

Integrating Deficit Irrigation Strategies and Soil-Management Systems in Almond Orchards for Resilient Agriculture

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Field Experimental Design
2.3. Sampling, Chemical Determinations, and Microbiological and Enzymatic Activity Measurements
2.4. Statistical Analysis
3. Results and Discussion
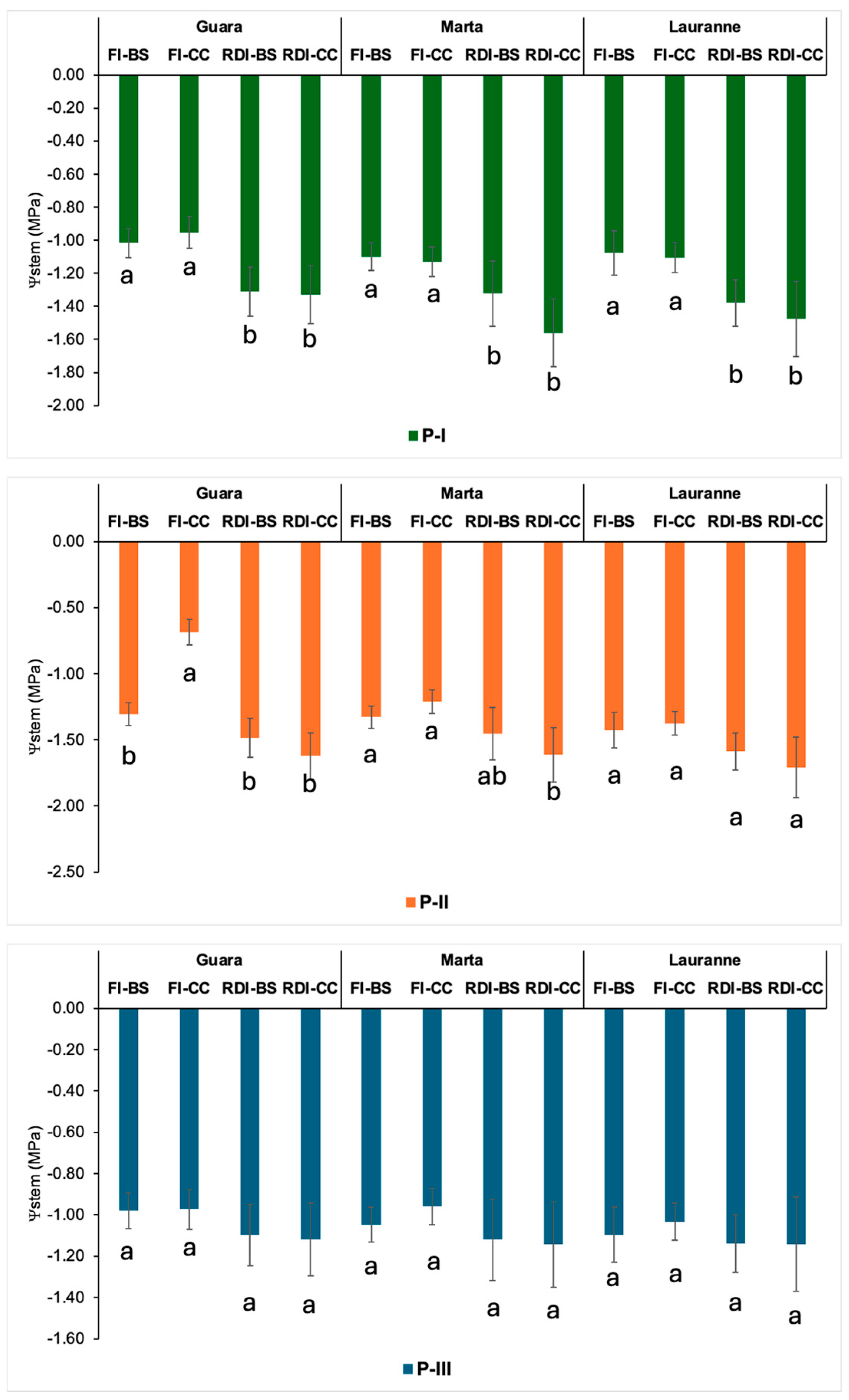
3.1. Crop Water Requirements, Irrigation Doses, Crop Water Status, and Leaf Nutrient Status
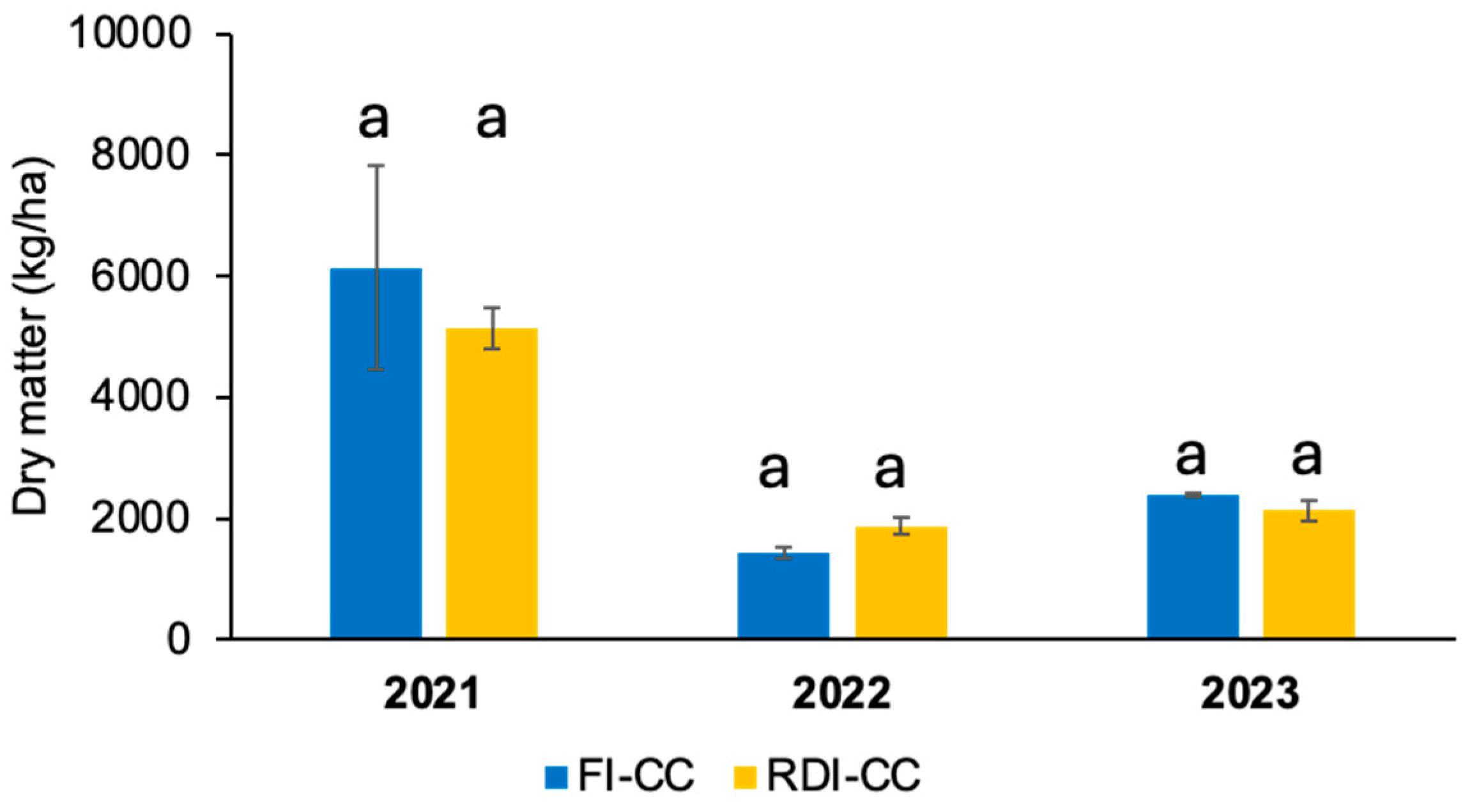
3.2. Cover Crop Dry Mass Production and Atmospheric Carbon Fixation
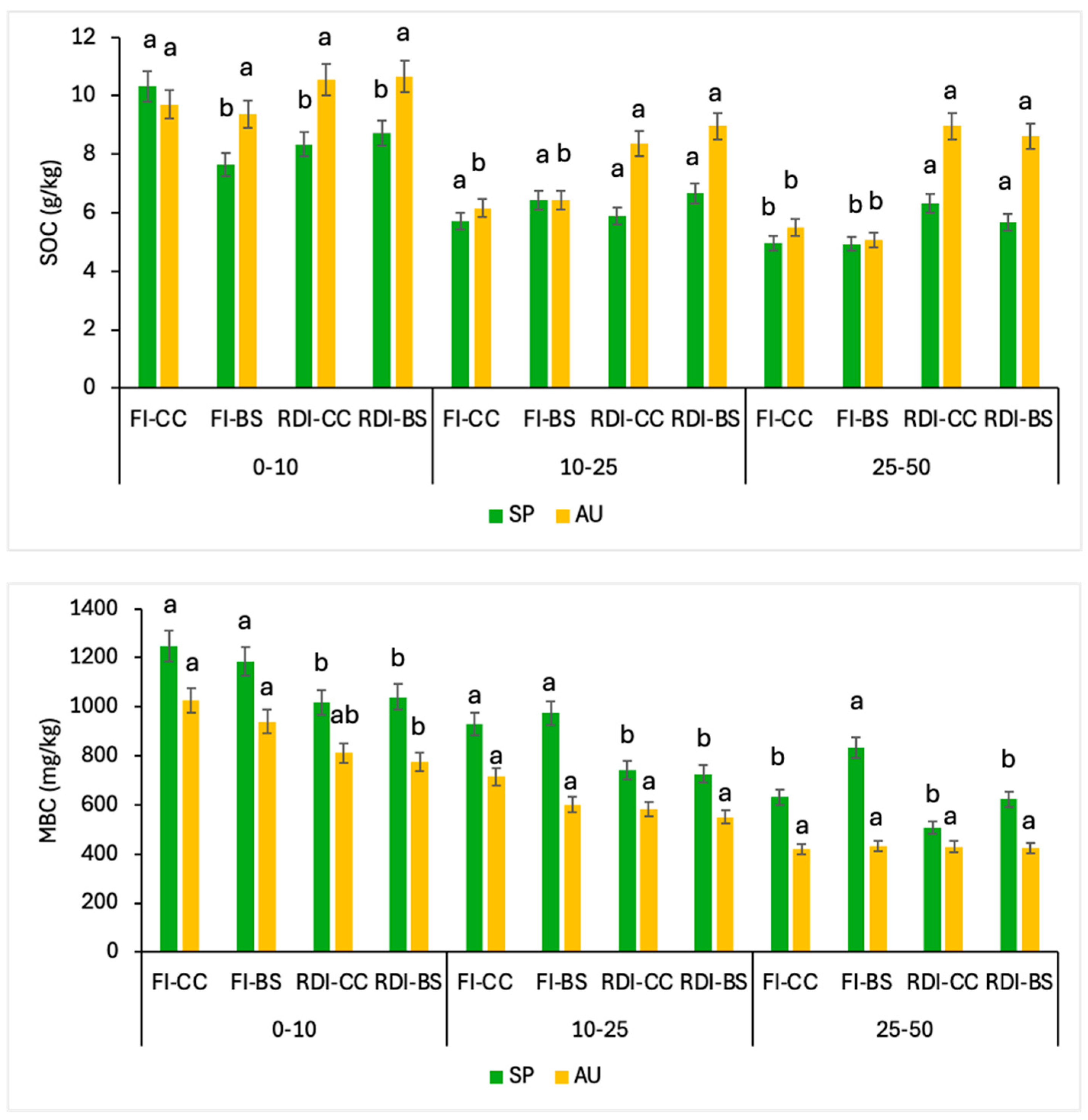
3.3. Soil Organic Carbon, Microbiological Biomass, and Enzymatic Activity
3.4. Almond Yield and Its Components
