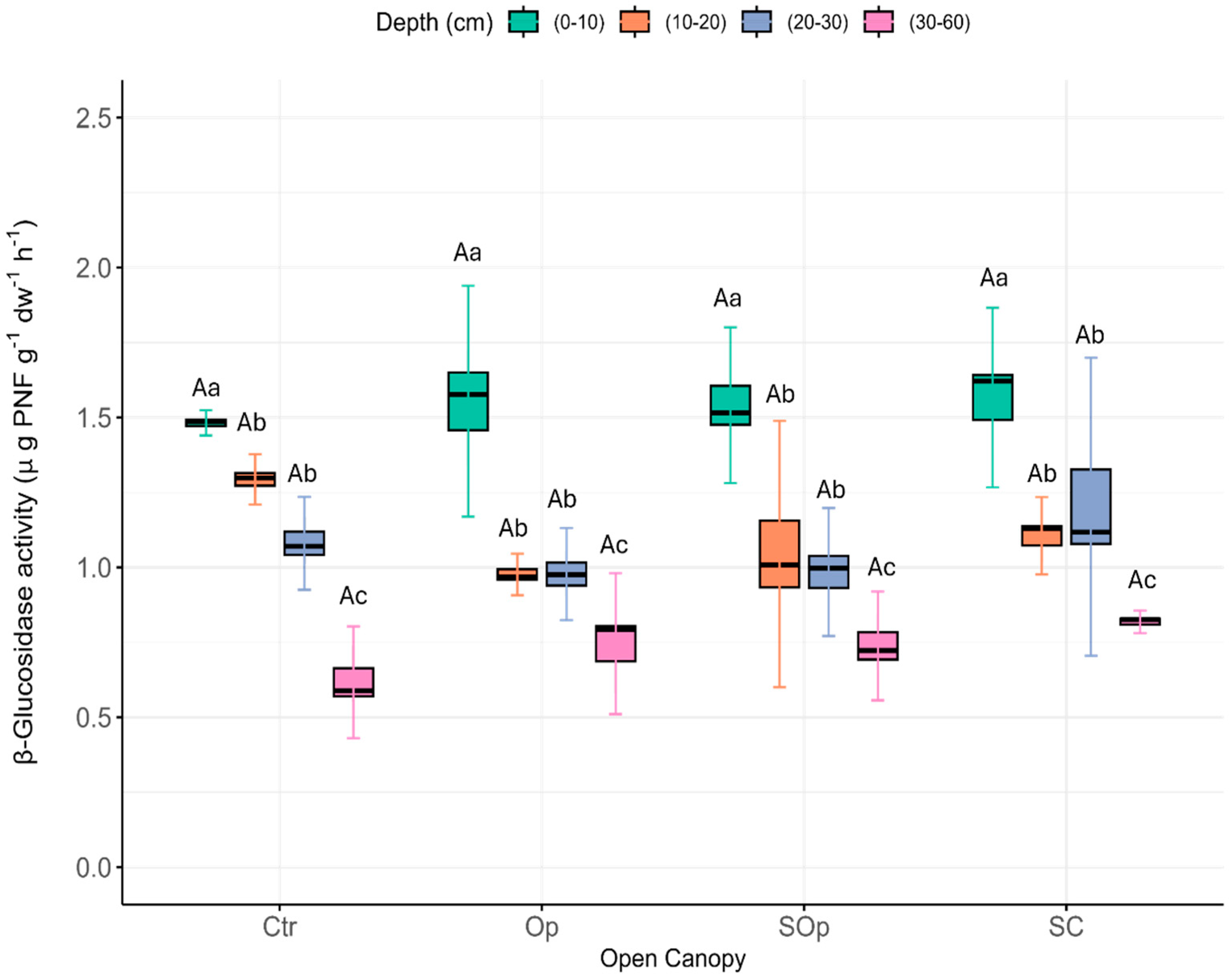
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Description |
| Mha | million hectares |
| SPSs | silvopastoral systems |
| C | carbon |
| SOC | soil organic carbon |
| IPCC | Intergovernmental Panel on Climate Change |
| FAO | Food and Agriculture Organization of the United Nations |
| SOM | soil organic matter |
| POM | particulate organic matter |
| MAOM | matrix-associated organic matter minerals |
| N | nitrogen |
| Op | open |
| SOp | semi-open |
| SC | semi-closed |
| Ctr | control |
| WFPS | water-filled pore space |
| BD | bulk density |
| PD | particle density |
| OM | organic matter |
| P | phosphorous |
| K+ | potassium |
| ECEC | effective cation exchange capacity |
| AlEXCH | exchangeable aluminum |
| Ca2+EXCH | exchangeable calcium |
| Mg2+EXCH | exchangeable magnesium |
| NH4+ | ammonium |
| NO3− | nitrate |
| K+EXCH | exchangeable potassium |
| Na+EXCH | exchangeable sodium |
| AlSAT | aluminum saturation |
| K+SAT | potassium saturation |
| Ca2+SAT | calcium saturation |
| Mg2+SAT | magnesium saturation |
| FDA | fluorescein diacetate |
| CO2 | carbon dioxide |
| TC | total carbon |
| Cox | oxidizable carbon |
| TOC | total organic carbon |
| Cnox | non-oxidizable carbon |
| C stock | carbon stock |
| PyC | pyrogenic carbon |
| C/N | C:N ratio |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Condition/ Depths | NH4+ | NO3− | K+EXCH | Na+EXCH | AlSAT | K+SAT | Ca2+SAT | Mg2+SAT |
|---|---|---|---|---|---|---|---|---|
| Ctr (0–30) | 3.06 ± 0.07 | 14.7 ± 4.88 | 0.28 ± 1.34 | 0.07 ± 0.00 | 3.85 ± 2.19 | 8.64 ± 2.77 | 70.79 ± 8.49 | 13.75 ± 2.04 |
| Ctr (30–60) | 2.50 ± 0.00 | 5.90 ± 3.18 | 0.10 ± 0.59 | 0.07 ± 0.00 | 2.20 ± 0.43 | 8.72 ± 3.78 | 64.84 ± 1.32 | 17.24 ± 2.29 |
| OP (0–30) | 9.53 ± 0.05 | 11.60 ± 0.05 | 0.23 ± 0.05 | 0.04 ± 0.05 | 9.40 ± 0.05 | 12.54 ± 0.05 | 63.41 ± 0.05 | 12.85 ± 0.05 |
| OP (30–60) | 8.63 ± 0.04 | 5.80 ± 0.04 | 0.12 ± 0.04 | 0.04 ± 0.04 | 8.69 ± 0.04 | 13.60 ± 0.04 | 54.12 ± 0.04 | 17.81 ± 0.04 |
| SOp (0–30) | 12.83 ± 1.87 | 7.77 ± 1.87 | 0.16 ± 0.05 | 0.03 ± 0.00 | 5.14 ± 3.49 | 8.51 ± 2.64 | 71.12 ± 10.20 | 12.89 ± 2.77 |
| SOp (30–60) | 6.47 ± 1.83 | 4.37 ± 1.83 | 0.12 ± 0.06 | 0.07 ± 0.00 | 1.46 ± 0.53 | 9.88 ± 1.77 | 65.83 ± 6.86 | 15.82 ± 4.46 |
| SC (0–30) | 12.23 ± 3.03 | 14.7 ± 3.03 | 0.23 ± 0.05 | 0.04 ± 0.00 | 6.98 ± 7.49 | 7.87 ± 2.51 | 71.37 ± 11.92 | 12.30 ± 1.63 |
| SC (30–60) | 7.23 ± 0.37 | 10.07 ± 0.37 | 0.21 ± 0.10 | 0.07 ± 0.03 | 8.08 ± 8.28 | 11.28 ± 1.88 | 61.77 ± 14.10 | 14.54 ± 2.89 |
Appendix B
References
- Casanova, M.; Salazar, O.; Seguel, O.; Luzio, W. Human-Induced Soil Degradation in Chile. In The Soils of Chile; World Soils Book Series; Springer: Dordrecht, The Netherlands, 2013; pp. 121–158. ISBN 978-94-007-5948-0. [Google Scholar]
- Flores, J.P.; Martínez, E.; Espinosa, M.; Avendaño, P.; Ahumada, I.; Henríquez, G.; Torres, P. Determinación de La Erosión Actual y Potencial de Los Suelos de Chile: Región Del Bío Bío. Síntesis de Resultados.(Pub. Ciren N° 148). 2010. Available online: https://bibliotecadigital.ciren.cl/items/114f1067-d6bb-4e0f-aa24-b259ad15e901 (accessed on 27 March 2025).
