
Calcium Allocation to the Tree Canopy and the Edible Part of Sweet Cherry Fruit Is Hindered by Boron Soil Deficiency

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
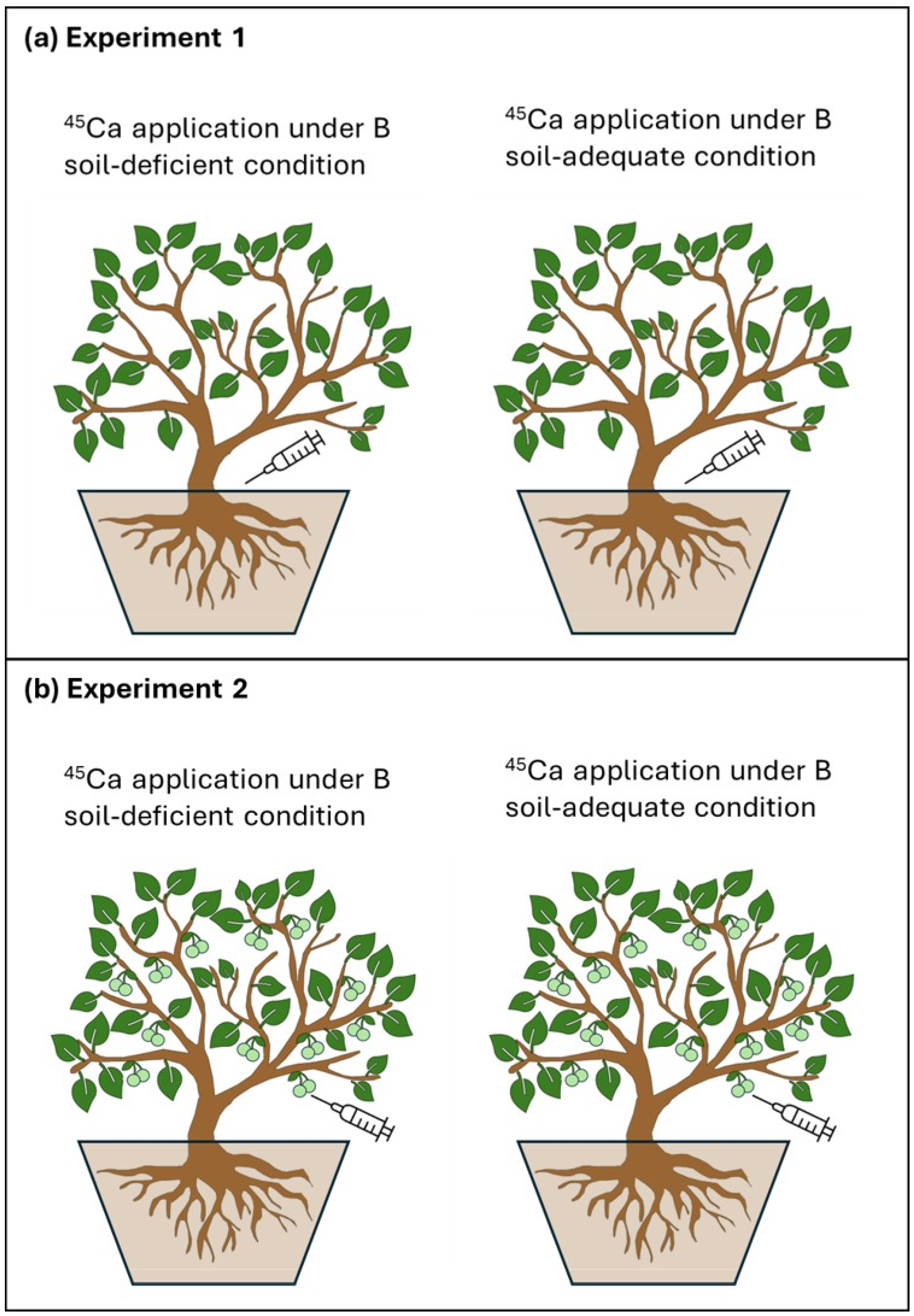
2.1. Plant Materials and Treatments
2.2. Isotopic 45Ca Application
2.3. Plant and Soil Samples for Analytical Determinations of Ca
3. Results and Discussion
3.1. Total Ca Recovery in the Trees
3.2. Ca Recovery in Different Tree Tissues
3.3. Ca Recovery in the Fruit
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Freitas, S.T.; Mitcham, E.J. Factors involved in fruit calcium deficiency disorders. Hortic. Rev. 2012, 40, 107–146. [Google Scholar] [CrossRef]
- Hocking, B.; Tyerman, S.D.; Burton, R.A.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef]
- Montanaro, G.; Dichio, B.; Xiloyannis, C. Significance of fruit transpiration on calcium nutrition in developing apricot fruit. J. Plant Nutr. Soil Sci. 2010, 173, 618–622. [Google Scholar] [CrossRef]
- Winkler, A.; Fiedler, B.; Knoche, M. Calcium physiology of sweet cherry fruits. Trees 2020, 34, 1157–1167. [Google Scholar] [CrossRef]
- Lötze, E.; Joubert, J.; Theron, K.I. Evaluating pre-harvest foliar calcium applications to increase fruit calcium and reduce bitter pit in ‘Golden Delicious’ apples. Sci. Hortic. 2008, 116, 299–304. [Google Scholar] [CrossRef]
- Fernández, V.; Sotiropoulos, T.; Brown, P.H. Foliar Fertilization: Scientific Principles and Field Practices, 1st ed.; International Fertilizer Industry Association (IFA): Paris, France, 2013; 140p. [Google Scholar]
- Robinson, T.; Lopez, S. Crop load affects ‘Honeycrisp’ fruit quality more than N, K, or irrigation. Acta Hortic. 2012, 940, 529–537. [Google Scholar] [CrossRef]
- Conway, W.S.; Sams, C.E.; Hickey, K.D. Pre- and postharvest calcium treatment of apple fruit and its effect on quality. Acta Hortic. 2002, 594, 413–419. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, X.; Wang, X.; Yan, L.; Hu, X.; Lian, M. Effect of foliar calcium fertilization on fruit quality, cell wall enzyme activity and expression of key genes in Chinese cherry. Int. J. Fruit Sci. 2023, 23, 200–216. [Google Scholar] [CrossRef]
- Brown, P.H.; Bellaloui, N.; Wimmer, M.A.; Bassil, E.S.; Ruiz, J.; Hu, H.; Pfeffer, H.; Dannel, F.; Romheld, V. Boron in plant biology. Plant Biol. 2002, 4, 205–223. [Google Scholar] [CrossRef]
- Liu, Y.; Riaz, M.; Yan, L.; Zeng, Y.; Cuncang, J. Boron and calcium deficiency disturbing the growth of trifoliate rootstock seedlings (Poncirus trifoliate L.) by changing root architecture and cell wall. Plant Physiol. Biochem. 2019, 144, 345–354. [Google Scholar] [CrossRef]
