
Zinc Oxide Nanoparticles: An Influential Element in Alleviating Salt Stress in Quinoa (Chenopodium quinoa L. Cv Atlas)

 ,
,  ,
, 
 ,
,  , , , ,
, , , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials, Growth Conditions, and Treatments
2.2. Synthesis of Zn Nanoparticles (ZnO-NPs)
2.3. Morpho-Physiological and Biochemical Properties
2.4. K/Na and Ca/Na Ratios
2.5. Antioxidant Enzyme Activities (CAT, POD, and SOD)
2.6. H2O2, MDA, Sucrose, and Proline
2.7. Statistical Analysis
3. Results
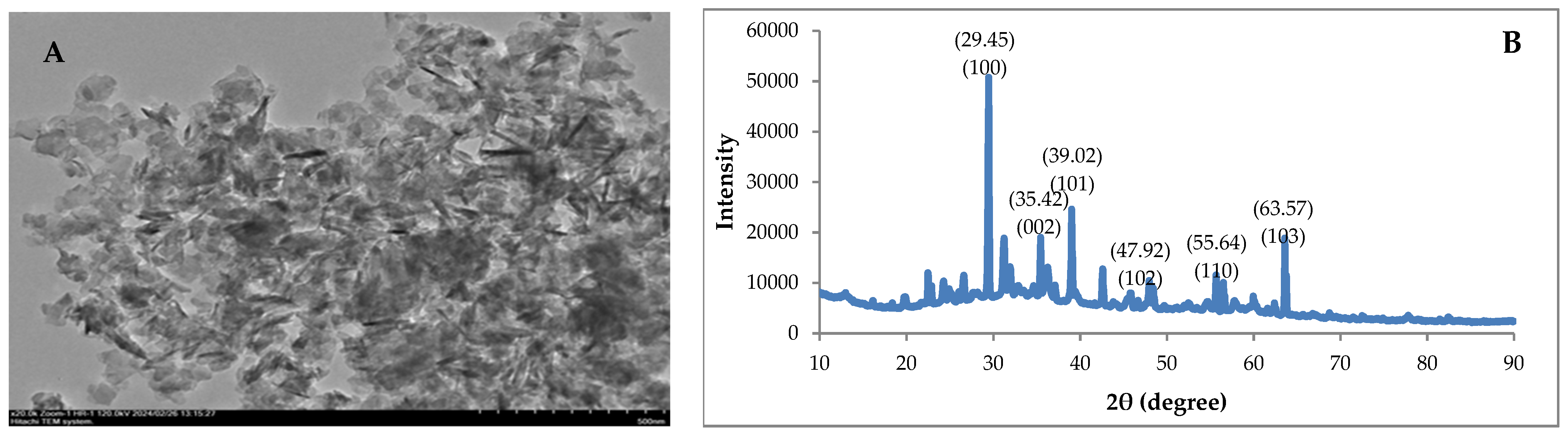
3.1. Characterization of ZnO-NPs
3.2. Plant Growth
3.3. K/Na and Ca/Na Ratios
3.4. Antioxidant Enzyme Activities
3.5. Hydrogen Peroxide (H2O2), Melondialdehyde (MDA), Proline, and Sucrose Contents
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, L.; Chhogyel, N.; Gopalakrishnan, T.; Hasan, M.K.; Jayasinghe, S.L.; Kariyawasam, C.S.; Kogo, B.K.; Ratnayake, S. Climate change and future of agri-food production. In Future Foods; Elsevier: Amsterdam, The Netherlands, 2022; pp. 49–79. [Google Scholar]
- Mishra, A.K.; Das, R.; George Kerry, R.; Biswal, B.; Sinha, T.; Sharma, S.; Arora, P.; Kumar, M. Promising management strategies to improve crop sustainability and to amend soil salinity. Front. Environ. Sci. 2023, 10, 962581. [Google Scholar] [CrossRef]
- Hussain, M.I.; Al-Dakheel, A.J.; Reigosa, M.J. Genotypic differences in agro-physiological, biochemical and isotopic responses to salinity stress in quinoa (Chenopodium quinoa Willd.) plants: Prospects for salinity tolerance and yield stability. Plant Physiol. Biochem. 2018, 129, 411–420. [Google Scholar] [CrossRef]
- Shahid, M.A.; Sarkhosh, A.; Khan, N.; Balal, R.M.; Ali, S.; Rossi, L.; Gómez, C.; Mattson, N.; Nasim, W.; Garcia-Sanchez, F. Insights into the physiological and biochemical impacts of salt stress on plant growth and development. Agronomy 2020, 10, 938. [Google Scholar] [CrossRef]
- Gul, Z.; Tang, Z.-H.; Arif, M.; Ye, Z. An insight into abiotic stress and influx tolerance mechanisms in plants to cope in saline environments. Biology 2022, 11, 597. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef]
- Hao, S.; Wang, Y.; Yan, Y.; Liu, Y.; Wang, J.; Chen, S. A review on plant responses to salt stress and their mechanisms of salt resistance. Horticulturae 2021, 7, 132. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Masud, A.A.C.; Rahman, K.; Nowroz, F.; Rahman, M.; Nahar, K.; Fujita, M. Regulation of reactive oxygen species and antioxidant defense in plants under salinity. Int. J. Mol. Sci. 2021, 22, 9326. [Google Scholar] [CrossRef]
- Sardar, H.; Khalid, Z.; Ahsan, M.; Naz, S.; Nawaz, A.; Ahmad, R.; Razzaq, K.; Wabaidur, S.M.; Jacquard, C.; Širić, I. Enhancement of salinity stress tolerance in lettuce (Lactuca sativa L.) via foliar application of nitric oxide. Plants 2023, 12, 1115. [Google Scholar] [CrossRef]
- Turan, M.; Ekinci, M.; Kul, R.; Boynueyri, F.G.; Yildirim, E. Mitigation of salinity stress in cucumber seedlings by exogenous hydrogen sulfide. J. Plant Res. 2022, 135, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77. [Google Scholar] [CrossRef]
- Shahzad, S.; Ali, S.; Ahmad, R.; Ercisli, S.; Anjum, M.A. Foliar application of silicon enhances growth, flower yield, quality and postharvest life of tuberose (Polianthes tuberosa L.) under saline conditions by improving antioxidant defense mechanism. Silicon 2022, 14, 1511–1518. [Google Scholar] [CrossRef]
- Alkharabsheh, H.M.; Seleiman, M.F.; Hewedy, O.A.; Battaglia, M.L.; Jalal, R.S.; Alhammad, B.A.; Schillaci, C.; Ali, N.; Al-Doss, A. Field crop responses and management strategies to mitigate soil salinity in modern agriculture: A review. Agronomy 2021, 11, 2299. [Google Scholar] [CrossRef]
- Ruiz, K.; Biondi, S.; Martínez, E.A.; Orsini, F.; Antognoni, F.; Jacobsen, S.-E. Quinoa–a model crop for understanding salt-tolerance mechanisms in halophytes. Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2016, 150, 357–371. [Google Scholar] [CrossRef]