- Lara, A.; Solari, M.E.; Prieto, M.D.R.; Peña, M.P. Reconstrucción de La Cobertura de La Vegetación y Uso Del Suelo Hacia 1550 y Sus Cambios a 2007 En La Ecorregión de Los Bosques Valdivianos Lluviosos de Chile (35°–43° 30′ S). Bosque Valdivia 2012, 33, 3–4. [Google Scholar] [CrossRef]
- Nair, P.K.R.; Kumar, B.M.; Nair, V.D. Carbon Sequestration and Climate Change Mitigation. In An Introduction to Agroforestry; Springer International Publishing: Cham, Switzerland, 2021; pp. 487–537. ISBN 978-3-030-75357-3. [Google Scholar]
- Udawatta, R.P.; Gantzer, C.J.; Jose, S. Agroforestry Practices and Soil Ecosystem Services. In Soil Health and Intensification of Agroecosytems; Elsevier: Amsterdam, The Netherlands, 2017; pp. 305–333. ISBN 978-0-12-805317-1. [Google Scholar]
- Yasin, G.; Nawaz, M.F.; Sinha, D.; Qadir, I.; Altaf, M.; Ashraf, M.N.; Soufan, W.; Mammadov, A.; Zulfiqar, U.; Rahman, S.U. Agroforestry Status, Services, and Its Role in Climate Change Mitigation through Carbon Sequestration under Semi-Arid Conditions. Trees For. People 2024, 17, 100640. [Google Scholar] [CrossRef]
- Ramachandran Nair, P.K.; Nair, V.D.; Mohan Kumar, B.; Showalter, J.M. Carbon Sequestration in Agroforestry Systems. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2010; Volume 108, pp. 237–307. ISBN 978-0-12-381031-1. [Google Scholar]
- Ortiz, J.; Dube, F.; Neira, P.; Hernández Valera, R.R.; De Souza Campos, P.M.; Panichini, M.; Pérez-San Martín, A.; Stolpe, N.B.; Zagal, E.; Curaqueo, G. Comparative Study between Silvopastoral and Agroforest Systems on Soil Quality in a Disturbed Native Forest of South-Central Chile. Agronomy 2023, 13, 2683. [Google Scholar] [CrossRef]
- IPCC. Summary for Policymakers; IPCC: Geneva, Switzerland, 2019. [Google Scholar]
- Cai, Z.; Aguilar, F.X. Economic Valuation of Agroforestry Ecosystem Services. In Agroforestry and Ecosystem Services; Udawatta, R.P., Jose, S., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 477–494. ISBN 978-3-030-80059-8. [Google Scholar]
- Kay, S.; Graves, A.; Palma, J.H.N.; Moreno, G.; Roces-Díaz, J.V.; Aviron, S.; Chouvardas, D.; Crous-Duran, J.; Ferreiro-Domínguez, N.; García De Jalón, S.; et al. Agroforestry Is Paying off—Economic Evaluation of Ecosystem Services in European Landscapes with and without Agroforestry Systems. Ecosyst. Serv. 2019, 36, 100896. [Google Scholar] [CrossRef]
- FAO. Scaling up Agroecology to Achieve the Sustainable Development Goals. In Proceedings of the 2nd FAO International Symposium on Agroecology, Rome, Italy, 3–5 April 2018; FAO: Rome, Italy, 2019. [Google Scholar]
- Smith, P.; Cotrufo, M.F.; Rumpel, C.; Paustian, K.; Kuikman, P.J.; Elliott, J.A.; McDowell, R.; Griffiths, R.I.; Asakawa, S.; Bustamante, M.; et al. Biogeochemical Cycles and Biodiversity as Key Drivers of Ecosystem Services Provided by Soils. Soil 2015, 1, 665–685. [Google Scholar] [CrossRef]
- Voltr, V.; Menšík, L.; Hlisnikovský, L.; Hruška, M.; Pokorný, E.; Pospíšilová, L. The Soil Organic Matter in Connection with Soil Properties and Soil Inputs. Agronomy 2021, 11, 779. [Google Scholar] [CrossRef]
- Hayes, M.H.B.; Swift, R.S. An Appreciation of the Contribution of Frank Stevenson to the Advancement of Studies of Soil Organic Matter and Humic Substances. J. Soils Sediments 2018, 18, 1212–1231. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Particulate Soil Organic-Matter Changes across a Grassland Cultivation Sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Ranalli, M.G.; Haddix, M.L.; Six, J.; Lugato, E. Soil Carbon Storage Informed by Particulate and Mineral-Associated Organic Matter. Nat. Geosci. 2019, 12, 989–994. [Google Scholar] [CrossRef]
- Follett, R.F.; Stewart, C.E.; Pruessner, E.G.; Kimble Retired, J.M. Great Plains Climate and Land-Use Effects on Soil Organic Carbon. Soil Sci. Soc. Am. J. 2015, 79, 261–271. [Google Scholar] [CrossRef]
- Heckman, K.; Hicks Pries, C.E.; Lawrence, C.R.; Rasmussen, C.; Crow, S.E.; Hoyt, A.M.; Von Fromm, S.F.; Shi, Z.; Stoner, S.; McGrath, C.; et al. Beyond Bulk: Density Fractions Explain Heterogeneity in Global Soil Carbon Abundance and Persistence. Glob. Change Biol. 2022, 28, 1178–1196. [Google Scholar] [CrossRef]
- Witzgall, K.; Vidal, A.; Schubert, D.I.; Höschen, C.; Schweizer, S.A.; Buegger, F.; Pouteau, V.; Chenu, C.; Mueller, C.W. Particulate Organic Matter as a Functional Soil Component for Persistent Soil Organic Carbon. Nat. Commun. 2021, 12, 4115. [Google Scholar] [CrossRef] [PubMed]
- Lavallee, J.M.; Soong, J.L.; Cotrufo, M.F. Conceptualizing Soil Organic Matter into Particulate and Mineral-associated Forms to Address Global Change in the 21st Century. Glob. Change Biol. 2020, 26, 261–273. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Lavallee, J.M. Soil Organic Matter Formation, Persistence, and Functioning: A Synthesis of Current Understanding to Inform Its Conservation and Regeneration. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2022; Volume 172, pp. 1–66. ISBN 978-0-323-98953-4. [Google Scholar]
- Rumpel, C.; Kögel-Knabner, I. Deep Soil Organic Matter—A Key but Poorly Understood Component of Terrestrial C Cycle. Plant Soil 2011, 338, 143–158. [Google Scholar] [CrossRef]
- Sandoval, E.; Dorner, F.; Seguel, S.; Cuevas, B.; Rivera, S. Métodos de Análisis Físicos del Suelo. 2012. Available online: https://hdl.handle.net/20.500.14001/59208 (accessed on 27 March 2025).
