
An Earthworm Peptide Alters Soil Nematode, Microbial, and Nutrient Dynamics: A Novel Mechanism of Soil Food Web Feedbacks

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sampling
2.2. Soil Nematodes and Physicochemical Analysis
2.3. DNA Extraction and PCR Amplification
2.4. Sequencing Data Analysis
2.5. Statistics and Analysis
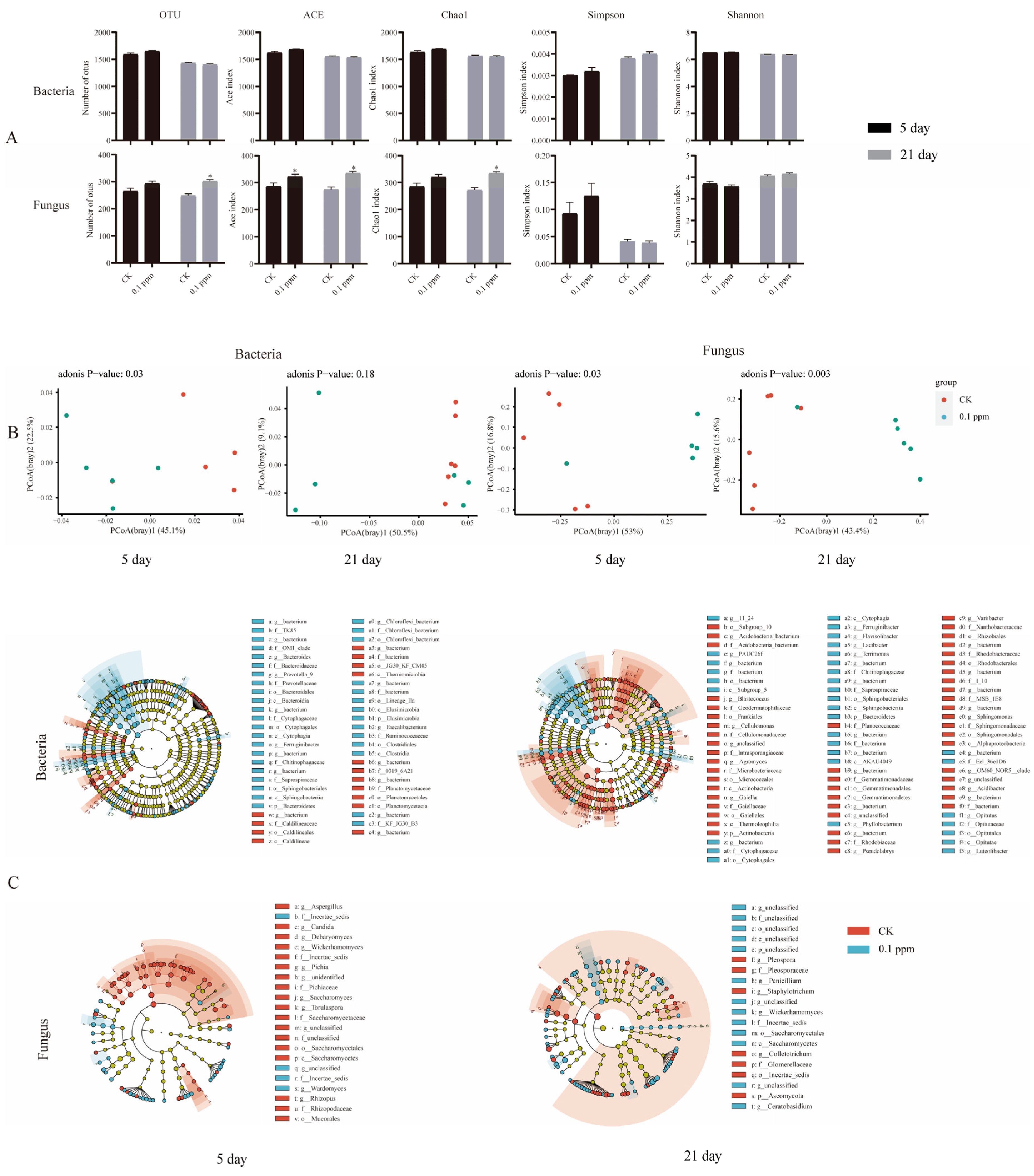
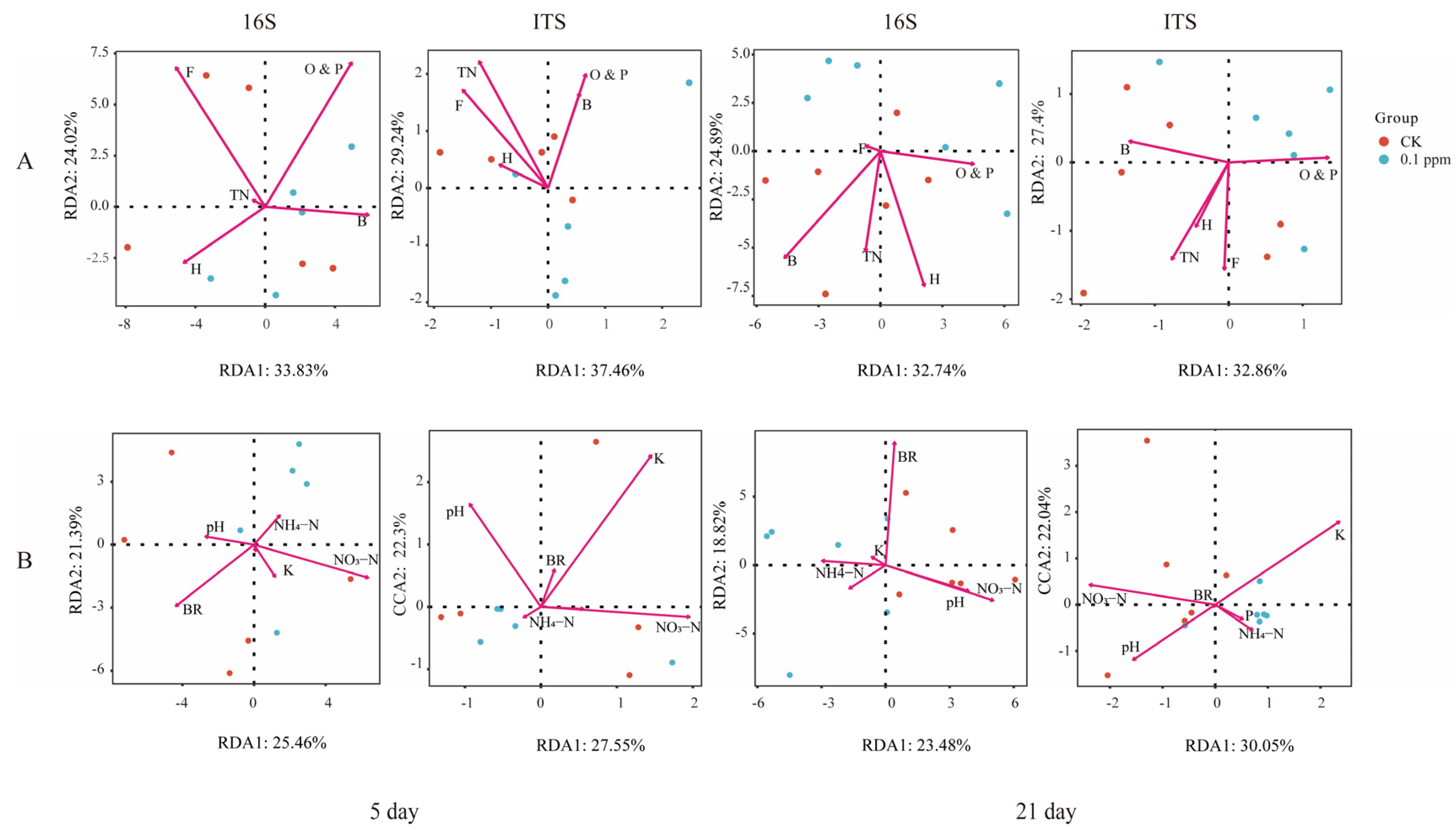
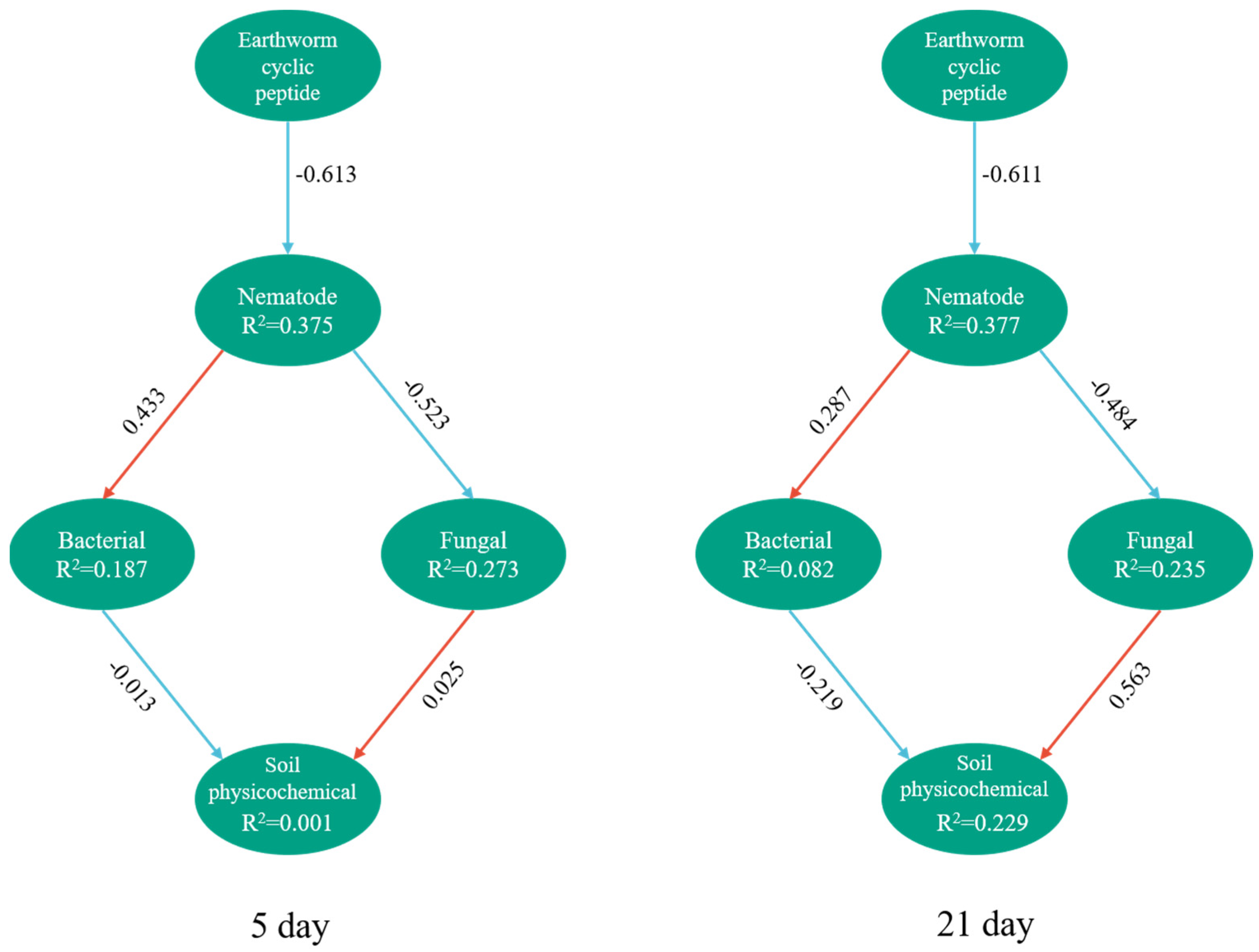
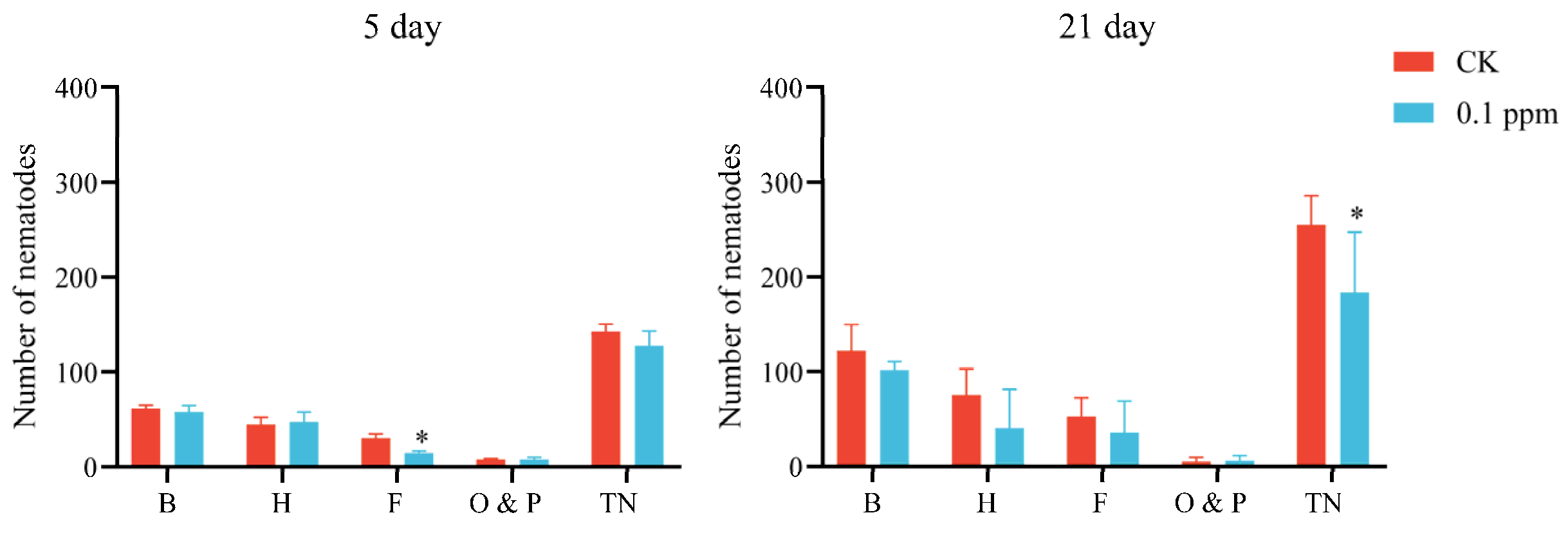
3. Results
3.1. Soil Physicochemical Properties
3.2. Soil Nematodes
3.3. Soil Microbial Community
3.4. Relationships between Soil Nematodes and Soil Factors with Microbial Communities
3.5. PLS-PM Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chapin, F.S.; Walker, B.H.; Hobbs, R.J.; Hooper, D.U.; Lawton, J.H.; Sala, O.E.; Tilman, D. Biotic Control over the Functioning of Ecosystems. Science 1997, 277, 500–504. [Google Scholar] [CrossRef]
- Chatterjee, S.; Isaia, M.; Venturino, E. Spiders as biological controllers in the agroecosystem. J. Theor. Biol. 2009, 258, 352–362. [Google Scholar] [CrossRef] [PubMed]
- Campanella, F.; Auster, P.J.; Taylor, J.C.; Muñoz, R.C. Dynamics of predator-prey habitat use and behavioral interactions over diel periods at sub-tropical reefs. PLoS ONE 2019, 14, e0211886. [Google Scholar] [CrossRef] [PubMed]
- Newsome, T.M.; Greenville, A.C.; Ćirović, D.; Dickman, C.R.; Johnson, C.N.; Krofel, M.; Letnic, M.; Ripple, W.J.; Ritchie, E.G.; Stoyanov, S.; et al. Top predators constrain mesopredator distributions. Nat. Commun. 2017, 8, 15469. [Google Scholar] [CrossRef] [PubMed]