- Dakhili, S.; Abdolalizadeh, L.; Hosseini, S.M.; Shojaee-Aliabadi, S.; Mirmoghtadaie, L. Quinoa protein: Composition, structure and functional properties. Food Chem. 2019, 299, 125161. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco-Valencia, R.A.-M.; Serna, L.A. Quinoa (Chenopodium quinoa Willd.) as a source of dietary fiber and other functional components. Food Sci. Technol. 2011, 31, 225–230. [Google Scholar] [CrossRef]
- Fischer, S.; Wilckens, R.; Jara, J.; Aranda, M.; Valdivia, W.; Bustamante, L.; Graf, F.; Obal, I. Protein and antioxidant composition of quinoa (Chenopodium quinoa Willd.) sprout from seeds submitted to water stress, salinity and light conditions. Ind. Crop. Prod. 2017, 107, 558–564. [Google Scholar] [CrossRef]
- Pathan, S.; Siddiqui, R.A. Nutritional composition and bioactive components in quinoa (Chenopodium quinoa Willd.) greens: A review. Nutrients 2022, 14, 558. [Google Scholar] [CrossRef] [PubMed]
- Vilcacundo, R.; Hernández-Ledesma, B. Nutritional and biological value of quinoa (Chenopodium quinoa Willd.). Curr. Opin. Food Sci. 2017, 14, 1–6. [Google Scholar] [CrossRef]
- Gómez-Pando, L.R.; Álvarez-Castro, R.; Eguiluz-De La Barra, A. Effect of salt stress on Peruvian germplasm of Chenopodium quinoa Willd.: A promising crop. J. Agron. Crop Sci. 2010, 196, 391–396. [Google Scholar] [CrossRef]
- Causin, H.F.; Bordón, D.A.; Burrieza, H. Salinity tolerance mechanisms during germination and early seedling growth in Chenopodium quinoa Wild. genotypes with different sensitivity to saline stress. Environ. Exp. Bot. 2020, 172, 103995. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, K.; Abd-Elsalam, K.A. Exploring the potential of nanofertilizers for a sustainable agriculture. Plant Nano Biol. 2023, 5, 100044. [Google Scholar] [CrossRef]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Badakhshan, H. Effects of zinc application on growth, absorption and distribution of mineral nutrients under salinity stress in soybean (Glycine max L.). J. Plant Nutr. 2014, 37, 2255–2269. [Google Scholar] [CrossRef]
- Tavallali, V.; Rahemi, M.; Maftoun, M.; Panahi, B.; Karimi, S.; Ramezanian, A.; Vaezpour, M. Zinc influence and salt stress on photosynthesis, water relations, and carbonic anhydrase activity in pistachio. Sci. Hortic. 2009, 123, 272–279. [Google Scholar] [CrossRef]
- Tolay, I. The impact of different Zinc (Zn) levels on growth and nutrient uptake of Basil (Ocimum basilicum L.) grown under salinity stress. PLoS ONE 2021, 16, e0246493. [Google Scholar] [CrossRef] [PubMed]
- Amooaghaie, R.; Norouzi, M.; Saeri, M. Impact of zinc and zinc oxide nanoparticles on the physiological and biochemical processes in tomato and wheat. Botany 2017, 95, 441–455. [Google Scholar] [CrossRef]
- Zafar, S.; Hasnain, Z.; Aslam, N.; Mumtaz, S.; Jaafar, H.Z.; Wahab, P.E.M.; Qayum, M.; Ormenisan, A.N. Impact of Zn nanoparticles synthesized via green and chemical approach on okra (Abelmoschus esculentus L.) growth under salt stress. Sustainability 2021, 13, 3694. [Google Scholar] [CrossRef]
- Elshoky, H.A.; Yotsova, E.; Farghali, M.A.; Farroh, K.Y.; El-Sayed, K.; Elzorkany, H.E.; Rashkov, G.; Dobrikova, A.; Borisova, P.; Stefanov, M. Impact of foliar spray of zinc oxide nanoparticles on the photosynthesis of Pisum sativum L. under salt stress. Plant Physiol. Biochem. 2021, 167, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Hosseinpour, A.; Haliloglu, K.; Tolga Cinisli, K.; Ozkan, G.; Ozturk, H.I.; Pour-Aboughadareh, A.; Poczai, P. Application of zinc oxide nanoparticles and plant growth promoting bacteria reduces genetic impairment under salt stress in tomato (Solanum lycopersicum L.‘Linda’). Agriculture 2020, 10, 521. [Google Scholar] [CrossRef]
- Hosseinpour, A.; Ilhan, E.; Özkan, G.; Öztürk, H.İ.; Haliloglu, K.; Cinisli, K.T. Plant growth-promoting bacteria (PGPBs) and copper (II) oxide (CuO) nanoparticle ameliorates DNA damage and DNA Methylation in wheat (Triticum aestivum L.) exposed to NaCl stress. J. Plant Biochem. Biotechnol. 2022, 31, 751–764. [Google Scholar] [CrossRef]
- Manaa, A.; Goussi, R.; Derbali, W.; Cantamessa, S.; Abdelly, C.; Barbato, R. Salinity tolerance of quinoa (Chenopodium quinoa Willd) as assessed by chloroplast ultrastructure and photosynthetic performance. Environ. Exp. Bot. 2019, 162, 103–114. [Google Scholar] [CrossRef]
