
Identification of Key Genes Associated with Lycopene Accumulation in Red Carrot Taproots Based on WGCNA



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Phenotype Evaluation
2.3. RNA Isolation and Transcriptome Sequencing
2.4. Clustering Analysis
2.5. Functional Enrichment of Module Genes
2.6. Construction of Gene Co-Expression Networks
2.7. Quantitative Real-Time PCR Validation
3. Results
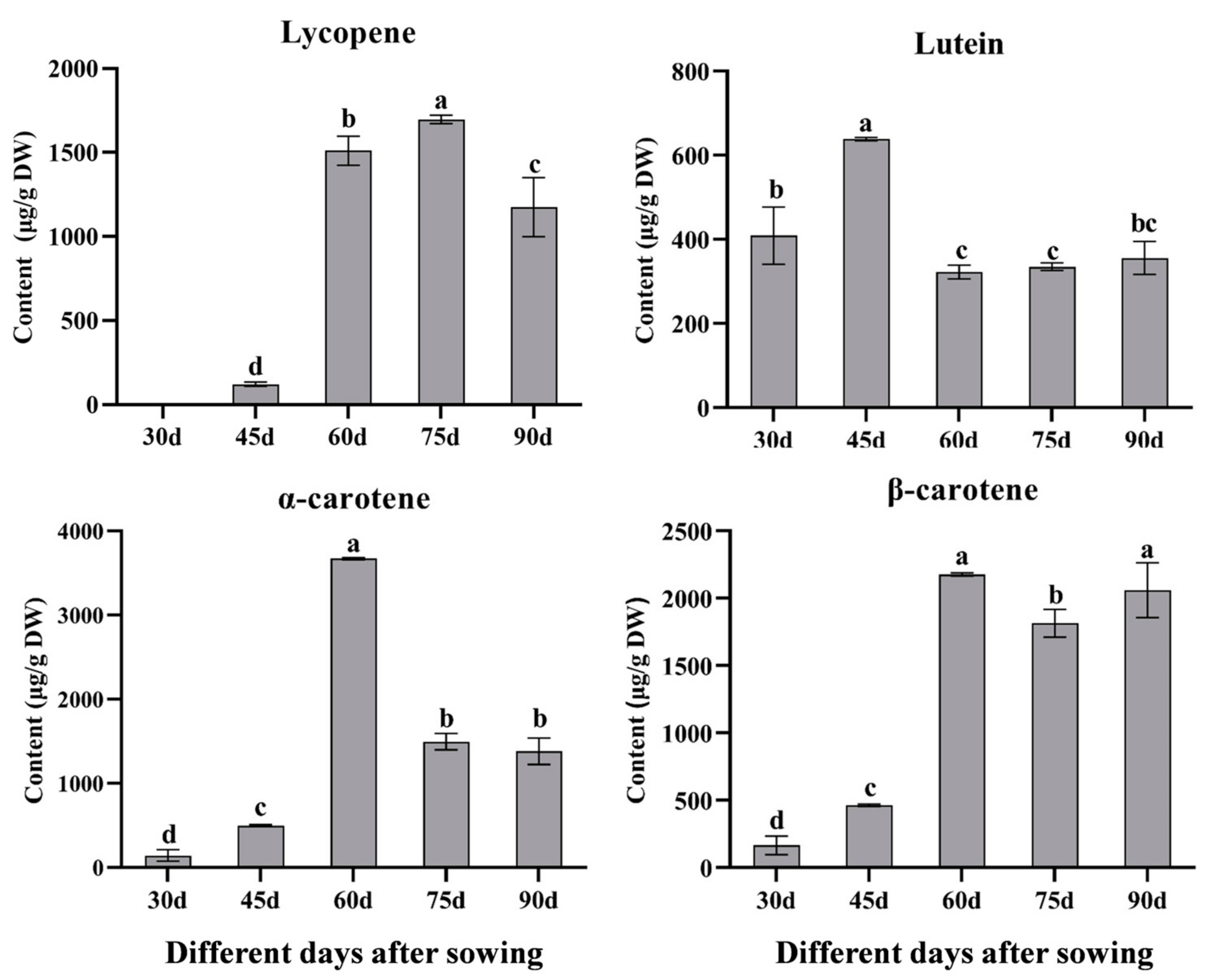
3.1. Dynamic Changes of Carotenoid Content at Different Developmental Stages in Red Carrot
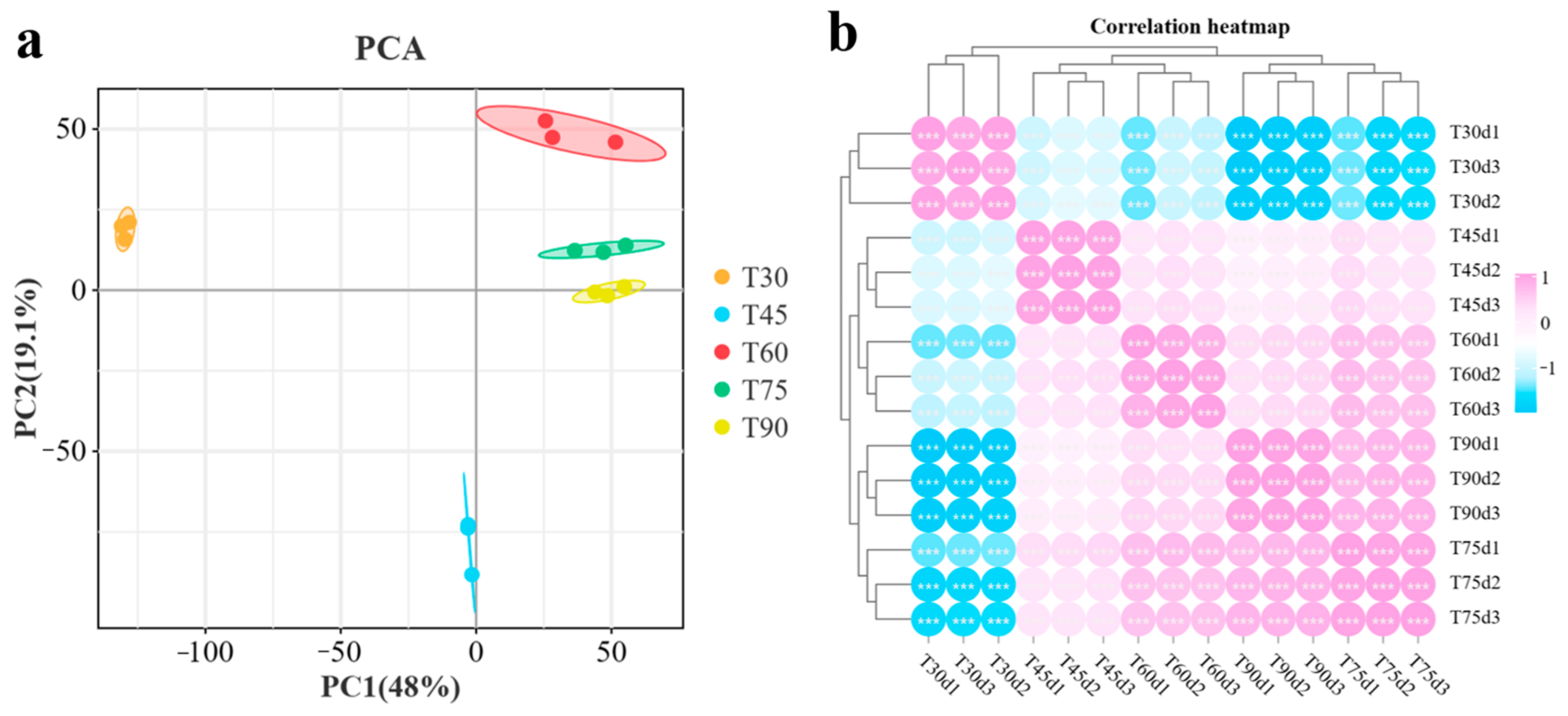
3.2. Transcriptome Analysis
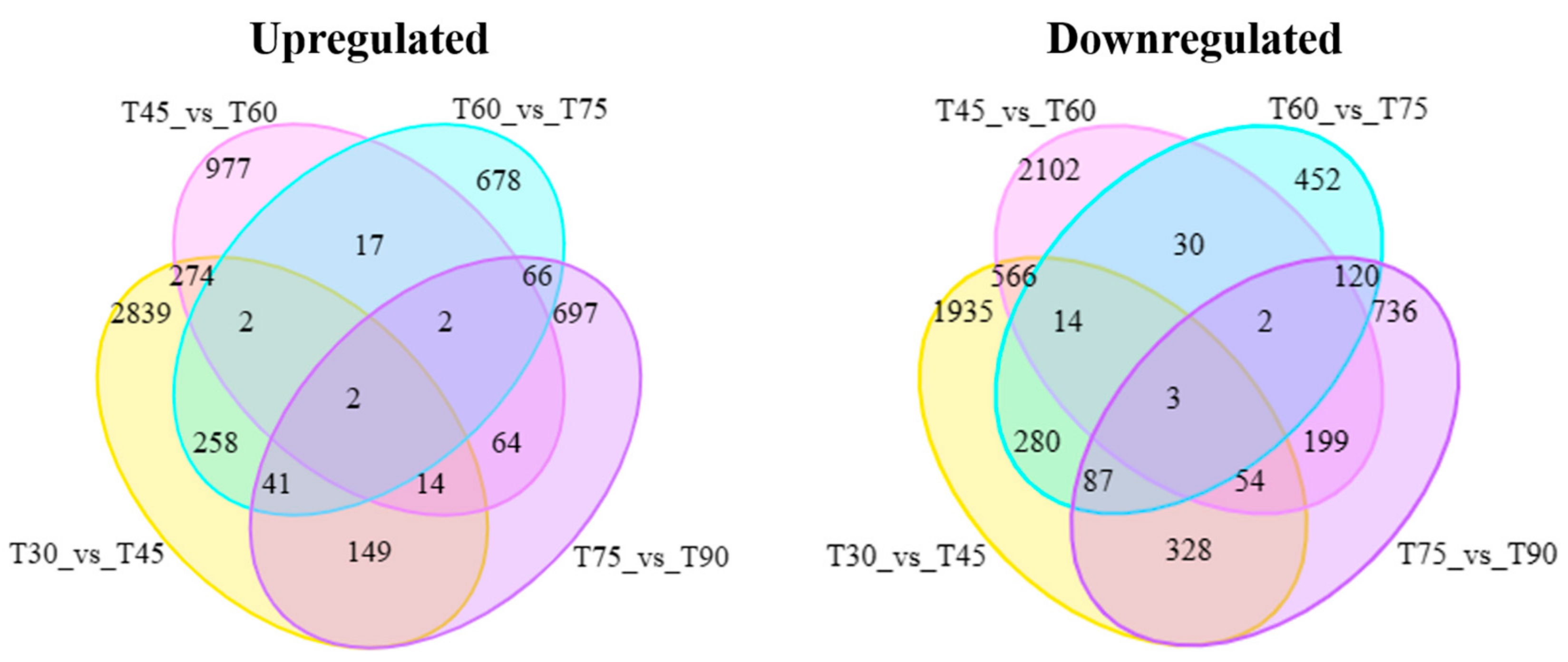
3.3. Screening of DEGs for Lycopene Accumulation
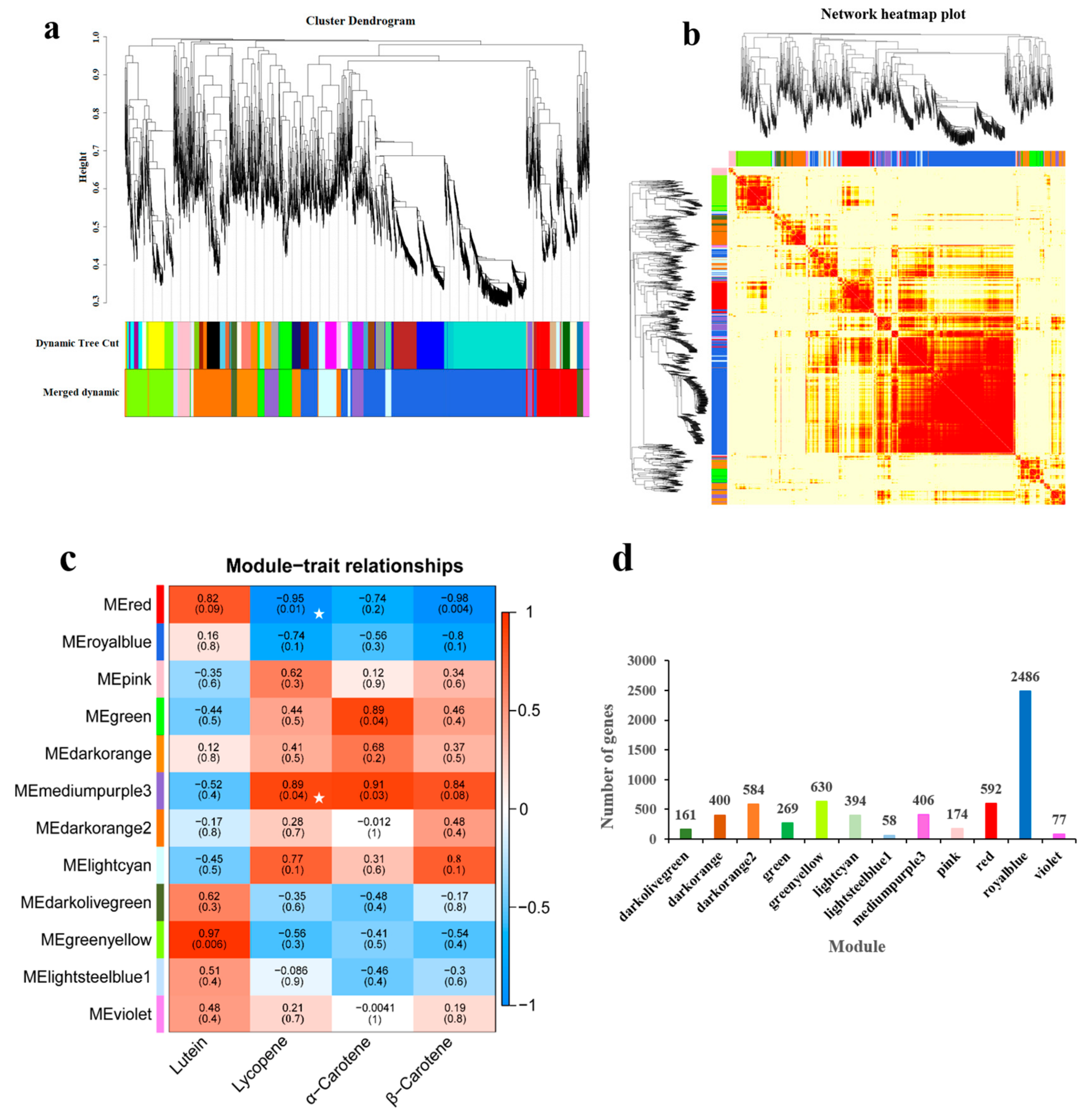
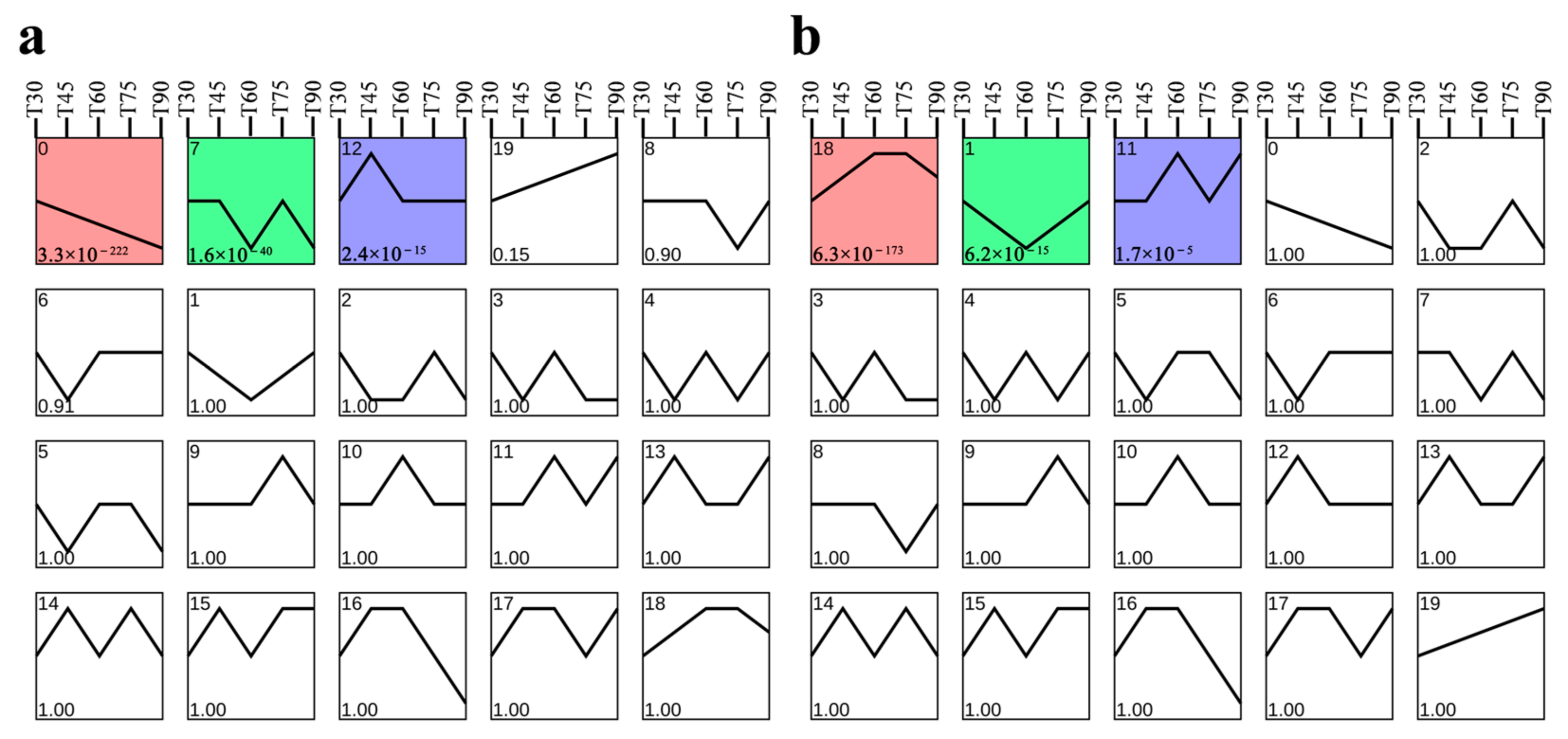
3.4. Weighted Gene Co-Expression Network Analysis
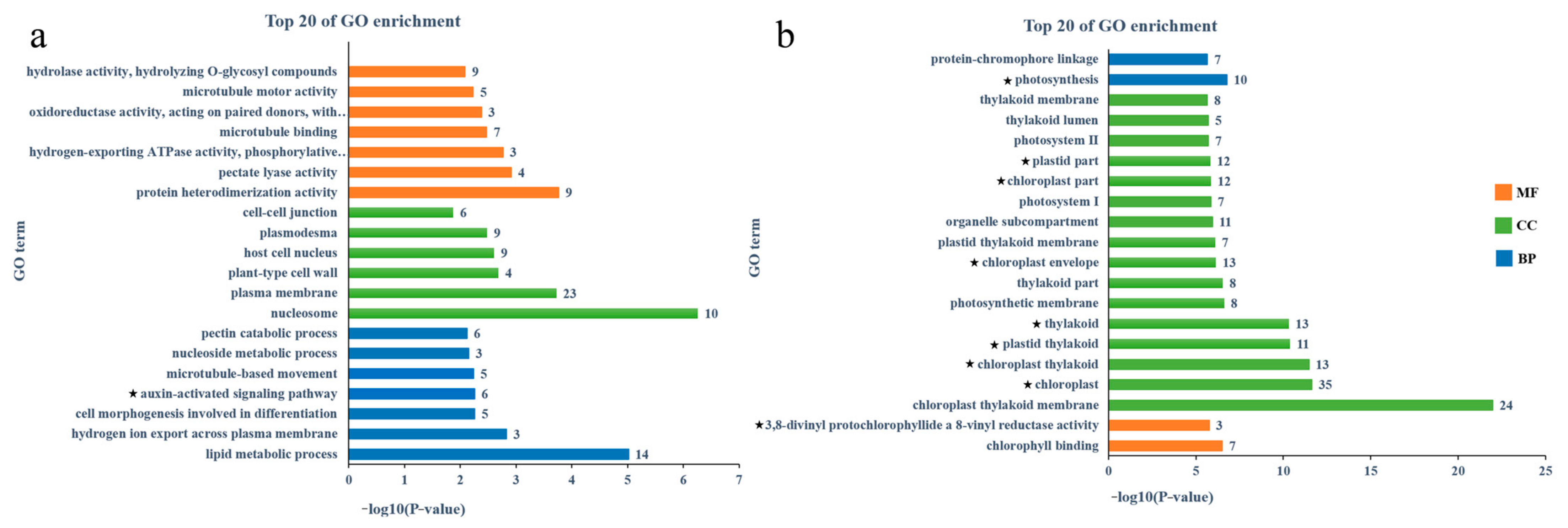
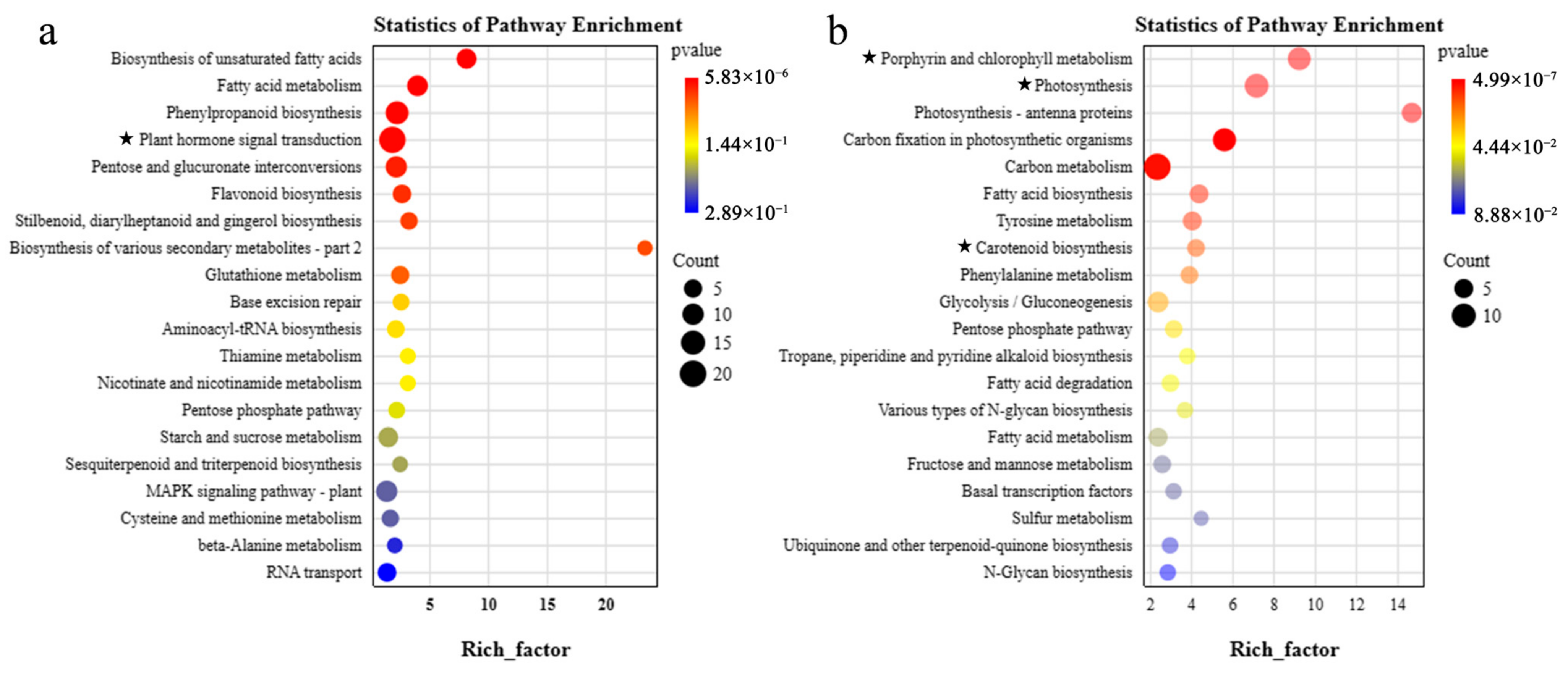
3.5. Screening of Related Genes in Co-Expression Module
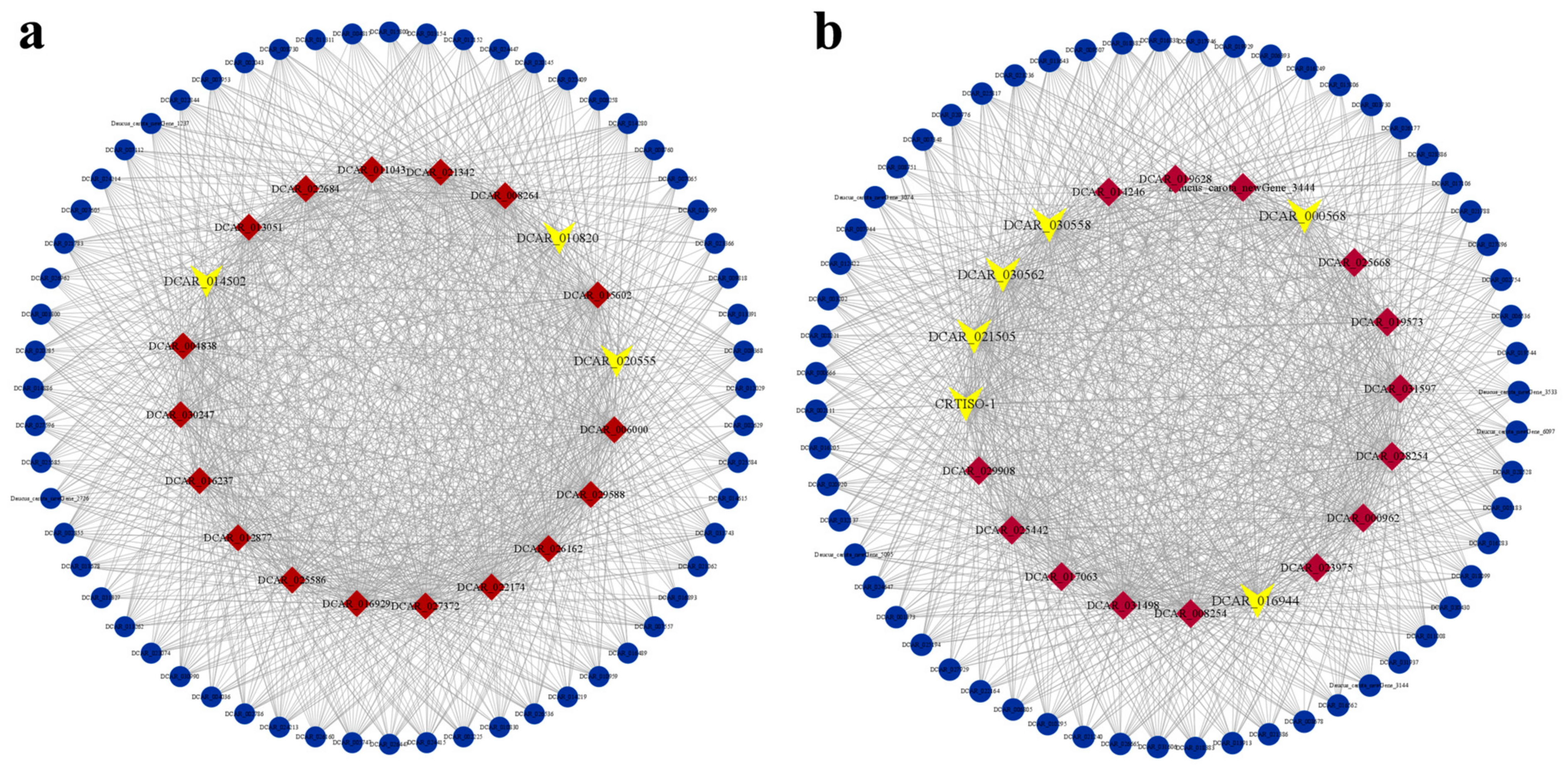
3.6. Hub Gene Screening and Interaction Network Construction
3.7. qRT-PCR of DEGs
4. Discussion
4.1. Excessive Water Supply Influences Lycopene Accumulation
4.2. DcCRTISO-1 Is a Promising Candidate Carotenoid Pathway Gene Involved in Controlling Lycopene Accumulation
4.3. Photosynthesis-Related Genes May Be Involved in Lycopene Accumulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.M.; Wu, R.H.; Wang, L.; Wang, Y.H.; Liu, H.; Xiong, A.S.; Xu, Z.S. Plastid diversity and chromoplast biogenesis in differently coloured carrots: Role of the DcOR3(Leu) gene. Planta 2022, 256, 104. [Google Scholar] [CrossRef] [PubMed]
- Dumas, Y.; Dadomo, M.; Lucca, D.G.; Grolier, P. Effects of environmental factors and agricultural techniques on antioxidantcontent of tomatoes. J. Sci. Food Agric. 2003, 83, 369–382. [Google Scholar] [CrossRef]
- Jing, Y.; Wang, Y.; Zhou, D.; Wang, J.; Li, J.; Sun, J.; Feng, Y.; Xin, F.; Zhang, W. Advances in the synthesis of three typical tetraterpenoids including β-carotene, lycopene and astaxanthin. Biotechnol. Adv. 2022, 61, 108033. [Google Scholar] [CrossRef]
