
Modeling the Effects of Extreme Temperatures on the Infection Rate of Botrytis cinerea Using Historical Climate Data (1951–2023) of Central Chile

 ,
,  , , and
, , and
Abstract
1. Introduction
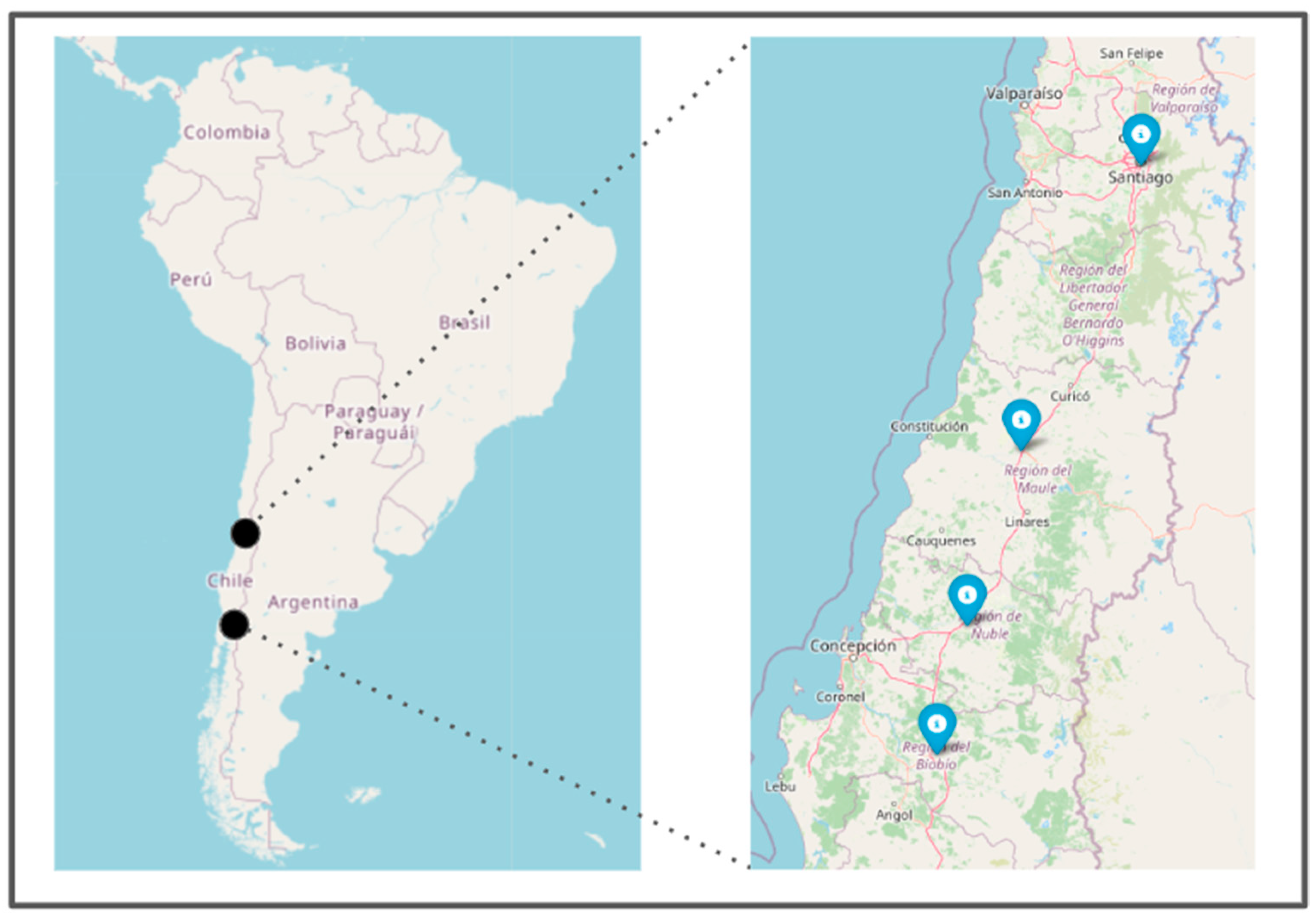
2. Materials and Methods
3. Results
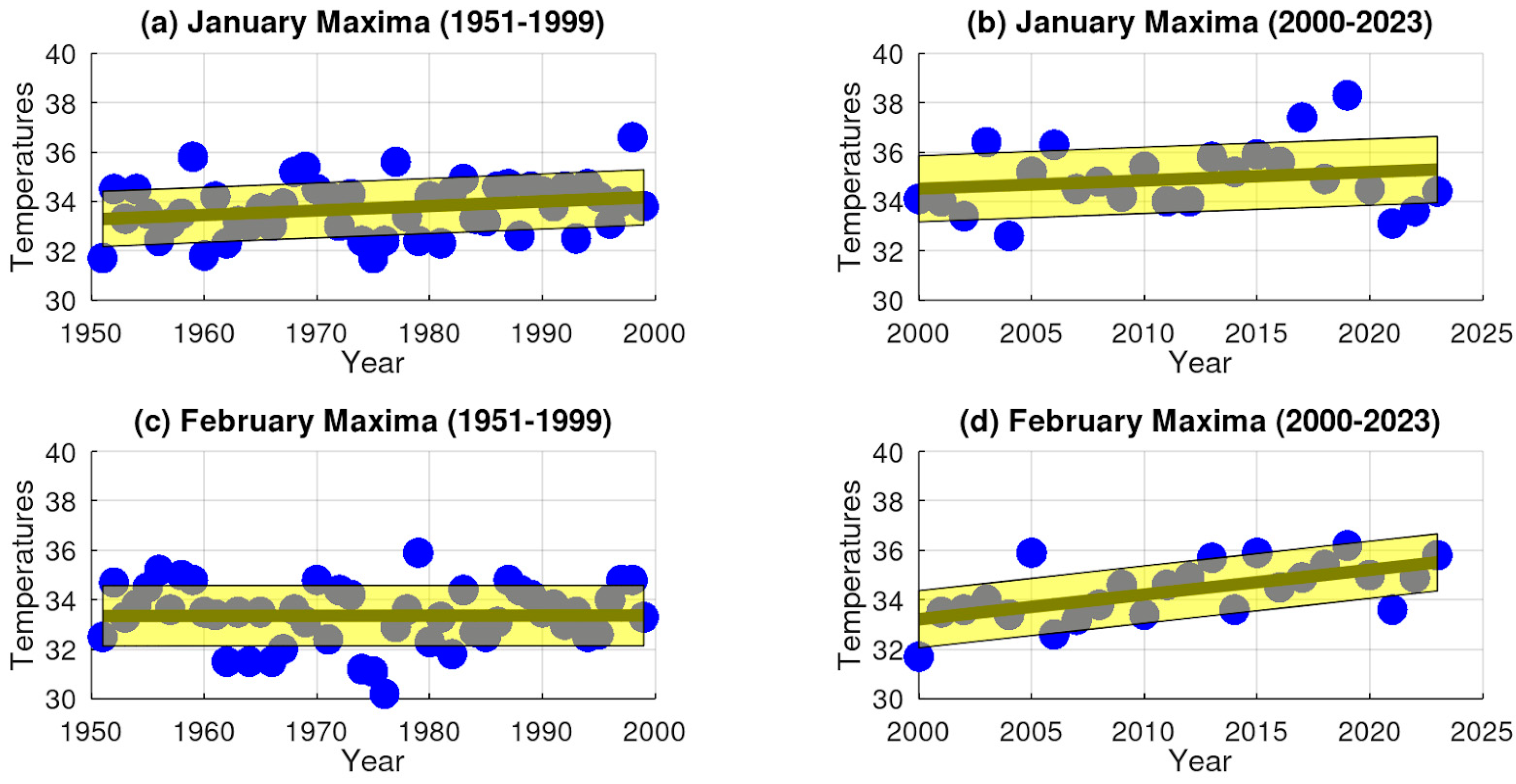
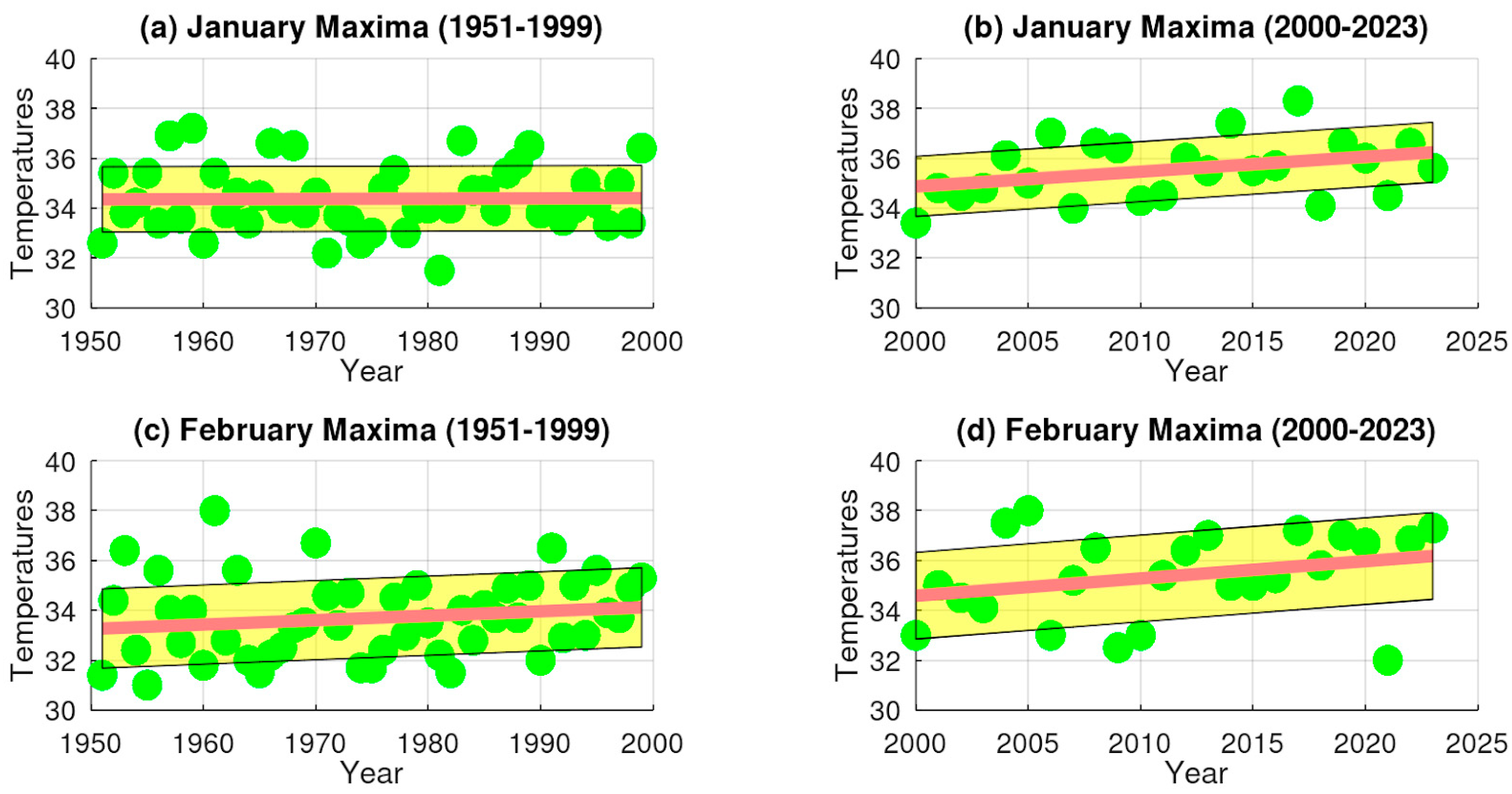
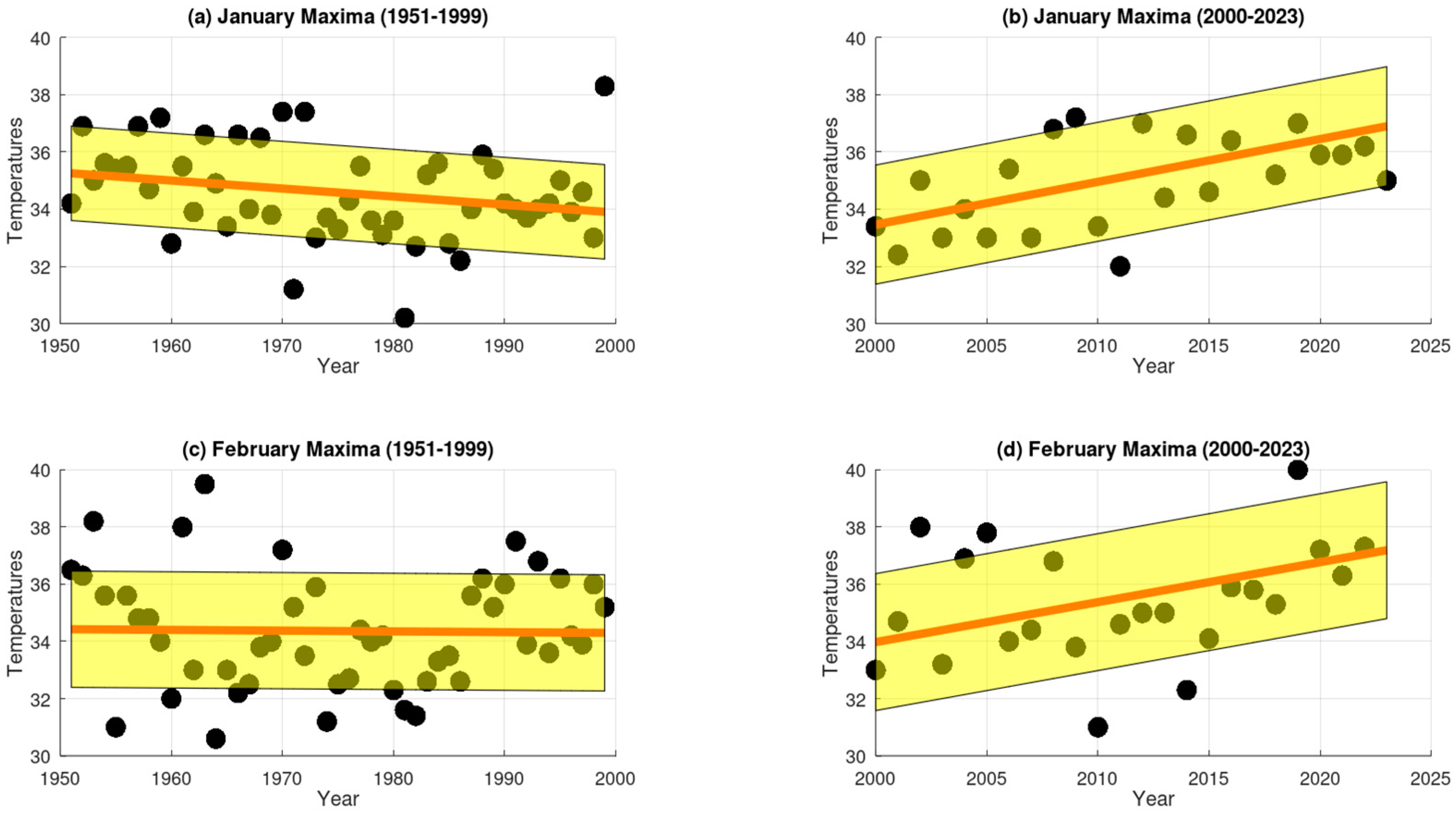
3.1. Extreme Temperature Trends
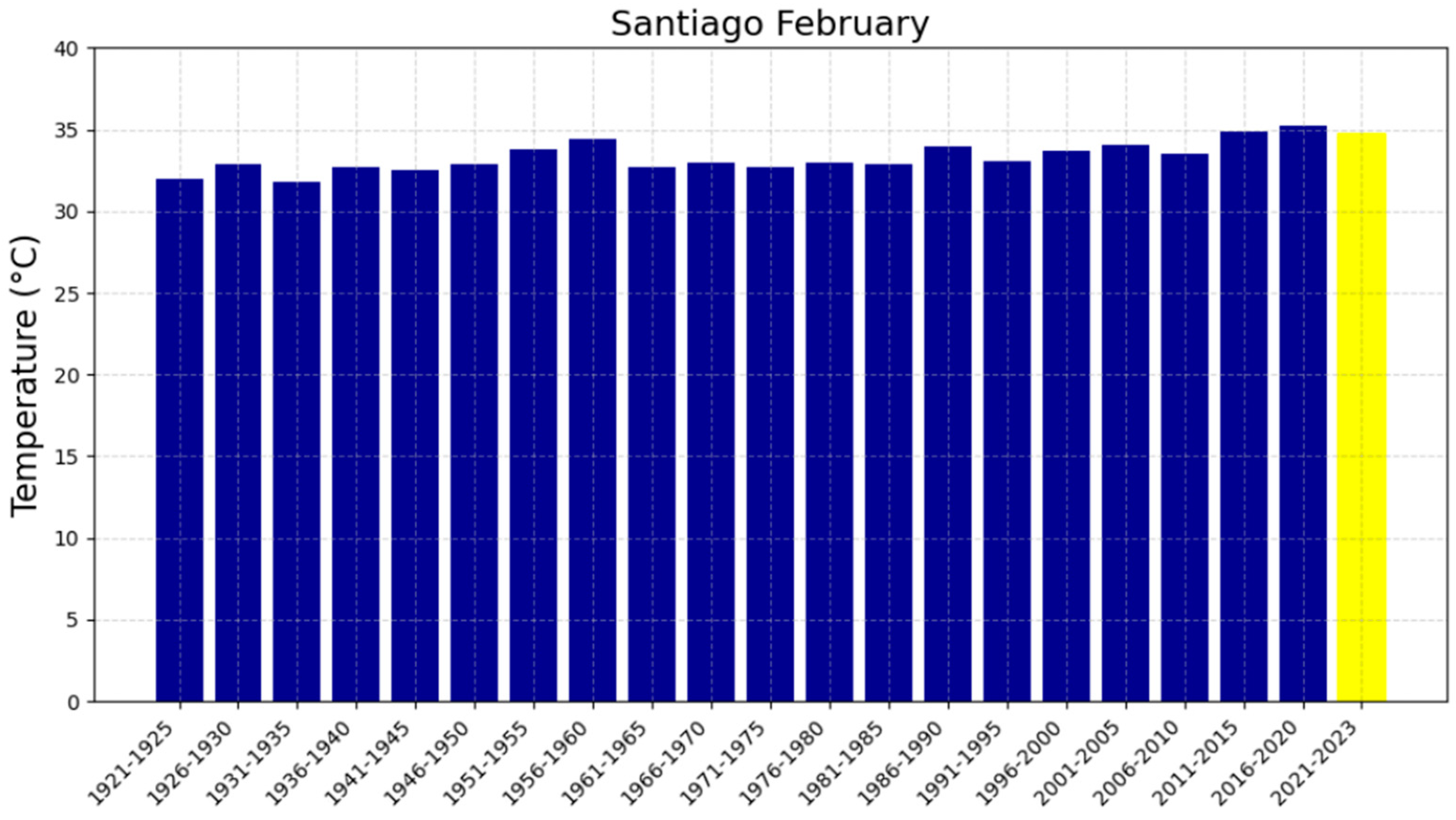
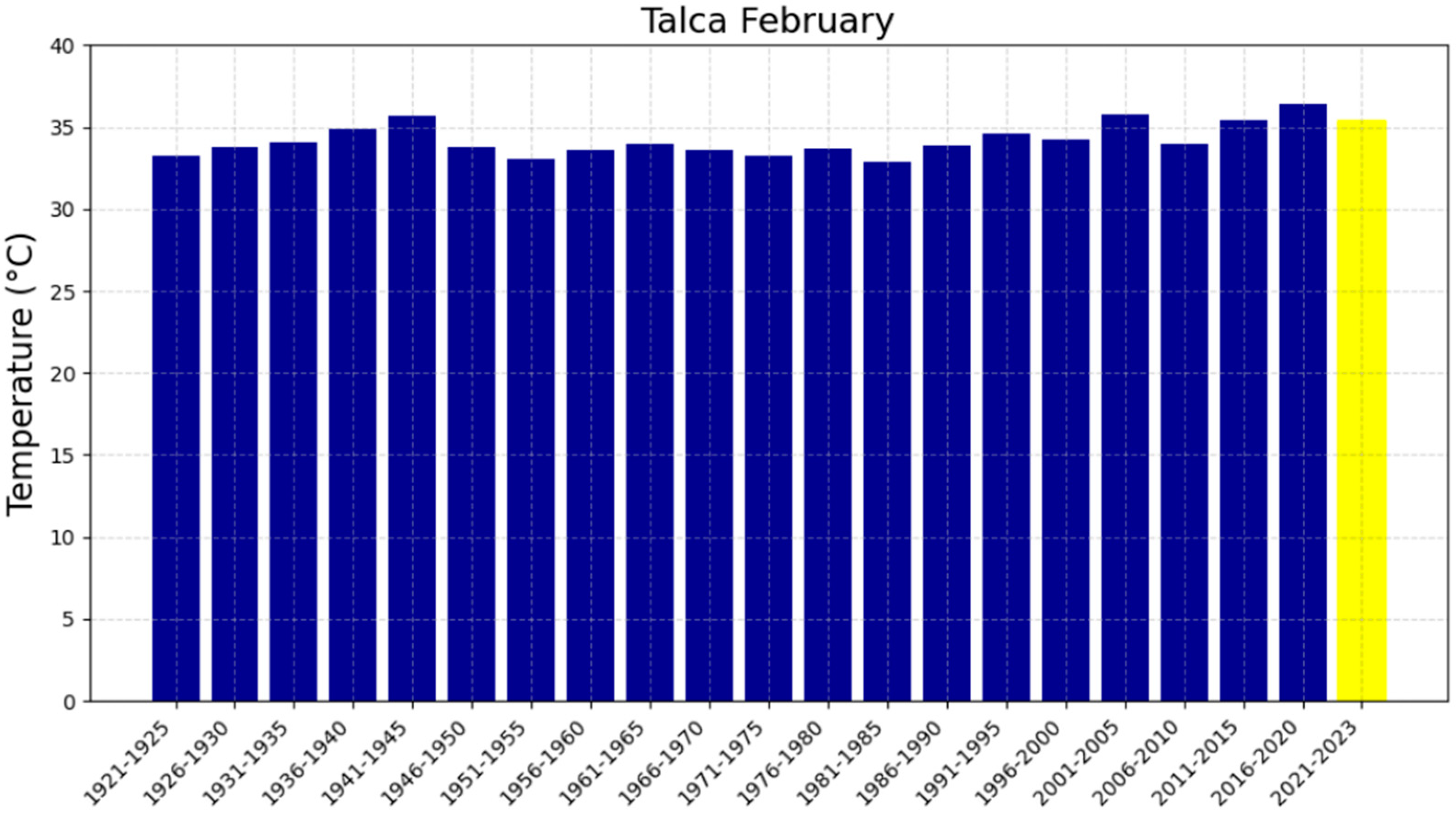
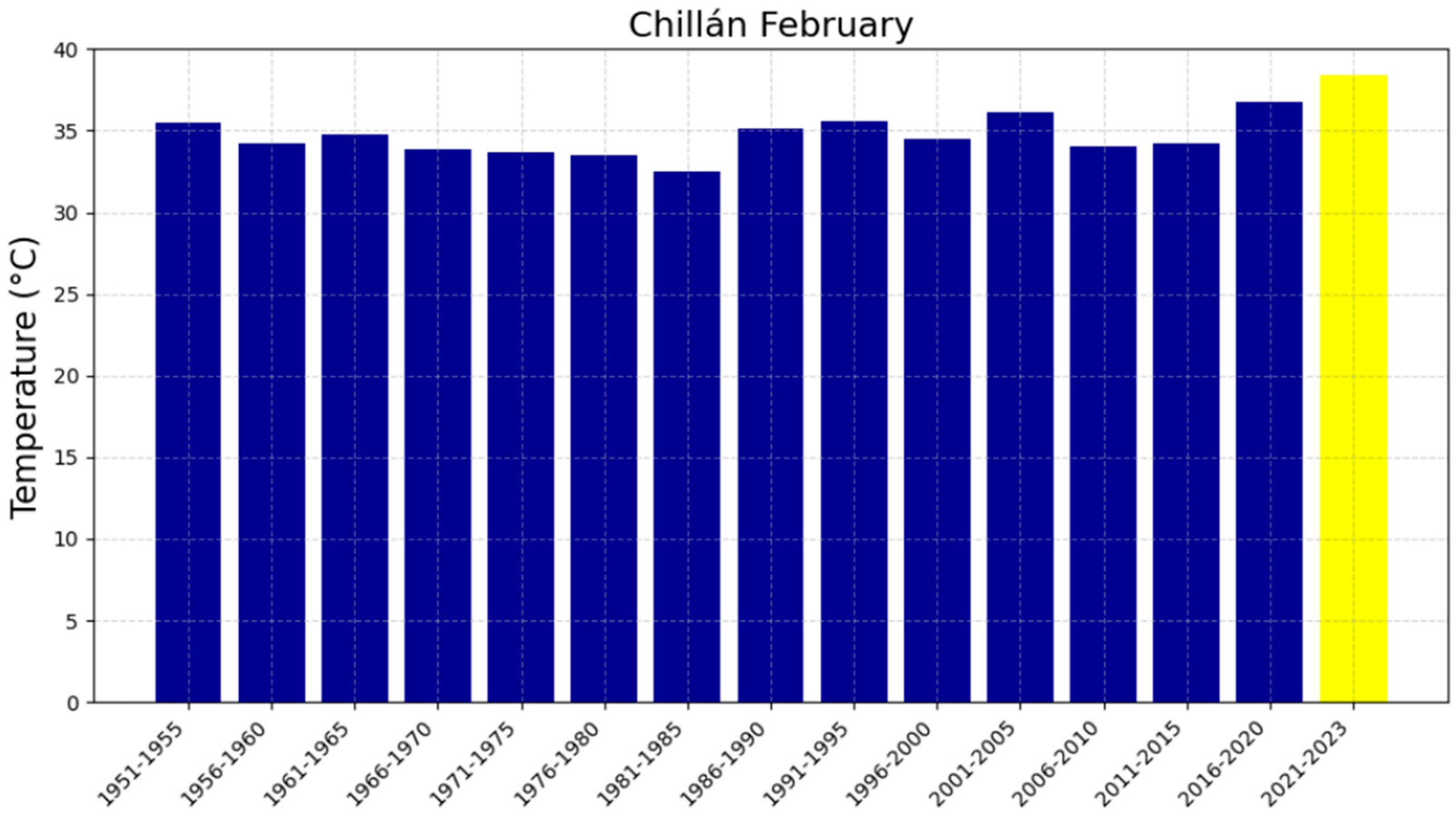
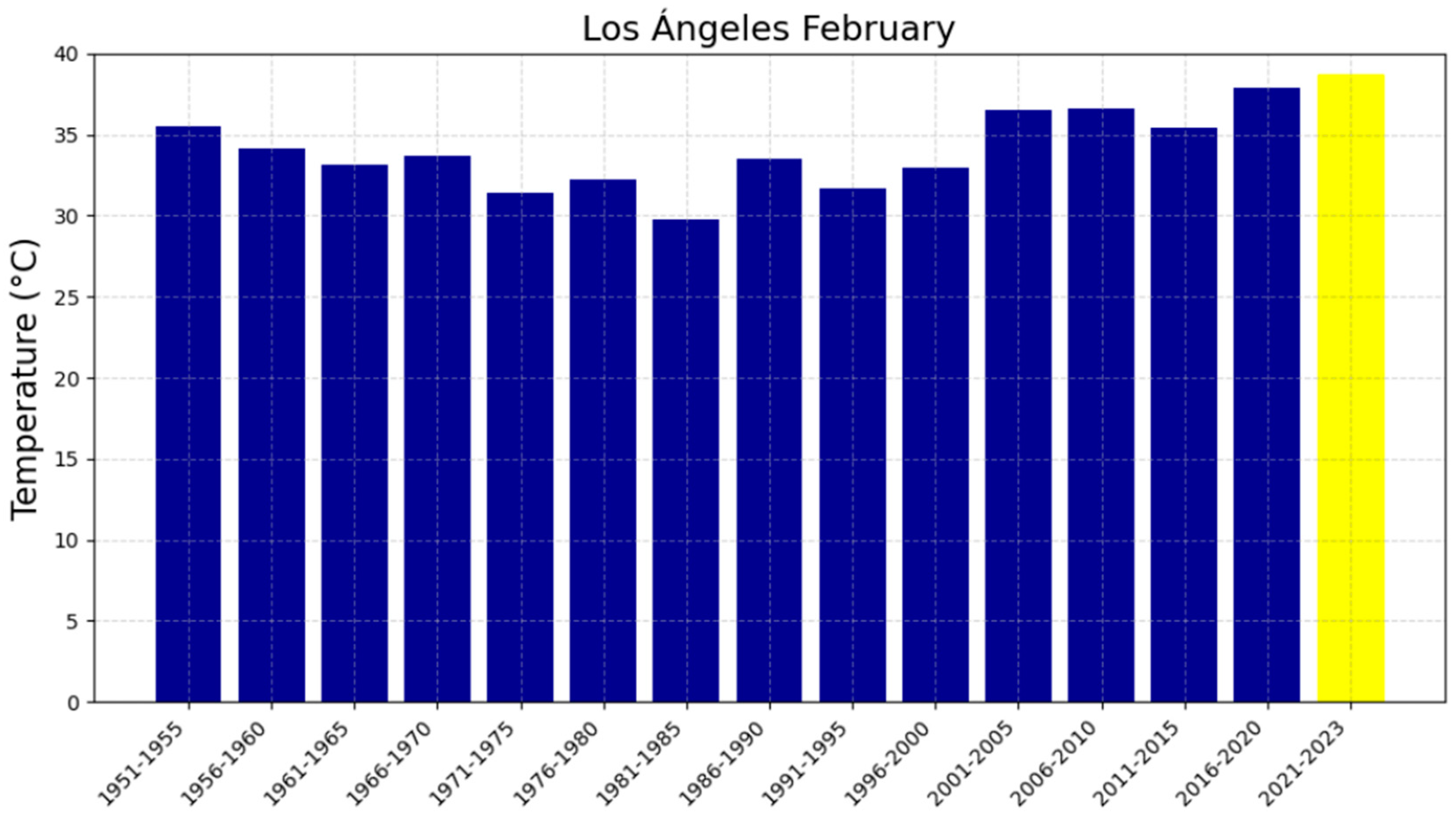
3.2. The Five-Year Averages of Extreme Maximum Temperatures (EMTs)
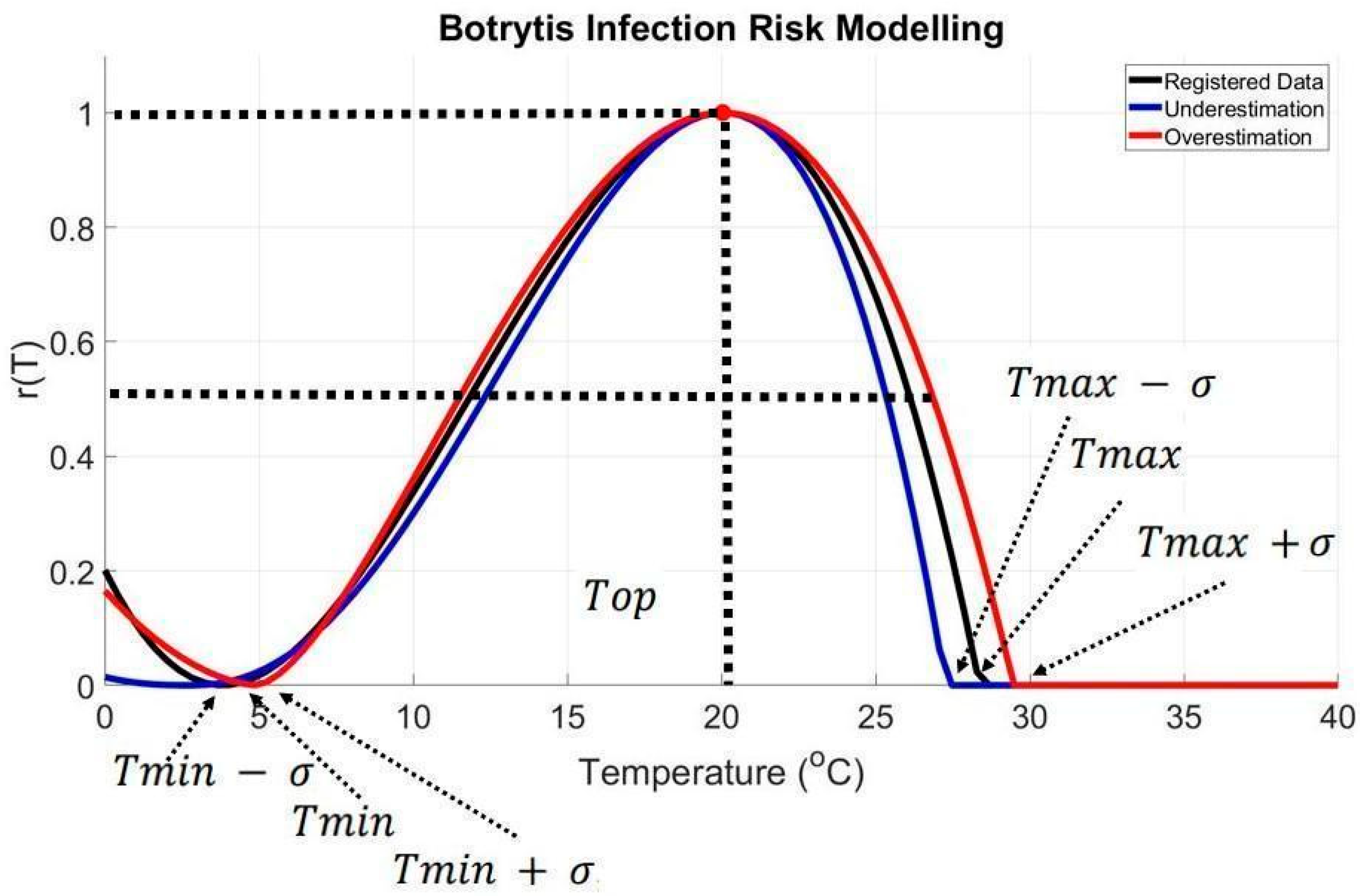
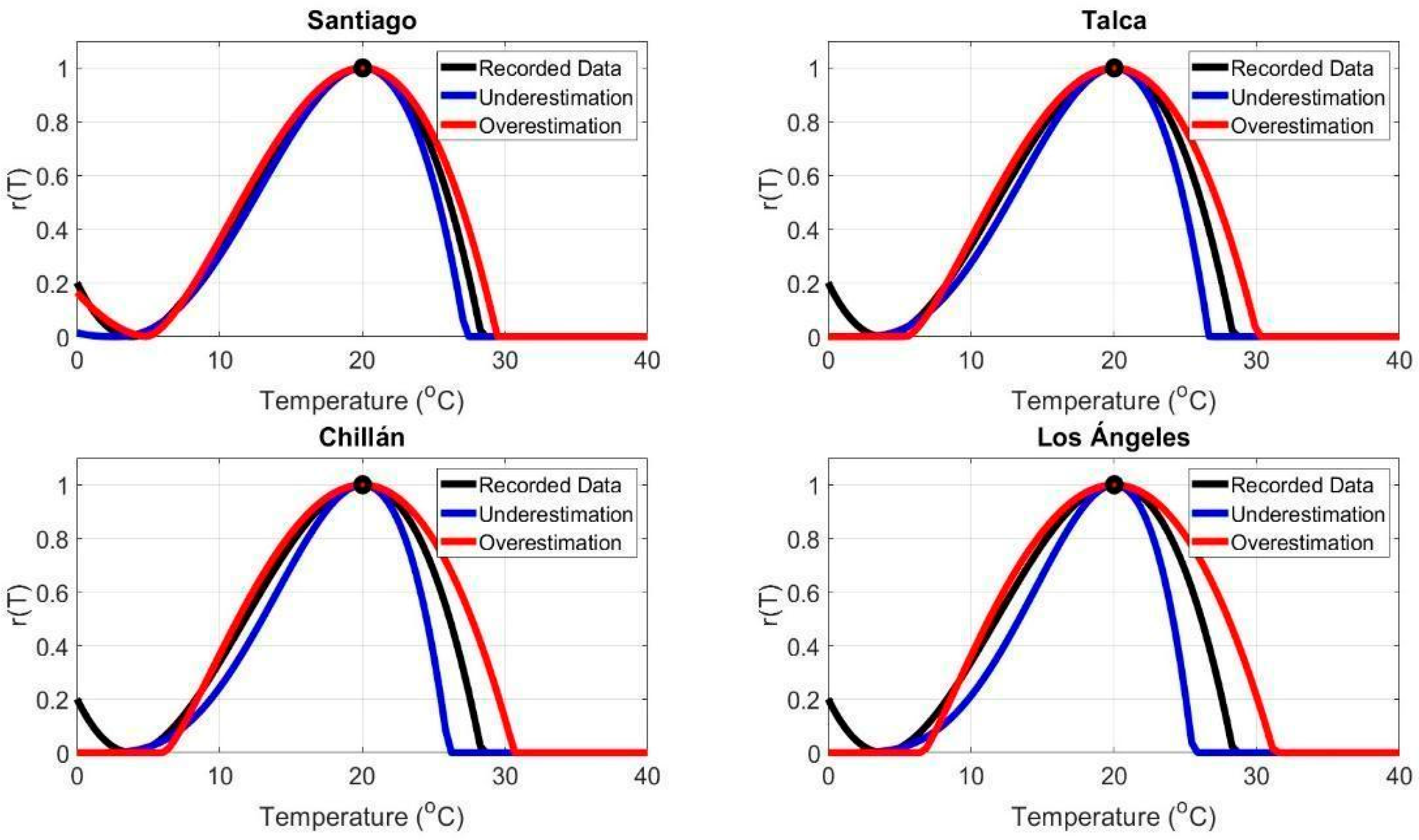
3.3. Effects of Extreme Temperatures on Infection Rate
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Molina-Montenegro, M.A. Climate change impacts and adaptation strategies of agriculture in Mediterranean-climate regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef]
- Vautard, R.; Cattiaux, J.; Happé, T.; Singh, J.; Bonnet, R.; Cassou, C.; Yiou, P. Heat extremes in Western Europe increasing faster than simulated due to atmospheric circulation trends. Nat. Commun. 2023, 14, 6803. [Google Scholar] [CrossRef]
- Campillay-Llanos, W.; Gonzalez-Colville, P.; Ortega-Farias, S.; López-Flores, M.M. Historical trends and future scenario projection of maximum summer temperatures in the southern hemisphere: Central-southern zone of Chile. Idesia 2024, 42, 43–54. [Google Scholar] [CrossRef]
- Hatfield, J.L.; Antle, J.; Garrett, K.A.; Izaurralde, R.C.; Mader, T.; Marshall, E.; Nearing, M.; Robertson, G.P.; Ziska, L. Indicators of climate change in agricultural systems. Clim. Change 2020, 163, 1719–1732. [Google Scholar] [CrossRef]
- Litchman, E.; Thomas, M.K. Are we underestimating the ecological and evolutionary effects of warming? Interactions with other environmental drivers may increase species vulnerability to high temperatures. Oikos 2023, 2023, e09155. [Google Scholar] [CrossRef]
- Chaloner, T.M.; Gurr, S.J.; Bebber, D.P. Plant pathogen infection risk tracks global crop yields under climate change. Nat. Clim. Change 2021, 11, 710–715. [Google Scholar] [CrossRef]
