
Transfer of the All-Stage Resistance Stripe Rust (Puccinia striifonnis f. sp. Tritici) Resistance Gene YrZH84 in Two Southwestern Chinese Wheat Cultivars

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Construction of Cross-Population and Offspring Screening
2.3. Genotyping Wheat Lines by Markers
2.4. Stripe Rust Reaction and Agronomic Traits in the Field
2.5. Statistical Analysis
3. Results
3.1. Molecular Marker Detection
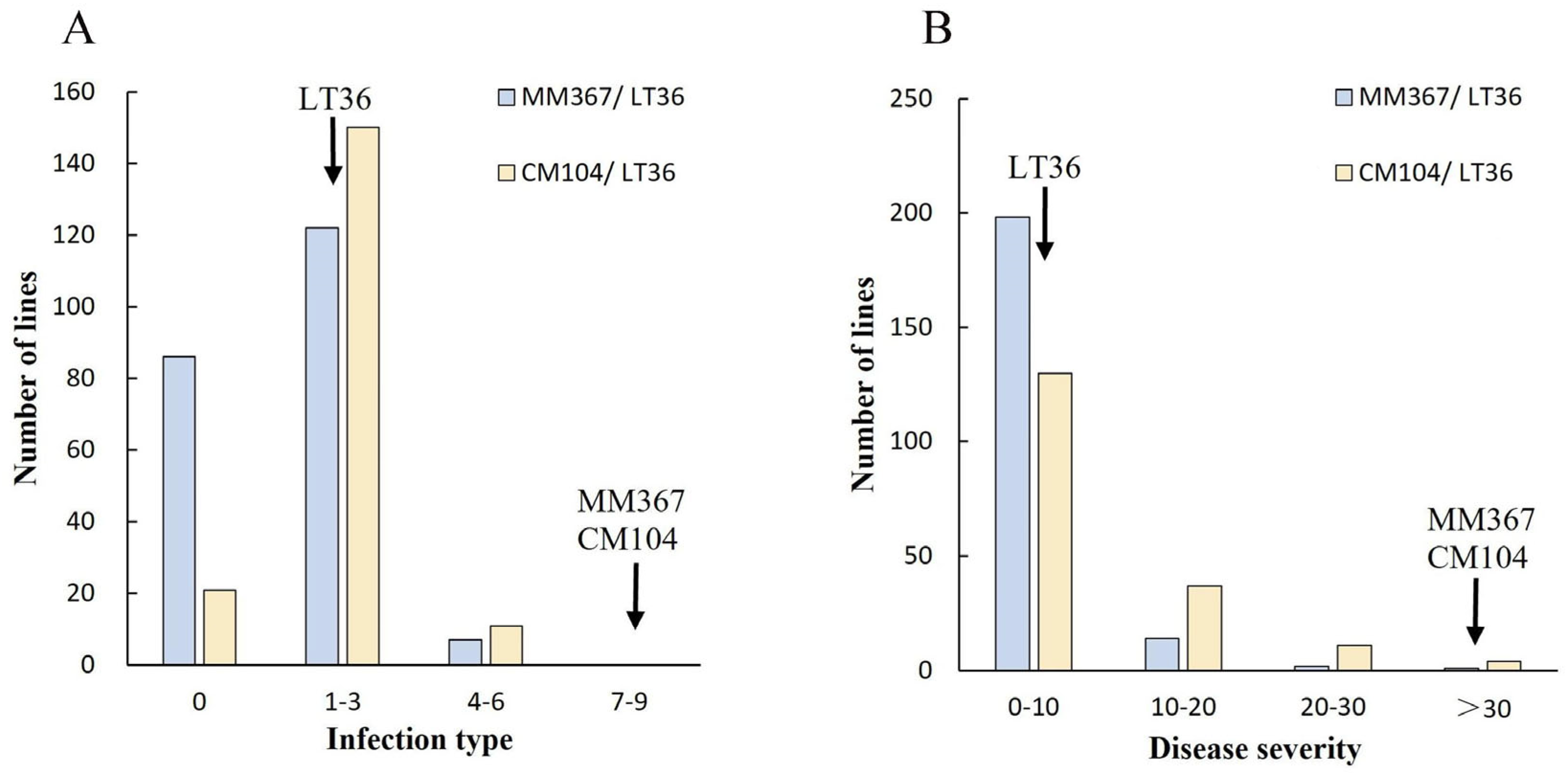
3.2. Stripe Rust Resistance Evaluation
3.3. Agronomic Traits Performance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Bouvet, L.; Holdgate, S.; James, L.; Thomas, J.; Mackay, I.J.; Cockram, J. The evolving battle between yellow rust and wheat: Implications for global food security. Theor. Appl. Genet. 2022, 135, 741–753. [Google Scholar] [CrossRef]
- Wan, A.M.; Zhao, Z.H.; Chen, X.M.; He, Z.H.; Jin, S.L.; Jia, Q.Z.; Yao, G.; X, Y.J.; Wang, B.T.; Li, G.B.; et al. Wheat stripe rust epidemic and virulence of Puccinia striiformis f. sp. tritici in China in 2002. Plant Dis. 2004, 88, 896–904. [Google Scholar] [CrossRef]