- Du, W.; Yang, J.; Peng, Q.; Liang, X.; Mao, H. Comparison study of zinc nanoparticles and zinc sulphate on wheat growth: From toxicity and zinc biofortification. Chemosphere 2019, 227, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Haliloğlu, K.; Türkoğlu, A.; Balpınar, Ö.; Nadaroğlu, H.; Alaylı, A.; Poczai, P. Effects of zinc, copper and iron oxide nanoparticles on induced DNA methylation, genomic instability and LTR retrotransposon polymorphism in wheat (Triticum aestivum L.). Plants 2022, 11, 2193. [Google Scholar] [CrossRef]
- Nalci, O.B.; Nadaroglu, H.; Pour, A.H.; Gungor, A.A.; Haliloglu, K. Effects of ZnO, CuO and γ-Fe3O4 nanoparticles on mature embryo culture of wheat (Triticum aestivum L.). Plant Cell Tissue Organ Cult. (PCTOC) 2019, 136, 269–277. [Google Scholar] [CrossRef]
- Palangi, V.; Macit, M.; Nadaroglu, H.; Taghizadeh, A. Effects of green-synthesized CuO and ZnO nanoparticles on ruminal mitigation of methane emission to the enhancement of the cleaner environment. Biomass Convers. Biorefinery 2024, 14, 5447–5455. [Google Scholar] [CrossRef]
- Rajendran, S.P.; Sengodan, K. Synthesis and characterization of zinc oxide and iron oxide nanoparticles using Sesbania grandiflora leaf extract as reducing agent. J. Nanosci. 2017, 2017, 8348507. [Google Scholar] [CrossRef]
- Mertens, D. Plants preparation of laboratory sample. In Official Methods of Analysis, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; AOAC: Gaithersburg, MD, USA, 2005; pp. 1–2. [Google Scholar]
- Mertens, D. AOAC Official Method 975.03. Metals in Plants and Pets Food Atomic Absorption Spectrophotometric Method, 18th ed.; Horwitz, W., Latimer, G.W., Eds.; Scientific Research Publishing: Wuhan, China, 2005; pp. 3–4. [Google Scholar]
- Liu, K.; Yuan, C.; Chen, Y.; Li, H.; Liu, J. Combined effects of ascorbic acid and chitosan on the quality maintenance and shelf life of plums. Sci. Hortic. 2014, 176, 45–53. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Khan, I.; Raza, M.A.; Awan, S.A.; Shah, G.A.; Rizwan, M.; Ali, B.; Tariq, R.; Hassan, M.J.; Alyemeni, M.N.; Brestic, M. Amelioration of salt induced toxicity in pearl millet by seed priming with silver nanoparticles (AgNPs): The oxidative damage, antioxidant enzymes and ions uptake are major determinants of salt tolerant capacity. Plant Physiol. Biochem. 2020, 156, 221–232. [Google Scholar] [CrossRef]
- Sharmin, S.; Lipka, U.; Polle, A.; Eckert, C. The influence of transpiration on foliar accumulation of salt and nutrients under salinity in poplar (Populus × canescens). PLoS ONE 2021, 16, e0253228. [Google Scholar] [CrossRef]
- Siddiqui, H.; Yusuf, M.; Faraz, A.; Faizan, M.; Sami, F.; Hayat, S. 24-Epibrassinolide supplemented with silicon enhances the photosynthetic efficiency of Brassica juncea under salt stress. S. Afr. J. Bot. 2018, 118, 120–128. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Tanveer, K.; Gilani, S.; Hussain, Z.; Ishaq, R.; Adeel, M.; Ilyas, N. Effect of salt stress on tomato plant and the role of calcium. J. Plant Nutr. 2020, 43, 28–35. [Google Scholar] [CrossRef]
- Hannachi, S.; Van Labeke, M.-C. Salt stress affects germination, seedling growth and physiological responses differentially in eggplant cultivars (Solanum melongena L.). Sci. Hortic. 2018, 228, 56–65. [Google Scholar] [CrossRef]
- Abbas, W.; Ashraf, M.; Akram, N.A. Alleviation of salt-induced adverse effects in eggplant (Solanum melongena L.) by glycinebetaine and sugarbeet extracts. Sci. Hortic. 2010, 125, 188–195. [Google Scholar] [CrossRef]
- Rakgotho, T.; Ndou, N.; Mulaudzi, T.; Iwuoha, E.; Mayedwa, N.; Ajayi, R.F. Green-synthesized zinc oxide nanoparticles mitigate salt stress in Sorghum bicolor. Agriculture 2022, 12, 597. [Google Scholar] [CrossRef]
- Solimaninya, Z.; Mohtadi, A.; Movahhedi Dehnavi, M. Response of some physiological and morphological properties of quinoa (Chenopodium quinoa Willd.) by zinc application under drought stress. J. Plant Process Funct. 2021, 10, 171–186. [Google Scholar]
- Jorfi, A.; Alavifazel, M.; Gilani, A.; Ardakani, M.R.; Lak, S. Quinoa (Chenopodium quinoa) root system development as affected by phosphorus and zinc sulfate application in an alkaline soil. Gesunde Pflanz. 2023, 75, 885–897. [Google Scholar] [CrossRef]
