
Thermal-Time Hazard Models of Seven Weed Species Germinability following Heat Treatment †


Abstract
1. Introduction
- (1)
- Determine the germination rates of important Californian weed species after exposure to solarization temperatures from 40 to 60 °C for a variety of time intervals;
- (2)
- Use the germination rates to create regression models simulating the responses of weed speeds to solarization temperatures.
2. Materials and Methods
2.1. Seed Collection and Preparation
2.2. Thermal Treatments
2.3. Seed Germination Tests
2.4. Verification of Germination Tests
2.5. Statistical Analysis
3. Results and Discussion
3.1. Tetrazolium Staining
3.2. Thermal Suppression of Germinability
3.3. Logistic Regression Models
3.4. Application to Field Conditions
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stapleton, J.J.; DeVay, J.E. Soil Solarization: A Non-Chemical Approach for Management of Plant Pathogens and Pests. Crop Prot. 1986, 5, 190–198. [Google Scholar] [CrossRef]
- Elmore, C.L.; Stapleton, J.J.; Bell, C.E. Soil Solarization: A Nonpesticidal Method for Controlling Diseases, Nematodes, and Weeds; FAO: Rome, Italy, 1997. [Google Scholar]
- Stapleton, J.J.; Wilen, C.A.; Molinar, R.H. Soil Solarization for Gardens and Landscapes; Taylor & Francis Ltd.: Abington, UK, 2008. [Google Scholar]
- Stapleton, J.J.; Molinar, R.H.; Lynn-Patterson, K.; McFeeters, S.K.; Shrestha, A. Methyl Bromide Alternatives… Soil Solarization Provides Weed Control for Limited-Resource and Organic Growers in Warmer Climates. Calif. Agric. 2008, 59, 84–89. [Google Scholar] [CrossRef]
- Dahlquist, R.M.; Prather, T.S.; Stapleton, J.J. Time and Temperature Requirements for Weed Seed Thermal Death. Weed Sci. 2007, 55, 619–625. [Google Scholar] [CrossRef]
- Hoyle, J.A.; Mcelroy, J.S. Relationship between Temperature and Heat Duration on Large Crabgrass (Digitaria sanguinalis), Virginia Buttonweed (Diodia virginiana), and Cock’s-Comb Kyllinga (Kyllinga squamulata) Seed Mortality. Weed Technol. 2012, 26, 800–806. [Google Scholar] [CrossRef]