- Sadzawka, R.; Carrasco, R.; Adriana, M.; Grez, Z.; Mora, G.; de la Luz, M.; Flores, P.; Neaman, A. Métodos de Análisis Recomendados para los Suelos de Chile. Revisión 2006; SIDALC: Turrialba, Costa Rica, 2006. [Google Scholar]
- Alef, K. Nitrogen Mineralization in Soils. In Methods Appl. Soil Microbiol. Biochem.; Academic Press: London, UK, 1995; pp. 234–245. [Google Scholar]
- Tabatabai, M.A. Soil Enzymes. In SSSA Book Series; Weaver, R.W., Angle, S., Bottomley, P., Bezdicek, D., Smith, S., Tabatabai, A., Wollum, A., Eds.; Soil Science Society of America: Madison, WI, USA, 2018; pp. 775–833. ISBN 978-0-89118-865-0. [Google Scholar]
- Craine, J.M.; Fierer, N.; McLauchlan, K.K. Widespread Coupling between the Rate and Temperature Sensitivity of Organic Matter Decay. Nat. Geosci. 2010, 3, 854–857. [Google Scholar] [CrossRef]
- Wright, A.F.; Bailey, J.S. Organic Carbon, Total Carbon, and Total Nitrogen Determinations in Soils of Variable Calcium Carbonate Contents Using a Leco CN-2000 Dry Combustion Analyzer. Commun. Soil Sci. Plant Anal. 2001, 32, 3243–3258. [Google Scholar] [CrossRef]
- Gomez, F.; Von Müller, A.; Tarabini, M.; La Manna, L. Resilient Andisols under Silvopastoral Systems. Geoderma 2022, 418, 115843. [Google Scholar] [CrossRef]
- Ortiz, J.; Dube, F.; Neira, P.; Panichini, M.; Stolpe, N.B.; Zagal, E.; Martínez-Hernández, P.A. Soil Quality Changes within a (Nothofagus Obliqua) Forest Under Silvopastoral Management in the Andes Mountain Range, South Central Chile. Sustainability 2020, 12, 6815. [Google Scholar] [CrossRef]
- Nissen, J.; Quiroz, C.; Seguel, O.; Mac Donald, R.; Sch, A.E. Variacion del potencial matrico durante el movimiento de agua en andisoles. Agro Sur 2005, 33, 36–47. [Google Scholar] [CrossRef]
- Vásquez, H.V.; Valqui, L.; Bobadilla, L.G.; Arbizu, C.I.; Alegre, J.C.; Maicelo, J.L. Influence of Arboreal Components on the Physical-Chemical Characteristics of the Soil under Four Silvopastoral Systems in Northeastern Peru. Heliyon 2021, 7, e07725. [Google Scholar] [CrossRef]
- Doetterl, S.; Stevens, A.; Six, J.; Merckx, R.; Van Oost, K.; Casanova Pinto, M.; Casanova-Katny, A.; Muñoz, C.; Boudin, M.; Zagal Venegas, E.; et al. Soil Carbon Storage Controlled by Interactions between Geochemistry and Climate. Nat. Geosci. 2015, 8, 780–783. [Google Scholar] [CrossRef]