- Paine, R.T. Food web complexity and species diversity. Am. Nat. 1966, 100, 65–75. [Google Scholar] [CrossRef]
- Holling, C.S. Cross-scale morphology, geometry, and dynamics of ecosystems. Ecol. Monogr. 1992, 62, 447–502. [Google Scholar] [CrossRef]
- Le Bayon, R.-C.; Bullinger-Weber, G.; Schomburg, A.; Turberg, P.; Schlaepfer, R.; Guenat, C. Earthworms as Ecosystem Engineers: A Review. In Earthworms: Types, Roles and Research; NOVA Science Publishers: New York, NY, USA, 2017; pp. 129–178. [Google Scholar]
- García-Palacios, P.; Maestre, F.T.; Bradford, M.A.; Reynolds, J.F. Earthworms modify plant biomass and nitrogen capture under conditions of soil nutrient heterogeneity and elevated atmospheric CO2 concentrations. Soil Biol. Biochem. 2014, 78, 182–188. [Google Scholar] [CrossRef]
- Edwards, C.A.; Arancon, N.Q.; Bohlen, P.J.; Hendrix, P. Biology and Ecology of Earthworms; Springer: Berlin/Heidelberg, Germany, 2022. [Google Scholar]
- Bernard, L.; Chapuis-Lardy, L.; Razafimbelo, T.; Razafindrakoto, M.; Pablo, A.-L.; Legname, E.; Poulain, J.; Brüls, T.; O’Donohue, M.; Brauman, A. Endogeic earthworms shape bacterial functional communities and affect organic matter mineralization in a tropical soil. ISME J. 2012, 6, 213–222. [Google Scholar] [CrossRef]
- Rui, X.U.E.; Chong, W.; Xuelian, L.I.U.; Mengli, L.I.U. Earthworm regulation of nitrogen pools and dynamics and marker genes of nitrogen cycling: A meta-analysis. Pedosphere 2022, 32, 131–139. [Google Scholar]
- Don, A.; Steinberg, B.; Schöning, I.; Pritsch, K.; Joschko, M.; Gleixner, G.; Schulze, E.-D. Organic carbon sequestration in earthworm burrows. Soil Biol. Biochem. 2008, 40, 1803–1812. [Google Scholar] [CrossRef]