- Marschner, P. Mineral Nutrition of Higher Plants, 3rd ed.; Academic Press: London, UK, 2012. [Google Scholar] [CrossRef]
- Kobayashi, M.; Nakagawa, H.; Asaka, T.; Matoh, T. Borate-rhamnogalacturonan II bonding reinforced by Ca2+ retains pectic polysaccharides in higher-plant cell walls. Plant Physiol. 1999, 119, 199–204. [Google Scholar] [CrossRef] [PubMed]
- Arredondo, G.; Bonomelli, C. Effect of three boron concentrations in soil on growth and physiology in sweet cherry trees. Plants 2023, 12, 1240. [Google Scholar] [CrossRef] [PubMed]
- Botelho, R.V.; Müller, M.M.L.; Umburanas, R.C.; Laconski, J.M.O.; Terra, M.M. Boron in fruit crops: Plant physiology, deficiency, toxicity, and sources for fertilization. In Boron in Plants and Agriculture; Aftab, T., Landi, M., Papadakis, I.E., Araniti, F., Brown, P.H., Eds.; Academic Press: London, UK, 2022; pp. 29–50. [Google Scholar] [CrossRef]
- Chen, Y.; Smagula, J.M.; Litten, W.; Dunham, S. Effect of Boron and Calcium Foliar Sprays on Pollen Germination and Development, Fruit Set, Seed Development, and Berry Yield and Quality in Lowbush Blueberry (Vaccinium angustifolium Ait.). J. Am. Soc. Hort. Sci. 1998, 123, 524–531. [Google Scholar] [CrossRef]
- Bhat, Z.A.; Rather, T.R.; Rather, G.H.; Kounser, J.; Itoo, H.U.; Sheikh, K.A.; Nisar, S. Effect of calcium and boron application on fruit set, yield and quality of apple cv Ambri. Int. J. Curr. Microbiol. Appl. Sci. 2020, 11, 4038–4042. [Google Scholar]
- Wójcik, P.; Akgül, H.; Demirtaş, İ.; Sarısu, C.; Aksu, M.; Gubbuk, H. Effect of preharvest sprays of calcium chloride and sucrose on cracking and quality of ‘Burlat’ sweet cherry fruit. J. Plant Nutr. 2013, 36, 1453–1465. [Google Scholar] [CrossRef]
- Knoche, M.; Winkler, A. Rain-induced cracking of sweet cherries. In Cherries: Botany, Production and Uses; Quero-García, J., Lezzoni, A., Puławska, J., Lang, G., Eds.; CABI: Wallingford, UK, 2017; pp. 140–165. [Google Scholar]
- Wojcik, P.; Wojcik, M. Effect of boron fertilization on sweet cherry tree yield and fruit quality. J. Plant Nutr. 2006, 29, 1755–1766. [Google Scholar] [CrossRef]
- von Uexküll, H.R.; Mutert, E. Global extent, development and economic impact of acid soils. Plant Soil 1995, 171, 1–15. [Google Scholar] [CrossRef]