- Bostani, A.; Badi, H.N.; Pakbaz, N.; Omidi, H. Foliar application of iron and zinc on quinoa under drought stress affects its seeds germination and biochemical properties. Iran. J. Plant Physiol. 2022, 12, 4153–4167. [Google Scholar]
- Lang, C.; Mission, E.G.; Fuaad, A.A.-H.A.; Shaalan, M. Nanoparticle tools to improve and advance precision practices in the Agrifoods Sector towards sustainability-A review. J. Clean. Prod. 2021, 293, 126063. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef]
- Sturikova, H.; Krystofova, O.; Huska, D.; Adam, V. Zinc, zinc nanoparticles and plants. J. Hazard. Mater. 2018, 349, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Vafa, Z.N.; Sirousmehr, A.R.; Ghanbari, A.; Khammari, I.; Falahi, N. Effects of nano zinc and humic acid on quantitative and qualitative characteristics of savory (Satureja hortensis L.). Int. J. Biosci. 2015, 6, 124–136. [Google Scholar] [CrossRef]
- Faizan, M.; Bhat, J.A.; Chen, C.; Alyemeni, M.N.; Wijaya, L.; Ahmad, P.; Yu, F. Zinc oxide nanoparticles (ZnO-NPs) induce salt tolerance by improving the antioxidant system and photosynthetic machinery in tomato. Plant Physiol. Biochem. 2021, 161, 122–130. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Sengar, R.S.; Shahi, U.P.; Rajput, V.D.; Minkina, T.; Ghazaryan, K.A. Prominent effects of zinc oxide nanoparticles on roots of rice (Oryza sativa L.) grown under salinity stress. Stresses 2022, 3, 33–46. [Google Scholar] [CrossRef]
- Nadeem, F.; Azhar, M.; Anwar-ul-Haq, M.; Sabir, M.; Samreen, T.; Tufail, A.; Awan, H.U.M.; Juan, W. Comparative response of two rice (Oryza sativa L.) cultivars to applied zinc and manganese for mitigation of salt stress. J. Soil Sci. Plant Nutr. 2020, 20, 2059–2072. [Google Scholar] [CrossRef]
- Rasouli, F.; Asadi, M.; Hassanpouraghdam, M.B.; Aazami, M.A.; Ebrahimzadeh, A.; Kakaei, K.; Dokoupil, L.; Mlcek, J. Foliar application of ZnO-NPs influences chlorophyll fluorescence and antioxidants pool in Capsicum annum L. under salinity. Horticulturae 2022, 8, 908. [Google Scholar] [CrossRef]
- Mahawar, L.; Živčák, M.; Barboricova, M.; Kovár, M.; Filaček, A.; Ferencova, J.; Vysoká, D.M.; Brestič, M. Effect of copper oxide and zinc oxide nanoparticles on photosynthesis and physiology of Raphanus sativus L. under salinity stress. Plant Physiol. Biochem. 2024, 206, 108281. [Google Scholar] [CrossRef] [PubMed]
- Seyed Sharifi, R.; Khalilzadeh, R.; Soltanmoradi, S. Effects of Mycorrhizal Fungi and Nano Zinc Oxide on Seed Yield, Na+ and K+ Content of Wheat (Triticum aestivum L.) under Salinity Stress. J. Crop Nutr. Sci. 2017, 3, 40–53. [Google Scholar]
- Manaa, A.; Ben Ahmed, H.; Valot, B.; Bouchet, J.-P.; Aschi-Smiti, S.; Causse, M.; Faurobert, M. Salt and genotype impact on plant physiology and root proteome variations in tomato. J. Exp. Bot. 2011, 62, 2797–2813. [Google Scholar] [CrossRef]
- Ekinci, M.; Yildirim, E.; Turan, M. Ameliorating effects of hydrogen sulfide on growth, physiological and biochemical characteristics of eggplant seedlings under salt stress. S. Afr. J. Bot. 2021, 143, 79–89. [Google Scholar] [CrossRef]
- Mahmud, S.; Sharmin, S.; Chowdhury, B.; Hossain, M.A. Research article effect of salinity and alleviating role of methyl jasmonate in some rice varieties. Asian J. Plant Sci 2017, 16, 87–93. [Google Scholar] [CrossRef]
- Orlovsky, N.; Japakova, U.; Zhang, H.; Volis, S. Effect of salinity on seed germination, growth and ion content in dimorphic seeds of Salicornia europaea L. (Chenopodiaceae). Plant Divers. 2016, 38, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Lu, Y.; Wu, M.; Liang, E.; Li, Y.; Zhang, D.; Yin, Z.; Ren, X.; Dai, Y.; Deng, D. Ability to remove Na+ and retain K+ correlates with salt tolerance in two maize inbred lines seedlings. Front. Plant Sci. 2016, 7, 1716. [Google Scholar] [CrossRef]
- AbdElgawad, H.; Zinta, G.; Hegab, M.M.; Pandey, R.; Asard, H.; Abuelsoud, W. High salinity induces different oxidative stress and antioxidant responses in maize seedlings organs. Front. Plant Sci. 2016, 7, 179763. [Google Scholar] [CrossRef] [PubMed]
