

Self-Organizing Map-Based Assessment of Compost Maturity and Cu/Zn Passivation in Biochar-Amended Pig Manure


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Acquisition of Compost Raw Substrate
2.2. Experimental Design and Sampling Procedure
2.3. Analysis of Composting Properties and Heavy Metals
2.4. Fluorescence Spectrometry and EEM-PARAFAC Analysis
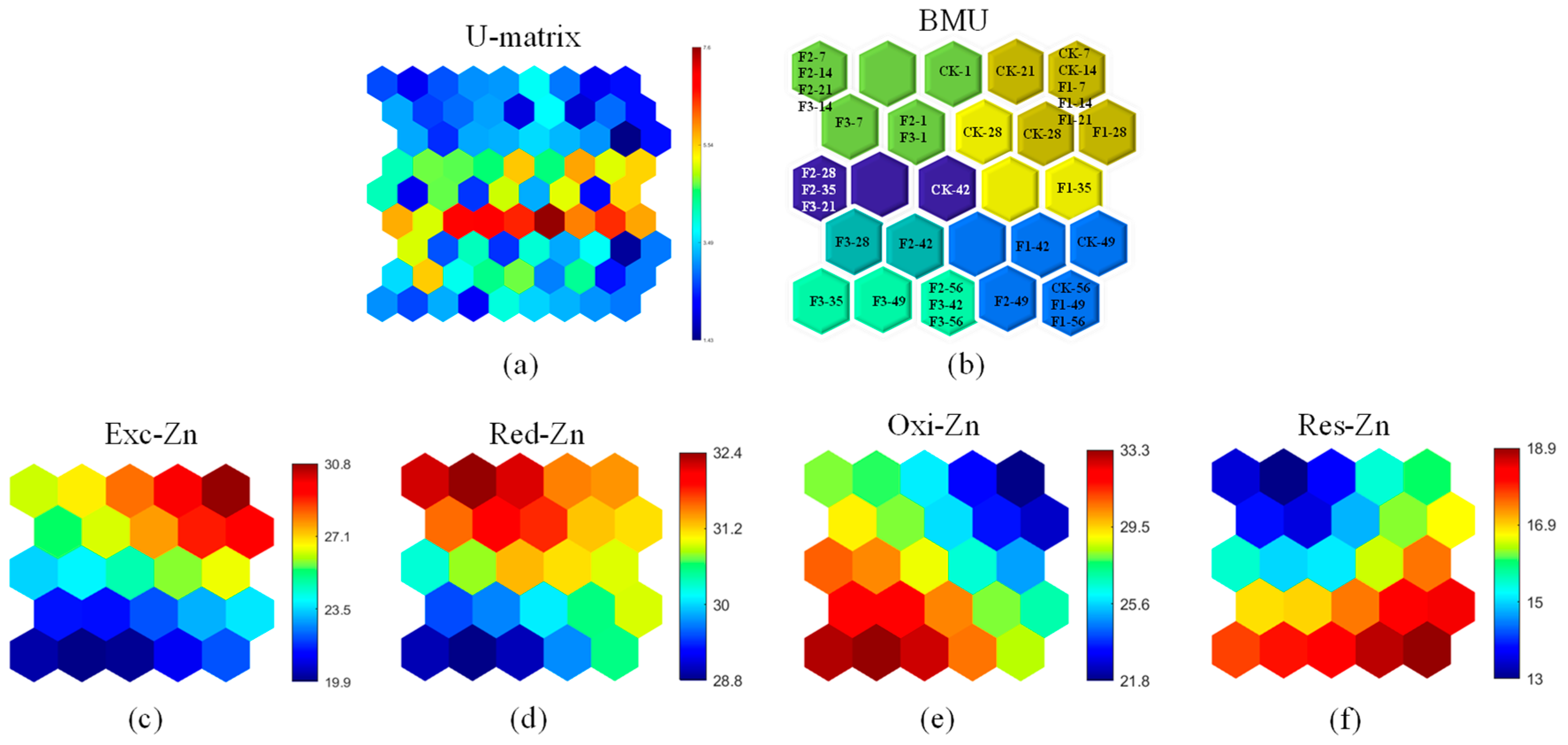
2.5. Integration of Self-Organizing Maps with PARAFAC
2.6. Microbial Analysis
2.7. Data Analysis and Software Tools
3. Results and Discussion
3.1. Physicochemical Properties
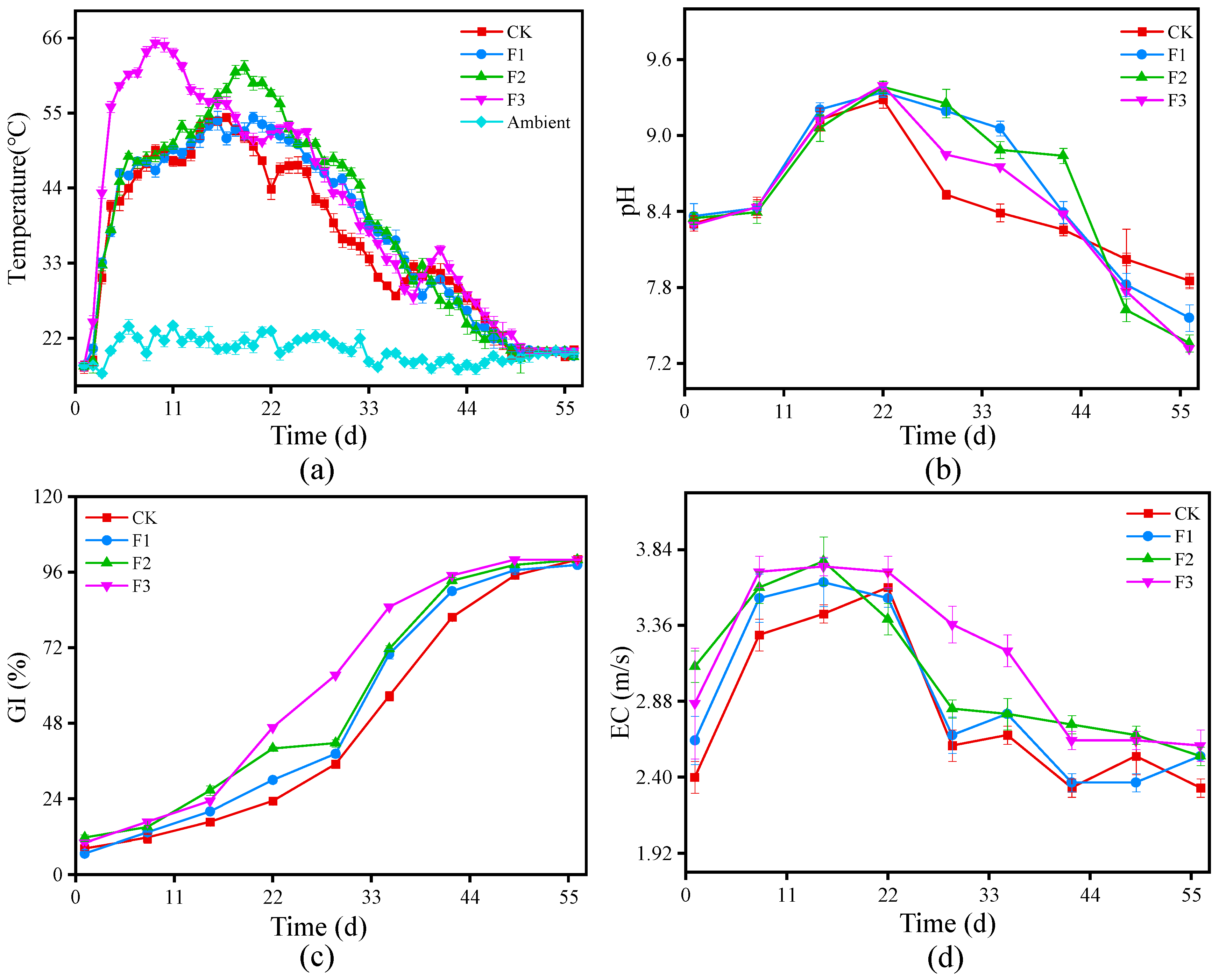
3.1.1. Temperature Dynamics
3.1.2. Changes in pH
3.1.3. Germination Index Variations
3.1.4. Electrical Conductivity Variations
3.2. Spectral Analysis of Dissolved Organic Matter
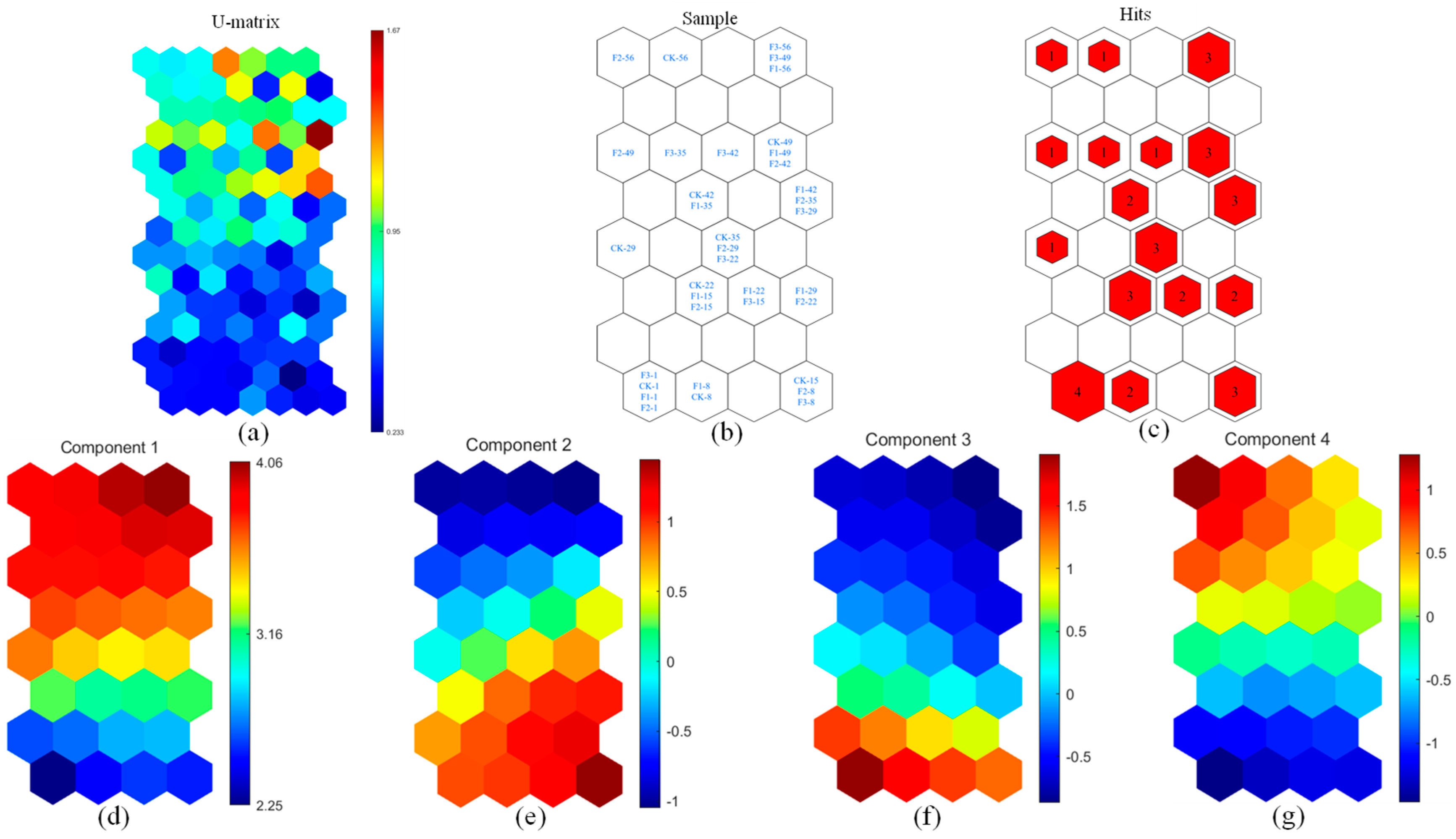
3.2.1. Identification of Fluorescent Components
3.2.2. Analysis of Fluorescent Components
3.3. Biochar-Induced Cu and Zn Passivation
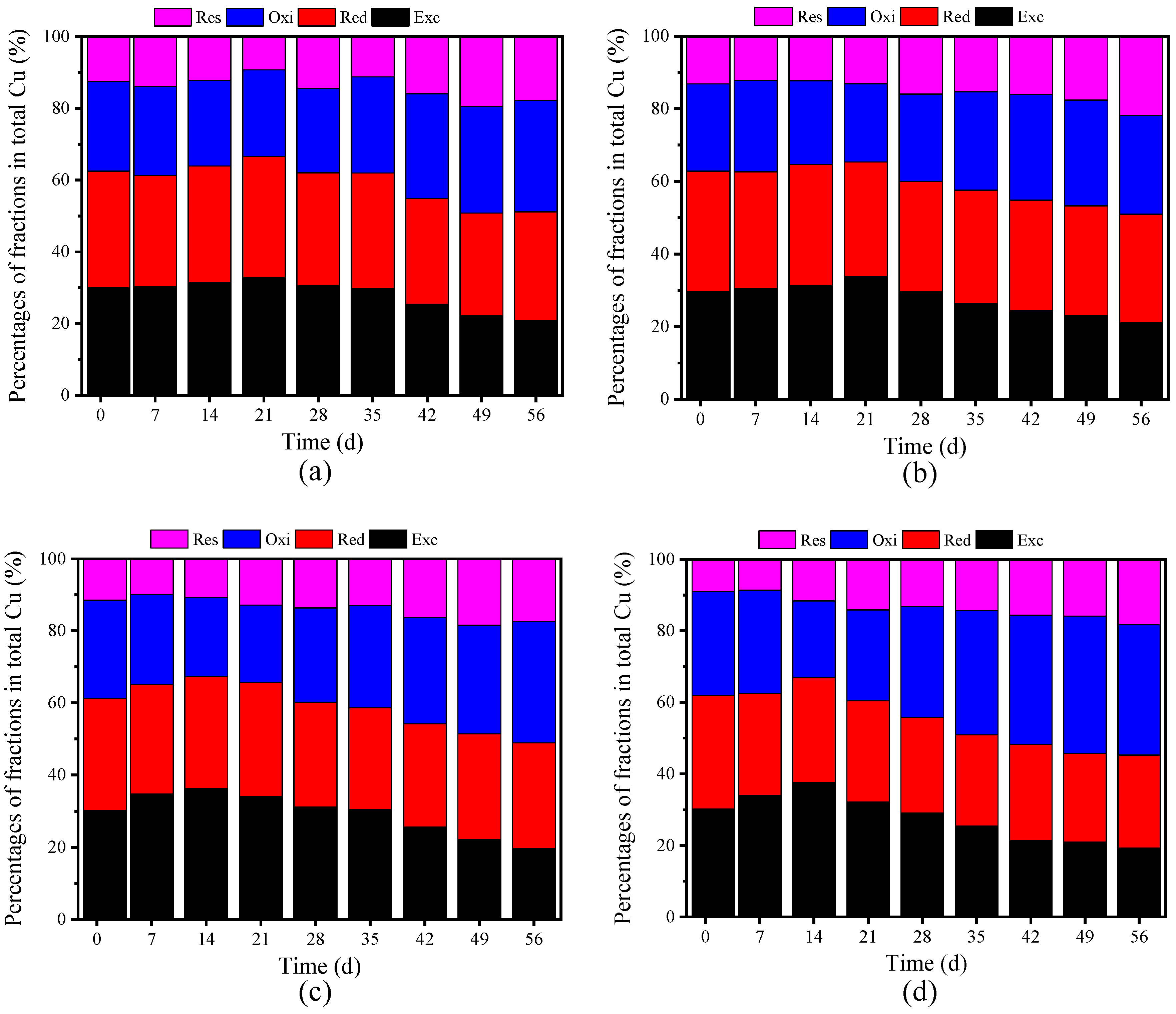
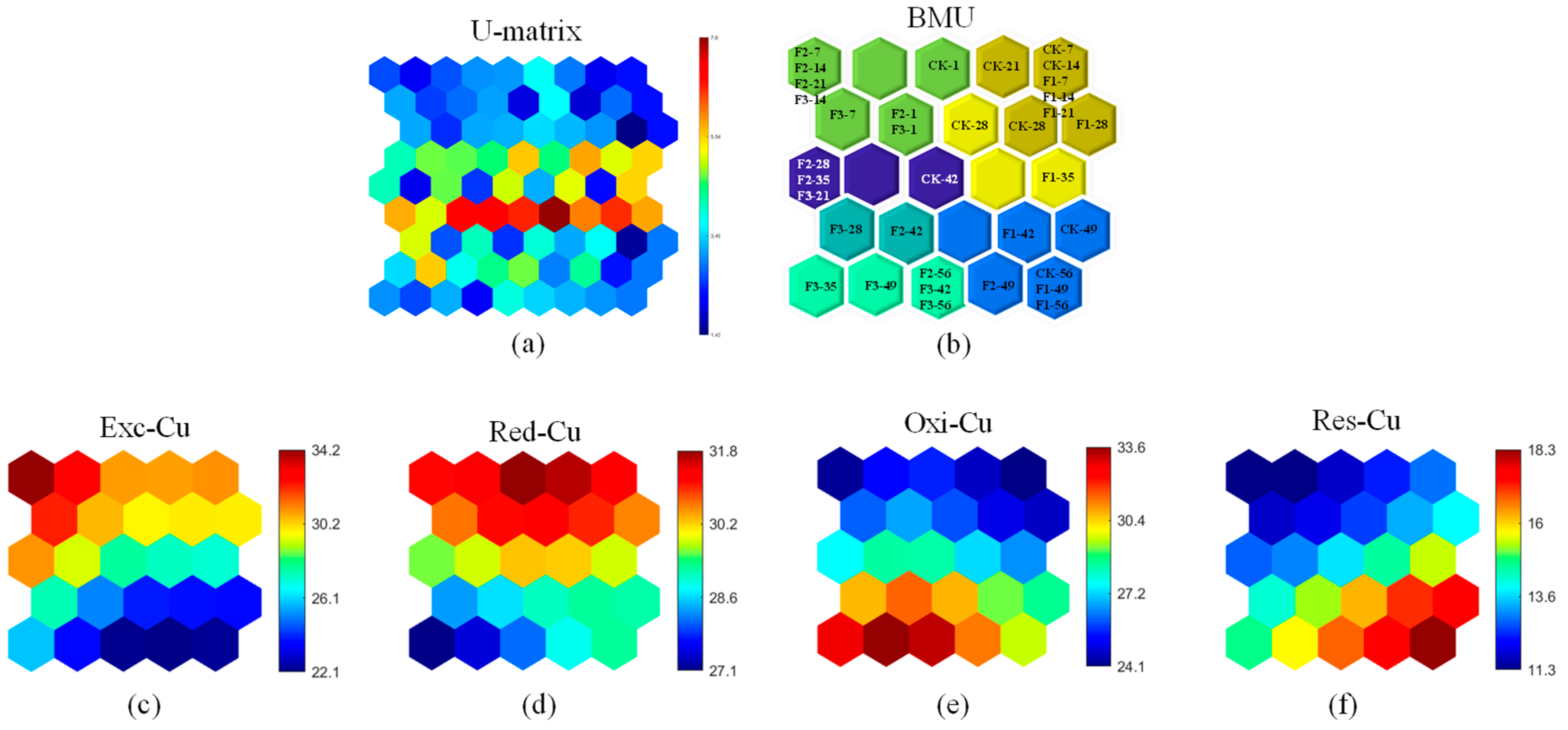
3.3.1. Passivation of Cu
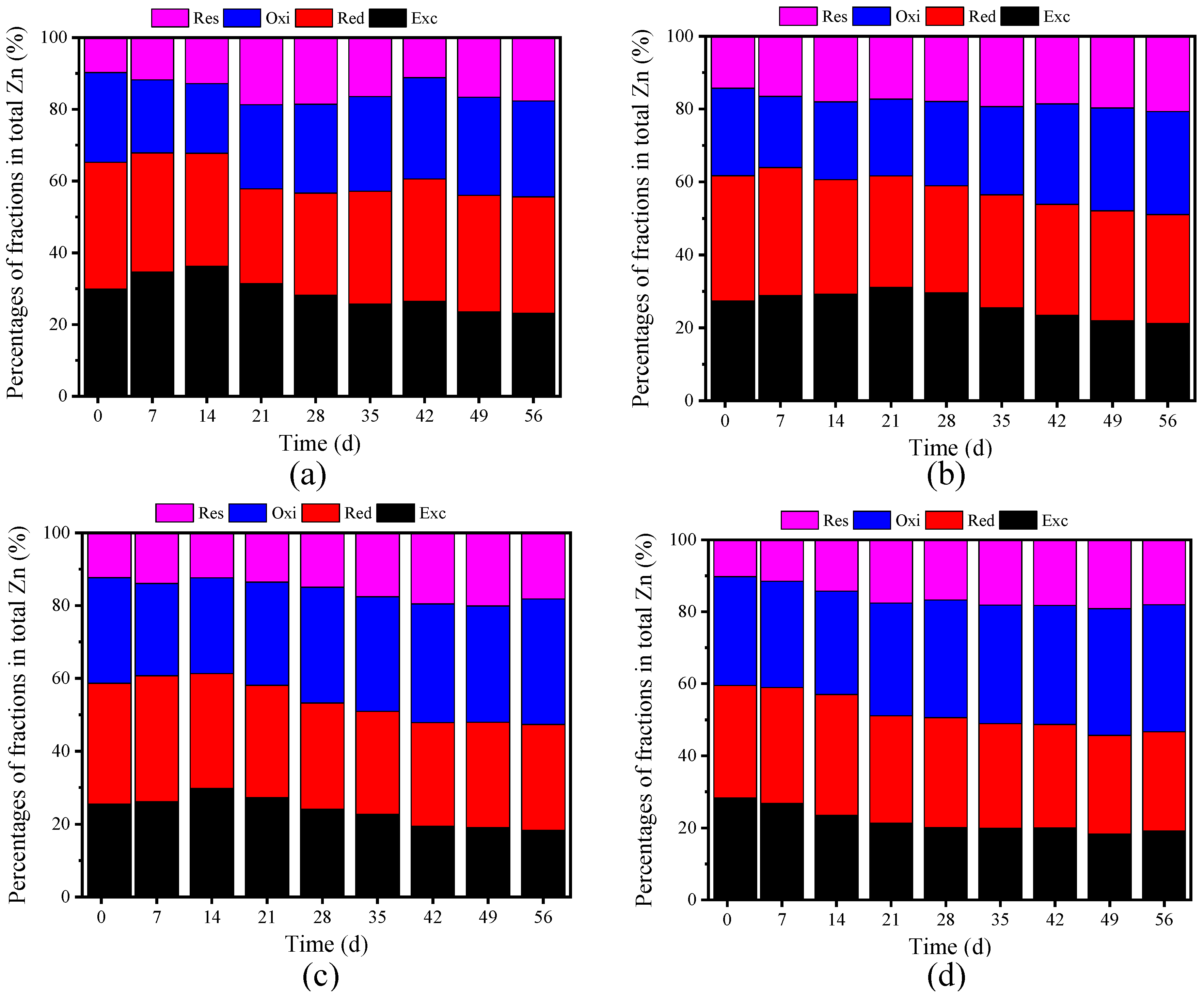
3.3.2. Passivation of Zn
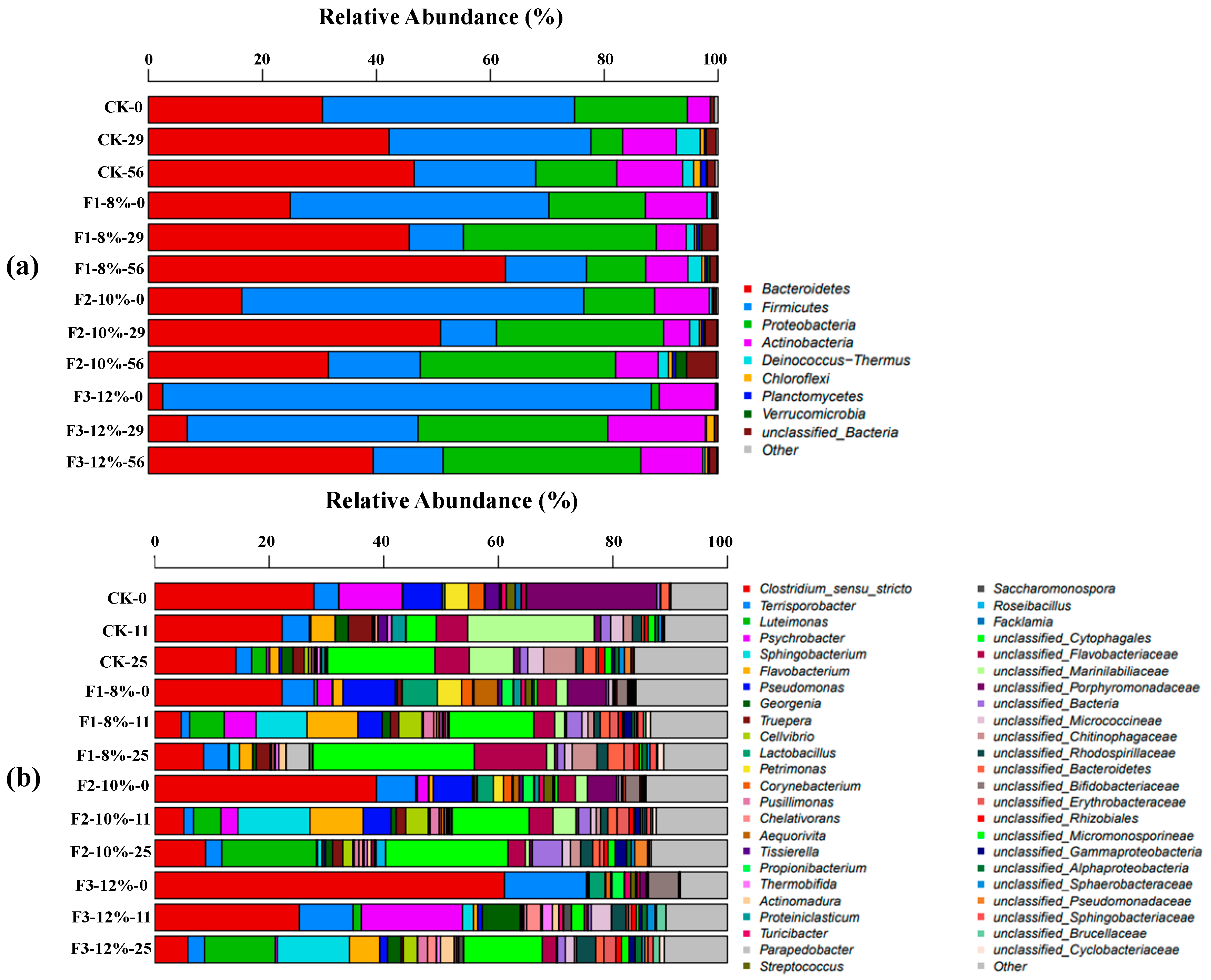
3.4. Microbial Composition Analysis
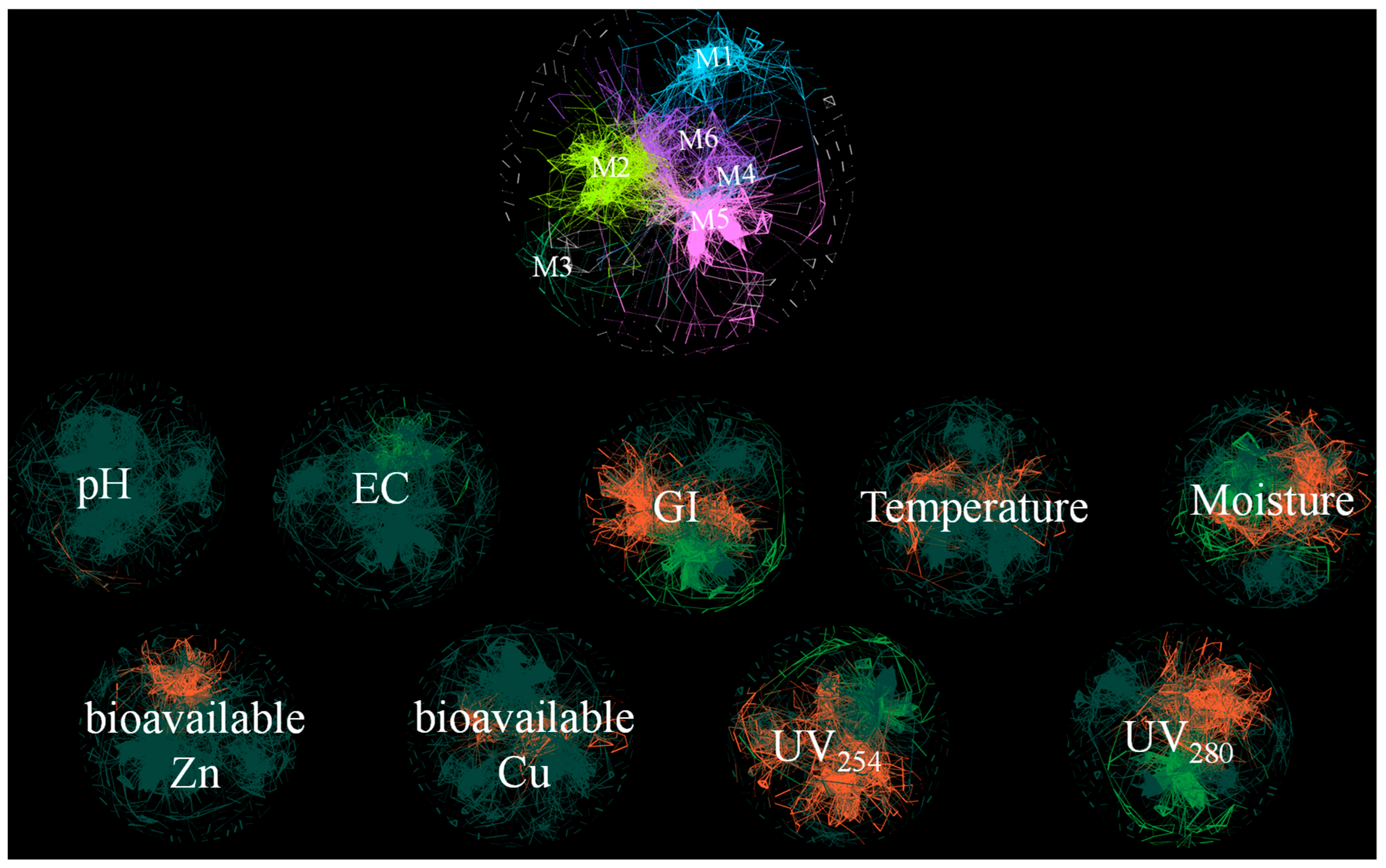
3.5. Bacterial Community Network Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhou, L.; Xue, J.; Xu, Y.; Tian, W.X.; Huang, G.W.; Liu, L.Q.; Zhang, Y.C. Effect of biochar addition on copper and zinc passivation pathways mediated by humification and microbial community evolution during pig manure composting. Bioresour. Technol. 2023, 370, 128575. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, L.X.; Wang, X.H.; Ou, Y.; Liu, H.P.; Hou, X.; Yan, L.M.; Li, X.Y. The various effect of cow manure compost on the degradation of imazethapyr in different soil types. Chemosphere 2023, 337, 139325. [Google Scholar] [CrossRef]
