

Impact of Low Light on Photosynthetic Characteristics, Antioxidant Activity, and Yield of Brassica napus L.

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Sampling and Measurements
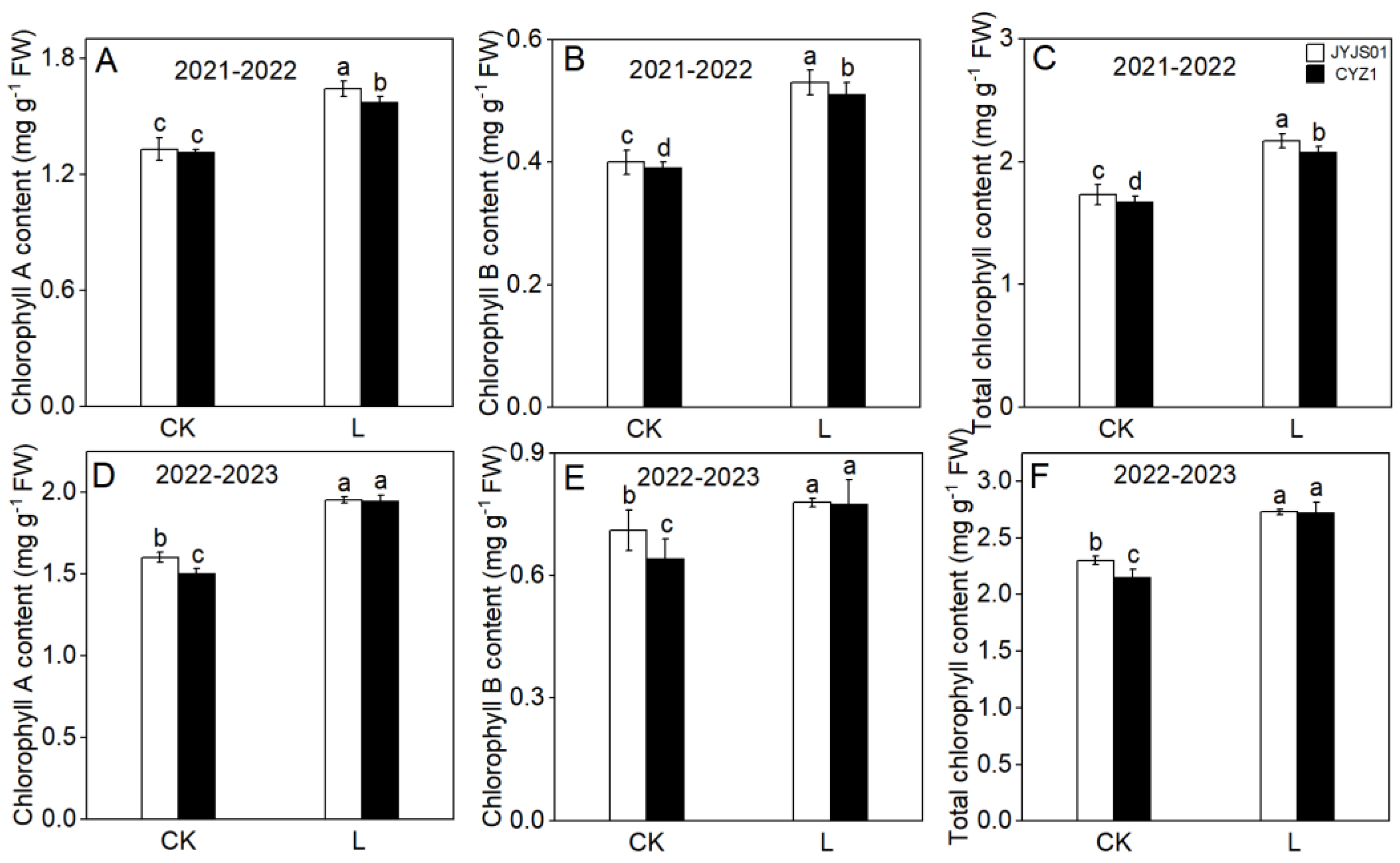
2.3.1. Chlorophyll Assessment
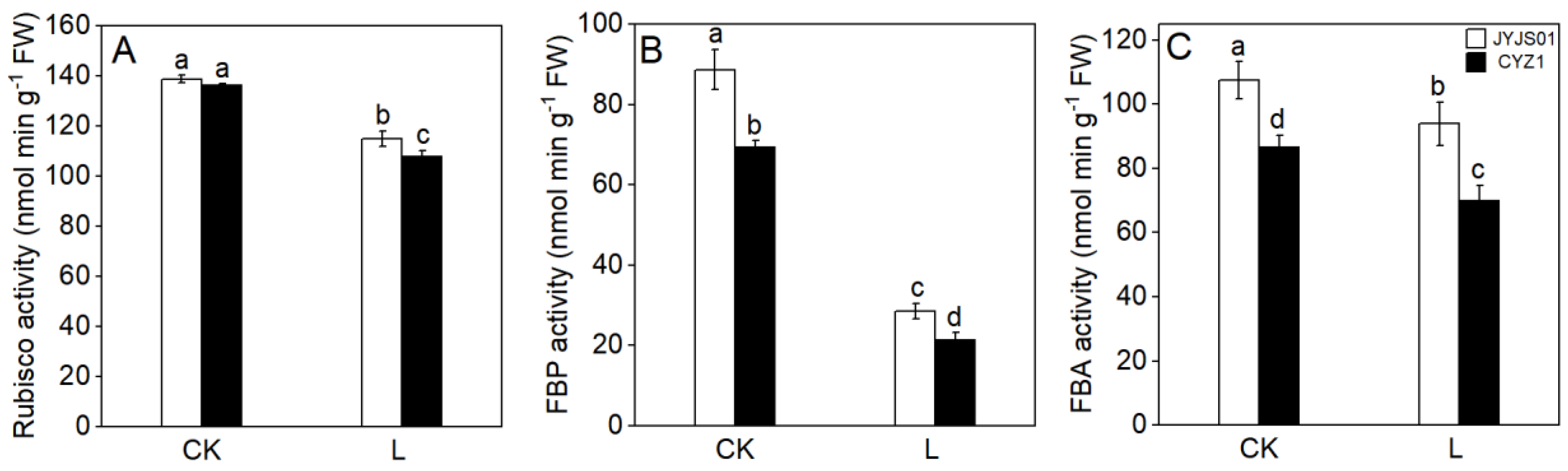
2.3.2. Activity of Key Photosynthetic and Antioxidant Enzymes
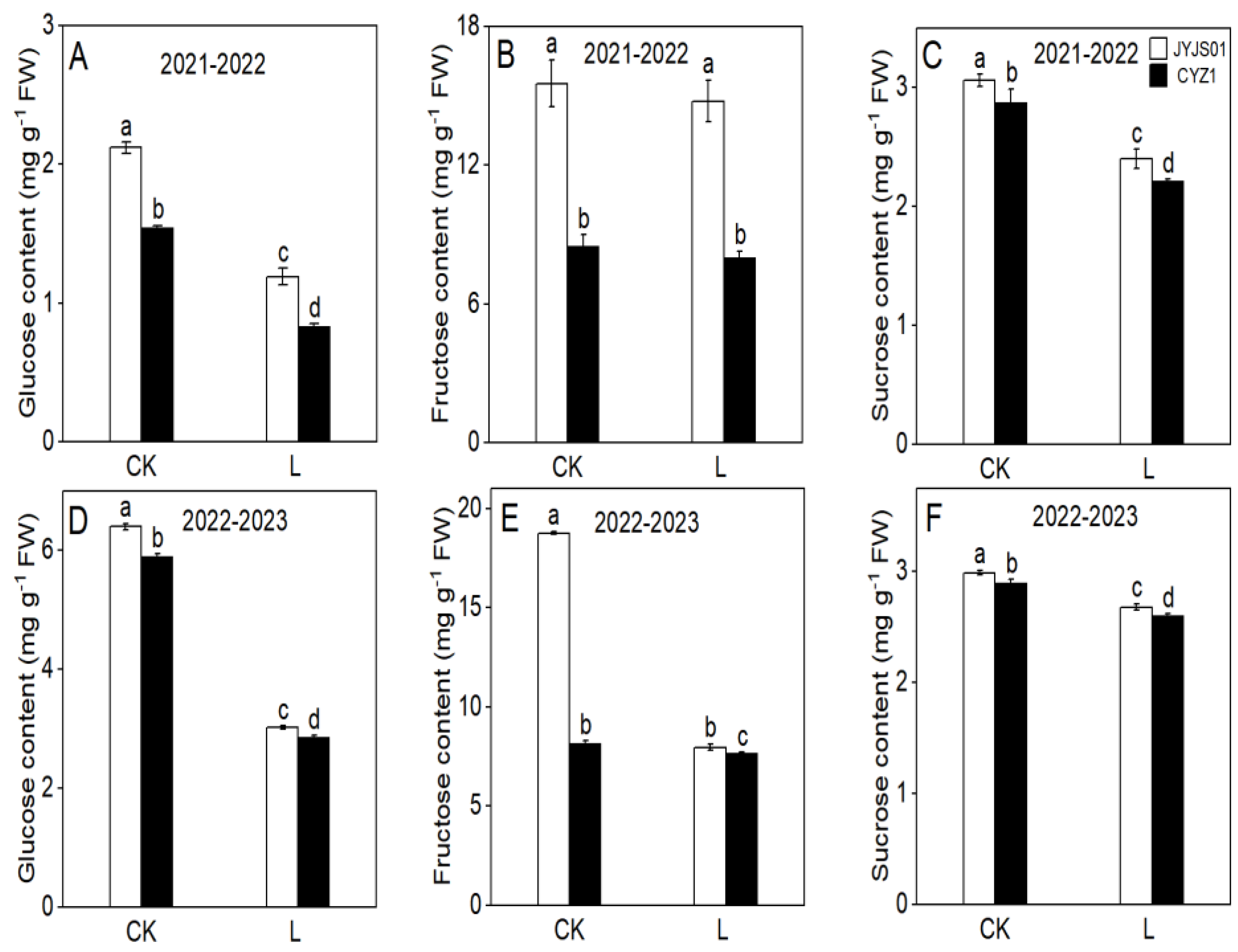
2.3.3. Carbohydrate Content
2.3.4. Amino Acid and Malondialdehyde Content
2.3.5. ATP Content
2.4. Statistics Analysis
3. Results
3.1. Chlorophyll Content
3.2. Photosynthetic Enzymatic Activity
3.3. Carbohydrate Content
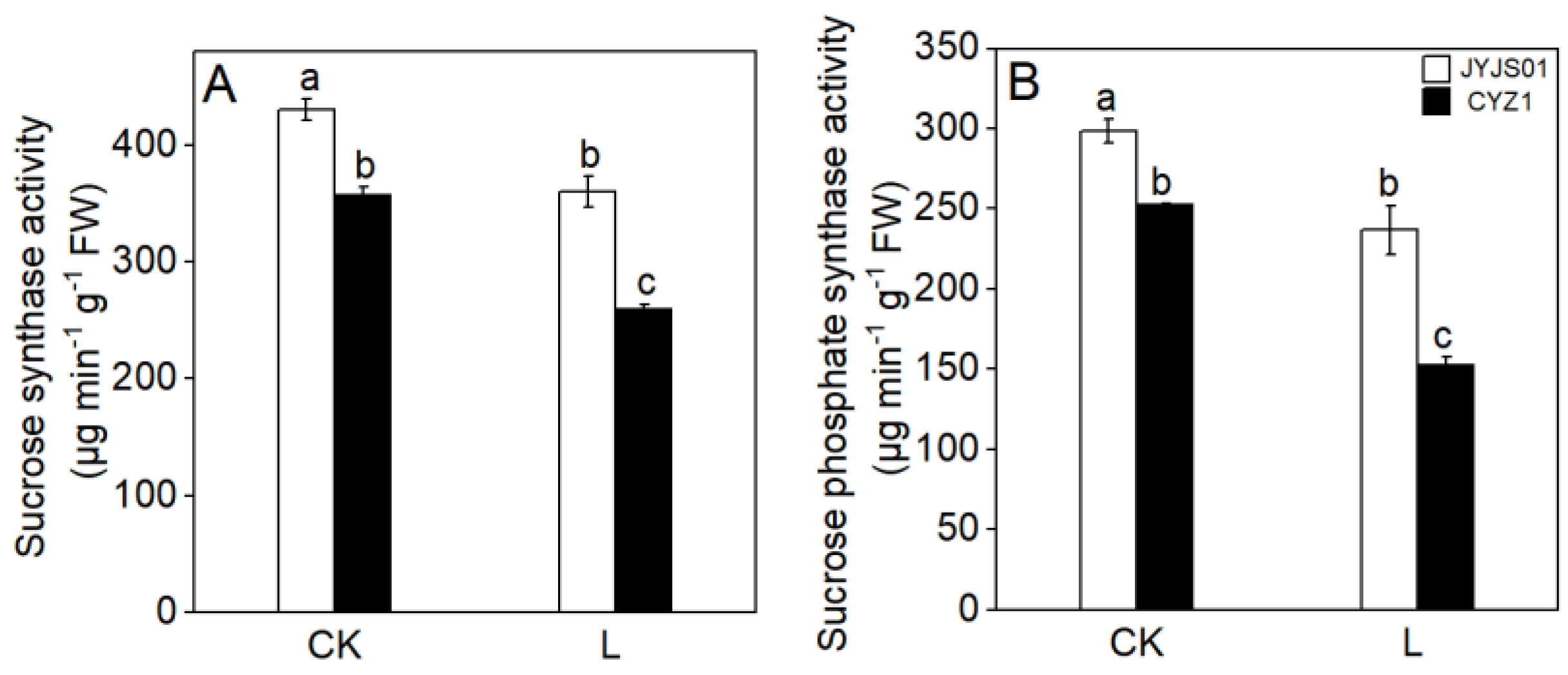
3.4. Sucrose Synthase Activity
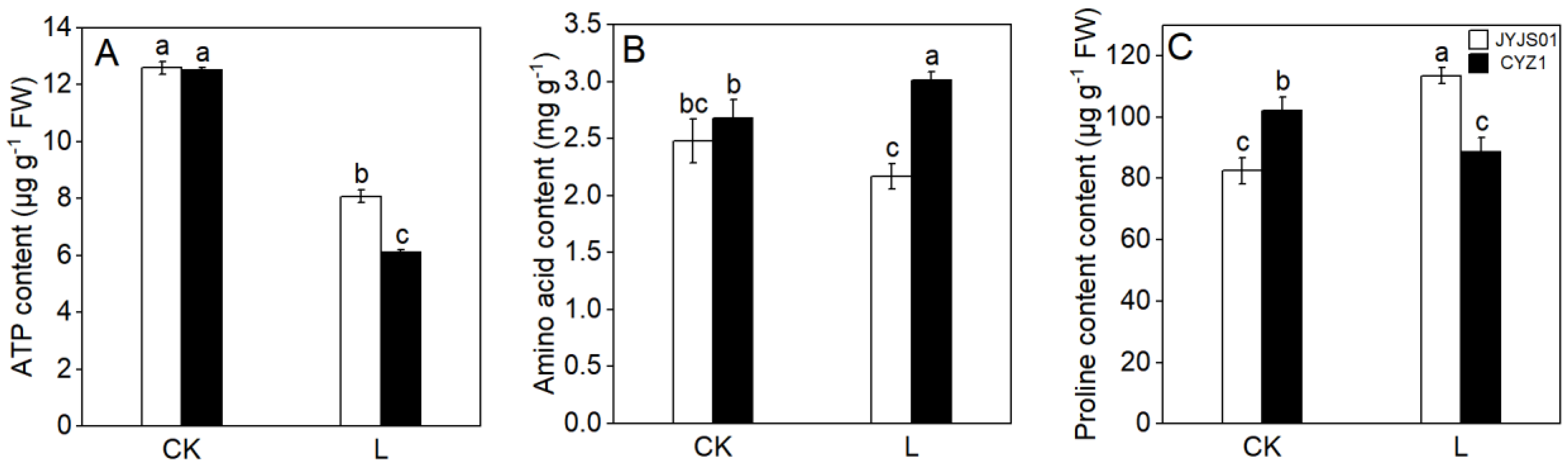
3.5. ATP, Amino Acid, and Proline Content
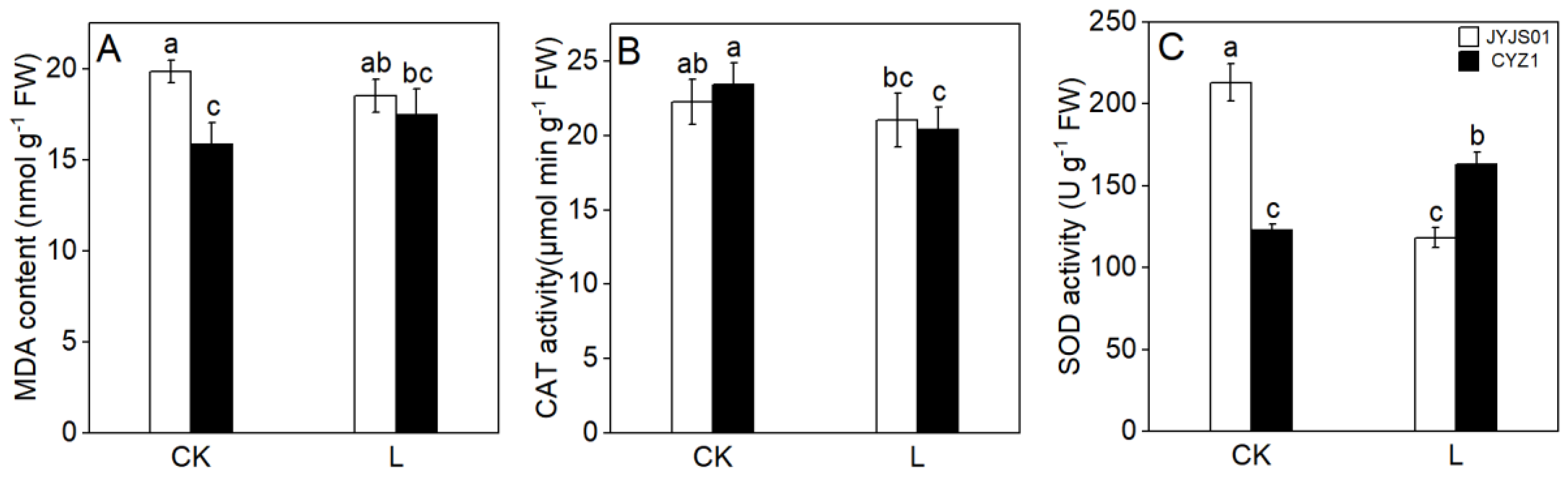
3.6. MDA Content CAT and SOD Activity
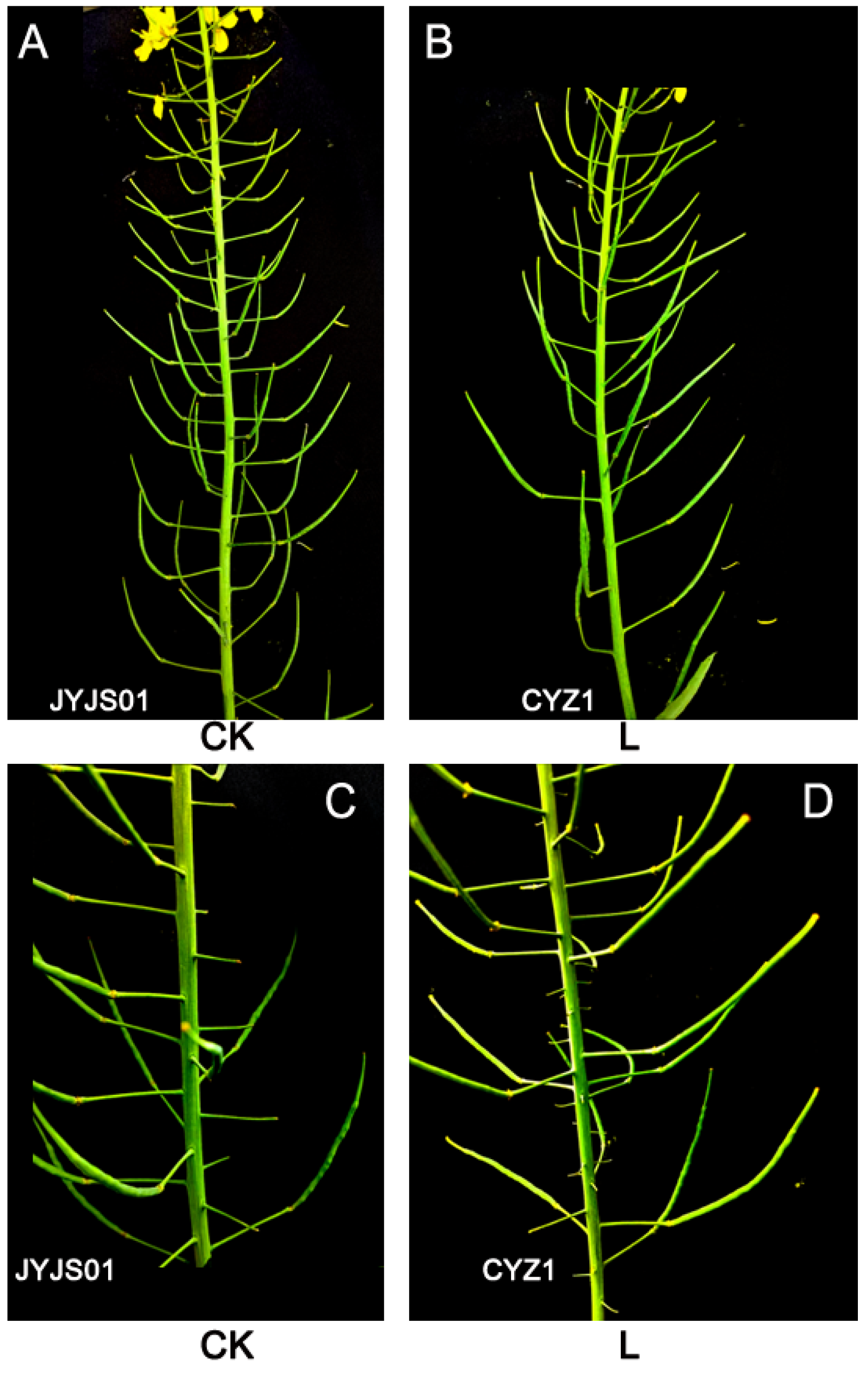
3.7. Yield Composition
3.8. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Al Murad, M.; Razi, K.; Jeong, B.R.; Samy, P.M.A.; Muneer, S. Light emitting diodes (LEDs) as agricultural lighting: Impact and its potential on improving physiology, flowering, and secondary metabolites of crops. Sustainability 2021, 13, 1985. [Google Scholar] [CrossRef]
- Xiao, L.; Shibuya, T.; Kato, K.; Nishiyama, M.; Kanayama, Y. Effects of light quality on plant development and fruit metabolism and their regulation by plant growth regulators in tomato. Sci. Hortic. 2022, 300, 111076. [Google Scholar] [CrossRef]
- Wang, N.; Ji, T.; Liu, X.; Li, Q.; Sairebieli, K.; Wu, P.; Song, H.; Wang, H.; Du, N.; Zheng, P. Defoliation significantly suppressed plant growth under low light conditions in two leguminosae species. Front. Plant Sci. 2022, 12, 777328. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, Ü.; Ntagkas, N.; Siebenkäs, A.; Mäenpää, M.; Matsubara, S.; Pons, T. A meta-analysis of plant responses to light intensity for 70 traits ranging from molecules to whole plant performance. New Phytol. 2019, 223, 1073–1105. [Google Scholar] [CrossRef]
- Paradiso, R.; Proietti, S. Light-quality manipulation to control plant growth and photomorphogenesis in greenhouse horticulture: The state of the art and the opportunities of modern LED systems. J. Plant Growth Regul. 2022, 41, 742–780. [Google Scholar] [CrossRef]