- Ge, B.; Wang, W.; Gao, Y.; Chen, X. Optimization of extraction of lycopene from carrot and determination of its antioxidant activity. J. Food Meas. Charact. 2023, 17, 5497–5505. [Google Scholar] [CrossRef]
- Long, Y.; Paengkoum, S.; Lu, S.; Niu, X.; Thongpea, S.; Taethaisong, N.; Han, Y.; Paengkoum, P. Physicochemical properties, mechanism of action of lycopene and its application in poultry and ruminant production. Front. Vet. Sci. 2024, 11, 1364589. [Google Scholar] [CrossRef]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef]
- Rowles, J.L.; Erdman, J.W. Carotenoids and their role in cancer prevention. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158613. [Google Scholar] [CrossRef]
- Rowles, J.L.; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose-response meta-analysis. Prostate Cancer Prostatic Dis. 2018, 21, 319–336. [Google Scholar] [CrossRef]
- Bae, J.M. Reinterpretation of the results of a pooled analysis of dietary carotenoid intake and breast cancer risk by using the interval collapsing method. Epidemiol. Health 2016, 38, e2016024. [Google Scholar] [CrossRef]
- Kulawik, A.; Cielecka-Piontek, J.; Zalewski, P. The importance of antioxidant activity for the health-promoting effect of lycopene. Nutrients 2023, 15, 3821. [Google Scholar] [CrossRef]
- Park, H.; Kreunen, S.S.; Cuttriss, A.J.; DellaPenna, D.; Pogson, B.J. Identification of the carotenoid isomerase provides insight into carotenoid biosynthesis, prolamellar body formation, and photomorphogenesis. Plant Cell 2002, 14, 321–323. [Google Scholar] [CrossRef] [PubMed]
- Lois, L.M.; Rodríguez-Concepción, M.; Gallego, F.; Campos, N.; Boronat, A. Carotenoid biosynthesis during tomato fruit development: Regulatory role of 1-deoxy-D-xylulose 5-phosphate synthase. Plant J. 2001, 22, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Guan, H.; Dai, Z.; Guo, J.; Shen, Y.; Cui, G.; Gao, W.; Huang, L. Functional analysis of the isopentenyl diphosphate isomerase of Salvia miltiorrhiza via color complementation and RNA interference. Molecules 2015, 20, 20206–20218. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development (Evidence for tissue-specific eene expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef]