- Tiunov, A.V.; Bonkowski, M.; Tiunov, J.A.; Scheu, S. Microflora, Protozoa and Nematoda in Lumbricus terrestris burrow walls: A laboratory experiment. Pedobiologia 2001, 45, 46–60. [Google Scholar] [CrossRef]
- Huang, K.; Xia, H. Role of earthworms’ mucus in vermicomposting system: Biodegradation tests based on humification and microbial activity. Sci. Total Environ. 2018, 610, 703–708. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Li, C.; Liu, T.; Li, T.; Hu, F.; Li, H.; Jiao, J. Earthworm mucus interfere in the behavior and physiology of bacterial-feeding nematodes. Appl. Soil Ecol. 2019, 143, 107–115. [Google Scholar] [CrossRef]
- Mulder, C.; Vonk, J.A. Nematode traits and environmental constraints in 200 soil systems: Scaling within the 60–6000 μm body size range: Ecological Archives E092-171. Ecology 2011, 92, 2004. [Google Scholar] [CrossRef]
- Bouwman, L.A.; Zwart, K.B. The ecology of bacterivorous protozoans and nematodes in arable soil. Agric. Ecosyst. Environ. 1994, 51, 145–160. [Google Scholar] [CrossRef]
- Jiang, Y.; Sun, B.; Jin, C.; Wang, F. Soil aggregate stratification of nematodes and microbial communities affects the metabolic quotient in an acid soil. Soil Biol. Biochem. 2013, 60, 1–9. [Google Scholar] [CrossRef]
- Djigal, D.; Baudoin, E.; Philippot, L.; Brauman, A.; Villenave, C. Shifts in size, genetic structure and activity of the soil denitrifier community by nematode grazing. Eur. J. Soil Biol. 2010, 46, 112–118. [Google Scholar] [CrossRef]
- Kane, J.L.; Kotcon, J.B.; Freedman, Z.B.; Morrissey, E.M. Fungivorous nematodes drive microbial diversity and carbon cycling in soil. Ecology 2023, 104, e3844. [Google Scholar] [CrossRef]