- Garcia-Caparros, P.; De Filippis, L.; Gul, A.; Hasanuzzaman, M.; Ozturk, M.; Altay, V.; Lao, M.T. Oxidative stress and antioxidant metabolism under adverse environmental conditions: A review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Wang, Z.; Hu, Y.; Zhang, S.; Sun, Y. Artificial photosynthesis systems for solar energy conversion and storage: Platforms and their realities. Chem. Soc. Rev. 2022, 51, 6704–6737. [Google Scholar] [CrossRef]
- Perchlik, M.; Tegeder, M. Leaf amino acid supply affects photosynthetic and plant nitrogen use efficiency under nitrogen stress. Plant Physiol. 2018, 178, 174–188. [Google Scholar] [CrossRef]
- Khan, S.; Yu, H.; Li, Q.; Gao, Y.; Sallam, B.N.; Wang, H.; Liu, P.; Jiang, W. Exogenous application of amino acids improves the growth and yield of lettuce by enhancing photosynthetic assimilation and nutrient availability. Agronomy 2019, 9, 266. [Google Scholar] [CrossRef]
- Sharkey, T.D. The end game (s) of photosynthetic carbon metabolism. Plant Physiol. 2024, 195, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Pu, X.; Han, Y. Promotion of carbon dioxide biofixation through metabolic and enzyme engineering. Catalysts 2022, 12, 399. [Google Scholar] [CrossRef]
- Gardeström, P.; Igamberdiev, A.U. The origin of cytosolic ATP in photosynthetic cells. Physiol. Plant. 2016, 157, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Kotilainen, T.; Robson, T.M.; Hernandez, R. Light quality characterization under climate screens and shade nets for controlled-environment agriculture. PLoS ONE 2018, 13, e0199628. [Google Scholar] [CrossRef]
- Andrade, F.H.; Ferreiro, M.A. Reproductive growth of maize, sunflower and soybean at different source levels during grain filling. Field Crops Res. 1996, 48, 155–165. [Google Scholar] [CrossRef]
- Bitterlich, M.; Franken, P.; Graefe, J. Atmospheric drought and low light impede mycorrhizal effects on leaf photosynthesis—A glasshouse study on tomato under naturally fluctuating environmental conditions. Mycorrhiza 2019, 29, 13–28. [Google Scholar] [CrossRef]