- Melander, B.; Kristensen, J.K. Soil Steaming Effects on Weed Seedling Emergence under the Influence of Soil Type, Soil Moisture, Soil Structure and Heat Duration. Ann. Appl. Biol. 2011, 158, 194–203. [Google Scholar] [CrossRef]
- Melander, B.; Jørgensen, M.H. Soil Steaming to Reduce Intrarow Weed Seedling Emergence. Weed Res. 2005, 45, 202–211. [Google Scholar] [CrossRef]
- Van Loenen, M.C.A.; Turbett, Y.; Mullins, C.E.; Feilden, N.E.H.; Wilson, M.J.; Leifert, C.; Seel, W.E. Low Temperature—Short Duration Steaming of Soil Kills Soil-Borne Pathogens, Nematode Pests and Weeds. Eur. J. Plant Pathol. 2003, 109, 993–1002. [Google Scholar] [CrossRef]
- Horowitz, M.; Taylorson, R.B. Effect of High Temperatures on Imbibition, Germination, and Thermal Death of Velvetleaf (Abutilon threophrasti) Seeds. Can. J. Bot. 1983, 61, 2269–2276. [Google Scholar] [CrossRef]
- Elmore, C.L.; Roncoroni, J.A.; Giraud, D.D. Perennial Weeds Respond to Control by Soil Solarization. Calif. Agric. 1993, 47, 19–22. [Google Scholar] [CrossRef]
- Webster, T.M. High Temperatures and Durations of Exposure Reduce Nutsedge (Cyperus spp.) Tuber Viability. Weed Sci. 2003, 51, 1010–1015. [Google Scholar] [CrossRef]
- El-Keblawy, A.; Al-Hamadi, F. Assessment of the Differential Response of Weeds to Soil Solarization by Two Methods. Weed Biol. Manag. 2009, 9, 72–78. [Google Scholar] [CrossRef]
- Egley, G. High-Temperature Effects on Germination and Survival of Weed Seeds in Soil. Weed Sci. 1990, 38, 429–435. [Google Scholar] [CrossRef]
- Rubin, B.; Benjamin, A. Solar Heating of the Soil: Involvement of Environmental Factors in the Weed Control Process. Weed Sci. 1984, 32, 138–142. [Google Scholar] [CrossRef]
- Pullman, G.S.; DeVay, J.E.; Garber, R.H. Soil Solarization and Thermal Death: A Logarithmic Relationship between Time and Temperature for Four Soilborne Plant Pathogens. Phytopathology 1981, 71, 959–964. [Google Scholar] [CrossRef]