- Valembois, P.; Roch, P.; Lassegues, M.; Cassand, P. Antibacterial activity of the hemolytic system from the earthworm Eisenia fetida andrei. J. Invertebr. Pathol. 1982, 40, 21–27. [Google Scholar] [CrossRef]
- Liu, Y.-Q.; Sun, Z.-J.; Wang, C.; Li, S.-J.; Liu, Y.-Z. Purification of a novel antibacterial short peptide in earthworm Eisenia foetida. Acta Biochim. Et Biophys. Sin. 2004, 36, 297–302. [Google Scholar] [CrossRef]
- Liebeke, M.; Strittmatter, N.; Fearn, S.; Morgan, A.J.; Kille, P.; Fuchser, J.; Wallis, D.; Palchykov, V.; Robertson, J.; Lahive, E.; et al. Unique metabolites protect earthworms against plant polyphenols. Nat. Commun. 2015, 6, 7869. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Yu, F.; Zhou, M.; Xu, J.; Wu, J.; Hu, F.; Li, H.; Whalen, J.K. Soil fauna actively change their diet to survive stress. Soil Biol. Biochem. 2021, 162, 108435. [Google Scholar] [CrossRef]
- Van Bezooijen, J. Methods and Techniques for Nematology; Wageningen University: Wageningen, The Netherlands, 2006. [Google Scholar]
- Liu, Y.-R.; Delgado-Baquerizo, M.; Wang, J.-T.; Hu, H.-W.; Yang, Z.; He, J.-Z. New insights into the role of microbial community composition in driving soil respiration rates. Soil Biol. Biochem. 2018, 118, 35–41. [Google Scholar] [CrossRef]
- Sparks, D.L.; Page, A.L.; Helmke, P.A.; Loeppert, R.H.; Mulvaney, R.L. Nitrogen—Inorganic forms. In Methods of Soil Analysis Part; SSSA: Madison, Wisconsin, 1996; pp. 1149–1224. [Google Scholar]
- Yu, F.; Yan, Y.; Dong, Q.; Jiang, C.; Zu, C.; Shen, J. The Changes in Rhizosphere Metabolome and Microbiota Are the Main Direct Obstacles to Continuous Cropping in Tobacco (Nicotiana tabacum L.). Agronomy 2023, 13, 964. [Google Scholar] [CrossRef]