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of climate change on crops adaptation and strategies to tackle its outcome: A review. Plants 2019, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The impact of climate change on agricultural insect pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.T.; Eisenhauer, N.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Change 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Ummenhofer, C.C.; Meehl, G.A. Extreme weather and climate events with ecological relevance: A review. Philos. Trans. R. Soc. B 2017, 372, 20160135. [Google Scholar] [CrossRef]
- Cassou, C.; Cattiaux, J. Disruption of the European climate seasonal clock in a warming world. Nat. Clim. Change 2016, 6, 589–594. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Fasullo, J.T. An apparent hiatus in global warming? Earth’s Future 2013, 1, 19–32. [Google Scholar] [CrossRef]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogée, J.; Allard, V.; Valentini, R. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2005, 437, 529–533. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Summary for Policymakers. In Climate Change 2013: The Physical Science Basis; Stocker, T.F., Qin, D., Plattner, G.-K., Tignor, M.M.B., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Barredo, J.I.; Mauri, A.; Caudullo, G.; Dosio, A. Assessing shifts of Mediterranean and arid climates under RCP4.5 and RCP8.5 climate projections in Europe. In Meteorology and Climatology of the Mediterranean and Black Seas; Springer: Berlin/Heidelberg, Germany, 2019; pp. 235–251. [Google Scholar] [CrossRef]
- Sippel, S.; Zscheischler, J.; Heimann, M.; Otto, F.E.L.; Peters, J.; Mahecha, M.D. Quantifying changes in climate variability and extremes: Pitfalls and their overcoming. Geophys. Res. Lett. 2015, 42, 9990–9998. [Google Scholar] [CrossRef]
- Singh, B.K.; Delgado-Baquerizo, M.; Egidi, E.; Guirado, E.; Leach, J.E.; Liu, H.; Trivedi, P. Climate change impacts on plant pathogens, food security and paths forward. Nat. Rev. Microbiol. 2023, 21, 640–656. [Google Scholar] [CrossRef] [PubMed]
- Rezende, E.L.; Bozinovic, F.; Szilágyi, A.; Santos, M. Predicting temperature mortality and selection in natural Drosophila populations. Science 2020, 369, 1242–1245. [Google Scholar] [CrossRef] [PubMed]
- Sillero, N.; Arenas-Castro, S.; Enriquez-Urzelai, U.; Vale, C.G.; Sousa-Guedes, D.; Martínez-Freiría, F.; Real, R.; Barbosa, A.M. Want to model a species niche? A step-by-step guideline on correlative ecological niche modelling. Ecol. Model. 2021, 456, 109671. [Google Scholar] [CrossRef]