- Giuliano, G.; Bartley, G.E.; Scolnik, P.A. Regulation of carotenoid biosynthesis during tomato development. Plant Cell 1993, 5, 379–387. [Google Scholar]
- Dahan-Meir, T.; Filler-Hayut, S.; Melamed-Bessudo, C.; Bocobza, S.; Czosnek, H.; Aharoni, A.; Levy, A.A. Efficient in planta gene targeting in tomato using geminiviral replicons and the CRISPR/Cas9 system. Plant J. 2018, 95, 5–16. [Google Scholar] [CrossRef]
- Costa, M.G.C.; Moreira, C.D.; Melton, J.R.; Otoni, W.C.; Moore, G.A. Characterization and developmental expression of genes encoding the early carotenoid biosynthetic enzymes in Citrus paradisi Macf. Mol. Biol. Rep. 2012, 39, 895–902. [Google Scholar] [CrossRef]
- Fang, X.; Gao, P.; Luan, F.; Liu, S. Identification and characterization roles of phytoene synthase (PSY) genes in watermelon development. Genes 2022, 13, 1189. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A STAY-GREEN protein SlSGR1 regulates lycopene and β-carotene accumulation by interacting directly with SlPSY1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef]
- Zou, L.; Gao, H.; Zhong, Y. Construction of overexpression vector for phytoene dehydrogenase gene and its expression identification in tomato. Hubei Agric. Sci. 2012, 51, 393–395+399. [Google Scholar]
- Ma, J.; Xu, Z.; Tan, G.; Wang, F.; Xiong, A. Distinct transcription profile of genes involved in carotenoid biosynthesis among six different color carrot (Daucus carota L.) cultivars. Acta Biochim. Biophys. Sin. 2017, 49, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Fantini, E.; Falcone, G.; Frusciante, S.; Giliberto, L.; Giuliano, G. Dissection of tomato lycopene biosynthesis through virus-induced gene silencing. Plant Physiol. 2013, 163, 986–998. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, T.; Ronen, G.; Zamir, D.; Hirschberg, J. Cloning of tangerine from tomato reveals a carotenoid isomerase essential for the production of β-carotene and xanthophylls in plants. Plant Cell 2002, 14, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.C.; Pizarro, L.; Fuentes, P.; Handford, M.; Cifuentes, V.; Stange, C. Levels of lycopene β-cyclase 1 modulate carotenoid gene expression and accumulation in Daucus carota. PLoS ONE 2013, 8, e58144. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, T.; Zhang, R.R.; Khadr, A.; Tian, Y.S.; Xu, Z.S.; Xiong, A.S. Transcript profiling of genes involved in carotenoid biosynthesis among three carrot cultivars with various taproot colors. Protoplasma 2020, 257, 949–963. [Google Scholar] [CrossRef]
- Wan, Q.; Zhang, X.G.; Song, M. Fruit-specific RNAi-mediated Lcy gene enhances content of lycopene in tomatoes silencing. Sheng Wu Gong Cheng Xue Bao 2007, 23, 429–433. [Google Scholar] [CrossRef]
- Sun, L.; Yuan, B.; Zhang, M.; Wang, L.; Cui, M.M.; Wang, Q.; Leng, P. Fruit-specific RNAi-mediated suppression of SlNCED1 increases both lycopene and β-carotene contents in tomato fruit. J. Exp. Bot. 2012, 63, 3097–3108. [Google Scholar] [CrossRef]
- Hashimoto, H.; Uragami, C.; Cogdell, R.J. Carotenoids and Photosynthesis. Subcell. Biochem. 2016, 79, 111–139. [Google Scholar]
- Dall’Osto, L.; Bassi, R.; Ruban, A.V. Photoprotective mechanisms: Carotenoids. In Plastid Biology; Theg, S., Wollman, F.A., Eds.; Springer: New York, NY, USA, 2014; Volume 5, pp. 393–435. [Google Scholar]
- Allorent, G.; Petroutsos, D. Photoreceptor-dependent regulation of photoprotection. Curr. Opin. Plant Biol. 2017, 37, 102–108. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Stange, C. Biosynthesis of carotenoids in carrot: An underground story comes to light. Arch. Biochem. Biophys. 2013, 539, 110–116. [Google Scholar] [CrossRef]