- Slattery, R.A.; Ort, D.R. Perspectives on improving light distribution and light use efficiency in crop canopies. Plant Physiol. 2021, 185, 34–48. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; Xie, Y.; Wang, B.; Kuai, J.; Zhou, G. An improvement in oilseed rape (Brassica napus L.) productivity through optimization of rice-straw quantity and plant density. Field Crop. Res. 2021, 273, 108290. [Google Scholar] [CrossRef]
- An, J.; Wei, X.; Huo, H. Transcriptome analysis reveals the accelerated expression of genes related to photosynthesis and chlorophyll biosynthesis contribution to shade-tolerant in Phoebe bournei. BMC Plant Biol. 2022, 22, 268. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Bhuyan, M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Mahmud, J.A.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Zhu, H.; Li, X.; Zhai, W.; Liu, Y.; Gao, Q.; Liu, J.; Ren, L.; Chen, H.; Zhu, Y. Effects of low light on photosynthetic properties, antioxidant enzyme activity, and anthocyanin accumulation in purple pak-choi (Brassica campestris ssp. Chinensis Makino). PLoS ONE 2017, 12, e0179305. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Fu, Y.; Liu, G.; Liu, H. Low light intensity effects on the growth, photosynthetic characteristics, antioxidant capacity, yield and quality of wheat (Triticum aestivum L.) at different growth stages in BLSS. Adv. Space Res. 2014, 53, 1557–1566. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Zhu, Z.H.; Sami, A.; Xu, Q.Q.; Wu, L.L.; Zheng, W.Y.; Chen, Z.P.; Jin, X.Z.; Zhang, H.; Li, Y.; Yu, Y. Effects of seed priming treatments on the germination and development of two rapeseed (Brassica napus L.) varieties under the co-influence of low temperature and drought. PLoS ONE 2021, 16, e0257236. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and carotenoids: Measurement and characterization by UV-VIS spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Javed, H.H.; Hu, Y.; Raza, A.; Alabdallah, N.M.; Asghar, M.A.; Khan, K.S.; Peng, X.; Ghafoor, A.Z.; Ullah, A.; Wu, Y.C. Low light at specific growth stage affects photoassimilates transportation, seed quality and yield in Brassica napus L. J. Agron. Crop Sci. 2024, 210, e12735. [Google Scholar] [CrossRef]