- Droby, S.; Lichter, A. Post-harvest Botrytis infection: Etiology, development and management. In Botrytis: Biology, Pathology and Control; Springer: Dordrecht, The Netherlands, 2007; pp. 349–367. [Google Scholar] [CrossRef]
- Magarey, R.D.; Sutton, T.B.; Thayer, C.L. A simple generic infection model for foliar fungal plant pathogens. Phytopathology 2005, 95, 92–100. [Google Scholar] [CrossRef]
- Ponti, R.; Sannolo, M. The importance of including phenology when modelling species ecological niche. Ecography 2023, 2023, e06143. [Google Scholar] [CrossRef]
- Bozinovic, F.; Cavieres, G.; Martel, S.I.; Alruiz, J.M.; Molina, A.N.; Roschzttardtz, H.; Rezende, E.L. Thermal effects vary predictably across levels of organization: Empirical results and theoretical basis. Proc. R. Soc. B 2020, 287, 20202508. [Google Scholar] [CrossRef]
- Villeneuve, A.R.; Komoroske, L.M.; Cheng, B.S. Environment and phenology shape local adaptation in thermal performance. Proc. R. Soc. B 2021, 288, 20210741. [Google Scholar] [CrossRef] [PubMed]
- Wooliver, R.; Vtipilthorpe, E.E.; Wiegmann, A.M.; Sheth, S.N. A viewpoint on ecological and evolutionary study of plant thermal performance curves in a warming world. AoB Plants 2022, 14, plac016. [Google Scholar] [CrossRef] [PubMed]
- Vidal, M.A.; Rezende, E.L.; Bacigalupe, L.D. Intra and interspecific variation in thermal performance and critical limits in anurans from southern Chile. J. Therm. Biol. 2024, 121, 103851. [Google Scholar] [CrossRef] [PubMed]
- Broome, J.C.; English, J.T.; Marois, J.J.; Latorre, B.A.; Aviles, J.C. Development of an infection model for Botrytis bunch rot of grapes based on wetness duration and temperature. Phytopathology 1995, 85, 97–102. [Google Scholar] [CrossRef]
- Latorre, B.A.; Spadaro, I.; Rioja, M.E. Occurrence of resistant strains of Botrytis cinerea to anilinopyrimidine fungicides in table grapes in Chile. Crop Prot. 2002, 21, 957–961. [Google Scholar] [CrossRef]
- Ferrada, E.E.; Latorre, B.A.; Zoffoli, J.P.; Castillo, A. Identification and characterization of Botrytis blossom blight of Japanese plums caused by Botrytis cinerea and B. prunorum sp. nov. in Chile. Phytopathology 2016, 106, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Cheung, N.; Tian, L.; Liu, X.; Li, X. The destructive fungal pathogen Botrytis cinerea: Insights from genes studied with mutant analysis. Pathogens 2020, 9, 923. [Google Scholar] [CrossRef] [PubMed]