- Coe, K.M.; Ellison, S.; Senalik, D.; Dawson, J.; Simon, P. The influence of the Or. and carotene hydroxylase genes on carotenoid accumulation in orange carrots [Daucus carota (L.)]. Theor. Appl. Genet. 2021, 134, 3351–3362. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Duan, C.; Tian, F.H.; Yao, L.; Lv, J.H.; Jia, C.W.; Li, C.T. Comparative transcriptome and WGCNA reveal key genes involved in lignocellulose degradation in Sarcomyxa edulis. Sci. Rep. 2022, 12, 18379. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, X.; Jia, H. WGCNA analysis of important modules and hub genes of compound probiotics regulating lipid metabolism in heat-stressed broilers. Animals 2022, 12, 2644. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Q.; Li, W.; Zhang, S.; Xi, W. Identification of key genes and regulators associated with carotenoid metabolism in apricot (Prunus armeniaca) fruit using weighted gene coexpression network analysis. BMC Genom. 2019, 20, 876. [Google Scholar] [CrossRef]
- Li, X.; Hou, R.; Li, D.; Wang, L.; Wang, T.; Chen, Q.; Qi, X.; Hou, L.; Li, M. Metabolism and transcriptional analyses reveal the mechanism of sucrose affecting the floral transition in pak choi (Brassica rapa ssp. chinensis Makino). Sci. Hortic. 2024, 328, 112968. [Google Scholar] [CrossRef]
- Liu, N.; Cheng, F.; Zhong, Y.; Guo, X. Comparative transcriptome and coexpression network analysis of carpel quantitative variation in Paeonia rockii. BMC Genom. 2019, 20, 683. [Google Scholar] [CrossRef]
- Schefe, J.H.; Lehmann, K.E.; Buschmann, I.R.; Unger, T.; Funke-Kaiser, H. Quantitative real-time RT-PCR data analysis: Current concepts and the novel “gene expression’s CT difference” formula. J. Mol. Med. 2006, 84, 901–910. [Google Scholar] [CrossRef]
- An, W.; Zhang, Y.; Chang, S.; Wu, Z. Cloning and expression analysis of DcZDS gene in carrot. North. Hortic. 2022, 22, 9–15. [Google Scholar]
- Opara, U.L.; Al-Ani, M.R.; Al-Rahbi, N.M. Effect of fruit ripening stage on physico-chemical properties, nutritional composition and antioxidant components of tomato (Lycopersicum esculentum) cultivars. Food Bioproc. Technol. 2012, 5, 3236–3243. [Google Scholar] [CrossRef]
- Schmidt-Szantner, B.; Égei, M.; Takács, S.; Helyes, L.; Ilahy, R.; Pék, Z. The effect of deficit irrigation in processing tomato for the important industrial parameters. Acta Hortic. 2022, 1351, 25–32. [Google Scholar] [CrossRef]