- Zhang, M.X.; Zhao, L.Y.; He, Y.Y.; Hu, J.P.; Hu, G.W.; Zhu, Y.; Khan, A.; Xiong, Y.C.; Zhang, J.L. Potential roles of iron nanomaterials in enhancing growth and nitrogen fixation and modulating rhizomicrobiome in alfalfa (Medicago sativa L.). Bioresour. Technol. 2024, 391, 129987. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qu, W.J. The Experimental Guide for Plant Physiology; Higher Education Press: Beijing, China, 2003. [Google Scholar]
- Chen, H.; Jin, Z.; Huang, R.; He, L.; Tian, W.; Zhao, L.; Zhang, Z. Promotion of growth of alfalfa by Erwinia persicina Cp2 exopolysaccharides under NaCl stress. Agronomy 2023, 13, 2129. [Google Scholar] [CrossRef]
- Cheng, B.; Zhou, M.; Tang, T.; Hassan, M.J.; Zhou, J.; Tan, M.; Li, Z.; Peng, Y. A Trifolium repens flavodoxin-like quinone reductase 1 (TrFQR1) improves plant adaptability to high temperature associated with oxidative homeostasis and lipids remodeling. Plant J. 2023, 115, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Servaites, J.C.; Parry, M.A.; Gutteridge, S.; Keys, A.J. Species variation in the predawn inhibition of ribulose-1, 5-bisphosphate carboxylase/oxygenase. Plant Physiol. 1986, 82, 1161–1163. [Google Scholar] [CrossRef]
- Hu, L.; Liao, W.; Dawuda, M.M.; Yu, J.; Lv, J. Appropriate NH4+: NO3− ratio improves low light tolerance of mini Chinese cabbage seedlings. BMC Plant Biol. 2017, 17, 1–14. [Google Scholar] [CrossRef]
- Seemann, J.R.; Kirschbaum, M.U.; Sharkey, T.D.; Pearcy, R.W. Regulation of ribulose-1, 5-bisphosphate carboxylase activity in Alocasia macrorrhiza in response to step changes in irradiance. Plant Physiol. 1988, 88, 148–152. [Google Scholar] [CrossRef]
- Ernstsen, J.; Woodrow, I.E.; Mott, K.A. Responses of Rubisco activation and deactivation rates to variations in growth-light conditions. Photosyn. Res. 1997, 52, 117–125. [Google Scholar] [CrossRef]
- Sun, J.L.; Sui, X.L.; Huang, H.Y.; Wang, S.H.; Wei, Y.X.; Zhang, Z.X. Low light stress down-regulated Rubisco gene expression and photosynthetic capacity during cucumber (Cucumis sativus L.) leaf development. J. Integr. Agric. 2014, 13, 997–1007. [Google Scholar] [CrossRef]