- Renwick, L.L.R.; Celedón, A.; Nájera, F.; Fuentes Espoz, J.-P.; Celedón, D.; Arellano, C.; Salazar, O. Integrated Crop-Livestock Farms Have Higher Topsoil Nitrogen and Carbon than Crop-Only Farms in Chilean Mediterranean Climate Volcanic Soils. Agric. Syst. 2025, 222, 104172. [Google Scholar] [CrossRef]
- Banerjee, A.; Jhariya, M.K.; Yadav, D.K.; Raj, A. (Eds.) Environmental and Sustainable Development Through Forestry and Other Resources; Apple Academic Press: New York, NY, USA, 2020; ISBN 978-0-429-27602-6. [Google Scholar]
- Borie, F.; Aguilera, P.; Castillo, C.; Valentine, A.; Seguel, A.; Barea, J.M.; Cornejo, P. Revisiting the Nature of Phosphorus Pools in Chilean Volcanic Soils as a Basis for Arbuscular Mycorrhizal Management in Plant P Acquisition. J. Soil Sci. Plant Nutr. 2019, 19, 390–401. [Google Scholar] [CrossRef]
- Lambers, H.; Finnegan, P.M.; Jost, R.; Plaxton, W.C.; Shane, M.W.; Stitt, M. Phosphorus Nutrition in Proteaceae and Beyond. Nat. Plants 2015, 1, 15109. [Google Scholar] [CrossRef]
- Poirier, V.; Basile-Doelsch, I.; Balesdent, J.; Borschneck, D.; Whalen, J.K.; Angers, D.A. Organo-Mineral Interactions Are More Important for Organic Matter Retention in Subsoil Than Topsoil. Soil Syst. 2020, 4, 4. [Google Scholar] [CrossRef]
- Reyes, F.; Lillo, A.; Ojeda, N.; Reyes, M.; Alvear, M. Efecto de La Exposición y La Toposecuencia Sobre Actividades Biológicas Del Suelo En Bosque Relicto Del Centro-Sur de Chile. Bosque Valdivia 2011, 32, 255–265. [Google Scholar] [CrossRef]
- Masebo, N.; Birhane, E.; Takele, S.; Lucena, J.J.; Araceli, P.-S.; Yunta, F.; Belay, Z.; Anjulo, A. Microbial Biomass Carbon Distribution under Agroforestry Practices and Soil Depth Variations in Southern Ethiopia. Agrofor. Syst. 2025, 99, 56. [Google Scholar] [CrossRef]
- Lavallee, J.M.; Conant, R.T.; Haddix, M.L.; Follett, R.F.; Bird, M.I.; Paul, E.A. Selective Preservation of Pyrogenic Carbon across Soil Organic Matter Fractions and Its Influence on Calculations of Carbon Mean Residence Times. Geoderma 2019, 354, 113866. [Google Scholar] [CrossRef]
