1. Introduction
Currently, an estimated 1 billion hectares (30%) of arable soils are affected worldwide by salt stress, resulting in crop losses of USD 27 billion annually [
1,
2]. In the Northern Great Plains (NGP) of North America, marine sediments buried below glaciated soils are a source of salts that cause saline seeps due to rising water tables [
3]. In South Dakota, salinity and sodicity lead to approximately USD 26.2 million in annual economic losses across over 45,870 hectares in the northeastern and central counties of the state.
Maize (
Zea mays L.) (47%), soybean (
Glycine max L.) (41%), and wheat (
Triticum aestivum) (11%) are the most common crops in eastern South Dakota [
3]. Maize yield losses start when saturated paste electrical conductivity (EC
e) values are about 1.7 dS m
−1, with additional 12% yield losses for each 1 dS m
−1 above that value [
4]. Currently, around 14% of corn farmers are witnessing some yield detriment due to ECe levels exceeding this value, with indications that soil ECs are on the rise [
5]. Soybean and wheat have higher EC
e threshold values of 5.0 and 6.0, respectively, [
4], but even these crops are at risk for yield losses due to the presence of a subsurface natric soil horizon that continues to supply salts to the surface. The elevated osmotic potential associated with saline–sodic soils restricts water availability to plants, thereby hindering growth [
6,
7]. Additionally, the high sodium concentration contributes to soil dispersion and sealing while increasing the likelihood of flooding and water erosion risks. Salts can also be transported by water to previously unaffected regions, leading to an expansion of saline–sodic zones, which in turn elevates soil EC levels and diminishes future soil productivity, even far from the initial source of the issue [
8].
Traditional remediation/restoration strategies for NGP saline–sodic soils, such as leaching of soil salts using non-saline water, chemical amendments such as gypsum and elemental sulfur application, and mechanical methods such as installing tile drains and deep tillage (ripping) to improve water infiltration, have been unsuccessful [
5,
8,
9] in either reducing the affected areas or lowering the EC. However, establishing salt-tolerant plants in saline–sodic areas may help restore soil functionality and health to near-barren areas [
3,
10,
11,
12], slow the expansion of saline seeps, and provide income to the farmers [
5,
13,
14]. Previous studies on plant tolerance to saline–sodic soils in South Dakota dryland areas have evaluated native plants [
15] and perennial grasses [
3,
10,
16], with less attention given to annual crops that have significantly greater economic importance.
Brassica spp. like canola (
B. napus), mustard (
B. juncea), and carinata (
B. carinata) have shown promise in studies as viable options for salt-tolerant annual crops [
13,
17,
18]. Some canola genotypes and other
Brassica spp. have been reported to have an EC
e threshold of 9.7 [
4,
19], much higher than the rotational crops now commonly grown in these areas.
Brassica spp. could be used as either cash (canola and carinata) or cover (brown mustard or Florida mustard) crops, although evaluation is needed since tolerance to higher EC
e values varies by species and genotypes. For example, amphidiploid species of
Brassica, such as canola and Florida broadleaf mustard (
B. juncea), are relatively more salt tolerant compared to
Brassica diploid species [
20,
21]. An added advantage of
Brassica species is that they have glucosinolate compounds with antimicrobial activity in the leaves and, therefore, have suppressive effects on major soilborne pathogens that cause root rot, such as
Pythium and
Rhizoctonia spp. [
22]. While microbial suppression may be desirable in some soil types, it may be undesirable in saline–sodic soils that tend to have low microbial populations [
12], with further suppression being undesirable for long-term ecosystem function restoration [
23].
The often-suggested chemical amendments of gypsum and elemental sulfur application have had minimal impact on soil chemical and physical properties of NGP saline–sodic soils [
5,
8]. Soils of South Dakota are often saturated with calcite and have high sulfate levels (mg SO
4−2 kg
−1), making the amendments valueless. However, biochar has attracted considerable attention as a soil amendment to remediate and improve the physical, chemical, and biological properties of degraded soil [
24] and promote plant growth [
25]. Despite the potential of biochar as an amendment for saline–sodic soils, it has not been evaluated on South Dakota saline–sodic soils.
Several types and sources of biochar are available, with pyrolysis temperature, time, and pressure, as well as source of feedstock, impacting the final product [
26,
27,
28,
29]. For example, hardwood feedstock pyrolyzed at high temperatures produces high carbon char with higher amounts of aromatic structures and greater cation exchange capacity, whereas soft and non-wood feedstocks, such as crop residues, manures, and straw biomass, produce biochar with lower carbon content [
28,
30], more aliphatic compounds, and lower sorption capacity [
31]. Biochar produced from pinewood chips at 500 °C under slow pyrolysis had 817 g kg
−1 C content, whereas poultry manure char pyrolyzed at the same temperatures had about half the carbon content (only 400 g kg
−1 C) [
28]. Biochar produced from herbaceous feedstock also tends to contain lower proportions of mesopores and macropores and exhibits a smaller surface area compared with biochar pyrolyzed from woody biomass, which also influences its activity when added to the soil [
29,
32].