- Tang, W.; Guo, H.; Baskin, C.C.; Xiong, W.; Yang, C.; Li, Z.; Song, H.; Wang, T.; Yin, J.; Wu, X. Effect of light intensity on morphology, photosynthesis and carbon metabolism of alfalfa (Medicago sativa) seedlings. Plants 2022, 11, 1688. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Cun, Z.; Chen, J.W. Photosynthetic performance and photosynthesis-related gene expression coordinated in a shade-tolerant species Panax notoginseng under nitrogen regimes. BMC Plant Biol. 2020, 20, 273. [Google Scholar] [CrossRef]
- Lazár, D.; Stirbet, A.; Björn, L.O.; Govindjee, G. Light quality, oxygenic photosynthesis and more. Photosynthetica 2022, 60, 25. [Google Scholar] [CrossRef]
- Mu, H.R.; Jiang, D.; Dai, T.B.; Jing, Q.; Cao, W.X. Effect of shading on photosynthesis and chlorophyll fluorescence characters in wheat flag leaves. Sci. Agric. Sin. 2008, 41, 599–606. [Google Scholar] [CrossRef]
- Shao, Q.; Wang, H.; Guo, H.; Zhou, A.; Huang, Y.; Sun, Y.; Li, M. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, S.; He, X.; Li, Y.; Zhang, Y.; Chen, W. Response of total phenols, flavonoids, minerals, and amino acids of four edible fern species to four shading treatments. PeerJ 2020, 8, e8354. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yang, J.; Li, J.; Zhou, X.; Xiao, Y.; Liao, Y.; Tang, J.; Dong, F.; Zeng, L. Effects of temperature and light on quality-related metabolites in tea [Camellia sinensis (L.) Kuntze] leaves. Food Res. Int. 2022, 161, 111882. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Yu, M.; Xiao, H.; Wang, H.; Xie, H.; Zeng, X. Influences of silicon on content of soluble sugars, amino acids in Turf-grasses under shading stress. J. Huazhong Agric. Univ. 2010, 29, 317–320. [Google Scholar] [CrossRef]
- Zhu, X.; Galili, G. Lysine metabolism is concurrently regulated by synthesis and catabolism in both reproductive and vegetative tissues. Plant Physiol. 2004, 135, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Galili, G.; Avin-Wittenberg, T.; Angelovici, R.; Fernie, A.R. The role of photosynthesis and amino acid metabolism in the energy status during seed development. Front. Plant Sci. 2014, 5, 447. [Google Scholar] [CrossRef]