- Yildiz, A.; Benlioǧlu, S.; Boz, Ö.; Benlioǧlu, K. Use of Different Plastics for Soil Solarization in Strawberry Growth and Time-Temperature Relationships for the Control of Macrophomina phaseolina and Weeds. Phytoparasitica 2010, 38, 463–473. [Google Scholar] [CrossRef]
- Fennimore, S.A. Weed control as part of soil disinfestation with fumigants and nonfumigants. In Proceedings of the 2012 CWSS Proceedings; California Weed Science Society: West Sacramento, CA, USA, 2012; pp. 118–123. [Google Scholar]
- Samtani, J.B.; Gilbert, C.; Ben Weber, J.; Subbarao, K.V.; Goodhue, R.E.; Fennimore, S.A. Effect of Steam and Solarization Treatments on Pest Control, Strawberry Yield, and Economic Returns Relative to Methyl Bromide Fumigation. HortScience 2012, 47, 64–70. [Google Scholar] [CrossRef]
- Wu, J.Y.; Dastgheib, F. Effects of Various Herbicides and Surfactants on Mallow (Malva Spp.). In Proceedings of the 18th Asian-Pacific Weed Science Society Conference; Asian-Pacific Weed Science Society: Beijing, China, 2001; pp. 589–593. [Google Scholar]
- Moravcova, L.; Dostalek, J. Contribution to the Biology of Germination of Four Species of Chenopodium album Agg. under Different Condition. Folia Geobot. Phytotaxon. 1989, 24, 431–439. [Google Scholar] [CrossRef]
- Mauromicale, G.; Lo Monaco, A.; Longo, A.M.G.; Restuccia, A. Soil Solarization, a Nonchemical Method to Control Branched Broomrape (Orobanche Ramosa) and Improve the Yield of Greenhouse Tomato. Weed Sci. 2005, 53, 877–883. [Google Scholar] [CrossRef]
- Nagelkerke, N.J.D. A Note on a General Definition of the Coefficient of Determination. Biometrika 1991, 78, 691–692. [Google Scholar] [CrossRef]