- Ali, S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K.; Hansen, J.G.; Lassen, P.; Nazari, K.; Hodson, D.P.; Justesen, A.F.; Hovmøller, M.S. Yellow rust epidemics worldwide were caused by pathogen paces from divergent genetic lineages. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef]
- Schwessinger, B. Fundamental wheat stripe rust research in the 21st century. New Phytol 2017, 213, 1625–1631. [Google Scholar] [CrossRef]
- Gebremariam, T.G.; Wang, F.T.; Lin, R.M.; Li, H.J. Comparative analysis of virulence and molecular diversity of Puccinia striiformis f. sp. tritici isolates collected in 2016 and 2023 in the western region of China. Genes 2024, 15, 542. [Google Scholar] [CrossRef]
- Erenstein, O.; Chamberlin, J.; Sonder, K. Estimating the global number and distribution of maize and wheat farms. Glob. Food Secur. 2021, 30, 100558. [Google Scholar] [CrossRef]
- Singh, J.; Chhabra, B.; Raza, A.; Yang, S.H.; Sandhu, K.S. Important wheat diseases in the US and their management in the 21st century. Front. Plant Sci. 2023, 13, 1010191. [Google Scholar] [CrossRef]
- Mcintosh, R.A.; Dubcovsky, J.; Rogers, W.J.; Xia, X.C.; Raupp, W.J. Catalogue of gene symbols for wheat: 2021 supplement. Annu. Wheat Newsl. 2021, 67, 104–113. Available online: https://wheat.pw.usda.gov/GG3/wgc (accessed on 10 June 2024).