- Altieri, V.; Rossi, V.; Fedele, G. Biocontrol of Botrytis cinerea as influenced by grapevine growth stages and environmental conditions. Plants 2023, 12, 3430. [Google Scholar] [CrossRef] [PubMed]
- Melo, O.; Foster, W. Agricultural and forestry land and labor use under long-term climate change in Chile. Atmosphere 2021, 12, 305. [Google Scholar] [CrossRef]
- Malhi, G.S.; Kaur, M.; Kaushik, P. Impact of climate change on agriculture and its mitigation strategies: A review. Sustainability 2021, 13, 1318. [Google Scholar] [CrossRef]
- Farooq, A.; Farooq, N.; Akbar, H.; Hassan, Z.U.; Gheewala, S.H. A critical review of climate change impact at a global scale on cereal crop production. Agronomy 2023, 13, 162. [Google Scholar] [CrossRef]
- Mirón, I.J.; Linares, C.; Díaz, J. The influence of climate change on food production and food safety. Environ. Res. 2023, 216, 114674. [Google Scholar] [CrossRef] [PubMed]
- Cabré, M.F.; Solman, S.; Núñez, M. Regional climate change scenarios over southern South America for future climate (2080–2099) using the MM5 model. Mean, interannual variability and uncertainties. Atmósfera 2016, 29, 35–60. [Google Scholar] [CrossRef]
- González, J.; Velasco, R. Evaluation of the impact of climatic change on the economic value of land in agricultural systems in Chile. Chil. J. Agric. Res. 2008, 68, 56–68. [Google Scholar] [CrossRef]
- Stigler, S.M. Studies in the History of Probability and Statistics. XXXIV: Napoleonic statistics: The work of Laplace. Biometrika 1975, 62, 503–517. [Google Scholar] [CrossRef]
- Debnath, L.; Basu, K. A short history of probability theory and its applications. Int. J. Math. Educ. Sci. Technol. 2015, 46, 13–39. [Google Scholar] [CrossRef]
- Mazzonia, E.; Vazquez, M. Desertification in Patagonia. Dev. Earth Surf. Process. 2009, 13, 351–377. [Google Scholar] [CrossRef]
- Meinander, O.; Dagsson-Waldhauserova, P.; Amosov, P.; Aseyeva, E.; Atkins, C.; Baklanov, A.; Vukovic Vimic, A. Newly identified climatically and environmentally significant high latitude dust sources. Atmos. Chem. Phys. Discuss. 2021, 22, 11889–11930. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, Y.; Wu, C.; Jardim, A.M.D.R.F.; Fang, M.; Yao, L.; Tang, X. Drought-induced stress on rainfed and irrigated agriculture: Insights from multi-source satellite-derived ecological indicators. Agric. Water Manag. 2025, 307, 109249. [Google Scholar] [CrossRef]