- Vera-Maldonado, P.; Aquea, F.; Reyes-Díaz, M.; Carcamo-Fincheira, P.; Soto-Cerda, B.; Nunes-Nesi, A.; Inostroza-Blancheteau, C. Role of boron and its interaction with other elements in plants. Front. Plant Sci. 2024, 15, 1332459. [Google Scholar] [CrossRef]
- Mathew, J.; Krishnakumar, V.; Srinivasan, V.; Bhat, R.; Namboothiri, C.; Haris, A. Standardization of critical boron level in soil and leaves of coconut palms grown in a tropical Entisol. J. Soil Sci. Plant Nutr. 2018, 18, 376–387. [Google Scholar] [CrossRef]
- Fadón, E.; Herrero, M.; Rodrigo, J. Flower development in sweet cherry framed in the BBCH scale. Sci. Hortic. 2015, 192, 141–147. [Google Scholar] [CrossRef]
- Videla, X.; Rojas-Silva, X.; Nario, A.; Arias, M.I.; Bonomelli, C. Calcination method of 45Ca samples for isotope ratio analysis via liquid scintillation. Commun. Soil Sci. Plant Anal. 2019, 50, 412–420. [Google Scholar] [CrossRef]
- Napier, N.; Combrink, J. Aspects of calcium nutrition to limit plant physiological disorders. Acta Hortic. 2006, 702, 107–116. [Google Scholar] [CrossRef]
- Poza-Viejo, L.; Abreu, I.; González-García, M.P.; Allauca, P.; Bonilla, I.; Bolaños, L.; Reguera, M. Boron deficiency inhibits root growth by controlling meristem activity under cytokinin regulation. Plant Sci. 2018, 270, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Wimmer, M.A.; Eichert, T. Mechanisms for boron deficiency-mediated changes in plant water relations. Plant Sci. 2013, 203, 25–32. [Google Scholar] [CrossRef]
- Yamauchi, T.; Hara, T.; Sonoda, Y. Effects of boron deficiency and calcium supply on the calcium metabolism in tomato plant. Plant Soil 1986, 93, 223–230. [Google Scholar] [CrossRef]
- Tepecik, M.; Şen, F.; Irget, M.E.; Karaçali, I.; Elmaci, Ö.L. Post-Harvest Calcium and Boron Treatments on Retranslocation of Ca from Senescent Apple Leaves. Fresenius Environ. Bull. 2014, 23, 2229–2234. [Google Scholar]
- White, P.; Broadley, M. Calcium in Plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef] [PubMed]
- Saure, M.C. Calcium translocation to fleshy fruit: Its mechanism and endogenous control. Sci. Hortic. 2005, 105, 65–89. [Google Scholar] [CrossRef]
- Bonomelli, C.; Gil, P.; Schaffer, B. Effect of soil type on calcium absorption and partitioning in young avocado (Persea americana Mill.) trees. Agronomy 2019, 9, 837. [Google Scholar] [CrossRef]
- Famiani, F.; Casulli, V.; Baldicchi, A.; Battistelli, A.; Moscatello, S.; Walker, R.P. Development and metabolism of the fruit and seed of the Japanese plum Ozark premier (Rosaceae). J. Plant Physiol. 2012, 169, 551–560. [Google Scholar] [CrossRef]