A review of the impact of soil biochar amendment on crop yields [
24] indicated mixed results, with yields ranging from −28 to +39% compared to non-amended soils, although there was an overall average yield increase of 10%. Some studies reported negative effects of biochar on soil organic carbon, nutrient mineralization and uptake, and soil microbial activity [
26,
32], whereas others showed negative effects on seed germination and plant growth due to the release of volatile organic compounds [
33]. However, biochar has been reported to reduce salinity impacts on soil EC
e [
34], improving soil water infiltration [
35] and sorbing sodium and sulfate, which can be present in very high amounts in NGP saline–sodic soils [
3]. These changes in soil properties may improve plant emergence, growth, and development.
Due to the variation in Brassica spp. salt and ECe tolerances and differences in biochar performance in past studies, it is worthwhile to screen genotypes and chars produced from various wood sources under controlled conditions to determine if positive results for seedling emergence and plant growth could be obtained prior to large-scale field experiments. Therefore, the objectives of these greenhouse studies were to (i) evaluate the emergence of Brassica spp. (canola and mustard genotypes) in a South Dakota saline–sodic soil and (ii) evaluate emergence and shoot biomass of canola and mustard genotypes in the saline–sodic soil amended with two biochar types (softwood and hardwood).
4. Discussion
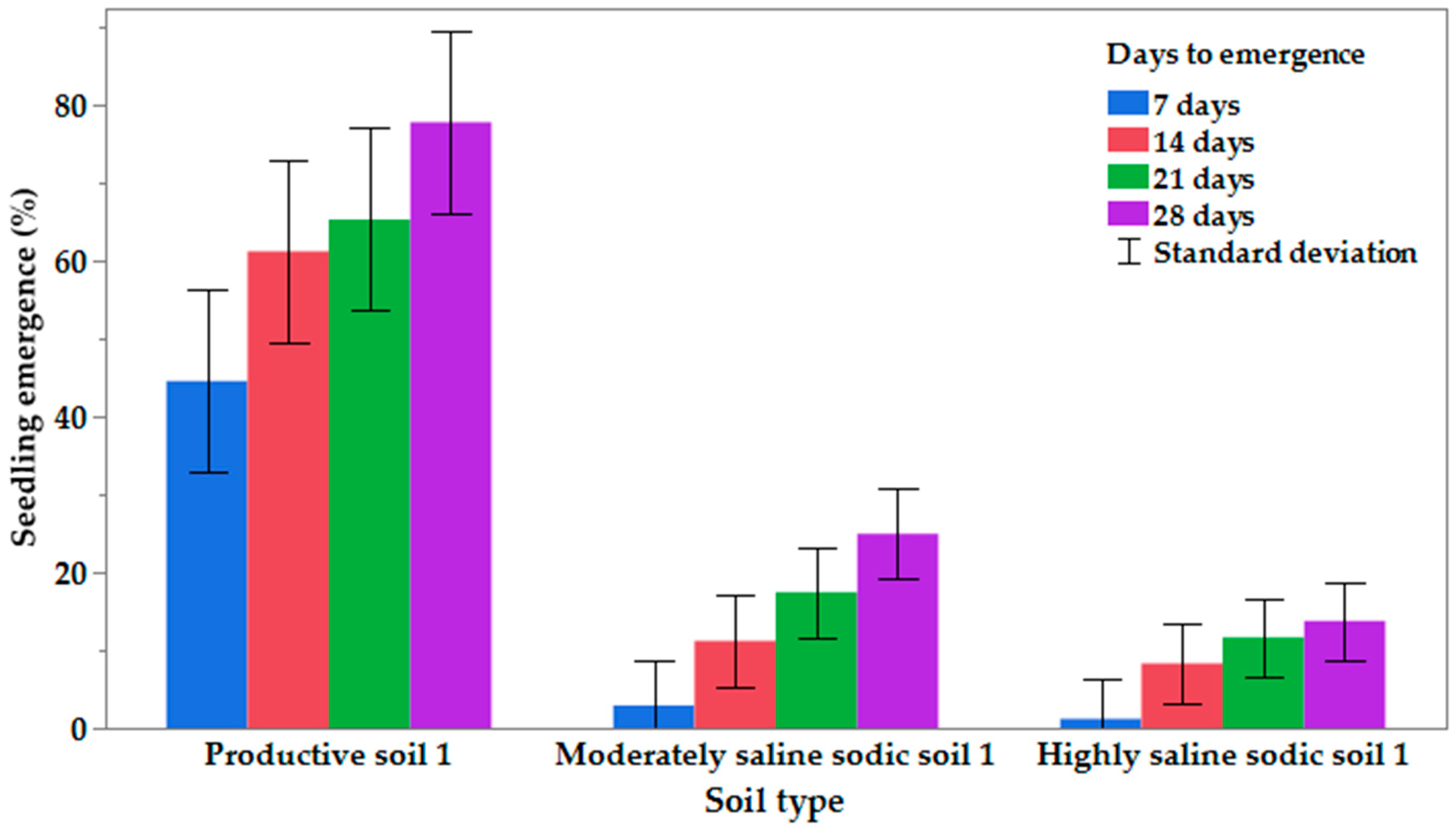
Although saline–sodic soils can exhibit high water content [
3], the elevated salt concentration, particularly sodium ions in the seed–soil contact zone, not only draws water away from the seed, thereby reducing the moisture available for hydrolyzing seed endosperm contents [
53,
54] but also directly disrupts critical physiological processes, such as enzyme activity and ion homeostasis, that are essential for germination, resulting in seedling desiccation due to reverse osmosis [
55]. Similarly, under greenhouse conditions, high salt content in the soil lowers osmotic potential, reducing plant available water for seed germination and seedling emergence [
55], and, as observed in the current study, seedling emergence was reduced. Even though we observed that watering the plants flushed some of the salts out of the pots, as observed by white crusting at the base of the pot, the amount of salt remaining in the soil was high enough to impact seedling emergence. The results in the current study are consistent with other studies using canola [