- Poblete-Grant, P.; Suazo-Hernández, J.; Condron, L.; Rumpel, C.; Demanet, R.; Malone, S.L.; Mora, M.D.L.L. Soil Available P, Soil Organic Carbon and Aggregation as Affected by Long-Term Poultry Manure Application to Andisols under Pastures in Southern Chile. Geoderma Reg. 2020, 21, e00271. [Google Scholar] [CrossRef]
- Crovo, O.; Costa-Reidel, C.D.; Rodríguez, R.; Aburto, F. Livestock grazing reduces soil quality and threatens recovery of a degraded andean araucaria forest. 2021. Authorea 2021, preprints. [Google Scholar] [CrossRef]
- Alfaro, M.; Dube, F.; Zagal, E. Soil Quality Indicators in an Andisol under Different Tree Covers in Disturbed Nothofagus Forests. Chil. J. Agric. Res. 2018, 78, 106–116. [Google Scholar] [CrossRef]
- Batjes, N.H. Total Carbon and Nitrogen in the Soils of the World. Eur. J. Soil Sci. 2014, 65, 10–21. [Google Scholar] [CrossRef]
- Katsumi, T. Soil Excavation and Reclamation in Civil Engineering: Environmental Aspects. Soil Sci. Plant Nutr. 2015, 61, 22–29. [Google Scholar] [CrossRef]
- Decker, K.L.M.; Boerner, R.E.J. Mass Loss and Nutrient Release from Decomposing Evergreen and Deciduous Nothofagus Litters from the Chilean Andes. Austral Ecol. 2006, 31, 1005–1015. [Google Scholar] [CrossRef]
- Hall, S.J.; Ye, C.; Weintraub, S.R.; Hockaday, W.C. Molecular Trade-Offs in Soil Organic Carbon Composition at Continental Scale. Nat. Geosci. 2020, 13, 687–692. [Google Scholar] [CrossRef]
- Manral, V.; Bargali, K.; Bargali, S.S.; Karki, H.; Chaturvedi, R.K. Seasonal Dynamics of Soil Microbial Biomass C, N and P along an Altitudinal Gradient in Central Himalaya, India. Sustainability 2023, 15, 1651. [Google Scholar] [CrossRef]
- Angst, G.; Mueller, K.E.; Nierop, K.G.J.; Simpson, M.J. Plant- or Microbial-Derived? A Review on the Molecular Composition of Stabilized Soil Organic Matter. Soil Biol. Biochem. 2021, 156, 108189. [Google Scholar] [CrossRef]
- Hall, S.J.; Silver, W.L. Reducing Conditions, Reactive Metals, and Their Interactions Can Explain Spatial Patterns of Surface Soil Carbon in a Humid Tropical Forest. Biogeochemistry 2015, 125, 149–165. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A.; Six, J.; Kaiser, M.; Benbi, D.; Chenu, C.; Cotrufo, M.F.; Derrien, D.; Gioacchini, P.; Grand, S.; et al. Isolating Organic Carbon Fractions with Varying Turnover Rates in Temperate Agricultural Soils—A Comprehensive Method Comparison. Soil Biol. Biochem. 2018, 125, 10–26. [Google Scholar] [CrossRef]