- Less, H.; Galili, G. Principal transcriptional programs regulating plant amino acid metabolism in response to abiotic stresses. Plant Physiol. 2008, 147, 316–330. [Google Scholar] [CrossRef]
- Feller, U.; Vaseva, I.I. Extreme climatic events: Impacts of drought and high temperature on physiological processes in agronomically important plants. Front. Environ. Sci. 2014, 2, 39. [Google Scholar] [CrossRef]
- van Kempen, M.M.; Smolders, A.J.; Bögemann, G.M.; Lamers, L.L.; Visser, E.J.; Roelofs, J.G. Responses of the Azolla filiculoides Stras.–Anabaena azollae Lam. association to elevated sodium chloride concentrations: Amino acids as indicators for salt stress and tipping point. Aquat. Bot. 2013, 106, 20–28. [Google Scholar] [CrossRef]
- Yu, A.; Xie, Y.; Pan, X.; Zhang, H.; Cao, P.; Su, X.; Chang, W.; Li, M. Photosynthetic phosphoribulokinase structures: Enzymatic mechanisms and the redox regulation of the Calvin-Benson-Bassham cycle. Plant Cell 2020, 32, 1556–1573. [Google Scholar] [CrossRef]
- Li, R.; He, Y.; Chen, J.; Zheng, S.; Zhuang, C. Research progress in improving photosynthetic efficiency. Int. J. Mol. Sci. 2023, 24, 9286. [Google Scholar] [CrossRef] [PubMed]
- Rott, M.; Martins, N.F.; Thiele, W.; Lein, W.; Bock, R.; Kramer, D.M.; Schöttler, M.A. ATP synthase repression in tobacco restricts photosynthetic electron transport, CO2 assimilation, and plant growth by overacidification of the thylakoid lumen. Plant Cell 2011, 23, 304–321. [Google Scholar] [CrossRef]
- Franklin, K.A. Light and temperature signal crosstalk in plant development. Curr. Opin. Plant Biol. 2009, 12, 63–68. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity stress affects photosynthesis, malondialdehyde formation, and proline content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.H.; Jaafar, H.Z. Primary, secondary metabolites, H2O2, malondialdehyde and photosynthetic responses of Orthosiphon stimaneus Benth. to different irradiance levels. Molecules 2012, 17, 1159–1176. [Google Scholar] [CrossRef]