- Chen, T.; Liu, Y.X.; Huang, L. ImageGP: An easy-to-use data visualization web server for scientific researchers. Imeta 2022, 1, e5. [Google Scholar] [CrossRef]
- Ren, C.; Zhou, Z.; Guo, Y.; Yang, G.; Zhao, F.; Wei, G.; Han, X.; Feng, L.; Feng, Y.; Ren, G. Contrasting patterns of microbial community and enzyme activity between rhizosphere and bulk soil along an elevation gradient. Catena 2021, 196, 104921. [Google Scholar] [CrossRef]
- Colgrave Michelle, L.; Huang, Y.-H.; Craik David, J.; Kotze Andrew, C. Cyclotide Interactions with the Nematode External Surface. Antimicrob. Agents Chemother. 2010, 54, 2160–2166. [Google Scholar] [CrossRef]
- Wright, K.A. Nematode chemosensilla: Form and function. J. Nematol. 1983, 15, 151–158. [Google Scholar]
- Basyoni, M.M.A.; Rizk, E.M.A. Nematodes ultrastructure: Complex systems and processes. J. Parasit. Dis. 2016, 40, 1130–1140. [Google Scholar] [CrossRef]
- Backeljau, T.; Brown, D.; Decraemer, W.; Karanastasi, E. Review of the ultrastructure of the nematode body cuticle and its phylogenetic interpretation. Biol. Rev. 2003, 78, 465–510. [Google Scholar] [CrossRef]
- Schafer, W. Nematode nervous systems. Curr. Biol. 2016, 26, R955–R959. [Google Scholar] [CrossRef]
- Neilson, R.; Caul, S.; Fraser, F.C.; King, D.; Mitchell, S.M.; Roberts, D.M.; Giles, M.E. Microbial community size is a potential predictor of nematode functional group in limed grasslands. Appl. Soil Ecol. 2020, 156, 103702. [Google Scholar] [CrossRef]