- Hussein, M.; Abou-Baker, N. The contribution of nano-zinc to alleviate salinity stress on cotton plants. R. Soc. Open Sci. 2018, 5, 171809. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.I.; Niño-Medina, G.; Olivares-Sáenz, E.; Lira-Saldivar, R.H.; Barriga-Castro, E.D.; Vázquez-Alvarado, R.; Rodríguez-Salinas, P.A.; Zavala-García, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compounds in habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A. Effects of zinc-oxide nanoparticles on soil, plants, animals and soil organisms: A review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84. [Google Scholar] [CrossRef]
- Yasmin, H.; Mazher, J.; Azmat, A.; Nosheen, A.; Naz, R.; Hassan, M.N.; Noureldeen, A.; Ahmad, P. Combined application of zinc oxide nanoparticles and biofertilizer to induce salt resistance in safflower by regulating ion homeostasis and antioxidant defence responses. Ecotoxicol. Environ. Saf. 2021, 218, 112262. [Google Scholar] [CrossRef]
- Alabdallah, N.M.; Alzahrani, H.S. The potential mitigation effect of ZnO nanoparticles on [Abelmoschus esculentus L. Moench] metabolism under salt stress conditions. Saudi J. Biol. Sci. 2020, 27, 3132–3137. [Google Scholar] [CrossRef]
- Ahmad, P.; Ahanger, M.A.; Alyemeni, M.N.; Wijaya, L.; Egamberdieva, D.; Bhardwaj, R.; Ashraf, M. Zinc application mitigates the adverse effects of NaCl stress on mustard [Brassica juncea (L.) Czern & Coss] through modulating compatible organic solutes, antioxidant enzymes, and flavonoid content. J. Plant Interact. 2017, 12, 429–437. [Google Scholar]

{kind=link}
| Sources of Variation | Plant Height | Plant Weight | Plant Diameter | Chlorophyll Content (SPAD) |
|---|---|---|---|---|
| F value (salinity) | 24.00 *** | 190.31 *** | 3.96 * | 163.27 *** |
| F value (Zn) | 9.81 ** | 66.94 *** | 51.32 *** | 52.82 *** |
| F value (concentration) | 2.15 ns | 3.40 * | 2.91 * | 2.72 ns |
| F value (S × Z) | 4.16 * | 0.55 ns | 0.39 ns | 0.01 ns |
| F value (S × C) | 3.22 ** | 4.14 ** | 0.34 ns | 10.09 *** |
| F value (Z × C) | 0.92 ns | 2.71 * | 0.60 ns | 0.23 ns |
| F value (S × Z × C) | 2.20 * | 0.57 ns | 0.32 ns | 0.25 ns |
| CV % | 12.20 | 14.08 | 13.41 | 4.18 |
| Salinity | Zn Treatments | Zinc Concentration (ppm) | Plant Height (cm) | Plant Weight (g) | Plant Diameter (mm) | Chlorophyll Content (SPAD) |
|---|---|---|---|---|---|---|
| 0 mM | Zn | 0.0 | 11.17 | 1.02 | 1.34 | 34.07 |
| 50.0 | 8.33 | 1.11 | 1.53 | 36.07 | ||
| 100.0 | 8.50 | 1.29 | 1.60 | 37.63 | ||
| 200.0 | 11.17 | 1.37 | 1.81 | 39.97 | ||
| Mean | 9.79 a | 1.20 b | 1.57 | 36.93 b | ||
| Zn-NPs | 0.0 | 9.83 | 1.53 | 1.85 | 36.70 | |
| 50.0 | 10.00 | 1.30 | 1.87 | 38.47 | ||
| 100.0 | 10.67 | 1.51 | 1.91 | 39.97 | ||
| 200.0 | 11.17 | 1.60 | 1.95 | 42.20 | ||
| Mean | 10.42 a | 1.48 a | 1.90 | 39.33 a | ||
| Mean | 0.0 | 10.50 | 1.27 | 1.60 | 35.38 | |
| 50.0 | 9.17 | 1.20 | 1.70 | 37.27 | ||
| 100.0 | 9.58 | 1.40 | 1.76 | 38.80 | ||
| 200.0 | 11.17 | 1.49 | 1.88 | 41.08 | ||
| Mean | 10.10 a | 1.34 a | 1.73 a | 38.13 a | ||
| 100 mM | Zn | 0.0 | 9.83 | 0.66 | 1.23 | 31.83 |
| 50.0 | 8.00 | 0.50 | 1.37 | 34.47 | ||
| 100.0 | 9.83 | 0.56 | 1.40 | 32.83 | ||
| 200.0 | 7.50 | 0.61 | 1.47 | 32.10 | ||
| Mean | 8.79 b | 0.58 d | 1.37 | 32.81 d | ||
| Zn-NPs | 0.0 | 10.00 | 1.00 | 1.68 | 35.10 | |
| 50.0 | 8.50 | 0.78 | 1.78 | 36.40 | ||
| 100.0 | 8.00 | 0.77 | 1.85 | 35.40 | ||
| 200.0 | 8.67 | 0.81 | 1.88 | 34.27 | ||
| Mean | 8.79 b | 0.84 c | 1.80 | 35.29 c | ||
| Mean | 0.0 | 9.92 | 0.83 | 1.45 | 33.47 | |
| 50.0 | 8.25 | 0.64 | 1.58 | 35.43 | ||
| 100.0 | 8.92 | 0.67 | 1.63 | 34.12 | ||
| 200.0 | 8.08 | 0.71 | 1.68 | 33.18 | ||
| Mean | 8.79 b | 0.71 b | 1.58 b | 34.05 b | ||
| 200 mM | Zn | 0.0 | 6.50 | 0.41 | 1.31 | 30.90 |
| 50.0 | 7.00 | 0.58 | 1.38 | 28.30 | ||
| 100.0 | 7.17 | 0.67 | 1.44 | 28.57 | ||
| 200.0 | 7.50 | 0.74 | 1.47 | 29.93 | ||
| Mean | 7.04 c | 0.60 d | 1.40 | 29.43 e | ||
| Zn-NPs | 0.0 | 8.17 | 0.71 | 1.82 | 33.23 | |
| 50.0 | 8.83 | 0.89 | 1.59 | 31.83 | ||
| 100.0 | 9.00 | 0.83 | 1.73 | 31.03 | ||
| 200.0 | 9.33 | 0.80 | 1.86 | 31.53 | ||
| Mean | 8.83 b | 0.81 c | 1.75 | 31.91 d | ||
| Mean | 0.0 | 7.33 | 0.56 | 1.57 | 32.07 | |