- Yang, M.; He, L.L.; Rao, Z.X.; Huang, D.Y.; Tian, Y.B.; Zhu, H.H.; Xu, C.; Zhang, Q. Effects of long-term fertilization on accumulation and availability of Cd, Zn and Cu in paddy soil. Res. Agric. Mod. 2021, 42, 302–310. [Google Scholar] [CrossRef]
- Li, X.X.; Wei, Y.F.; Wang, L.J.; Jin, S.J.; Wang, P.; Chang, J.; Yin, Q.Q.; Liu, C.Q.; Li, M.L.; Liu, Y.Y.; et al. Effects of multi-component passivator on heavy metal passivation, compost quality andplant growth. Resour. Environ. Sustain. 2024, 17, 100166. [Google Scholar] [CrossRef]
- Zheng, X.R.; Wu, K.H.; Sun, P.J.; Zhouyang, S.Y.; Wang, Y.P.; Wang, H.T.; Zheng, Y.M.; Li, Q.B. Effects of substrate types on the transformation of heavy metal speciation and bioavailability in an anaerobic digestion system. J. Environ. Sci. 2021, 101, 361–372. [Google Scholar] [CrossRef]
- Shen, Y.J.; Zhao, L.X.; Meng, H.B.; Hou, Y.Q.; Zhou, H.B.; Wang, F.; Cheng, H.S.; Liu, H.B. Effect of aeration rate, moisture content and composting period on availability of copper and lead during pig manure composting. Waste Manag. Res. 2016, 34, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.L.; Li, J.L.; Lin, L.; Qin, W.K.; Gao, Y.C.; Hu, E.D.; Jiang, J.G. Occurrence, fate and control strategies of heavy metals and antibiotics in livestock manure compost land application: A review. Sci. Total Environ. 2024, 957, 177381. [Google Scholar] [CrossRef] [PubMed]
- Song, H.R.; Chen, S.F.; Si, G.L.; Bhatt, K.; Chen, S.H.; Chen, W.J. Removal of environmental pollutants using biochar: Current status and emerging opportunities. Environ. Geochem. Health 2024, 46, 384. [Google Scholar] [CrossRef]
- Li, Y.; Awasthi, M.K.; Sindhu, R.; Binod, P.; Zhang, Z.Q.; Taherzadeh, M.J. Biochar preparation and evaluation of its effect in composting mechanism: A review. Bioresour. Technol. 2023, 384, 129329. [Google Scholar] [CrossRef]