- Kim, Y.X.; Kwon, M.C.; Lee, S.; Jung, E.S.; Lee, C.H.; Sung, J. Effects of nutrient and water supply during fruit development on metabolite composition in tomato fruits (Solanum lycopersicum L.) grown in magnesium excess soils. Front. Plant Sci. 2020, 11, 562399. [Google Scholar] [CrossRef]
- Takács, S.; Pék, Z.; Csányi, D.; Daood, H.G.; Szuvandzsiev, P.; Palotás, G.; Helyes, L. Influence of water stress levels on the yield and lycopene content of tomato. Water 2020, 12, 2165. [Google Scholar] [CrossRef]
- Wang, C.; Qiao, A.; Fang, X.; Sun, L.; Gao, P.; Davis, A.R.; Liu, S.; Luan, F. Fine mapping of lycopene content and flesh color related gene and development of molecular marker-assisted selection for flesh color in watermelon (Citrullus lanatus). Front. Plant Sci. 2019, 10, 1240. [Google Scholar] [CrossRef]
- Leskovar, D.; Bang, H.; Kolenda, K.; Perkins, P.; Franco, J. Deficit irrigation influences yield and lycopene content of diploid and triploid watermelon. Acta Hort. 2003, 628, 147–151. [Google Scholar] [CrossRef]
- Cazzonelli, C.I.; Pogson, B.J. Source to sink: Regulation of carotenoid biosynthesis in plants. Trends Plant Sci. 2010, 15, 266–274. [Google Scholar] [CrossRef]
- Efremov, G.I.; Dzhos, E.A.; Ashikhmin, A.A.; Kochieva, E.Z.; Shchennikova, A.V. Effect of the carotenoid content and activity of the carotene cis-trans isomerase CRTISO on tomato fruit color. Russ. J. Plant Physiol. 2022, 69, 64. [Google Scholar] [CrossRef]
- Pinheiro, T.T.; Peres, L.E.P.; Purgatto, E.; Latado, R.R.; Maniero, R.A.; Martins, M.M.; Figueira, A. Citrus carotenoid isomerase gene characterization by complementation of the “Micro-Tom” tangerine mutant. Plant Cell Rep. 2019, 38, 623–636. [Google Scholar] [CrossRef]
- Simpson, K.; Cerda, A.; Stange, C. Carotenoid biosynthesis in daucus carota. Subcell. Biochem. 2016, 79, 199–217. [Google Scholar] [PubMed]