- Zhang, J.; Li, H.; Xu, B.; Li, J.; Huang, B. Exogenous melatonin suppresses dark-induced leaf senescence by activating the superoxide dismutase-catalase antioxidant pathway and down-regulating chlorophyll degradation in excised leaves of perennial ryegrass (Lolium perenne L.). Front. Plant Sci. 2016, 7, 1500. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, C.; Nan, S.; Li, Y.; Hu, J.; Zhao, K.; Guo, J.; Wang, S. PagSOD2a improves poplar salt tolerance by elevating superoxide dismutase activity and decreasing malondialdehyde contents. Front. Plant Sci. 2024, 15, 1456249. [Google Scholar] [CrossRef]
- Jie, Z.; Liu, J.; Shu, M.; Ying, Y.; Yang, H. Detection strategies for superoxide anion: A review. Talanta 2022, 236, 122892. [Google Scholar] [CrossRef] [PubMed]
- Kirkegaard, J.A.; Lilley, J.M.; Brill, R.D.; Ware, A.H.; Walela, C.K. The critical period for yield and quality determination in canola (Brassica napus L.). Field Crop. Res. 2018, 222, 180–188. [Google Scholar] [CrossRef]
- Rondanini, D.P.; del Pilar Vilariño, M.; Roberts, M.E.; Polosa, M.A.; Botto, J.F. Physiological responses of spring rapeseed (Brassica napus) to red/far-red ratios and irradiance during pre-and post-flowering stages. Physiol. Plant. 2014, 152, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Labra, M.H.; Struik, P.C.; Evers, J.B.; Calderini, D.F. Plasticity of seed weight compensates reductions in seed number of oilseed rape in response to shading at flowering. Eur. J. Agron. 2017, 84, 113–124. [Google Scholar] [CrossRef]
- Huang, Y.; Schnurbusch, T. The birth and death of floral organs in cereal crops. Annu. Rev. Plant Biol. 2024, 75, 427–458. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Indexes | V | L | V × L |
|---|---|---|---|---|
| 2021–2022 | Chlorophyll a | * | * | ns |
| Chlorophyll b | ns | * | ns | |
| Total chlorophyll | * | * | ns | |
| Sucrose | * | * | ns | |
| Glucose | * | * | ns | |
| Fructose | ns | * | * | |
| 2022–2023 | Chlorophyll a | * | * | ns |
| Chlorophyll b | ns | * | ns | |
| Total chlorophyll | * | * | ns | |
| Sucrose | * | * | * | |
| Glucose | * | * | * | |
| Fructose | ns | * | ns |
| Treatments | Rubisco | FBP | FBA | SS | SPS |
|---|---|---|---|---|---|
| V | * | * | * | * | * |
| L | * | * | * | * | * |
| V × L | * | * | ns | * | * |