- Duan, Y.M.; Yang, J.F.; Guo, Y.R.; Wu, X.P.; Tian, Y.L.; Li, H.K.; Awasthi, M.K. Pollution control in biochar-driven clean composting: Emphasize on heavy metal passivation and gaseous emissions mitigation. J. Hazard. Mater. 2021, 420, 126635. [Google Scholar] [CrossRef]
- Zhou, Y.W.; Awasthi, S.K.; Liu, T.; Verma, S.; Zhang, Z.Q.; Pandey, A.; Varjani, S.; Li, R.H.; Taherzadeh, M.J.; Awasthi, M.K. Patterns of heavy metal resistant bacterial community succession influenced by biochar amendment during poultry manure composting. J. Hazard. Mater. 2021, 420, 126562. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Ou, Y.; Wang, L.X.; Yan, B.X.; Li, Y.X.; Ding, D.W. The passivation effect of heavy metals during biochar-amended composting: Emphasize on bacterial communities. Waste Manag. 2020, 118, 360–368. [Google Scholar] [CrossRef]
- Kong, Y.L.; Zhang, J.; Zhang, X.S.; Gao, X.; Yin, J.; Wang, G.Y.; Li, J.M.; Li, G.X.; Cui, Z.L.; Yuan, J. Applicability and limitation of compost maturity evaluation indicators: A review. Chem. Eng. J. 2024, 489, 151386. [Google Scholar] [CrossRef]
- Zhu, Y.P.; Cao, Y.X.; Fu, B.B.; Wang, C.J.; Shu, S.H.; Zhu, P.J.; Wang, D.F.; Xu, H.; Zhong, N.Q.; Cai, D.Q. Waste milk humification product can be used as a slow release nano-fertilizer. Nat. Commun. 2024, 15, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rajapaksha, A.U.; Ok, Y.S.; El-Naggar, A.; Kim, H.; Song, F.H.; Kang, S.; Tsang, Y.F. Dissolved organic matter characterization of biochars produced from different feedstock materials. J. Environ. Manag. 2019, 233, 393–399. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Ni, J.Z.; Wei, R.; Chen, W.F. Water-soluble organic carbon (WSOC) from vegetation fire and its differences from WSOC in natural media: Spectral comparison and self-organizing maps (SOM) classification. Sci. Total Environ. 2023, 895, 165180. [Google Scholar] [CrossRef]