- Klymiuk, V.; Chawla, H.S.; Wiebe, K.; Ens, J.; Fatiukha, A.; Govta, L.; Fahima, T.; Pozniak, C.J. Discovery of stripe rust resistance with incomplete dominance in wild emmer wheat using bulked segregant analysis sequencing. Commun. Biol. 2022, 5, 826. [Google Scholar] [CrossRef]
- Wang, Y.; Xie, J.Z.; Zhang, H.Z.; Guo, B.; Ning, S.Z.; Chen, Y.X.; Lu, P.; Wu, Q.H.; Li, M.M.; Zhang, D.Y.; et al. Mapping stripe rust resistance gene YrZH22 in Chinese wheat cultivar Zhoumai22 by bulked segregant RNA-Seq (BSR-Seq) and comparative genomics analyses. Theor. Appl. Genet. 2017, 130, 2191–2201. [Google Scholar] [CrossRef]
- Grover, G.; Sharma, A.; Mackay, I.; Srivastava, P.; Kaur, S.; Kaur, J.; Burridge, A.; Allen, S.P.; Bentley, A.R.; Chhuneja, P.; et al. Identification of a novel stripe rust resistance gene from the European winter wheat cultivar ‘Acienda’: A step towards rust proofing wheat cultivation. PLoS ONE 2022, 17, e0264027. [Google Scholar] [CrossRef]
- Chen, C.; He, Z.H.; Lu, J.L.; Li, J.; Ren, Y.; Ma, C.X.; Xia, X.C. Molecular mapping of stripe rust resistance gene YrJ22 in Chinese wheat cultivar Jimai22. Mol. Breed. 2016, 36, 118. [Google Scholar] [CrossRef]
- Li, Z.Q.; Zeng, S.M. Wheat Rust in China; China Agriculture Press: Beijing, China, 2002; pp. 164–167. [Google Scholar]
- Tian, J.C. Genetic Analyses of Wheat Major Traits and Molecular Marker-Assisted Breeding; Science Press: Beijing, China, 2015; pp. 474–476. [Google Scholar]
- Hasan, N.; Choudhary, S.; Naaz, N.; Sharma, N.; Laskar, R.A. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. J. Genet. Eng. Biotechnol. 2021, 19, 128. [Google Scholar] [CrossRef]
- Nana, V.; Peter Skov, K.; Jeppe Reitan, A.; Ahmed, J.; Jihad, O. Marker-assisted breeding in wheat. In Next Generation Plant Breeding; Yelda Özden, Ç., Ed.; IntechOpen: Rijeka, Croatia, 2018; pp. 4–10. [Google Scholar]
- Varshney, R.K.; Langridge, P.; Graner, A. Application of genomics to molecular breeding of wheat and barley. Adv. Genet. 2007, 58, 121–155. [Google Scholar] [CrossRef]
- Utomo, H.S.; Linscombe, S.D. Current patents and future development underlying marker-assisted breeding in major grain crops. Recent Pat. DNA Gene Seq. 2009, 3, 53–62. [Google Scholar] [CrossRef]
- Qutub, M.; Chandran, S.; Rathinavel, K.; Sampathrajan, V.; Rajasekaran, R.; Manickam, S.; Adhimoolam, K.; Muniyandi, S.J.; Natesan, S. Improvement of a Yairipok Chujak Maize Landrace from north eastern Himalayan region for β-Carotene content through molecular marker-assisted backcross breeding. Genes 2021, 12, 762. [Google Scholar] [CrossRef]
- Zhou, J.N.; Zheng, X.C.; Zhong, X.; Tan, W.J.; Ma, C.H.; Wang, Y.Q.; Tian, R.; Yang, S.Z.; Li, X.; Xia, C.J.; et al. Transfer of the high-temperature adult-plant stripe rust resistance gene Yr62 in four Chinese wheat cultivars. Mol. Breed. 2023, 43, 44. [Google Scholar] [CrossRef]