| 50.0 | 7.92 | 0.74 | 1.48 | 30.07 | ||
| 100.0 | 8.08 | 0.75 | 1.59 | 29.80 | ||
| 200.0 | 8.42 | 0.77 | 1.67 | 30.73 | ||
| Mean | 7.94 c | 0.71 b | 1.58 b | 30.67 c | ||
| Means of Zn treatments | Zn | 0.0 | 9.17 ab | 0.69 | 1.29 | 32.27 |
| 50.0 | 7.78 c | 0.73 | 1.43 | 32.94 | ||
| 100.0 | 8.50 bc | 0.84 | 1.48 | 33.01 | ||
| 200.0 | 8.72 abc | 0.91 | 1.58 | 34.00 | ||
| Mean | 8.54 b | 0.79 b | 1.45 b | 33.06 b | ||
| Zn-NPs | 0.0 | 9.33 ab | 1.08 | 1.78 | 35.01 | |
| 50.0 | 9.11 ab | 0.99 | 1.75 | 35.57 | ||
| 100.0 | 9.22 ab | 1.04 | 1.83 | 35.47 | ||
| 200.0 | 9.72 a | 1.07 | 1.90 | 36.00 | ||
| Mean | 9.35 a | 1.04 a | 1.82 a | 35.51 a | ||
| Means of Zn concentration (ppm) | 0.0 | 9.25 | 0.89 b | 1.54 b | 33.64 b | |
| 50.0 | 8.44 | 0.86 b | 1.59 ab | 34.26 ab | ||
| 100.0 | 8.86 | 0.94 ab | 1.66 ab | 34.24 ab | ||
| 200.0 | 9.22 | 0.99 a | 1.74 a | 35.00 a | ||
| Sources of Variation | K/Na | Ca/Na |
|---|---|---|
| F value (salinity) | 547.46 *** | 1548.31 ** |
| F value (Zn) | 12.26 ** | 179.32 ** |
| F value (concentration) | 9.61 *** | 19.67 ** |
| F value (S × Z) | 129.49 *** | 798.88 ** |
| F value (S × C) | 2.26 ns | 10.57 ** |
| F value (Z × C) | 0.67 ns | 2.49 ns |
| F value (S × Z × C) | 4.26 ** | 14.95 ** |
| CV % | 7.21 | 5.13 |
| Salinity | Zn Treatments | Zinc Concentration (ppm) | K/Na Value | Ca/Na Value |
|---|---|---|---|---|
| 0 mM | Zn | 0.0 | 46.98 cd | 56.15 de |
| 50.0 | 50.23 a–d | 56.54 de | ||
| 100.0 | 46.04 d | 51.98 e | ||
| 200.0 | 50.93 abc | 55.58 de | ||
| Mean | 48.55 b | 55.06 d | ||
| Zn-NPs | 0.0 | 48.64 bcd | 98.67 b | |
| 50.0 | 54.96 a | 105.33 a | ||
| 100.0 | 52.22 ab | 104.29 a | ||
| 200.0 | 53.56 a | 107.66 a | ||
| Mean | 52.34 a | 103.99 a | ||
| Mean | 0.0 | 47.81 | 77.41 b | |
| 50.0 | 52.59 | 80.94 ab | ||
| 100.0 | 49.13 | 78.14 ab | ||
| 200.0 | 52.25 | 81.62 a | ||
| Mean | 50.45 a | 79.53 a | ||
| 100 mM | Zn | 0.0 | 22.28 jk | 55.02 de |
| 50.0 | 21.18 jkl | 54.96 de | ||
| 100.0 | 29.08 i | 77.97 c | ||
| 200.0 | 29.89 hi | 76.56 c | ||
| Mean | 25.60 e | 66.13 b | ||
| Zn-NPs | 0.0 | 36.47 fg | 53.02 e | |
| 50.0 | 39.80 ef | 59.50 d | ||
| 100.0 | 38.59 efg | 59.19 d | ||
| 200.0 | 41.41 e | 58.83 d | ||
| Mean | 39.07 c | 57.63 c | ||
| Mean | 0.0 | 29.37 | 54.02 d | |
| 50.0 | 30.49 | 57.23 d | ||
| 100.0 | 33.84 | 68.58 c | ||
| 200.0 | 35.65 | 67.70 c | ||
| Mean | 32.34 b | 61.88 b | ||
| 200 mM | Zn | 0.0 | 30.89 hi | 39.84 f |
| 50.0 | 34.62 gh | 40.33 f | ||
| 100.0 | 30.32 hi | 36.79 f | ||
| 200.0 | 31.20 hi | 36.73 f | ||
| Mean | 31.76 d | 38.42 e | ||
| Zn-NPs | 0.0 | 17.51 l | 21.12 i | |
| 50.0 | 18.58 kl | 24.45 hi | ||
| 100.0 | 24.39 j | 30.62 g | ||
| 200.0 | 23.52 j | 28.36 gh | ||
| Mean | 21.00 f | 26.14 f | ||
| Mean | 0.0 | 24.20 | 30.48 e | |
| 50.0 | 26.60 | 32.39 e | ||
| 100.0 | 27.35 | 33.71 e | ||
| 200.0 | 27.36 | 32.54 e | ||
| Mean | 26.38 c | 32.28 c | ||
| Means of Zn treatments | Zn | 0.0 | 33.38 | 50.34 |
| 50.0 | 35.34 | 50.61 | ||
| 100.0 | 35.15 | 55.58 | ||
| 200.0 | 37.34 | 56.29 | ||
| Mean | 35.30 b | 53.20 b | ||
| Zn-NPs | 0.0 | 34.20 | 57.60 | |
| 50.0 | 37.78 | 63.09 | ||
| 100.0 | 38.40 | 64.70 | ||
| 200.0 | 39.49 | 64.95 | ||
| Mean | 37.47 a | 62.59 a | ||
| Means of Zn concentration (ppm) | 0.0 | 33.79 c | 53.97 c | |
| 50.0 | 36.56 b | 56.85 b | ||
| 100.0 | 36.77 ab | 60.14 a | ||
| 200.0 | 38.42 a | 60.62 a | ||
| Sources of Variation | SOD | CAT | POD |
|---|---|---|---|
| F value (salinity) | 9730.50 *** | 200.56 *** | 319.76 *** |
| F value (Zn) | 853.05 *** | 39.66 *** | 59.33 *** |
| F value (concentration) | 402.66 *** | 5.22 ** | 11.26 *** |
| F value (S × Z) | 714.33 *** | 25.64 *** | 269.76 *** |
| F value (S × C) | 145.52 *** | 5.16 *** | 9.87 *** |
| F value (Z × C) | 130.28 *** | 4.13 * | 6.36 ** |
| F value (S × Z × C) | 195.06 *** | 10.75 *** | 10.25 *** |
| CV % | 2.38 | 7.38 | 6.80 |
| Salinity | Zn Treatments | Zinc Concentration (ppm) | SOD (EU g−1 Leaf) | CAT (EU g−1 Leaf) | POD (EU g−1 Leaf) |
|---|---|---|---|---|---|
| 0 mM | Zn | 0.0 | 182.45 hi | 4621.41 i | 128.98 fg |
| 50.0 | 191.29 gh | 4909.01 ghi | 124.62 fg | ||
| 100.0 | 182.91 hi | 4682.49 hi | 110.17 g | ||