- Dahlquist-Willard, R.M.; Marshall, M.N.; Betts, S.L.; Tuell-Todd, C.C.; VanderGheynst, J.S.; Stapleton, J.J. Development and Validation of a Weibull–Arrhenius Model to Predict Thermal Inactivation of Black Mustard (Brassica nigra) Seeds under Fluctuating Temperature Regimens. Biosyst. Eng. 2016, 151, 350–360. [Google Scholar] [CrossRef]
- Peachey, A.R.E.; Pinkerton, J.N.; Ivors, K.L.; Miller, M.L.; Moore, L.W. Effect of Soil Solarization, over Crops, and Metham on Field Emergence and Survival of Buried Annual Bluegrass (Poa annua). Weed Technol. 2001, 15, 81–88. [Google Scholar] [CrossRef]
- Gamliel, A.; Austerweil, M.; Kritzman, G. Non-Chemical Approach to Soilborne Pest Management—Organic Amendments. Crop Prot. 2000, 19, 847–853. [Google Scholar] [CrossRef]
- Gamliel, A.; Stapleton, J.J. Effect of Chicken Compost or Ammonium Phosphate and Solarization on Pathogen Control, Rhizosphere Microorganisms, and Lettuce Growth. Plant Dis. 1993, 77, 886–891. [Google Scholar] [CrossRef]
- Simmons, C.W.; Higgins, B.; Staley, S.; Joh, L.D.; Simmons, B.A.; Singer, S.W.; Stapleton, J.J.; VanderGheynst, J.S. The Role of Organic Matter Amendment Level on Soil Heating, Organic Acid Accumulation, and Development of Bacterial Communities in Solarized Soil. Appl. Soil Ecol. 2016, 106, 37–46. [Google Scholar] [CrossRef]
- Marshall, M.N.; Rumsey, T.R.; Stapleton, J.J.; Vandergheynst, J.S. A Predictive Model for Soil Temperature during Solarization and Model Validation at Two California Field Sites. Trans. ASABE 2013, 56, 117–133. [Google Scholar] [CrossRef]
- Medina, C.; Cardemil, L. Prosopis chilensis Is a Plant Highly Tolerant to Heat Shock. Plant Cell Environ. 1993, 16, 305–310. [Google Scholar] [CrossRef]
- Coca, M.A.; Almoguera, C.; Jordano, J. Expression of Sunflower Low-Molecular-Weight Heat-Shock Proteins during Embryogenesis and Persistence after Germination: Localization and Possible Functional Implications. Plant Mol. Biol. 1994, 25, 479–492. [Google Scholar] [CrossRef]
- Horowitz, M.; Regev, Y.; Herzlinger, G. Solarization for Weed Control. Weed Sci. 1983, 31, 170–179. [Google Scholar] [CrossRef]