- Yu, W.; Huang, W.; Weintraub-Leff, S.R.; Hall, S.J. Where and Why Do Particulate Organic Matter (POM) and Mineral-Associated Organic Matter (MAOM) Differ among Diverse Soils? Soil Biol. Biochem. 2022, 172, 108756. [Google Scholar] [CrossRef]
- Sierra, M.; Martínez, F.J.; Verde, R.; Martín, F.J.; Macías, F. Soil-Carbon Sequestration and Soil-Carbon Fractions, Comparison between Poplar Plantations and Corn Crops in South-Eastern Spain. Soil Tillage Res. 2013, 130, 1–6. [Google Scholar] [CrossRef]
- Kraus, T.E.C.; Dahlgren, R.A.; Zasoski, R.J. Tannins in Nutrient Dynamics of Forest Ecosystems—A Review. Plant Soil 2003, 256, 41–66. [Google Scholar] [CrossRef]
- Tegelaar, E.W.; De Leeuw, J.W.; Holloway, P.J. Some Mechanisms of Flash Pyrolysis of Naturally Occurring Higher Plant Polyesters. J. Anal. Appl. Pyrolysis 1989, 15, 289–295. [Google Scholar] [CrossRef]
- Goering, H.K.; Soest, P.J.V. Forage Fiber Analyses (Apparatus, Reagents, Procedures, and Some Applications); U.S. Agricultural Research Service: Washington, DC, USA, 1970.
- Wasner, D.; Abramoff, R.; Griepentrog, M.; Venegas, E.Z.; Boeckx, P.; Doetterl, S. The role of climate, mineralogy and stable aggregates for soil organic carbon dynamics along a geoclimatic gradient. Glob. Biogeochem. Cycles 2024, 38, e2023GB007934. [Google Scholar] [CrossRef]
- Matus, F.; Salazar, O.; Aburto, F.; Zamorano, D.; Nájera, F.; Jovanović, R.; Guerra, C.; Reyes-Rojas, L.; Seguel, O.; Pfeiffer, M.; et al. Perspective of soil carbon sequestration in Chilean volcanic soils. npj Mater. Sustain. 2024, 2, 32. [Google Scholar] [CrossRef]
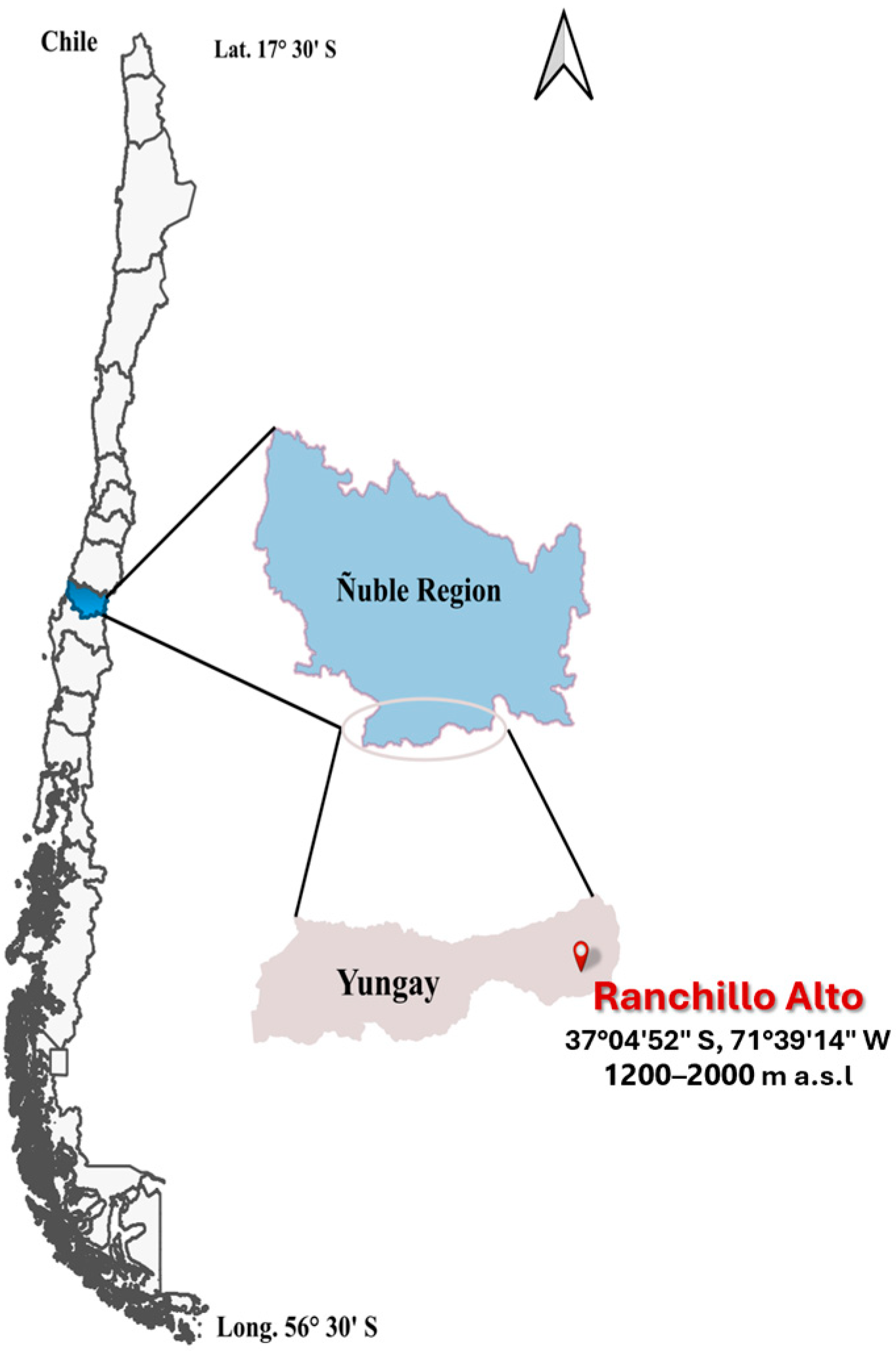


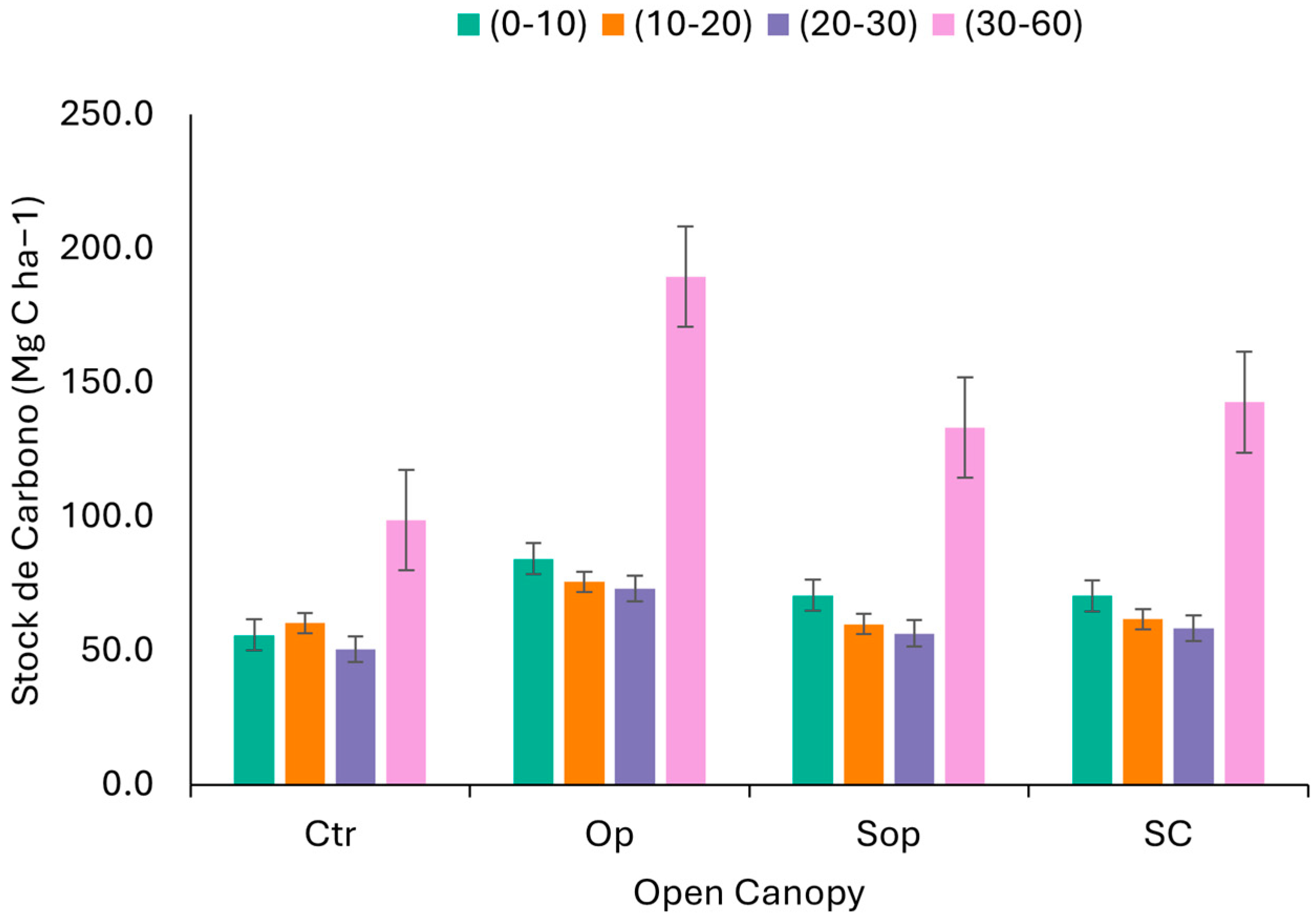
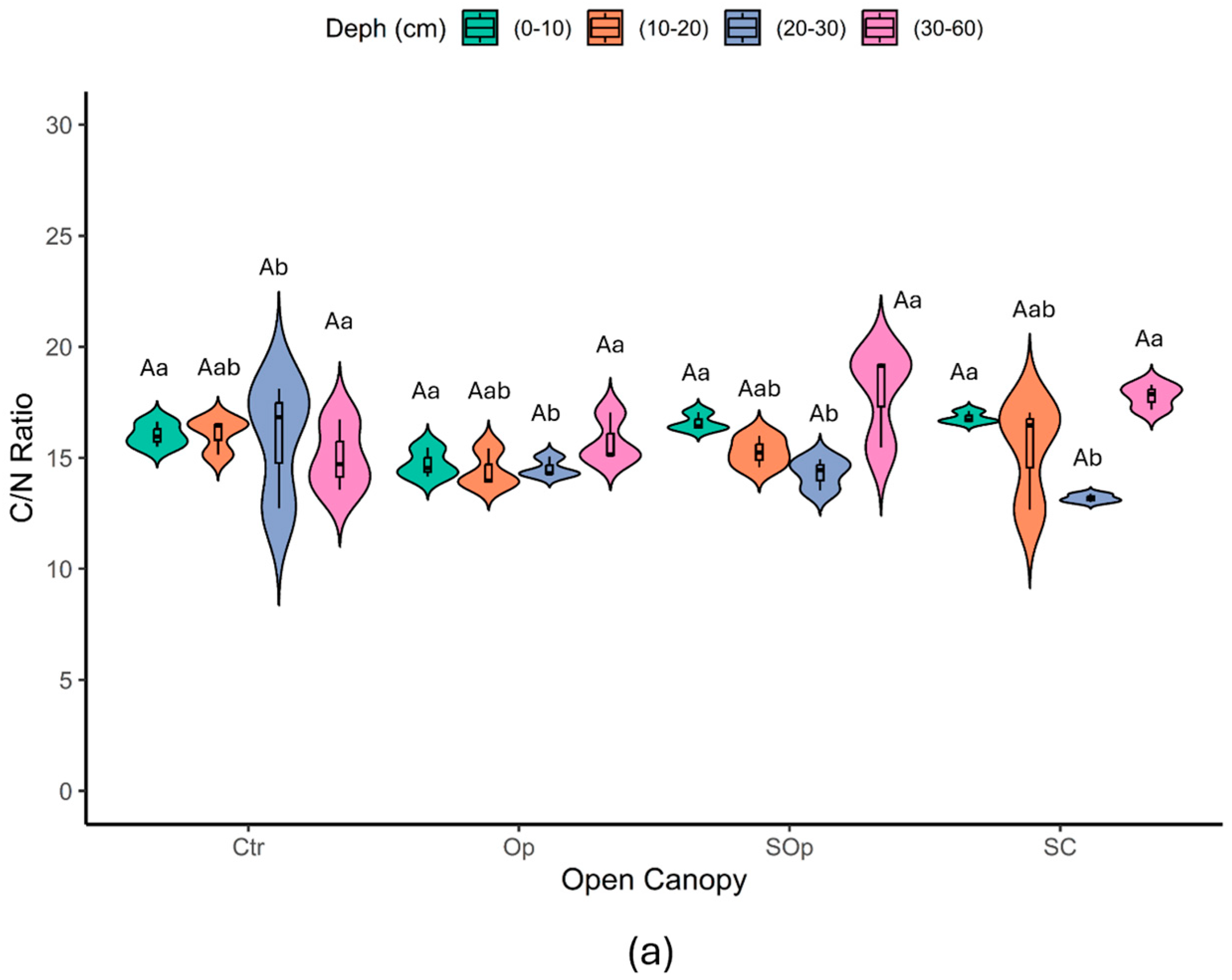

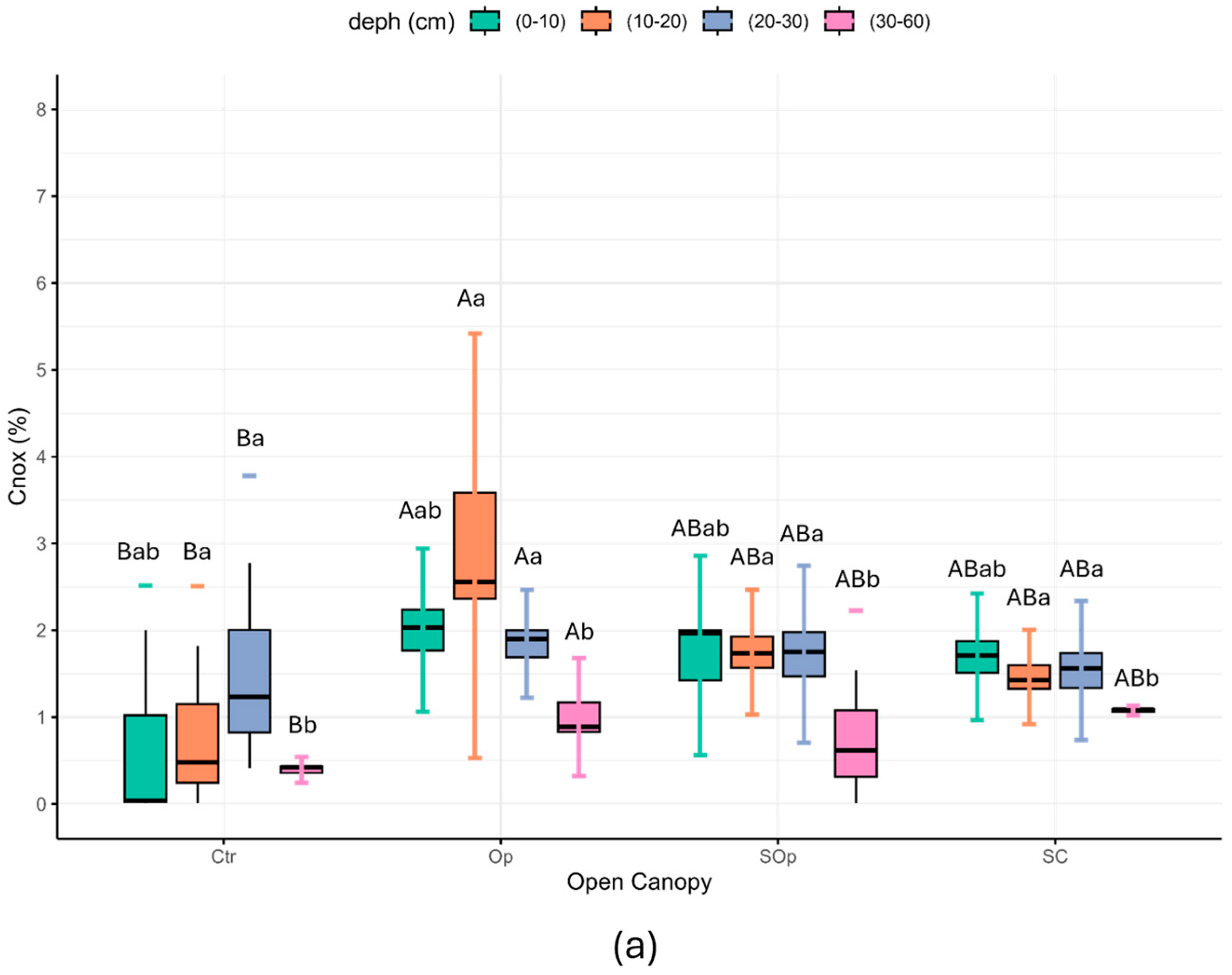
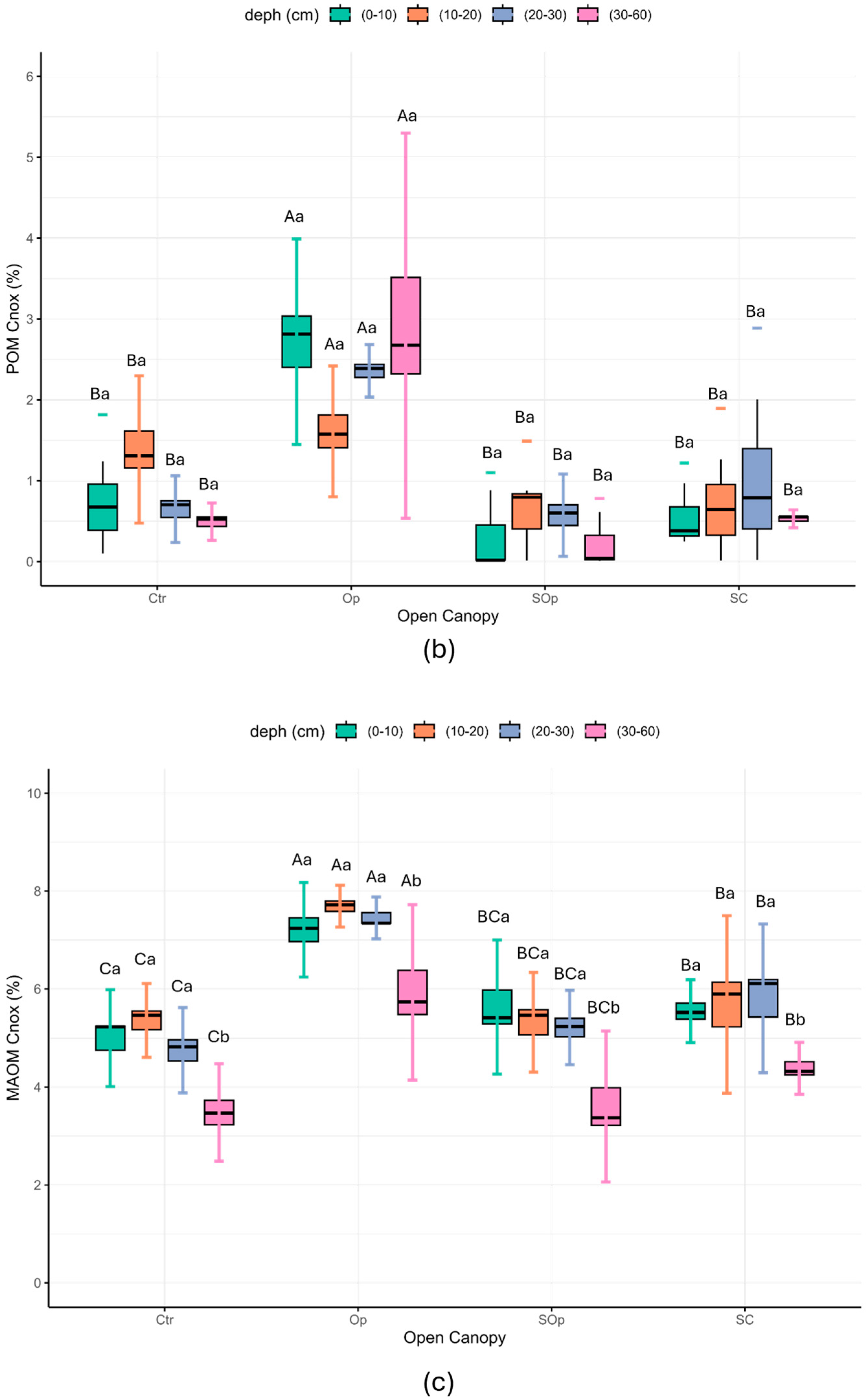
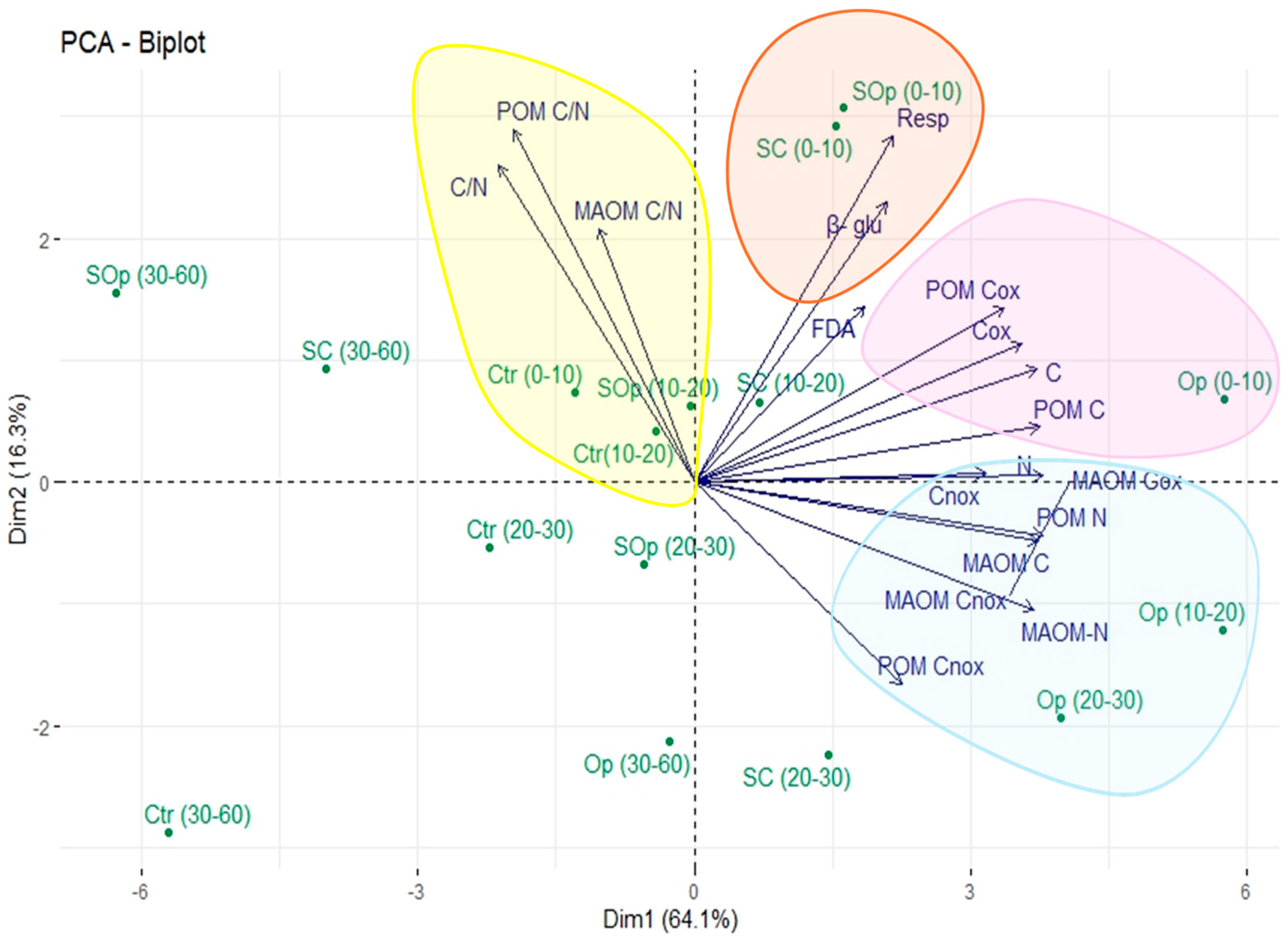
| Open Canopy | Location | NP/or Surface (ha) | Tree Density (ha−1) | Forest Species | Percentage of Light | Degradation | Soil Sampling |
|---|---|---|---|---|---|---|---|