- Famiani, F.; Bonghi, C.; Chen, Z.-H.; Drincovich, M.F.; Farinelli, D.; Lara, M.V.; Proietti, S.; Rosati, A.; Vizzotto, G.; Walker, R.P. Stone fruits: Growth and nitrogen and organic acid metabolism in the fruits and seeds—A review. Front. Plant Sci. 2020, 11, 572601. [Google Scholar] [CrossRef] [PubMed]
- Grimm, E.; Pflugfelder, D.; van Dusschoten, D.; Winkler, A.; Knoche, M. Physical rupture of the xylem in developing sweet cherry fruit causes progressive decline in xylem sap inflow rate. Planta 2017, 246, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Gärtner, J.; Grimm, E.; Knoche, M. Xylogenesis and phloemogenesis in the flesh of sweet cherry fruit are limited to early-stage development. Sci. Rep. 2022, 12, 12274. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| B Soil Availability | Total Plant 45Ca Recovery (%) |
|---|---|
| Deficient | 29.9 |
| Adequate | 54.5 |
| p-value * | 0.05 |
| Boron Soil Availability | 45Ca Allocation in Sweet Cherry Tree (%) | |||
|---|---|---|---|---|
| Shoots | Fruits | Trunk | Roots | |
| Deficient | 31 b | 11 a | 13 a | 45 a |
| Adequate | 45 a | 13 a | 14 a | 28 b |
| p-value * | 0.03 | 0.76 | 0.81 | 0.02 |
| B Soil Availability | 45Ca Allocation in Sweet Cherry Fruit (%) | ||
|---|---|---|---|
| Pedicel | Flesh + Peel | Pit | |
| Deficient | 22.6 a | 21.3 b | 56.1 a |
| Adequate | 24.3 a | 25.1 a | 50.6 a |
| p-value * | 0.78 | 0.04 | 0.53 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonomelli, C.; Arredondo, G.; Nario, A.; Artacho, P.; Contreras, C. Calcium Allocation to the Tree Canopy and the Edible Part of Sweet Cherry Fruit Is Hindered by Boron Soil Deficiency. Agronomy 2025, 15, 691. https://doi.org/10.3390/agronomy15030691
Bonomelli C, Arredondo G, Nario A, Artacho P, Contreras C. Calcium Allocation to the Tree Canopy and the Edible Part of Sweet Cherry Fruit Is Hindered by Boron Soil Deficiency. Agronomy. 2025; 15(3):691. https://doi.org/10.3390/agronomy15030691
Chicago/Turabian StyleBonomelli, Claudia, Gerardo Arredondo, Adriana Nario, Pamela Artacho, and Carolina Contreras. 2025. "Calcium Allocation to the Tree Canopy and the Edible Part of Sweet Cherry Fruit Is Hindered by Boron Soil Deficiency" Agronomy 15, no. 3: 691. https://doi.org/10.3390/agronomy15030691
APA StyleBonomelli, C., Arredondo, G., Nario, A., Artacho, P., & Contreras, C. (2025). Calcium Allocation to the Tree Canopy and the Edible Part of Sweet Cherry Fruit Is Hindered by Boron Soil Deficiency. Agronomy, 15(3), 691. https://doi.org/10.3390/agronomy15030691