- Zhang, T.Z.; Yuan, Y.L.; Yu, J.; Guo, W.Z.; Kohel, R.J. Molecular tagging of a major QTL for fiber strength in upland cotton and its marker-assisted selection. Theor. Appl. Genet. 2003, 106, 262–268. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, F.Y.; Luo, X.X.; Kong, D.Y.; Zhang, A.N.; Wang, F.M.; Pan, Z.Q.; Wang, J.H.; Bi, J.G.; Luo, L.J.; et al. Molecular breeding of a novel PTGMS line of WDR for broad-spectrum resistance to Blast using Pi9, Pi5, and Pi54 genes. Rice 2021, 14, 96. [Google Scholar] [CrossRef]
- De Mori, G.; Cipriani, G. Marker-assisted selection in breeding for fruit trait improvement: A review. Int. J. Mol. Sci. 2023, 24, 8984. [Google Scholar] [CrossRef]
- Li, Z.F.; Zheng, T.C.; He, Z.H.; Li, G.Q.; Xu, S.C.; Li, X.P.; Yang, G.Y.; Singh, R.P.; Xia, X.C. Molecular tagging of stripe rust resistance gene YrZH84 in Chinese wheat line Zhou8425B. Theor. Appl. Genet. 2006, 112, 1098–1103. [Google Scholar] [CrossRef]
- Yin, G.H.; Wang, J.W.; Wen, W.E.; He, Z.H.; Li, Z.F.; Wang, H.; Xia, X.C. Mapping of wheat stripe rust resistance gene YrZH84 with RGAP markers and its application. Acta Agron. Sin. 2009, 35, 1274–1281. [Google Scholar] [CrossRef]
- Zhang, H.; Ren, Y.; He, Y.J.; Zheng, S.H.; Wu, G.; Zhou, F.L.; Lei, J.R.; Du, X.Y.; Tao, J.; Ou, J.M. Molecular detection of disease resistance genes in 153 Sichuan wheat varieties and lines. J. Triticeae Crops 2022, 42, 26–35. [Google Scholar] [CrossRef]
- Zhang, H.P.; Ye, X.L.; Guan, F.N.; Huang, L.; Li, W.; Deng, M.; Wei, Y.M.; Jiang, Y.F.; Chen, G.Y. Identification and evaluation of stripe rust resistance in 220 Sichuan wheat germplasms. J. Sichuan Agric. Univ. 2023, 41, 1020–1031. [Google Scholar] [CrossRef]
- McIntosh, R.; Mu, J.M.; Han, D.J.; Kang, Z.S. Wheat stripe rust resistance gene Yr24/Yr26: A retrospective review. Crop J. 2018, 6, 321–329. [Google Scholar] [CrossRef]
- Cheng, B.; Gao, X.; Cao, N.; Ding, Y.; Gao, Y.; Chen, T.; Xin, Z.; Zhang, L. Genome-wide association analysis of stripe rust resistance loci in wheat accessions from southwestern China. J. Appl. Genet. 2020, 61, 37–50. [Google Scholar] [CrossRef]
- Liu, W.; Frick, M.; Huel, R.; Nykiforuk, C.L.; Wang, X.; Gaudet, D.A.; Eudes, F.; Conner, R.L.; Kuzyk, A.; Chen, Q.; et al. The stripe rust resistance gene Yr10 encodes an evolutionary-conserved and unique CC-NBS-LRR sequence in wheat. Mol. Plant. 2014, 7, 1740–1755. [Google Scholar] [CrossRef]
- Wang, C.M.; Zhang, Y.P.; Han, D.J.; Kang, Z.S.; Li, G.P.; Cao, A.Z.; Chen, P.D. SSR and STS markers for wheat stripe rust resistance gene Yr26. Euphytica 2008, 159, 359–366. [Google Scholar] [CrossRef]
- Mace, E.S.; Buhariwalla, K.K.; Buhariwalla, H.K.; Crouch, J.H. A high-throughput DNA extraction protocol for tropical molecular breeding programs. Plant Mol. Biol. Report. 2003, 21, 459–460. [Google Scholar] [CrossRef]
- Qayoum, A.; Line, R.F. High-temperature, adult-plant resistance to stripe rust of wheat. Phytopathology 1985, 75, 1121–1125. [Google Scholar] [CrossRef]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 2011, 26, 496–500. [Google Scholar] [CrossRef]