53,
56,
57,
58], where negative associations between seedling emergence and soil salt content were observed.
All
Brassica species used in this study are amphidiploids and have been reported to be relatively more salt tolerant compared to diploid species [
17,
21]. However, salt tolerance threshold levels within amphidiploids vary [
21], with, for example, canola (
B. napus) reported as more salt tolerant than Brown mustard (
B. juncea). The genetic variation, therefore, explains part of the variation in salt tolerance among
Brassica species used in the current study. This assertion is also consistent with Francois [
56], who reported the threshold for growth in saline soil to be an EC
e of 10 dS/m
−1 for most canola genotypes. In the current study, on average, only 36% of the seeds emerged at an EC
1:1 of 5.16 dS m
−1. However, as EC
1:1 increased, the percentage of seedlings that emerged declined. It must be remembered that the EC
e value is determined on a saturated paste, whereas the EC
1:1 is quantified on a soil slurry. To convert the EC
1:1 to the EC
e, the EC
1:1 value needs to be multiplied by 1.8 to 2.1, depending on soil type [
59,
60]. This means that if the soils used in this study had been analyzed by the saturated paste method, which is expensive, labor-intensive, and not conducted in commercial labs, the EC
e of our saline–sodic soil would be about 12 dS m
−1 [
59] and pH 6.9, which is then comparable to the EC
e threshold of 10 dS m
−1, reported by Francois [
56].
When we separated the entries by species,
B. napus emergence averaged 35.1% (ranging from 20.2% to 48.2% for 10 entries). Emergence for the two
B. juncea species averaged 42.2%, ranging from 33.2% (Brown mustard) to 52.2% (Florida broadleaf mustard). The interaction of canola genotype/
Brassica spp. with soil salinity indicates that although emergence was not 100% under saline–sodic conditions, carefully chosen genotypes/species may be useful to provide stability from roots and vegetative shoots to the saline–sodic areas. For example, Green Cover (
https://store.greencover.com/ accessed 18 March 2025) categorizes African cabbage and Florida broadleaf mustard tolerance to saline soils as very good and fair, respectively, and recommends them for cover cropping in areas impacted by salinity.