| Treatments | ATP | Amino Acid | Proline |
|---|---|---|---|
| V | ns | ns | * |
| L | * | * | ns |
| V × L | * | * | * |
| Treatments | MDA | CAT | SOD |
|---|---|---|---|
| V | ns | * | * |
| L | * | ns | * |
| V × L | * | ns | * |
| Year | Varieties | Treatments | Number of Pods per Plant | Number of Seeds per Pod | 1000 Seed Weight (g) | Yield (kg plot−1) |
|---|---|---|---|---|---|---|
| 2021–2022 | JYJS01 | CK | 277.50 a | 22.50 a | 4.04 c | 6.49 a |
| L | 191.33 d | 18.17 c | 4.44 b | 5.96 bc | ||
| CYZ1 | CK | 237.33 bc | 20.17 b | 4.08 c | 3.70 d | |
| L | 135.17 e | 14.33 e | 4.78 a | 2.88 e | ||
| 2022–2023 | JYJS01 | CK | 260.17 ab | 22.17 a | 3.99 d | 6.25 ab |
| L | 202.83 cd | 16.50 d | 4.39 b | 5.86 c | ||
| CYZ1 | CK | 220.50 cd | 20.33 b | 4.10 c | 3.82 d | |
| L | 128.00 e | 13.50 e | 4.72 a | 2.69 e | ||
| p-value | Y | ns | * | ns | ns | |
| V | * | ns | ns | * | ||
| T | * | * | * | * | ||
| Y × V | ns | ns | ns | ns | ||
| Y × T | ns | * | ns | ns | ||
| V × T | ns | ns | ns | ns | ||
| Y × V × T | ns | ns | ns | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Y.; Javed, H.H.; Liu, L.; Liu, Y.; Yang, X.; Xu, F.; Liu, Y.; Peng, X.; Wu, Y. Impact of Low Light on Photosynthetic Characteristics, Antioxidant Activity, and Yield of Brassica napus L. Agronomy 2025, 15, 214. https://doi.org/10.3390/agronomy15010214
Hu Y, Javed HH, Liu L, Liu Y, Yang X, Xu F, Liu Y, Peng X, Wu Y. Impact of Low Light on Photosynthetic Characteristics, Antioxidant Activity, and Yield of Brassica napus L. Agronomy. 2025; 15(1):214. https://doi.org/10.3390/agronomy15010214
Chicago/Turabian StyleHu, Yue, Hafiz Hassan Javed, Li Liu, Yalong Liu, Xiaohui Yang, Feifei Xu, Ying Liu, Xiao Peng, and Yongcheng Wu. 2025. "Impact of Low Light on Photosynthetic Characteristics, Antioxidant Activity, and Yield of Brassica napus L." Agronomy 15, no. 1: 214. https://doi.org/10.3390/agronomy15010214
APA StyleHu, Y., Javed, H. H., Liu, L., Liu, Y., Yang, X., Xu, F., Liu, Y., Peng, X., & Wu, Y. (2025). Impact of Low Light on Photosynthetic Characteristics, Antioxidant Activity, and Yield of Brassica napus L. Agronomy, 15(1), 214. https://doi.org/10.3390/agronomy15010214