- Yan, C.X.; Wang, W.Y.; Nie, M.H.; Ding, M.J.; Wang, P.; Zhang, H.; Huang, G.X. Characterization of copper binding to biochar-derived dissolved organic matter: Effects of pyrolysis temperature and natural wetland plants. J. Hazard. Mater. 2023, 442, 130076. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Shi, T.; Zhang, X.M.; Wei, J.S.; Lu, J.; Zhu, J.X.; Wu, Z.H.; Liu, Q.; Liu, M. Implementing in-situ self-organizing maps with memristor crossbar arrays for data mining and optimization. Nat. Commun. 2022, 13, 1–10. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Li, G.X.; Gu, J.; Wang, G.Q.; Li, Y.Y.; Zhang, D.F. Influence of aeration on volatile sulfur compounds (VSCs) and NH3 emissions during aerobic composting of kitchen waste. Waste Manag. 2016, 58, 369–375. [Google Scholar] [CrossRef]
- Bieroza, M.; Baker, A.; Bridgeman, J. Exploratory analysis of excitation–emission matrix fluorescence spectra with self-organizing maps—A tutorial. Educ. Chem. Eng. 2012, 7, 22–31. [Google Scholar] [CrossRef]
- Ke, W.S.; Liu, Z.; Zhu, F.; Xie, Y.; Hartley, W.; Li, X.; Wu, H.; Xue, S.G. Remediation potential of magnetic biochar in lead smelting sites: Insight from the complexation of dissolved organic matter with potentially toxic elements. J. Environ. Manag. 2023, 344, 118556. [Google Scholar] [CrossRef]
- Yu, S.H.; Zhang, H.Y.; Ni, J.Z.; Xiang, Y.; Wei, R.; Qian, W.; Chen, W.F. Spectral characteristics coupled with self-organizing maps analysis on different molecular size-fractionated water-soluble organic carbon from biochar. Sci. Total Environ. 2023, 857, 159424. [Google Scholar] [CrossRef] [PubMed]
- Fleming, M.; Tai, Y.P.; Zhuang, P.; McBride, M.B. Extractability and bioavailability of Pb and As in historically contaminated orchard soil: Effects of compost amendments. Environ. Pollut. 2013, 177, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Bi, Z.T.; Zhang, Y.C.; Wu, H.; Zhou, L.; Zhang, H.Q. Impact of wine grape pomace on humification performance and microbial dynamics during pig manure composting. Bioresour. Technol. 2022, 358, 127380. [Google Scholar] [CrossRef]