- Djigal, D.; Brauman, A.; Diop, T.A.; Chotte, J.L.; Villenave, C. Influence of bacterial-feeding nematodes (Cephalobidae) on soil microbial communities during maize growth. Soil Biol. Biochem. 2004, 36, 323–331. [Google Scholar] [CrossRef]
- Dackman, C.; Jansson, H.-B.; Nordbring-Hertz, B. Nematophagous fungi and their activities in soil. In Soil biochemistry; CRC Press: Boca Raton, FL, USA, 2021; pp. 95–130. [Google Scholar]
- Li, N.; Pan, F.-j.; Han, X.-Z.; Zhang, B. Development of soil food web of microbes and nematodes under different agricultural practices during the early stage of pedogenesis of a Mollisol. Soil Biol. Biochem. 2016, 98, 208–216. [Google Scholar] [CrossRef]
- Zhou, D.; Feng, H.; Schuelke, T.; De Santiago, A.; Zhang, Q.; Zhang, J.; Luo, C.; Wei, L. Rhizosphere Microbiomes from Root Knot Nematode Non-infested Plants Suppress Nematode Infection. Microb. Ecol. 2019, 78, 470–481. [Google Scholar] [CrossRef]
- Song, L.; Ping, X.; Mao, Z.; Zhao, J.; Yang, Y.; Li, Y.; Xie, B.; Ling, J. Variation and stability of rhizosphere bacterial communities of Cucumis crops in association with root-knot nematodes infestation. Front. Plant Sci. 2023, 14. [Google Scholar] [CrossRef]
- Okada, H.; Harada, H.; Kadota, I. Fungal-feeding habits of six nematode isolates in the genus Filenchus. Soil Biol. Biochem. 2005, 37, 1113–1120. [Google Scholar] [CrossRef]
- Zheng, Q.; Hu, Y.; Zhang, S.; Noll, L.; Böckle, T.; Dietrich, M.; Herbold, C.W.; Eichorst, S.A.; Woebken, D.; Richter, A.; et al. Soil multifunctionality is affected by the soil environment and by microbial community composition and diversity. Soil Biol. Biochem. 2019, 136, 107521. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Smith, P.; Smith, J.U.; Moncrieff, J.B. Incorporating microorganisms as decomposers into models to simulate soil organic matter decomposition. Geoderma 2005, 129, 139–146. [Google Scholar] [CrossRef]