- Candido, V.; D’addabbo, T.; Miccolis, V.; Castronuovo, D. Weed Control and Yield Response of Soil Solarization with Different Plastic Films in Lettuce. Sci. Hortic. 2011, 130, 491–497. [Google Scholar] [CrossRef]
- Powles, S.B.; Charman, N.; Poole, F. Solar Heating (Solarization) of the Soil Surface: Effect on Weed Control, and Yield of Phaseolus vulgaris. Plant Prot. Q. 1988, 3, 31–35. [Google Scholar]
- Stapleton, J.J. Fumigation and Solarization Practice in Plasticulture Systems. HortTechnology 1996, 6, 189–192. [Google Scholar] [CrossRef]
- DeVay, J.E.; Stapleton, J.J.; Elmore, C.L. Soil solarization. In Proceedings of the First International Conference on Soil Solarization, Amman, Jordan, 19–25 February 1990; FAO: Rome, Italy, 1991. [Google Scholar]
- Porter, I.J.; Merriman, P.R. Effects of Solarization of Soil on Nematode and Fungal Pathogens at Two Sites in Victoria. Soil Biol. Biochem. 1983, 15, 39–44. [Google Scholar] [CrossRef]

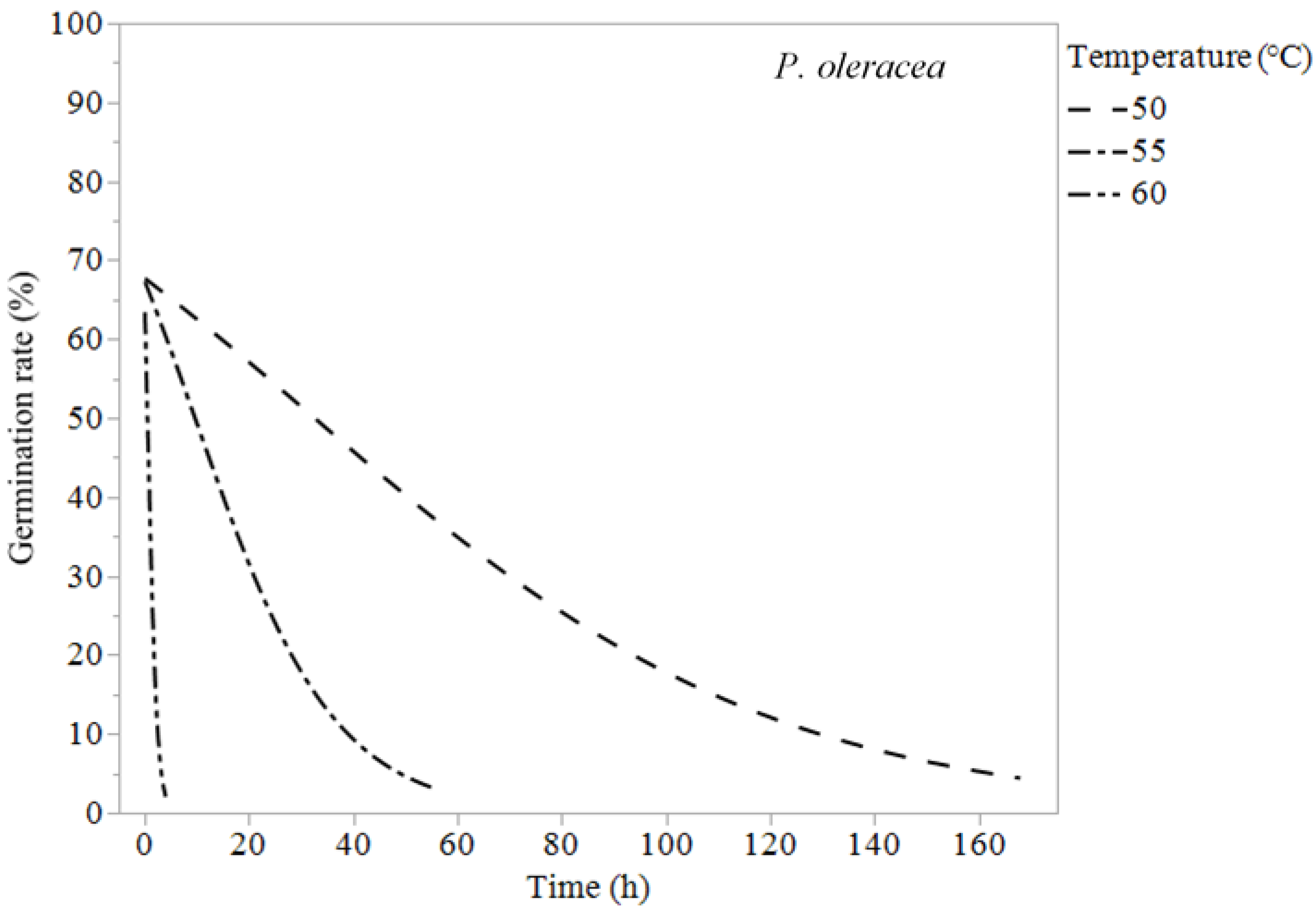
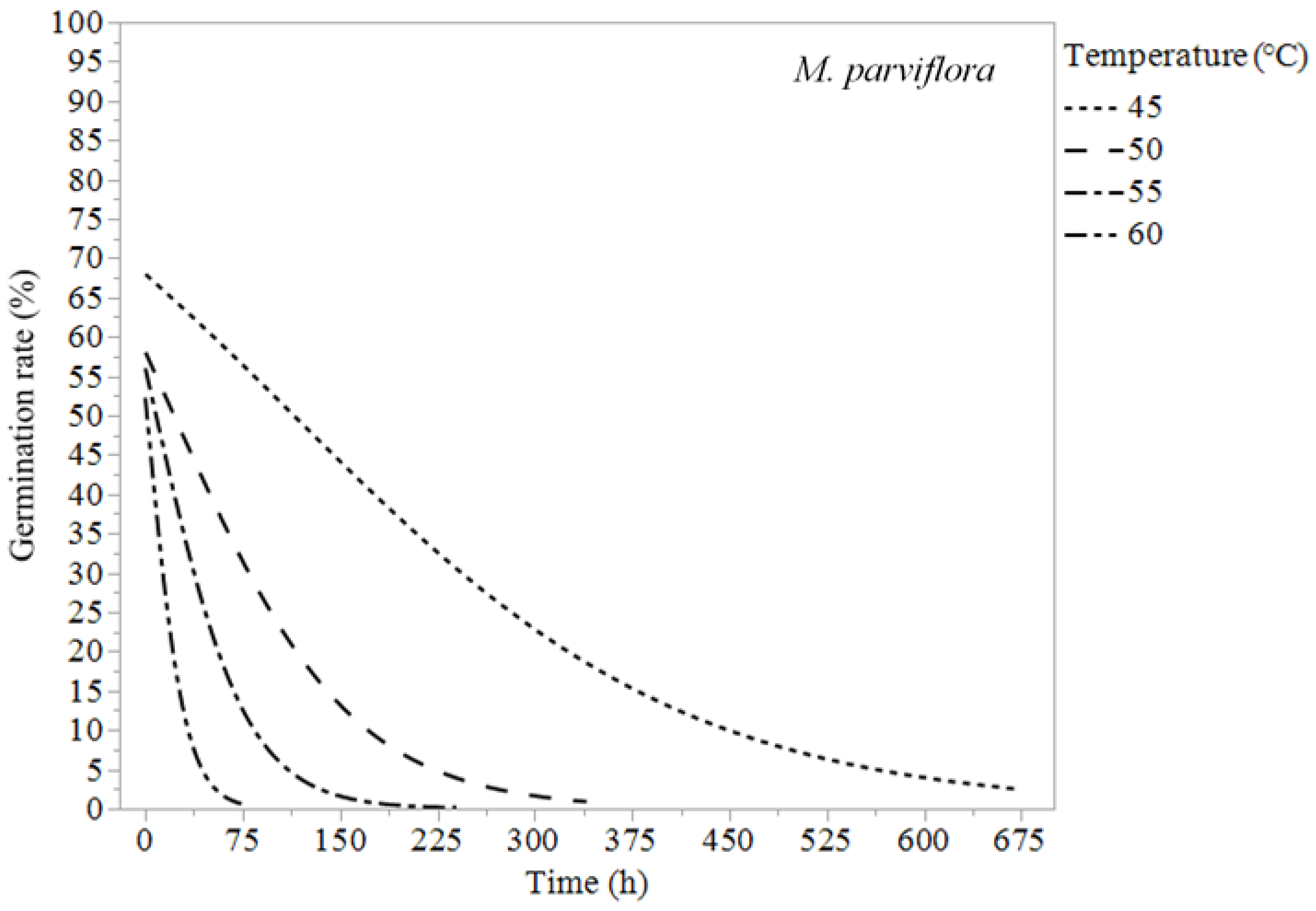

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Temperature (°C) | Treatment Germination Rate (%) | Control Germination Rate (%) | Exposure Time (h) | Pr > F |
|---|---|---|---|---|---|
| Amaranthus retroflexus | 40 | 39.6 | 47.4 | 340 | 0.757 |
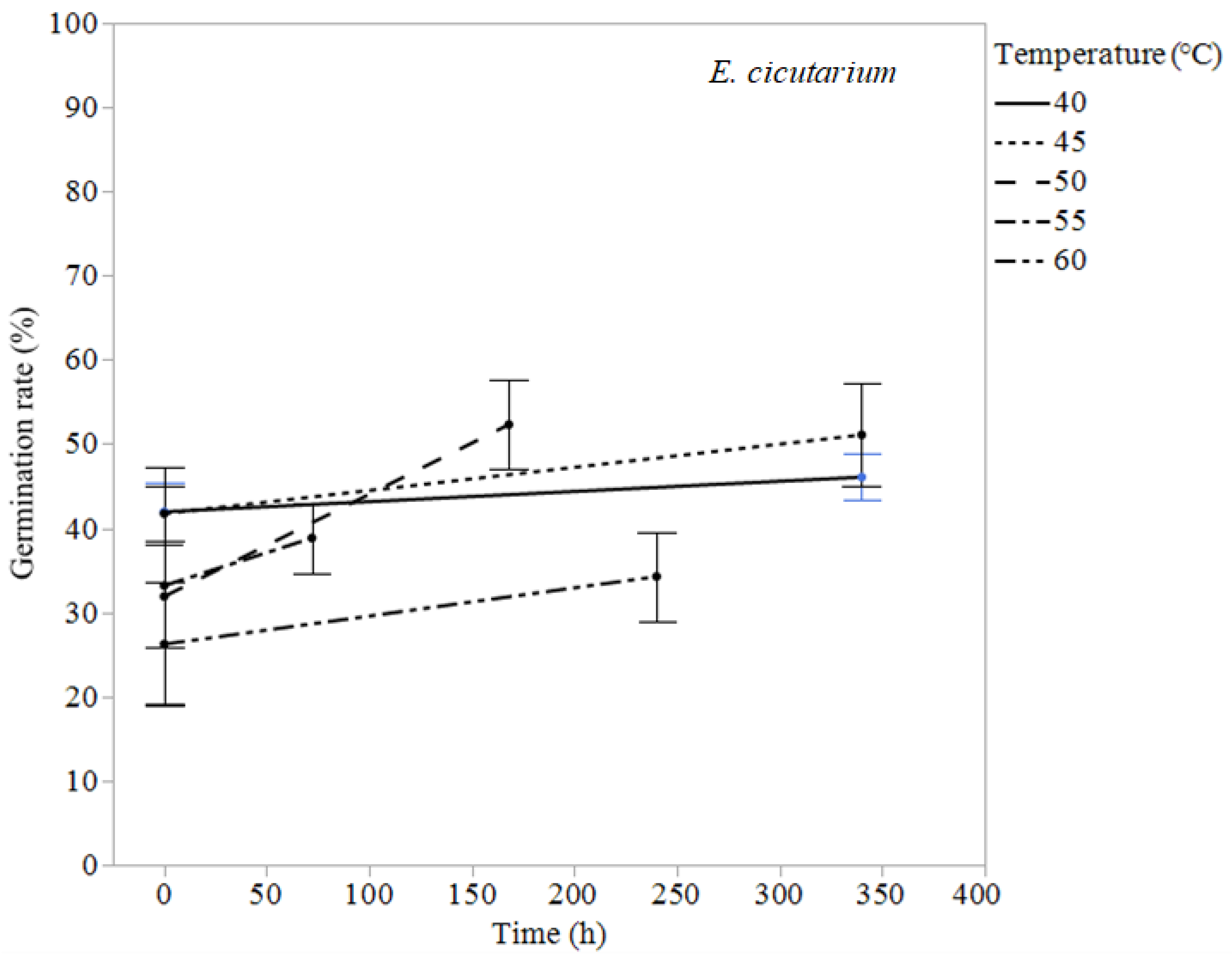
| Erodium cicutarium | 60 | 34.3 | 26.2 | 240 | 0.405 |
| 55 | 38.9 | 33.2 | 72 | 0.63 | |
| 50 | 52.3 | 31.9 | 168 | 0.051 | |
| 45 | 51.1 | 41.8 | 340 | 0.340 | |
| 40 | 46.1 | 42.0 | 340 | 0.446 | |
| Malva parviflora | 40 | 39.1 | 54.1 | 340 | 0.051 |
| Portulaca oleracea | 45 1 | 37.6 | 16.6 | 96 | 0.033 |
| Temperature (°C) | |||||
|---|---|---|---|---|---|
| 60 | 55 | 50 | 45 | 40 | |
| Species | Percentage | ||||
| Amaranthus retroflexus | 3.33 | 0.00 | 3.13 | 0.00 | N/a |
| Chenopodium murale | 0.00 | 2.78 | 7.14 | 5.66 | 2.44 |
| Malva parviflora | 0.00 | 8.57 | 8.57 | 0.00 | N/a |