| Ctr | 37°4′50″ S, 72°26′30″ W 1250 m.a.s.l | 3 × 1.33 | - | Roble (Nothofagus obliqua) | - | + | 4 depths (0–10, 10–20, 20–30 and 30–60 cm) |
| Op | 37°14′51″ S, 72°26′30″ O 1250 m.a.s.l | 3 × 1.33 | 60 | Roble (Nothofagus obliqua) | 85–95% | +++ | 4 depths (0–10, 10–20, 20–30 and 30–60 cm) |
| SOp | 37°14′50″ S, 72°26′30″ W 1250 m.a.s.l | 3 × 1.33 | 134 | Roble (Nothofagus obliqua) | 65–75% | ++ | 4 depths (0–10, 10–20, 20–30 and 30–60 cm) |
| SC | 37°14′49″ S, 72°26′30″ W 1250 m.a.s.l | 3 × 1.33 | 258 | Roble (Nothofagus obliqua) | 45–55% | + | 4 depths (0–10, 10–20, 20–30 and 30–60 cm) |
| Open Canopy/Depth | BD (g cm3) | PD (g cm3) | Sand (%) | Silt (%) | Clay (%) |
|---|---|---|---|---|---|
| Ctr (0–30) | 0.66 ± 0.02 Aa | 2.26 ± 0.00 Aa | 34.63 ± 1.10 Aa | 42.57 ± 0.40 Ab | 22.79 ± 0.77 Aa |
| Ctr (30–60) | 0.63 ± 0.02 Ab | 2.24 ± 0.01 Ab | 36.60 ± 4.09 Aa | 46.13 ± 3.77 Aa | 17.27 ± 0.40 Ab |
| Op (0–30) | 0.61 ± 0.02 Aa | 2.14 ± 0.02 ABb | 37.17 ± 0.37 Aa | 45.80 ± 0.37 Ab | 17.03 ± 0.68 Ba |
| Op (30–60) | 0.76 ± 0.08 Ab | 2.21 ± 0.00 ABa | 33.80 ± 2.36 Aa | 48.87 ± 0.37 Aa | 17.33 ± 1.41 Bb |
| SOp (0–30) | 0.61 ± 0.03 Aa | 2.09 ± 0.01 Bb | 40.57 ± 1.23 Aa | 43.90 ± 1.33 Ab | 15.50 ± 0.99 Ba |
| SOp (30–60) | 0.72 ± 0.06 Ab | 2.12 ± 0.09 Ba | 35.70 ± 1.04 Aa | 50.03 ± 1.59 Aa | 14.27 ± 0.37 Bb |
| SC (0–30) | 0.60 ± 0.01 Aa | 2.06 ± 0.01 Bb | 39.77 ± 3.01 Aa | 43.03 ± 0.82 Ab | 17.20 ± 2.12 ABa |
| SC (30–60) | 0.64 ± 0.06 Ab | 2.20 ± 0.05 Ba | 37.13 ± 2.53 Aa | 45.07 ± 1.17 Aa | 17.80 ± 2.47 ABb |
| Open Canopy /Depth /Depths | pH (Water) | OM 1 (%) | N 2 (mg kg−1) | P (mg kg−1) | K+ (mg kg−1) | ECEC (cmol kg−1) | AlEXCH * (cmol kg−1) | Ca2+EXCH * (cmol kg−1) | Mg2+EXCH * (cmol kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| Ctr (0–30) | 5.45 ± 0.05 Aa | 14.51 ± 1.56 Ca | 17.7 ± 0.30 Ba | 1.88 ± 17.76 Aa | 110.28 ± 0.04 Aa | 3.96 ± 1.52 Aa | 0.09 ± 0.01 ABa | 3.01 ± 1.34 Aa | 0.50 ± 0.14 Aa |
| Ctr (30–60) | 5.66 ± 0.04 Aa | 9.07 ± 1.44 Cb | 8.30 ± 3.18 Bb | 1.23 ± 8.03 Aa | 37.60 ± 0.02 Ab | 1.37 ± 0.72 Ab | 0.03 ± 0.01 Abb | 0.95 ± 0.59 Ab | 0.22 ± 0.10 Ab |
| Op (0–30) | 5.45 ± 0.05 Aa | 22.07 ± 0.52 Aa | 21.10 ± 0.05 ABa | 0.67 ± 0.05 Aa | 91.17 ± 0.05 Aa | 2.14 ± 0.05 Aa | 0.18 ± 0.05 Aa | 1.43 ± 0.05 Aa | 0.26 ± 0.05 Aa |