- Birrell, J.H.; Frakes, J.I.; Shah, A.A.; Woods, H.A. Mechanisms underlying thermal breadth differ by species in insects from adjacent but thermally distinct streams—A test of the climate variability hypothesis. J. Therm. Biol. 2023, 112, 103435. [Google Scholar] [CrossRef]
- Chevin, L.M.; Hoffmann, A.A. Evolution of phenotypic plasticity in extreme environments. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160138. [Google Scholar] [CrossRef] [PubMed]
- Wadia, K.D.R.; Butler, D.R. Relationships between temperature and latent periods of rust and leaf-spot diseases of groundnut. Plant Pathol. 1994, 43, 121–129. [Google Scholar] [CrossRef]
- Ciliberti, N.; Fermaud, M.; Roudet, J.; Rossi, V. Environmental conditions affect Botrytis cinerea infection of mature grape berries more than the strain or transposon genotype. Phytopathology 2015, 105, 1090–1096. [Google Scholar] [CrossRef] [PubMed]
- Beniston, M. The 2003 heat wave in Europe: A shape of things to come? An analysis based on Swiss climatological data and model simulations. Geophys. Res. Lett. 2004, 31, L02202. [Google Scholar] [CrossRef]
- García-Herrera, R.; Díaz, J.; Trigo, R.M.; Luterbacher, J.; Fischer, E.M. A review of the European summer heat wave of 2003. Crit. Rev. Environ. Sci. Technol. 2010, 40, 267–306. [Google Scholar] [CrossRef]
- Stott, P.A.; Stone, D.A.; Allen, M.R. Human contribution to the European heatwave of 2003. Nature 2004, 432, 610–614. [Google Scholar] [CrossRef] [PubMed]
- Matsueda, M. Predictability of Euro-Russian blocking in summer of 2010. Geophys. Res. Lett. 2011, 38, L06801. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Fasullo, J.T. Climate extremes and climate change: The Russian heat wave and other climate extremes of 2010. J. Geophys. Res. Atmos. 2012, 117, D17103. [Google Scholar] [CrossRef]
- Hoerling, M.; Kumar, A.; Dole, R.; Nielsen-Gammon, J.W.; Eischeid, J.; Perlwitz, J.; Chen, M. Anatomy of an extreme event. J. Clim. 2013, 26, 2811–2832. [Google Scholar] [CrossRef]
- Christidis, N.; Jones, G.S.; Stott, P.A. Dramatically increasing chance of extremely hot summers since the 2003 European heatwave. Nat. Clim. Change 2015, 5, 46–50. [Google Scholar] [CrossRef]