- Gai, J.Y. Methods and Standards for Recording Major Agronomic Traits of Wheat; China Agriculture Press: Beijing, China, 2009. [Google Scholar]
- Wang, Z.M. Regularity of wheat stripe rust and integrated control strategy. Neijiang Ke Ji 2003, 2, 19–20. [Google Scholar]
- Wu, X.L.; Tang, Y.L.; Li, C.S.; McHugh, A.D.; Li, Z.; Wu, C.X. Individual and combined effects of soil waterlogging and compaction on physiological characteristics of wheat in southwestern China. Field Crops Res. 2018, 215, 163–172. [Google Scholar] [CrossRef]
- Li, S.Z.; Wang, Q.; Zheng, J.M.; Zhu, H.Z.; Li, J.; Wan, H.S.; Luo, J.T.; Liu, Z.H.; Wu, L. Analysis of genetic components in the major wheat cultivar Chuanmai104 in southwest wheat region based on three wheat SNP arrays. J. Triticeae Crops 2021, 41, 665–672. [Google Scholar] [CrossRef]
- Li, Y.J.; Xu, H.; Yu, S.N.; Tang, J.W.; Li, Q.Y.; Gao, Y.; Zheng, J.Z.; Dong, C.H.; Yuan, Y.H.; Zheng, T.C.; et al. Genetic analysis of elite stripe rust resistance genes of founder parent Zhou8425B in its derived varieties. Acta Agron. Sin. 2024, 50, 16–31. [Google Scholar] [CrossRef]
- Bai, B.; Zhang, H.Z.; Du, J.Y.; Zhang, X.Y.; He, R.; Wu, L.; Zhang, Z.; Zhang, Y.H.; Cao, S.Q.; Liu, Z.Y. Current situation and strategy of stripe rust resistance genes untilization in winter wheat cultivars of northwestern oversummering region for Puccinia striiformis f. sp. tritici in China. Sci. Agric. Sin. 2024, 57, 4–17. [Google Scholar] [CrossRef]
- LI, Z.; Huang, Y.C.; Zhang, C.L.; Yu, R.; Liu, S.J.; Wu, J.H.; Li, C.L.; Zheng, W.J.; Zeng, Q.D.; Kang, Z.S.; et al. Evaluation of stripe rust resistance and analysis of disease resistance genes of the new wheat lines in the middle and lower reaches of the Yangtze River. J. Triticeae Crops 2024, 44, 835–845. [Google Scholar] [CrossRef]
- Atkinson, N.J.; Urwin, P.E. The interaction of plant biotic and abiotic stresses: From genes to the field. J. Exp. Bot. 2012, 63, 3523–3543. [Google Scholar] [CrossRef]
- Rakszegi, M.; Molnár, I.; Darkó, É.; Tiwari, V.K.; Shewry, P. Editorial: Aegilops: Promising genesources to improve agronomical and quality traits of wheat. Front. Plant Sci. 2020, 11, 1060. [Google Scholar] [CrossRef]
- Yan, H.H.; Yu, K.Q.; Xu, Y.h.; Zhou, P.P.; Zhao, J.; Li, Y.; Liu, X.M.; Ren, C.Z.; Peng, Y.Y. Position validation of the dwarfing gene Dw6 in oat (Avena sativa L.) and its correlated effects on agronomic traits. Front. Plant Sci. 2021, 12, 668847. [Google Scholar] [CrossRef]
- Burdon, J.J.; Barrett, L.G.; G, R.; Thrall, P.H. Guiding deployment of resistance in cereals using evolutionary principles. Evol. Appl. 2014, 7, 609–624. [Google Scholar] [CrossRef]
- Alma, K.; Aralbek, R.; Angelina, M.; Makpal, A.; Madina, K.; Zhenis, K. Identification of stripe rust resistance genes in common wheat cultivars and breeding lines from Kazakhstan. Plants 2021, 10, 2303. [Google Scholar] [CrossRef]