| Op (30–60) | 5.41 ± 0.04 Aa | 14.38 ± 0.27 Ab | 14.40 ± 0.04 ABb | 0.53 ± 0.04 Aa | 45.27 ± 0.04 Ab | 0.86 ± 0.04 Ab | 0.07 ± 0.04 Ab | 0.49 ± 0.04 Ab | 0.14 ± 0.04 Ab |
| SOp (0–30) | 5.62 ± 0.21 Aa | 17.49 ± 1.44 BCa | 20.57 ± 0.09 ABa | 0.73 ± 19.36 Aa | 62.57 ± 0.05 Aa | 2.36 ± 1.57 Aa | 0.07 ± 0.04 Ba | 1.83 ± 1.43 Aa | 0.27 ± 0.12 Aa |
| SOp (30–60) | 5.63 ± 0.08 Aa | 10.52 ± 1.73 BCb | 10.83 ± 0.09 ABb | 0.67 ± 22.66 Aa | 46.60 ± 0.06 Ab | 1.20 ± 0.56 Ab | 0.01 ± 0.00 Bb | 0.83 ± 0.48 Ab | 0.16 ± 0.02 Ab |
| SC (0–30) | 5.50 ± 0.18 Aa | 18.37 ± 1.48 Ba | 26.97 ± 0.14 Aa | 0.90 ± 21.32 Aa | 90.50 ± 0.05 Aa | 3.43 ± 1.52 Aa | 0.14 ± 0.09 ABa | 2.62 ± 1.43 Aa | 0.39 ± 0.13 Aa |
| SC (30–60) | 5.44 ± 0.12 Aa | 12.67 ± 1.39 Bb | 17.30 ± 0.05 Ab | 0.57 ± 38.09 Aa | 81.87 ± 0.06 Ab | 1.92 ± 0.84 Ab | 0.09 ± 0.05 ABb | 1.29 ± 0.69 Ab | 0.25 ± 0.08 Ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, C.; Zagal, E.; Timmusk, S.; Dube, F.; Paulino, L.; Ortiz, J.; Intriago-Ávila, J.; Fuentes, J.P. Soil Carbon Sequestration in Nothofagus obliqua Forests with Different Canopy Cover Levels Under Silvopastoral Management. Agronomy 2025, 15, 855. https://doi.org/10.3390/agronomy15040855
Ramos C, Zagal E, Timmusk S, Dube F, Paulino L, Ortiz J, Intriago-Ávila J, Fuentes JP. Soil Carbon Sequestration in Nothofagus obliqua Forests with Different Canopy Cover Levels Under Silvopastoral Management. Agronomy. 2025; 15(4):855. https://doi.org/10.3390/agronomy15040855
Chicago/Turabian StyleRamos, Camila, Erick Zagal, Salme Timmusk, Francis Dube, Leandro Paulino, Juan Ortiz, Jean Intriago-Ávila, and Juan Pablo Fuentes. 2025. "Soil Carbon Sequestration in Nothofagus obliqua Forests with Different Canopy Cover Levels Under Silvopastoral Management" Agronomy 15, no. 4: 855. https://doi.org/10.3390/agronomy15040855
APA StyleRamos, C., Zagal, E., Timmusk, S., Dube, F., Paulino, L., Ortiz, J., Intriago-Ávila, J., & Fuentes, J. P. (2025). Soil Carbon Sequestration in Nothofagus obliqua Forests with Different Canopy Cover Levels Under Silvopastoral Management. Agronomy, 15(4), 855. https://doi.org/10.3390/agronomy15040855