- Costa, R.L.; de Mello Baptista, G.M.; Gomes, H.B.; dos Santos Silva, F.D.; da Rocha Júnior, R.L.; de Araújo Salvador, M.; Herdies, D.L. Analysis of climate extremes indices over northeast Brazil from 1961 to 2014. Weather Clim. Extrem. 2020, 28, 100254. [Google Scholar] [CrossRef]
- Lovino, M.A.; Müller, O.V.; Berbery, E.H.; Müller, G.V. How have daily climate extremes changed in the recent past over northeastern Argentina? Glob. Planet. Change 2018, 168, 78–97. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The central Chile mega drought (2010–2018): A climate dynamics perspective. Int. J. Climatol. 2020, 40, 421–439. [Google Scholar] [CrossRef]
- Gulizia, C.N.; Raggio, G.A.; Camilloni, I.A.; Saurral, R.I. Changes in mean and extreme climate in southern South America under global warming of 1.5 °C, 2 °C, and 3 °C. Theor. Appl. Climatol. 2022, 150, 787–803. [Google Scholar] [CrossRef]
- Meseguer-Ruiz, O.; Ponce-Philimon, P.I.; Quispe-Jofré, A.S.; Guijarro, J.A.; Sarricolea, P. Spatial behaviour of daily observed extreme temperatures in Northern Chile (1966–2015): Data quality, warming trends, and its orographic and latitudinal effects. Stoch. Environ. Res. Risk Assess. 2018, 32, 3503–3523. [Google Scholar] [CrossRef]
- Mutz, S.G.; Scherrer, S.; Muceniece, I.; Ehlers, T.A. Twenty-first century regional temperature response in Chile based on empirical-statistical downscaling. Clim. Dyn. 2021, 56, 2881–2894. [Google Scholar] [CrossRef]
- Whan, K.; Zscheischler, J.; Orth, R.; Shongwe, M.; Rahimi, M.; Asare, E.O.; Seneviratne, S.I. Impact of soil moisture on extreme maximum temperatures in Europe. Weather Clim. Extrem. 2015, 9, 57–67. [Google Scholar] [CrossRef]
- Meehl, G.A.; Tebaldi, C.; Adams-Smith, D. US daily temperature records past, present, and future. Proc. Natl. Acad. Sci. USA 2016, 113, 13977–13982. [Google Scholar] [CrossRef]
- Huth, R.; Pokorná, L. Parametric versus non-parametric estimates of climatic trends. Theor. Appl. Climatol. 2004, 77, 107–112. [Google Scholar] [CrossRef]
- Schumacher, D.; Singh, J.; Hauser, M.; Fischer, E.; Seneviratne, S. Why climate models underestimate the exacerbated warming in Western Europe. Nat. Commun. 2023, 14, 6803. [Google Scholar] [CrossRef]
- Zhan, J.; McDonald, B.A. Thermal adaptation in the fungal pathogen Mycosphaerella graminicola. Mol. Ecol. 2011, 20, 1689–1701. [Google Scholar] [CrossRef]
- Donatelli, M.; Magarey, R.D.; Bregaglio, S.; Willocquet, L.; Whish, J.P.; Savary, S. Modelling the impacts of pests and diseases on agricultural systems. Agric. Syst. 2017, 155, 213–224. [Google Scholar] [CrossRef]