- Zheng, S.G.; Li, Y.F.; Lu, L.; Liu, Z.H.; Zhang, C.H.; Ao, D.H.; Li, L.R.; Zhang, C.Y.; Liu, R.; Luo, C.P.; et al. Evaluating the contribution of Yr genes to stripe rust resistance breeding through marker-assisted detection in wheat. Euphytica 2017, 213, 50. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, J.; Abbas, H.M.K.; Liu, Y.; Cheng, M.; Huang, J.; Cheng, W.; Wang, B.; Bai, C.; Wang, G.; et al. Pyramiding of nine transgenes in maize generates high-level resistance against necrotrophic maize pathogens. Theor. Appl. Genet. 2018, 131, 2145–2156. [Google Scholar] [CrossRef]
- Mbinda, W.; Masaki, H. Breeding strategies and challenges in the improvement of blast disease resistance in finger millet. A current review. Front. Plant Sci. 2021, 11, 602882. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Marker | Type | Primer Sequence | Tm (°C) | References |
|---|---|---|---|---|---|
| YrZH84 | Xcfa2040 | SSR | F: TCAAATGATTTCAGGTAACCACTA R: TTCCTGATCCCACCAAACAT | 52.2 | [25] |
| Xbarc32 | SSR | F: GCGTGAATCCGGAAACCCAATCTGTG R: TGGAGAACCTTCGCATTGTGTCATTA | 60.5 | ||
| Yr10 | Yr10 | SCAR | F: TCAAAGACATCAAGAGCCGC R: TGGCCTACATGAACTCTGGAT | 60.0 | [31] |
| Yr26 | Xbarc181 | SSR | F: CGCTGGAGGGGGTAAGTCATCAC R: CGCAAATCAAGAACACGGGAGAAAGAA | 60.5 | [32] |
| We173 | STS | F: GGGACAAGGGGAGTTGAAGC R: GAGAGTTCCAAGCAGAACAC | 56.0 |
| Crosses | YrZH84 (+) | YrZH84 (−) | Total |
|---|---|---|---|
| MM367/LT36 | 99 | 116 | 215 |
| CM104/LT36 | 136 | 46 | 182 |
| Crosses | Number of the Lines | ||||
|---|---|---|---|---|---|
| Immune (IT = 0) | Resistant (IT = 1–3) | Intermediate (IT = 4–6) | Susceptible (IT = 7–9) | Total | |
| MM367/LT36 | 86 | 122 | 7 | 0 | 215 |
| CM104/LT36 | 21 | 150 | 11 | 0 | 182 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, M.; Yang, X.; Feng, X.; Luo, X.; Chen, Q.; Yu, B.; Chen, C.; Huang, K.; Yang, S. Transfer of the All-Stage Resistance Stripe Rust (Puccinia striifonnis f. sp. Tritici) Resistance Gene YrZH84 in Two Southwestern Chinese Wheat Cultivars. Agronomy 2024, 14, 2672. https://doi.org/10.3390/agronomy14112672
Huang M, Yang X, Feng X, Luo X, Chen Q, Yu B, Chen C, Huang K, Yang S. Transfer of the All-Stage Resistance Stripe Rust (Puccinia striifonnis f. sp. Tritici) Resistance Gene YrZH84 in Two Southwestern Chinese Wheat Cultivars. Agronomy. 2024; 14(11):2672. https://doi.org/10.3390/agronomy14112672
Chicago/Turabian StyleHuang, Min, Xue Yang, Xianli Feng, Xiaoqin Luo, Qilin Chen, Boxun Yu, Caihong Chen, Kebing Huang, and Suizhuang Yang. 2024. "Transfer of the All-Stage Resistance Stripe Rust (Puccinia striifonnis f. sp. Tritici) Resistance Gene YrZH84 in Two Southwestern Chinese Wheat Cultivars" Agronomy 14, no. 11: 2672. https://doi.org/10.3390/agronomy14112672
APA StyleHuang, M., Yang, X., Feng, X., Luo, X., Chen, Q., Yu, B., Chen, C., Huang, K., & Yang, S. (2024). Transfer of the All-Stage Resistance Stripe Rust (Puccinia striifonnis f. sp. Tritici) Resistance Gene YrZH84 in Two Southwestern Chinese Wheat Cultivars. Agronomy, 14(11), 2672. https://doi.org/10.3390/agronomy14112672