| Picris echioides | 0.00 | 0.00 | 0.00 | 2.22 | 0.00 |
| Sonchus oleraceus | 0.00 | 0.00 | 3.33 | 2.22 | 0.00 |
| Temperature (°C) | |||||
|---|---|---|---|---|---|
| Species | 60 | 55 | 50 | 45 | 40 |
| Amaranthus retroflexus | 2 | 6 | 72 | 240 | N/a |
| Chenopodium murale | 2 | 4 | 72 2 | 196 | 432 |
| Erodium cicutarium | N/a | N/a | N/a | N/a | N/a |
| Malva parviflora | 74 | 240 2 | 340 | 672 2 | N/a |
| Picris echioides | 0.25 | 1 | 6 | 40 | 48 |
| Portulaca oleracea | 4 | 68 | 168 | N/a | N/a |
| Sonchus oleraceus | 0.25 | 2 | 6 | 24 | 48 |
| Species | Temp. (°C) | Parameter | Parameter Estimates (±SE) | Chi Square | p-Value | R2 |
|---|---|---|---|---|---|---|
| Malva parviflora | 45 | b0 | 0.7514 ± 0.0794 | 89.6 | <0.0001 | 0.2929 |
| b1 | −0.0066 ± 0.0005 | 194.94 | <0.0001 | |||
| 50 | b0 | 0.3256 ± 0.0894 | 13.26 | 0.0003 | 0.3362 | |
| b1 | −0.0147 ± 0.0011 | 164.9 | <0.0001 | |||
| 55 | b0 | 0.2241 ± 0.0830 | 7.28 | 0.007 | 0.346 | |
| b1 | −0.0295 ± 0.0025 | 144.02 | <0.0001 | |||
| 60 | b0 | 0.0913 ± 0.0895 | 1.04 | 0.3079 | 0.2521 | |
| b1 | −0.0694 ± 0.0078 | 79.05 | <0.0001 | |||
| Picris echioides | 40 | b0 | 0.8202 ± 0.1356 | 36.57 | <0.0001 | 0.4889 |
| b1 | −0.0810 ± 0.0064 | 157 | <0.0001 | |||
| 45 | b0 | 0.7037 ± 1161 | 36.7 | <0.0001 | 0.477 | |
| b1 | −0.1707 ± 0.0153 | 124.55 | <0.0001 | |||
| 50 | b0 | 1.0200 ± 0.1274 | 64.13 | <0.0001 | 0.5929 | |
| b1 | −1.9358 ± 0.1848 | 109.74 | <0.0001 | |||
| 55 | b0 | 0.5759 ± 0.2405 | 5.73 | 0.0167 | 0.6731 | |
| b1 | −9.7963 ± 2.0503 | 22.83 | <0.0001 | |||
| Portulaca oleracea | 50 | b0 | 0.7385 ± 0.1043 | 50.15 | <0.0001 | 0.3375 |
| b1 | −0.0227 ± 0.0017 | 181.13 | <0.0001 | |||
| 55 | b0 | 0.7194 ± 0.1118 | 41.40 | <0.0001 | 0.3941 | |
| b1 | −0.0749 ± 0.0061 | 149.66 | <0.0001 | |||
| 60 | b0 | 0.5493 ± 0.1170 | 22.04 | <0.0001 | 0.3592 | |
| b1 | −1.1040 ± 0.1026 | 115.84 | <0.0001 | |||