- Peralta-Maraver, I.; Rezende, E.L. Heat tolerance in ectotherms scales predictably with body size. Nat. Clim. Change 2021, 11, 58–63. [Google Scholar] [CrossRef]
- Gallegos, C.; Chevin, L.M.; Hodgins, K.A.; Monro, K. Predicting adaptation and evolution of plasticity from temporal environmental change. Methods Ecol. Evol. 2024, 16, 84–96. [Google Scholar] [CrossRef]
- González-Domínguez, E.; Caffi, T.; Ciliberti, N.; Rossi, V. A mechanistic model of Botrytis cinerea on grapevines that includes weather, vine growth stage, and the main infection pathways. PLoS ONE 2015, 10, e0140444. [Google Scholar] [CrossRef]
- Latorre, B.A.; Elfar, K.; Ferrada, E.E. Gray mold caused by Botrytis cinerea limits grape production in Chile. Cienc. Investig. Agrar. 2015, 42, 305–330. [Google Scholar] [CrossRef]
- Detka, J.; Jafari, M.; Gomez, M.; Gilbert, G.S. Machine learning vs. empirical models: Estimating leaf wetness patterns in a wildland landscape for plant disease management. Agric. For. Meteorol. 2025, 362, 110392. [Google Scholar] [CrossRef]
- Lichter, A.; Zhou, H.-W.; Vaknin, M.; Dvir, O.; Zutchi, Y.; Kaplunov, T.; Lurie, S. Survival and responses of Botrytis cinerea after exposure to ethanol and heat. J. Phytopathol. 2003, 151, 553–563. [Google Scholar] [CrossRef]
- Yin, X.; Kropff, M.J.; McLaren, G.; Visperas, R.M. A nonlinear model for crop development as a function of temperature. Agric. For. Meteorol. 1995, 77, 1–16. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A. An equation for modelling the temperature response of plants using only the cardinal temperatures. Ann. Bot. 1999, 84, 607–614. [Google Scholar] [CrossRef]
- Rosenzweig, C.; Elliott, J.; Deryng, D.; Ruane, A.C.; Müller, C.; Arneth, A.; Jones, J.W. Assessing agricultural risks of climate change in the 21st century in a global gridded crop model intercomparison. Proc. Natl. Acad. Sci. USA 2014, 111, 3268–3273. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.; Torres, R.; Campillay-Llanos, W.; Guevara-Morales, F. Applications of proportional calculus and a non-Newtonian logistic growth model. Proyecciones 2020, 39, 1471–1513. [Google Scholar] [CrossRef]