| 200.0 | 200.54 g | 4955.91 ghi | 148.96 f | ||
| Mean | 189.30 e | 4792.21 e | 128.18 f | ||
| Zn-NPs | 0.0 | 327.24 d | 5905.06 cde | 244.50 cde | |
| 50.0 | 189.96 gh | 5032.16 f–i | 129.71 fg | ||
| 100.0 | 203.34 g | 4461.25 i | 134.65 fg | ||
| 200.0 | 195.71 gh | 5296.80 e- i | 142.32 f | ||
| Mean | 229.06 d | 5173.82 d | 162.79 d | ||
| Mean | 0.0 | 254.84 d | 5263.24 def | 186.74 d | |
| 50.0 | 190.62 f | 4970.58 fg | 127.17 f | ||
| 100.0 | 193.13 f | 4571.87 g | 122.41 f | ||
| 200.0 | 198.12 f | 5126.36 ef | 145.64 e | ||
| Mean | 209.18 c | 4983.01 c | 145.49 c | ||
| 100 mM | Zn | 0.0 | 301.09 ef | 6573.88 c | 276.80 ab |
| 50.0 | 295.15 f | 5712.01 d–g | 283.29 a | ||
| 100.0 | 292.48 f | 5816.25 c–f | 256.72 bc d | ||
| 200.0 | 195.75 gh | 6387.83 cd | 267.08 ab c | ||
| Mean | 271.12 c | 6122.49 c | 270.97 a | ||
| Zn-NPs | 0.0 | 175.89 i | 4867.29 hi | 139.39 f | |
| 50.0 | 197.69 g | 4816.63 hi | 147.22 f | ||
| 100.0 | 202.05 g | 4725.70 hi | 138.48 f | ||
| 200.0 | 189.79 gh | 4820.66 hi | 144.49 f | ||
| Mean | 191.35 e | 4807.57 e | 142.39 e | ||
| Mean | 0.0 | 238.49 e | 5720.59 d | 208.09 bc | |
| 50.0 | 246.42 de | 5264.32 def | 215.25 b | ||
| 100.0 | 247.26 de | 5270.98 def | 197.60 cd | ||
| 200.0 | 192.77 f | 5604.24 de | 205.79 bc | ||
| Mean | 231.24 b | 5465.03 b | 206.68 b | ||
| 200 mM | Zn | 0.0 | 556.58 a | 8557.88 a | 232.90 de |
| 50.0 | 524.01 b | 7724.36 b | 237.23 de | ||
| 100.0 | 527.41 b | 7840.75 ab | 236.21 de | ||
| 200.0 | 499.78 c | 7430.72 b | 224.12 e | ||
| Mean | 526.94 a | 7888.43 a | 232.62 c | ||
| Zn-NPs | 0.0 | 526.13 b | 5922.03 cde | 252.52 bc d | |
| 50.0 | 510.34 c | 5488.63 e–h | 263.94 ab c | ||
| 100.0 | 310.24 e | 8078.36 ab | 243.68 cd e | ||
| 200.0 | 322.62 d | 7988.93 ab | 252.37 bc d | ||
| Mean | 417.33 b | 6869.49 b | 253.13 b | ||
| Mean | 0.0 | 541.35 a | 7239.95 b | 242.71 a | |
| 50.0 | 517.17 b | 6606.50 c | 250.59 a | ||
| 100.0 | 418.83 c | 7959.55 a | 239.95 a | ||
| 200.0 | 411.20 c | 7709.82 ab | 238.24 a | ||
| Mean | 472.14 a | 7378.96 a | 242.87 a | ||
| Mean of Zn treatments | Zn | 0.0 | 346.71 a | 6258.15 a | 212.89 a |
| 50.0 | 336.82 bc | 6584.39 bc | 215.05 a | ||
| 100.0 | 334.27 c | 6113.17 bc | 201.03 a | ||
| 200.0 | 298.69 d | 6115.12 ab | 213.39 a | ||
| Mean | 329.12 a | 6267.71 a | 210.59 a | ||
| Zn-NPs | 0.0 | 343.09 a | 6035.46 d | 212.14 a | |
| 50.0 | 299.33 d | 5564.79 e | 180.29 b | ||
| 100.0 | 238.54 e | 5755.10 cd | 172.27 b | ||
| 200.0 | 236.04 e | 5112.47 bc | 179.73 b | ||
| Mean | 279.25 b | 5616.96 b | 186.11 b | ||
| Means of Zn concentration (ppm) | 0.0 | 344.90 a | 6146.81 a | 212.51 a | |
| 50.0 | 318.07 b | 6074.59 b | 197.67 b | ||
| 100.0 | 286.41 c | 5934.13 a | 186.65 b | ||
| 200.0 | 267.37 d | 5613.80 a | 196.56 c | ||
| Sources of Variation | H2O2 | MDA | Proline | Sucrose |
|---|---|---|---|---|
| F value (Salinity) | 415.53 *** | 164.03 *** | 255.97 *** | 37.15 *** |
| F value (Zn) | 30.02 *** | 265.35 *** | 53.38 *** | 26.09 *** |
| F value (concentration) | 24.55 *** | 39.91 *** | 14.95 *** | 27.12 *** |
| F value (S × Z) | 82.48 *** | 41.08 *** | 68.11 *** | 36.69 *** |
| F value (S × C) | 8.45 *** | 11.01 *** | 9.77 *** | 9.85 *** |
| F value (Z × C) | 33.04 *** | 11.35 *** | 30.91 *** | 66.54 *** |
| F value (S × Z × C) | 17.92 *** | 26.41 *** | 30.19 *** | 17.70 *** |
| CV % | 5.77 | 4.04 | 7.29 | 6.33 |
| Salinity | Zn Treatments | Zinc Concentration (ppm) | H2O2 (mmol kg−1) | MDA (nmol g−1 DW) | Proline (µg g−1 FW) | Sucrose (g L−1) |
|---|---|---|---|---|---|---|
| 0 mM | Zn | 0.0 | 10.01 e–h | 5.27 ef | 151.66 ijk | 33.76 hij |
| 50.0 | 8.53 i | 5.75 e | 134.18 k | 32.64 ijk | ||
| 100.0 | 9.24 hi | 5.17 ef | 144.04 jk | 45.58 ab | ||
| 200.0 | 10.09 efg | 5.27 ef | 196.67 fg | 35.14 f–i | ||
| Mean | 9.21 de | 5.37 e | 156.64 d | 36.78 ab | ||
| Zn-NPs | 0.0 | 9.34 gh | 5.45 e | 206.49 f | 27.24 lm | |
| 50.0 | 10.39 ef | 6.81 d | 171.24 g–i | 27.00 lm | ||
| 100.0 | 9.70 fgh | 7.34 d | 168.38 hij | 29.86 ijk | ||
| 200.0 | 7.41 j | 8.19 c | 142.98 jk | 27.42 lm | ||
| Mean | 9.47 e | 6.95 c | 172.27 c | 27.88 c | ||
| Mean | 0.0 | 9.68 f | 5.36 f | 179.07 d | 30.50 f | |