| Chenopodium murale | 40 | b0 | −0.5608 ± 0.0995 | 31.75 | <0.0001 | 0.1117 |
| b1 | −0.0054 ± 0.0007 | 63.78 | <0.0001 | |||
| 45 | b0 | −0.1601 ± 0.0979 | 2.68 | 0.1019 | 0.2176 | |
| b1 | −0.0158 ± 0.0016 | 98.45 | <0.0001 | |||
| 50 | b0 | −0.5661 ± 0.1041 | 29.55 | <0.0001 | 0.219 | |
| b1 | −0.0616 ± 0.0077 | 63.52 | <0.0001 | |||
| 55 | b0 | −0.3616 ± 0.1221 | 8.77 | 0.0031 | 0.27 | |
| b1 | −1.0843 ± 0.1387 | 61.09 | <0.0001 | |||
| 60 | b0 | −0.4465 ± 0.2156 | 4.29 | 0.0384 | 0.3161 | |
| b1 | −3.0051 ± 0.5978 | 25.27 | <0.0001 | |||
| Amaranthus retroflexus | 45 | b0 | 0.0063 ± 0.1001 | 0 | 0.9501 | 0.2958 |
| b1 | −0.0165 ± 0.0014 | 132.58 | <0.0001 | |||
| 50 | b0 | −0.0197 ± 0.0994 | 0.04 | 0.8426 | 0.2322 | |
| b1 | −0.0515 ± 0.0053 | 94.09 | <0.0001 | |||
| 55 | b0 | −0.3605 ± 0.1154 | 9.76 | 0.0018 | 0.2182 | |
| b1 | −0.6664 ± 0.0916 | 52.92 | <0.0001 | |||
| 60 | b0 | −0.7851 ± 0.1830 | 18.4 | <0.0001 | 0.3212 | |
| b1 | −3.5358 ± 0.6574 | 28.93 | <0.0001 | |||
| Sonchus oleraceus | 40 | b0 | 0.3914 ± 0.1130 | 12.01 | 0.0005 | 0.4121 |
| b1 | −0.1310 ± 0.0106 | 151.93 | <0.0001 | |||
| 45 | b0 | 1.2919 ± 0.1399 | 85.32 | <0.0001 | 0.5751 | |
| b1 | −0.2394 ± 0.0191 | 156.92 | <0.0001 | |||
| 50 | b0 | 0.2591 ± 0.1116 | 5.39 | 0.0202 | 0.4634 | |
| b1 | −1.6811 ± 0.1766 | 90.58 | <0.0001 | |||
| 55 | b0 | 0.6289 ± 0.1806 | 12.13 | 0.0005 | 0.6624 | |
| b1 | −8.0668 ± 1.2038 | 44.91 | <0.0001 |
| Temperature (°C) | LD20 ± 95% CI (h) | LD50 ± 95%CI (h) | LD90 ± 95% CI (h) | |
|---|---|---|---|---|
| Malva parviflora | 45 | 87.8 ± 19.1 | 215 ± 23.1 | 513 ± 54.0 |
| 50 | 31.8 ± 9.95 | 82.9 ± 10.9 | 211 ± 24.2 | |
| 55 | 15.5 ± 4.83 | 40.8 ± 5.58 | 105 ± 13.1 | |
| 60 | 6.07 ± 2.29 | 16.3 ± 2.81 | 43.1 ± 7.07 | |
| Picris echioides | 40 | 7.38 ± 2.63 | 17.9 ± 2.62 | 42.2 ± 4.64 |
| 45 | 3.30 ± 1.08 | 8.15 ± 1.17 | 19.6 ± 2.47 | |
| 50 | 0.34 ± 0.11 | 0.81 ± 0.12 | 1.84 ± 0.25 | |
| 55 | 0.05 ± 0.06 | 0.14 ± 0.06 | 0.33 ± 0.10 | |