- Liu, H.X.; Yang, Y.J.; Yang, Y.Z.; Zhong, X.B.; Lv, J.L. Dynamics of fungal and bacterial communities in different types of soil ageing with different dosages of cadmium. Ecotox. Environ. Saf. 2022, 242, 113860. [Google Scholar] [CrossRef]
- Liu, Y.; Ma, R.N.; Li, D.Y.; Qi, C.R.; Han, L.N.; Chen, M.; Fu, F.; Yuan, J.; Li, G.X. Effects of calcium magnesium phosphate fertilizer, biochar and spent mushroom substrate on compost maturity and gaseous emissions during pig manure composting. J. Environ. Manag. 2020, 267, 110649. [Google Scholar] [CrossRef]
- Wang, G.Y.; Kong, Y.L.; Yang, Y.; Ma, R.N.; Shen, Y.J.; Li, G.X.; Yuan, J. Superphosphate, biochar, and a microbial inoculum regulate phytotoxicity and humification during chicken manure composting. Sci. Total Environ. 2022, 824, 153958. [Google Scholar] [CrossRef]
- Chen, Y.X.; Huang, X.D.; Han, Z.Y.; Huang, X.; Hu, B.; Shi, D.Z.; Wu, W.X. Effects of bamboo charcoal and bamboo vinegar on nitrogen conservation and heavy metals immobility during pig manure composting. Chemosphere 2010, 78, 1177–1181. [Google Scholar] [CrossRef]
- Li, Y.; Luo, W.H.; Li, G.X.; Wang, K.; Gong, X.Y. Performance of phosphogypsum and calcium magnesium phosphate fertilizer for nitrogen conservation in pig manure composting. Bioresour. Technol. 2018, 250, 53–59. [Google Scholar] [CrossRef]
- Bernal, M.P.; Alburquerque, J.A.; Moral, R. Composting of animal manures and chemical criteria for compost maturity assessment. A review. Bioresour. Technol. 2009, 100, 5444–5453. [Google Scholar] [CrossRef]
- Kong, Y.L.; Wang, G.Y.; Chen, W.J.; Yang, Y.; Ma, R.N.; Li, D.Y.; Shen, Y.J.; Li, G.X.; Yuan, J. Phytotoxicity of farm livestock manures in facultative heap composting using the seed germination index as indicator. Ecotox. Environ. Saf. 2022, 247, 114251. [Google Scholar] [CrossRef]