- Campillay-Llanos, W.; Guevara, F.; Pinto, M.; Torres, R. Differential and integral proportional calculus: How to find a primitive for f(x) = (1/2) * π * e^(-(1/2) * x^2). Int. J. Math. Educ. Sci. Technol. 2021, 52, 463–476. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| City | Slope (a) | Coef. (b) | 2024 | 2025 | 2026 | Prob (%) |
|---|---|---|---|---|---|---|
| Santiago | 0.033913 | −33.32 | 35.32 ± 1.30 | 35.35 ± 1.30 | 35.39 ± 1.30 | 70 |
| Talca | 0.059391 | −83.92 | 36.29 ± 1.20 | 36.35 ± 1.20 | 36.41 ± 1.20 | 75 |
| Chillán | 0.14952 | −265.58 | 37 ± 2.00 | 37.19 ± 2.00 | 37.34 ± 2.00 | 87 |
| Los Ángeles | 0.21096 | −388.54 | 38.44 ± 2.80 | 38.65 ± 2.80 | 38.86 ± 2.80 | 83 |
| City | Slope (a) | Coef. (b) | 2024 | 2025 | 2026 | Prob(%) |
|---|---|---|---|---|---|---|
| Santiago | 0.10004 | −166.87 | 35.6 ± 1.10 | 35.7 ± 1.10 | 35.8 ± 1.10 | 83 |
| Talca | 0.069304 | −104.02 | 36.2 ± 1.70 | 36.31 ± 1.70 | 36.38 ± 1.70 | 83 |
| Chillán | 0.13974 | −245.50 | 37.33 ± 2.30 | 37.47 ± 2.30 | 37.61 ± 2.30 | 75 |
| Los Ángeles | 0.13161 | −228.06 | 38.31 ± 2.84 | 38.45 ± 2.84 | 38.58 ± 2.84 | 83 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campillay-Llanos, W.; Ortega-Farías, S.; González-Colville, P.; Díaz, G.A.; López-Flores, M.M.; López-Olivari, R. Modeling the Effects of Extreme Temperatures on the Infection Rate of Botrytis cinerea Using Historical Climate Data (1951–2023) of Central Chile. Agronomy 2025, 15, 608. https://doi.org/10.3390/agronomy15030608
Campillay-Llanos W, Ortega-Farías S, González-Colville P, Díaz GA, López-Flores MM, López-Olivari R. Modeling the Effects of Extreme Temperatures on the Infection Rate of Botrytis cinerea Using Historical Climate Data (1951–2023) of Central Chile. Agronomy. 2025; 15(3):608. https://doi.org/10.3390/agronomy15030608
Chicago/Turabian StyleCampillay-Llanos, William, Samuel Ortega-Farías, Patricio González-Colville, Gonzalo A. Díaz, Marlon M. López-Flores, and Rafael López-Olivari. 2025. "Modeling the Effects of Extreme Temperatures on the Infection Rate of Botrytis cinerea Using Historical Climate Data (1951–2023) of Central Chile" Agronomy 15, no. 3: 608. https://doi.org/10.3390/agronomy15030608
APA StyleCampillay-Llanos, W., Ortega-Farías, S., González-Colville, P., Díaz, G. A., López-Flores, M. M., & López-Olivari, R. (2025). Modeling the Effects of Extreme Temperatures on the Infection Rate of Botrytis cinerea Using Historical Climate Data (1951–2023) of Central Chile. Agronomy, 15(3), 608. https://doi.org/10.3390/agronomy15030608