- Caravaca, F.; Lozano, Z.; Rodríguez-Caballero, G.; Roldán, A. Spatial shifts in soil microbial activity and degradation of pasture cover caused by prolonged exposure to cement dust. Land Degrad. Dev. 2017, 28, 1329–1335. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, D.; Huang, R.; Huang, G.; Yuan, Y.; Fan, H. Effects of bacterial-feeding nematodes on soil microbial activity and the microbial community in oil-contaminated soil. J. Environ. Manag. 2019, 234, 424–430. [Google Scholar] [CrossRef]
- Eylar, O.R.; Schmidt, E.L. A Survey of Heterotrophic Micro-Organisms from Soil for Ability to Form Nitrite and Nitrate. Microbiology 1959, 20, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Maeda, K.; Spor, A.; Edel-Hermann, V.; Heraud, C.; Breuil, M.-C.; Bizouard, F.; Toyoda, S.; Yoshida, N.; Steinberg, C.; Philippot, L. N2O production, a widespread trait in fungi. Sci. Rep. 2015, 5, 9697. [Google Scholar] [CrossRef] [PubMed]
- Kasana, R.C.; Panwar, N.R.; Burman, U.; Pandey, C.B.; Kumar, P. Isolation and identification of two potassium solubilizing fungi from arid soil. Int. J. Curr. Microbiol. App. Sci. 2017, 6, 1752–1762. [Google Scholar] [CrossRef]
- Jordan, C.F.; Jordan, C.F. Ecosystem Control: A Top-Down View. Evolution from a Thermodynamic Perspective. In Implications for Species Conservation and Agricultural Sustainability; Springer: Berlin/Heidelberg, Germany, 2022; pp. 47–62. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Treatment | pH | NH4-N | NO3-N | AK | AP | BR |