- Ji, Z.Y.; Zhang, L.Y.; Liu, Y.W.; Li, X.Q.; Li, Z.J. Evaluation of composting parameters, technologies and maturity indexes for aerobic manure composting: A meta-analysis. Sci. Total Environ. 2023, 886, 163929. [Google Scholar] [CrossRef]
- Ravindran, B.; Awasthi, M.K.; Karmegam, N.; Chang, S.W.; Chaudhary, D.K.; Selvam, A.; Nguyen, D.D.; Milon, A.R.; Munuswamy-Ramanujam, G. Co-composting of food waste and swine manure augmenting biochar and salts: Nutrient dynamics, gaseous emissions and microbial activity. Bioresour. Technol. 2022, 344, 126300. [Google Scholar] [CrossRef]
- Liu, C.; Du, Y.H.; Yin, H.B.; Fan, C.X.; Chen, K.N.; Zhong, J.C.; Gu, X.Z. Exchanges of nitrogen and phosphorus across the sediment-water interface influenced by the external suspended particulate matter and the residual matter after dredging. Environ. Pollut. 2019, 246, 207–216. [Google Scholar] [CrossRef]
- Lu, K.T.; Gao, H.J.; Yu, H.B.; Liu, D.P.; Zhu, N.M.; Wan, K.L. Insight into variations of DOM fractions in different latitudinal rural black-odor waterbodies of eastern China using fluorescence spectroscopy coupled with structure equation model. Sci. Total Environ. 2022, 816, 151531. [Google Scholar] [CrossRef]
- Li, J.Y.; Wu, S.W.; Zheng, J.X.; Sun, X.C.; Hu, C.X. Combining citrus waste-derived function microbes with biochar promotes humus formation by enhancing lignocellulose degradation in citrus waste compost. Chemosphere 2024, 368, 143754. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.W.; Awasthi, M.K.; Syed, A.; Bahkali, A.H. Effect of cyanobacteria biochar addition on humification, fungal dynamics and its mechanism of action in pig manure composting. J. Environ. Chem. Eng. 2024, 12, 113755. [Google Scholar] [CrossRef]
- Catalá, T.S.; Reche, I.; Fuentes, L.A.; Romera-Castillo, C.; Nieto-Cid, M.; Ortega-Retuerta, E.; Calvo, E.; Álvarez, M.; Marrasé, C.; Stedmon, C.A.; et al. Turnover time of fluorescent dissolved organic matter in the dark global ocean. Nat. Commun. 2015, 6, 5986. [Google Scholar] [CrossRef]