| 50.0 | 9.46 f | 6.28 e | 152.71 e | 29.82 f | ||
| 100.0 | 9.47 f | 6.26 e | 156.21 e | 37.72 bc | ||
| 200.0 | 8.75 g | 6.73 de | 169.83 de | 31.28 ef | ||
| Mean | 9.34 c | 6.16 c | 164.45 c | 32.33 c | ||
| 100 mM | Zn | 0.0 | 10.10 efg | 5.09 ef | 280.60 bcd | 29.77 jkl |
| 50.0 | 9.52 gh | 5.16 ef | 266.01 cde | 24.67 m | ||
| 100.0 | 9.21 hi | 4.51 f | 197.10 fg | 43.01 bcd | ||
| 200.0 | 9.86 e–h | 8.78 abc | 172.13 ghi | 48.58 a | ||
| Mean | 9.67 d | 5.89 d | 228.96 b | 36.51 b | ||
| Zn-NPs | 0.0 | 11.26 d | 8.22 c | 132.17 k | 40.64 cde | |
| 50.0 | 9.89 e–h | 8.79 abc | 179.27 fgh | 37.78 e–h | ||
| 100.0 | 10.36 ef | 8.84 abc | 139.75 k | 34.81 ghi | ||
| 200.0 | 10.67 de | 8.53 bc | 139.87 k | 29.01 kl | ||
| Mean | 10.55 c | 8.60 a | 147.77 d | 35.56 b | ||
| Mean | 0.0 | 10.68 d | 6.65 de | 206.39 c | 35.21 cd | |
| 50.0 | 9.71 f | 6.98 d | 222.64 c | 31.23 ef | ||
| 100.0 | 9.78 ef | 6.68 de | 168.42 de | 38.91 b | ||
| 200.0 | 10.26 de | 8.65 b | 156.00 e | 38.80 b | ||
| Mean | 10.11 b | 7.24 b | 188.36 b | 36.03 b | ||
| 200 mM | Zn | 0.0 | 14.12 ab | 6.90 d | 315.03 a | 38.61 efg |
| 50.0 | 13.36 bc | 6.84 d | 289.37 bc | 44.49 bc | ||
| 100.0 | 13.75 ab | 9.55 a | 186.76 fgh | 42.94 bcd | ||
| 200.0 | 14.52 a | 9.03 ab | 262.14 de | 28.63 klm | ||
| Mean | 13.94 a | 8.08 b | 263.32 a | 36.91 ab | ||
| Zn-NPs | 0.0 | 13.98 ab | 8.45 bc | 203.02 f | 33.45 ij | |
| 50.0 | 12.64 c | 8.69 bc | 251.40 e | 34.72 ghi | ||
| 100.0 | 9.99 e–h | 8.89 abc | 297.66 ab | 40.53 cd | ||
| 200.0 | 9.92 e–h | 8.38 bc | 256.75 de | 38.95 def | ||
| Mean | 11.63 b | 8.60 a | 252.21 a | 38.67 a | ||
| Mean | 0.0 | 14.05 a | 7.67 c | 259.02 ab | 36.03 cd | |
| 50.0 | 13.00 b | 7.76 c | 270.38 a | 39.61 ab | ||
| 100.0 | 11.87 c | 9.22 a | 242.21 b | 41.74 a | ||
| 200.0 | 12.22 c | 8.71 b | 259.44 ab | 33.79 de | ||
| Mean | 12.79 a | 8.34 a | 257.77 a | 37.79 a | ||
| Means of Zn treatments | Zn | 0.0 | 11.41 a | 5.75 d | 249.10 a | 32.33 d |
| 50.0 | 10.47 c | 5.92 d | 229.85 b | 30.68 d | ||
| 100.0 | 10.73 bc | 6.41 c | 175.96 d | 43.04 a | ||
| 200.0 | 11.49 a | 7.70 b | 210.31 c | 40.89 b | ||
| Mean | 11.03 a | 6.44 b | 216.31 a | 36.73 a | ||
| Zn-NPs | 0.0 | 11.53 a | 7.37 b | 180.56 d | 35.50 c | |
| 50.0 | 10.98 b | 8.09 a | 200.64 c | 36.43 c | ||
| 100.0 | 10.02 d | 8.36 a | 201.93 c | 35.87 c | ||
| 200.0 | 9.34 e | 8.37 a | 179.87 d | 28.35 e | ||
| Mean | 10.46 b | 8.05 a | 190.75 b | 34.04 b | ||
| Means of Zn concentration (ppm) | 0.0 | 11.47 a | 6.56 d | 214.83 a | 33.91 b | |
| 50.0 | 10.72 b | 7.01 c | 215.24 a | 33.55 b | ||
| 100.0 | 10.38 c | 7.39 b | 188.95 b | 39.46 a | ||
| 200.0 | 10.41 c | 8.03 a | 195.09 b | 34.62 b | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Türkoğlu, A.; Haliloğlu, K.; Ekinci, M.; Turan, M.; Yildirim, E.; Öztürk, H.İ.; Stansluos, A.A.L.; Nadaroğlu, H.; Piekutowska, M.; Niedbała, G. Zinc Oxide Nanoparticles: An Influential Element in Alleviating Salt Stress in Quinoa (Chenopodium quinoa L. Cv Atlas). Agronomy 2024, 14, 1462. https://doi.org/10.3390/agronomy14071462
Türkoğlu A, Haliloğlu K, Ekinci M, Turan M, Yildirim E, Öztürk Hİ, Stansluos AAL, Nadaroğlu H, Piekutowska M, Niedbała G. Zinc Oxide Nanoparticles: An Influential Element in Alleviating Salt Stress in Quinoa (Chenopodium quinoa L. Cv Atlas). Agronomy. 2024; 14(7):1462. https://doi.org/10.3390/agronomy14071462
Chicago/Turabian StyleTürkoğlu, Aras, Kamil Haliloğlu, Melek Ekinci, Metin Turan, Ertan Yildirim, Halil İbrahim Öztürk, Atom Atanasio Ladu Stansluos, Hayrunnisa Nadaroğlu, Magdalena Piekutowska, and Gniewko Niedbała. 2024. "Zinc Oxide Nanoparticles: An Influential Element in Alleviating Salt Stress in Quinoa (Chenopodium quinoa L. Cv Atlas)" Agronomy 14, no. 7: 1462. https://doi.org/10.3390/agronomy14071462
APA StyleTürkoğlu, A., Haliloğlu, K., Ekinci, M., Turan, M., Yildirim, E., Öztürk, H. İ., Stansluos, A. A. L., Nadaroğlu, H., Piekutowska, M., & Niedbała, G. (2024). Zinc Oxide Nanoparticles: An Influential Element in Alleviating Salt Stress in Quinoa (Chenopodium quinoa L. Cv Atlas). Agronomy, 14(7), 1462. https://doi.org/10.3390/agronomy14071462