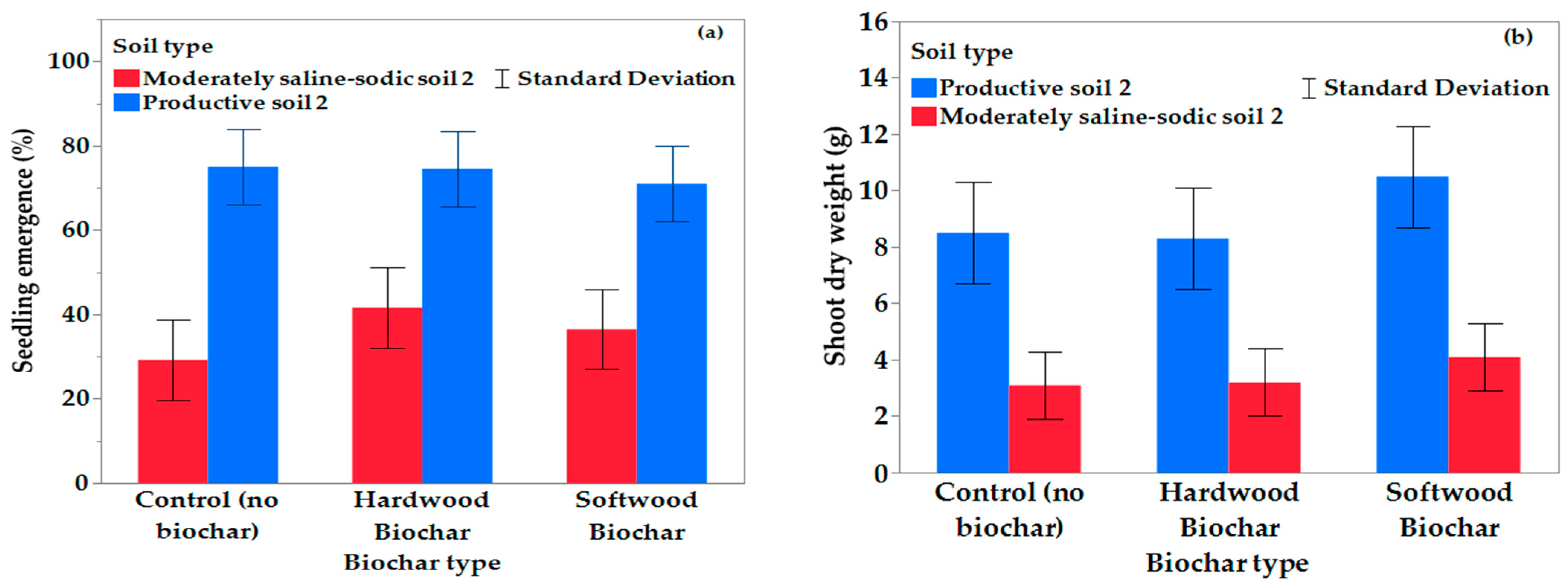
Biochar application enhanced seedling emergence in the current study. This is likely due to biochar sorbing sodium from the saline soils [
25,
26] or due to its capacity to increase plant available water (lowering ionic content) to reduce the salinity stress [
61,
62]. In this specific study, we did not analyze biochar-treated soil to determine changes in soil properties. However, prior studies have shown a reduction in the soil EC and Na+ content of saline soil with the addition of biochar [
63] in addition to increasing plant available moisture [
62,
64].
Although softwood biochar had over 20-fold the surface area of the hardwood biochar (
Table 3), the hardwood biochar had the greatest impact on emergence in the saline–sodic soil. The small surface area of the hardwood biochar likely improved plant available water and nutrients, also reducing osmotic stress, resulting in better conditions for seedling emergence [
65,
66]. Additionally, the hardwood had a smaller particle size, which likely increased porosity, improving drainage and reducing surface crusting compared to softwood biochar, explaining the improved seedling emergence [
66]. In a comparative study, three application rates of hardwood and softwood biochars were evaluated for their effects on plant growth, maize yield and moisture availability under different irrigation regimes. The study showed that there were substantial differences between hardwood and softwood biochar and that hardwood biochar reduced soil bulk density and increased soil porosity, but it had a marginal effect on the water retention characteristics. Other studies have observed large differences in the chemistry of softwood and hardwood biochars, which influence their impact on soil physical, chemical, and biological properties [
46,
67,
68]. These differences explain the variations observed in the current study results.
Microbial biomass and diversity are important in maintaining soil health and ecosystem sustainability [
12,
69]. The relative proportion of soil bacterial and fungal biomass is an indicator of how long it will take to rebuild the soil structure [
69,
70] because fungi and bacteria release exudates that are important in the formation and strengthening of soil aggregates [
69,
71,
72]. The low microbial biomass in the saline–sodic soil types used in this study indicates that carbon and nutrients will have lower turnover rates, and these soils would provide no buffering to plants for abiotic and biotic stresses [
23]. This highlights the importance of the current study and others aimed at identifying plant species for remediating these salt-impacted soils, as the presence of plants increases the fungi-to-bacteria ratio (from 0.063 to 0.094) [
3] and improves the overall biological health, suggesting that vegetative remediation will improve soil microbial diversity and ultimately rebuild the soil structure of these degraded soils.





{kind=link}
{kind=link}