- Kong, Y.L.; Ma, R.N.; Li, G.X.; Wang, G.Y.; Liu, Y.; Yuan, J. Impact of biochar, calcium magnesium phosphate fertilizer and spent mushroom substrate on humification and heavy metal passivation during composting. Sci. Total Environ. 2022, 824, 153755. [Google Scholar] [CrossRef]
- Chen, L.J.; Lin, Y.; Liu, C.W.; Zhang, H.; Lin, C.Q. Improving the Utilization of Flammulina velutipes Waste during Biochar-Amended Composting: Emphasis on Bacterial Communities. Agronomy 2024, 14, 1046. [Google Scholar] [CrossRef]
- Jiang, K.H.; Jiang, D.M.; Li, S.; Guo, Z.Z.; Zhao, L.B.; Wang, J.; Hao, X.X.; Bai, L.; Qiu, S.X.; Kang, B. Impacts of mixed ferrous sulfate-biochar additives on humification and bacterial community during electric field-assisted aerobic composting. Bioresour. Technol. 2024, 404, 130901. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Liu, L.Q.; Huang, G.W.; Yang, C.H.; Tian, W.X.; Ge, Z.Y.; Zhang, B.H.; Wang, S.F.; Zhang, H.Q. Enhancing humification and microbial interactions during co-composting of pig manure and wine grape pomace: The role of biochar and Fe2O3. Bioresour. Technol. 2024, 393, 130120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.C.; Xu, Y.; Yu, X.L.; Li, J.K.; Chen, G.; Wang, S.J.; Xu, Y.P.; Xu, R.; Zhang, B.H.; Zhang, H.Q. Microbial metabolism and humic acid formation in response to enhanced copper and zinc passivation during composting of wine grape pomace and pig manure. Bioresour. Technol. 2023, 384, 129226. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Zhao, Y.; Wang, R.X.; Lu, Q.; Wu, J.Q.; Zhang, D.Y.; Nie, Z.F.; Wei, Z.M. Effect of the addition of exogenous precursors on humic substance formation during composting. Waste Manag. 2018, 79, 462–471. [Google Scholar] [CrossRef]
- Wei, Y.H.; Liang, Z.W.; Zhang, Y. Evolution of physicochemical properties and bacterial community in aerobic composting of swine manure based on a patent compost tray. Bioresour. Technol. 2022, 343, 126136. [Google Scholar] [CrossRef]