- Stange, C.; Fuentes, P.; Handford, M.; Pizarro, L. Daucus carota as a novel model to evaluate the effect of light on carotenogenic gene expression. Biol. Res. 2008, 41, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Arias, D.; Maldonado, J.; Silva, H.; Stange, C. A de novo transcriptome analysis revealed that photomorphogenic genes are required for carotenoid synthesis in the dark-grown carrot taproot. Mol. Genet. Genom. 2020, 295, 1379–1392. [Google Scholar] [CrossRef] [PubMed]
- Iorizzo, M.; Ellison, S.; Senalik, D.; Zeng, P.; Satapoomin, P.; Huang, J.; Bowman, M.; Iovene, M.; Sanseverino, W.; Cavagnaro, P.; et al. A high-quality carrot genome assembly provides new insights into carotenoid accumulation and asterid genome evolution. Nat. Genet. 2016, 48, 657–666. [Google Scholar] [CrossRef]
- Wu, Z.; Xu, H.; Yang, X.; Li, L.; Luo, D.; Liu, Z.; Jia, L. Transcriptome and re-sequencing analyses reveal photosynthesis-related genes involvement in lutein accumulation in yellow taproot mutants of carrot. Agronomy 2022, 12, 1866. [Google Scholar] [CrossRef]
- Wang, Y.G.; Zhang, Y.M.; Wang, Y.H.; Zhang, K.; Ma, J.; Hang, J.X.; Su, Y.T.; Tan, S.S.; Liu, H.; Xiong, A.S.; et al. The Y locus encodes a REPRESSOR OF PHOTOSYNTHETIC GENES protein that represses carotenoid biosynthesis via interaction with APRR2 in carrot. Plant Cell 2024, 36, 2798–2817. [Google Scholar] [CrossRef]
- Hung, C.Y.; Sun, Y.H.; Chen, J.J.; Darlington, D.E.; Williams, A.L.; Burkey, K.O.; Xie, J.H. Identification of a Mg-protoporphyrin IX monomethyl ester cyclase homologue, EaZIP, differentially expressed in variegated Epipremnum aureum ‘Golden Pothos’ is achieved through a unique method of comparative study using tissue regenerated plants. J. Exp. Bot. 2010, 61, 1483–1493. [Google Scholar] [CrossRef]
- Bang, W.Y.; Jeong, I.S.; Kim, D.W.; Im, C.H.; Ji, C.; Hwang, S.M.; Kim, S.W.; Son, Y.S.; Jeong, J.; Shiina, T.; et al. Role of Arabidopsis CHL27 protein for photosynthesis, chloroplast development and gene expression profiling. Plant Cell Physiol. 2008, 49, 1350–1363. [Google Scholar] [CrossRef]