| Portulaca oleracea | 50 | 25.2 ± 6.91 | 62.1 ± 7.11 | 148 ± 15.1 |
| 55 | 7.58 ± 2.39 | 18.7 ± 2.55 | 44.7 ± 5.22 | |
| 60 | 0.47 ± 0.17 | 1.19 ± 0.17 | 2.94 ± 0.38 | |
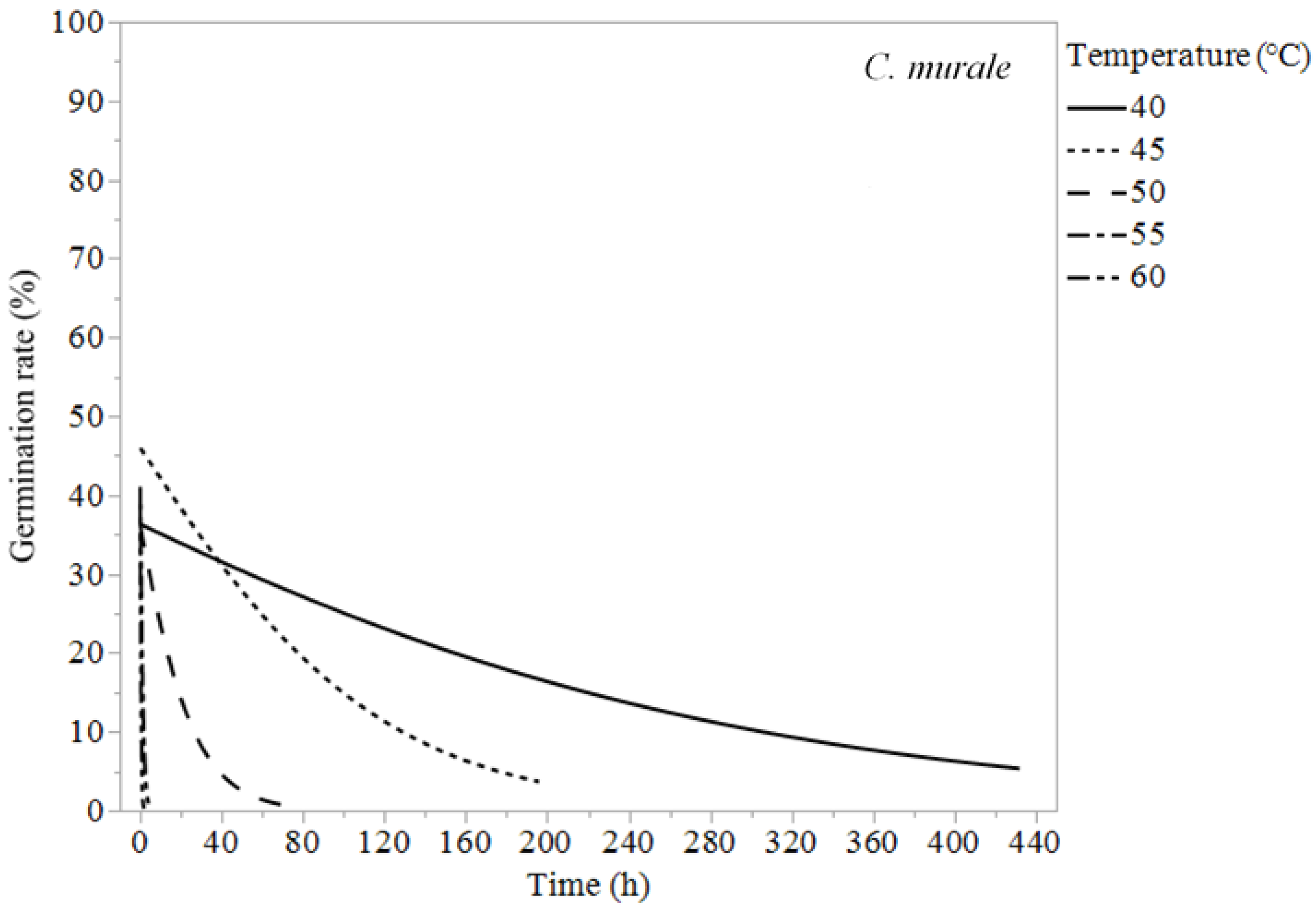
| Chenopodium murale | 40 | 61.9 ± 28.3 | 176 ± 31.0 | 508 ± 85.5 |
| 45 | 24.0 ± 10.2 | 66.2 ± 11.1 | 181 ± 26.2 | |
| 50 | 5.37 ± 2.95 | 15.3 ± 3.23 | 44.1 ± 7.89 | |
| 55 | 0.33 ± 0.19 | 0.91 ± 0.2 | 2.57 ± 0.46 | |
| 60 | 0.11 ± 0.14 | 0.32 ± 0.13 | 0.92 ± 0.25 | |
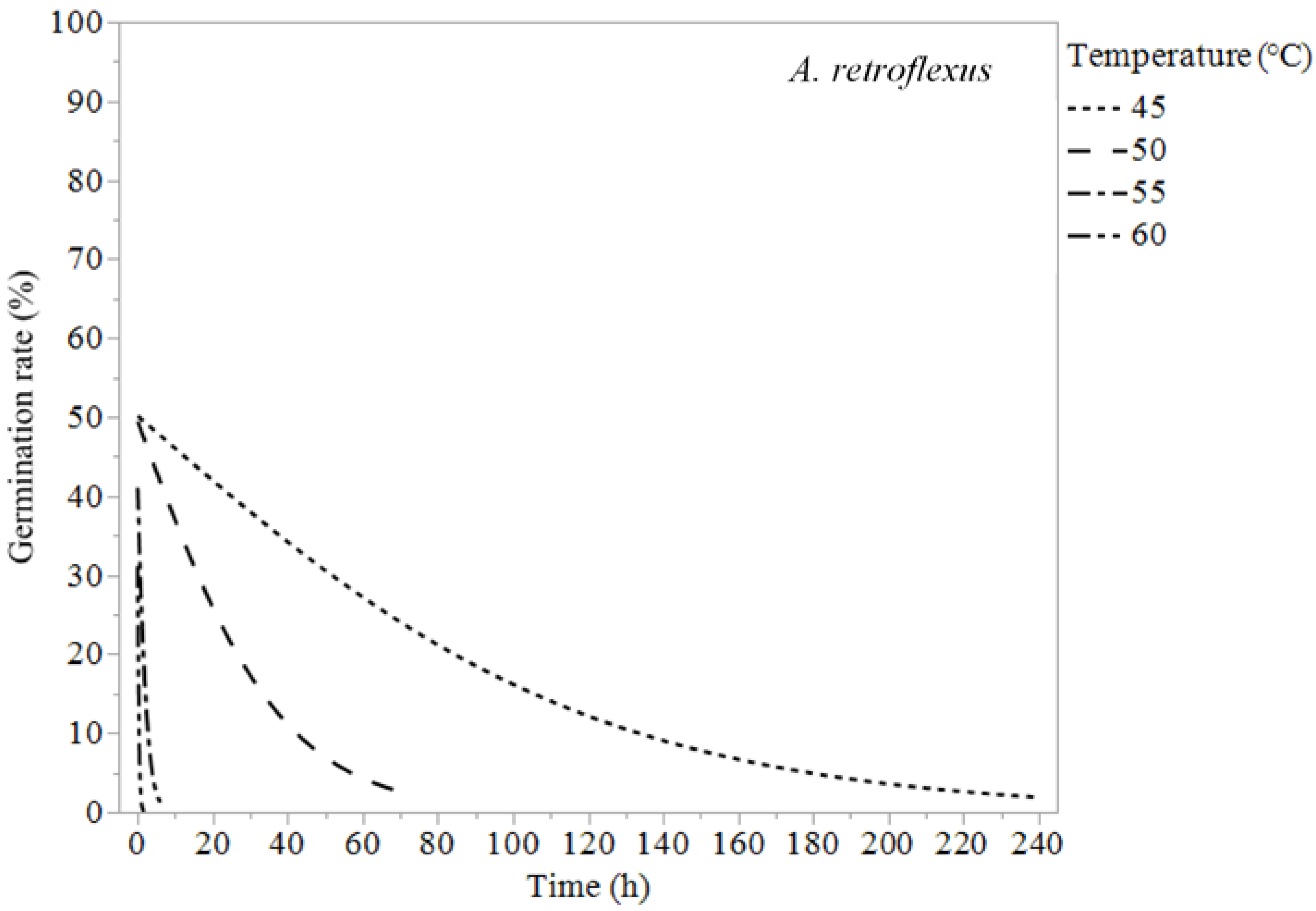
| Amaranthus retroflexus | 45 | 24.6 ± 9.86 | 66.7 ± 10.3 | 179 ± 22.2 |
| 50 | 7.81 ± 3.14 | 21.2 ± 3.46 | 57.0 ± 8.34 | |
| 55 | 0.53 ± 0.29 | 1.49 ± 0.32 | 4.19 ± 0.79 | |
| 60 | 0.09 ± 0.10 | 0.25 ± 0.09 | 0.75 ± 0.19 | |
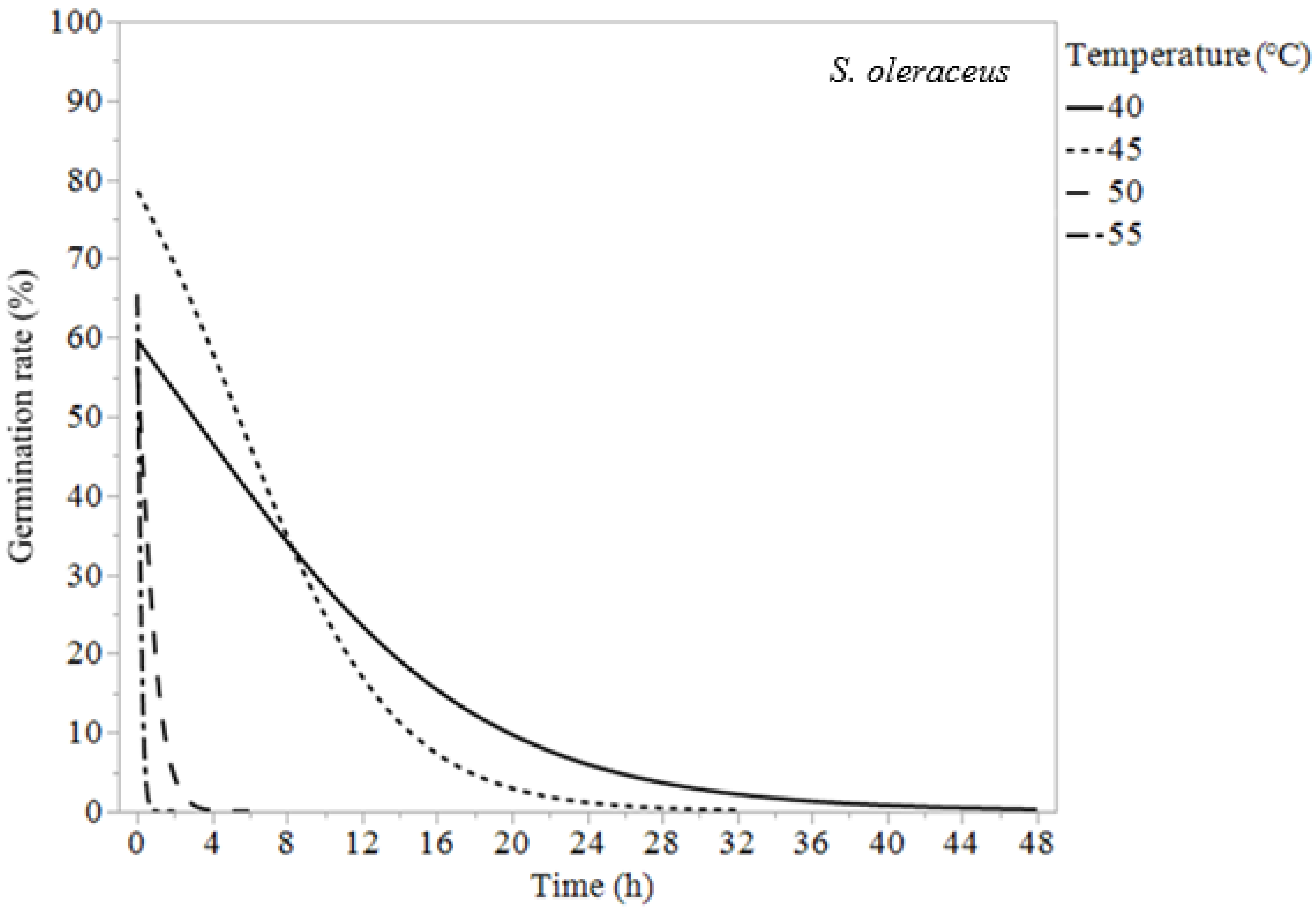
| Sonchus oleraceus | 40 | 3.68 ± 1.40 | 9.52 ± 1.42 | 24.0 ± 2.78 |
| 45 | 3.22 ± 0.87 | 7.22 ± 0.89 | 15.7 ± 1.69 | |
| 50 | 0.27 ± 0.11 | 0.71 ± 0.13 | 1.83 ± 0.28 | |
| 55 | 0.07 ± 0.05 | 0.17 ± 0.05 | 0.41 ± 0.09 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacobs, T.M.; Tubeileh, A.M.; Steinmaus, S.J. Thermal-Time Hazard Models of Seven Weed Species Germinability following Heat Treatment. Agronomy 2024, 14, 275. https://doi.org/10.3390/agronomy14020275
Jacobs TM, Tubeileh AM, Steinmaus SJ. Thermal-Time Hazard Models of Seven Weed Species Germinability following Heat Treatment. Agronomy. 2024; 14(2):275. https://doi.org/10.3390/agronomy14020275
Chicago/Turabian StyleJacobs, Timothy M., Ashraf M. Tubeileh, and Scott J. Steinmaus. 2024. "Thermal-Time Hazard Models of Seven Weed Species Germinability following Heat Treatment" Agronomy 14, no. 2: 275. https://doi.org/10.3390/agronomy14020275
APA StyleJacobs, T. M., Tubeileh, A. M., & Steinmaus, S. J. (2024). Thermal-Time Hazard Models of Seven Weed Species Germinability following Heat Treatment. Agronomy, 14(2), 275. https://doi.org/10.3390/agronomy14020275