- Ulrich, K.; Becker, R.; Behrendt, U.; Kube, M.; Schneck, V.; Ulrich, A. Physiological and genomic characterisation of Luteimonas fraxinea sp. nov., a bacterial species associated with trees tolerant to ash dieback. Syst. Appl. Microbiol. 2022, 45, 126333. [Google Scholar] [CrossRef] [PubMed]
- Hur, J.; Lee, B.M. Characterization of binding site heterogeneity for copper within dissolved organic matter fractions using two-dimensional correlation fluorescence spectroscopy. Chemosphere 2011, 83, 1603–1611. [Google Scholar] [CrossRef]
- Lu, M.L.; Lin, B.F.; Zhang, Y.; Hao, Y.H.; Li, K.; Huang, Z.; Li, J.B. Insight into the molecular transformation pathways of humic acid in the co-composting of bagasse and cow manure after adding compound microorganisms. Process Biochem. 2024, 143, 23–33. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Yu, X.; Luo, L.; Sun, Y.; Zhou, L.; Fu, H. Self-Organizing Map-Based Assessment of Compost Maturity and Cu/Zn Passivation in Biochar-Amended Pig Manure. Agronomy 2025, 15, 778. https://doi.org/10.3390/agronomy15040778
Zhang H, Yu X, Luo L, Sun Y, Zhou L, Fu H. Self-Organizing Map-Based Assessment of Compost Maturity and Cu/Zn Passivation in Biochar-Amended Pig Manure. Agronomy. 2025; 15(4):778. https://doi.org/10.3390/agronomy15040778
Chicago/Turabian StyleZhang, Hongqiong, Xinlong Yu, Lina Luo, Yong Sun, Ling Zhou, and Haimei Fu. 2025. "Self-Organizing Map-Based Assessment of Compost Maturity and Cu/Zn Passivation in Biochar-Amended Pig Manure" Agronomy 15, no. 4: 778. https://doi.org/10.3390/agronomy15040778
APA StyleZhang, H., Yu, X., Luo, L., Sun, Y., Zhou, L., & Fu, H. (2025). Self-Organizing Map-Based Assessment of Compost Maturity and Cu/Zn Passivation in Biochar-Amended Pig Manure. Agronomy, 15(4), 778. https://doi.org/10.3390/agronomy15040778