- Lintala, M.; Allahverdiyeva, Y.; Kangasjärvi, S.; Lehtimäki, N.; Keränen, M.; Rintamäki, E.; Aro, E.M.; Mulo, P. Comparative analysis of leaf-type ferredoxin-NADP oxidoreductase isoforms in Arabidopsis thaliana. Plant J. 2009, 57, 1103–1115. [Google Scholar] [CrossRef]
- Sakata, S.; Mizusawa, N.; Kubota-Kawai, H.; Sakurai, I.; Wada, H. Psb28 is involved in recovery of photosystem II at high temperature in Synechocystis sp. PCC 6803. Biochim. Biophys. Acta 2013, 1827, 50–59. [Google Scholar] [CrossRef]
- Zhao, Y.; Deng, L.; Last, R.L.; Hua, W.; Liu, J. Psb28 protein is indispensable for stable accumulation of PSII core complexes in Arabidopsis. Plant J. 2024, 119, 1226–1238. [Google Scholar] [CrossRef]
- Tachibana, R.; Yamagami, A.; Miyagi, S.; Nakazawa-Miklasevica, M.; Matsui, M.; Sakuta, M.; Tanaka, R.; Asami, T.; Nakano, T. BRZ-INSENSITIVE-PALE GREEN 1 is encoded by chlorophyll biosynthesis enzyme gene that functions in the downstream of brassinosteroid signaling. Biosci. Biotechnol. Biochem. 2022, 86, 1041–1048. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
An, X.; Zhang, Y.; Zhang, H.; Zhang, X.; Li, L.; Luo, D.; Wu, Z. Identification of Key Genes Associated with Lycopene Accumulation in Red Carrot Taproots Based on WGCNA. Agronomy 2025, 15, 530. https://doi.org/10.3390/agronomy15030530
An X, Zhang Y, Zhang H, Zhang X, Li L, Luo D, Wu Z. Identification of Key Genes Associated with Lycopene Accumulation in Red Carrot Taproots Based on WGCNA. Agronomy. 2025; 15(3):530. https://doi.org/10.3390/agronomy15030530
Chicago/Turabian StyleAn, Xiaoning, Yanan Zhang, Hang Zhang, Xinyu Zhang, Lixia Li, Dan Luo, and Zhe Wu. 2025. "Identification of Key Genes Associated with Lycopene Accumulation in Red Carrot Taproots Based on WGCNA" Agronomy 15, no. 3: 530. https://doi.org/10.3390/agronomy15030530
APA StyleAn, X., Zhang, Y., Zhang, H., Zhang, X., Li, L., Luo, D., & Wu, Z. (2025). Identification of Key Genes Associated with Lycopene Accumulation in Red Carrot Taproots Based on WGCNA. Agronomy, 15(3), 530. https://doi.org/10.3390/agronomy15030530