|---|---|---|---|---|---|---|---|
| 5 days | CK | 7.9 ± 0.03 | 4.84 ± 0.14 | 38.71 ± 0.94 | 284.78 ± 2.51 | 12.63 ± 0.40 | 2.34 ± 0.08 |
| 0.1 ppm | 7.84 ± 0.02 | 4.86 ± 0.22 | 38.85 ± 0.44 | 284.53 ± 1.95 | 11.91 ± 0.20 | 1.90 ± 0.18 * | |
| 21 days | CK | 7.83 ± 0.04 | 2.46 ± 0.15 | 59.32 ± 1.58 | 239.59 ± 2.43 | 15.20 ± 0.38 | 2.19 ± 0.13 |
| 0.1 ppm | 7.77 ± 0.01 | 2.75 ± 0.25 | 51.88 ± 1.58 * | 240.80 ± 1.87 | 15.68 ± 0.23 | 2.06 ± 0.26 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Qi, Y.; Yan, Y.; Xia, H.; Dong, Q.; Jiang, C.; Zu, C.; Shen, J. An Earthworm Peptide Alters Soil Nematode, Microbial, and Nutrient Dynamics: A Novel Mechanism of Soil Food Web Feedbacks. Agronomy 2024, 14, 435. https://doi.org/10.3390/agronomy14030435
Yu F, Qi Y, Yan Y, Xia H, Dong Q, Jiang C, Zu C, Shen J. An Earthworm Peptide Alters Soil Nematode, Microbial, and Nutrient Dynamics: A Novel Mechanism of Soil Food Web Feedbacks. Agronomy. 2024; 14(3):435. https://doi.org/10.3390/agronomy14030435
Chicago/Turabian StyleYu, Fei, Yaocheng Qi, Yifeng Yan, Hao Xia, Qing Dong, Chaoqiang Jiang, Chaolong Zu, and Jia Shen. 2024. "An Earthworm Peptide Alters Soil Nematode, Microbial, and Nutrient Dynamics: A Novel Mechanism of Soil Food Web Feedbacks" Agronomy 14, no. 3: 435. https://doi.org/10.3390/agronomy14030435
APA StyleYu, F., Qi, Y., Yan, Y., Xia, H., Dong, Q., Jiang, C., Zu, C., & Shen, J. (2024). An Earthworm Peptide Alters Soil Nematode, Microbial, and Nutrient Dynamics: A Novel Mechanism of Soil Food Web Feedbacks. Agronomy, 14(3), 435. https://doi.org/10.3390/agronomy14030435