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Zagaria, C.; Schulp, C.J.E.; Malek, Ž.; Verburg, P.H. Potential for land and water management adaptations in Mediterranean croplands under climate change. Agric. Syst. 2023, 205, 103586. [Google Scholar] [CrossRef]
- García-Tejero, I.F.; Durán, Z.V.H. Water Scarcity and Sustainable Agriculture. In Semiarid Environment. Tools, Strategies, and Challenges for Woody Crops; Academic Press: London, UK, 2018; p. 624. [Google Scholar]
- del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef]
- Cárceles, R.B.; Durán, Z.V.H.; Soriano, R.M.; García-Tejero, I.F.; Gálvez, R.B.; Cuadros, T.S. Conservation agriculture as a sustainable system for soil health: A review. Soil Syst. 2022, 6, 87. [Google Scholar] [CrossRef]
- Hossard, L.; Blanc, L.; Lambarraa-Lehnhardt, F.; Dordas, C.; Papakaloudis, P.; Michalitsis, A.; Lampurlanes, J.; Latati, M.; Touama, R.; Kherif, O.; et al. Co-design of diversified cropping systems in the Mediterranean area. Eur. J. Agron. 2024, 153, 127050. [Google Scholar] [CrossRef]
- Reckling, M.; Watson, C.A.; Whitbread, A.; Helming, K. Diversification for sustainable and resilient agricultural landscape systems. Agron. Sustain. Dev. 2023, 4, 44. [Google Scholar] [CrossRef]
- Hellin, J.; Fisher, E.; Taylor, M.; Bhasme, S.; Loboguerrero, A.M. Transformative adaptation: From climate-smart to climate-resilient agriculture. CABI Agric. Biosci. 2023, 4, 30. [Google Scholar] [CrossRef]
- IEEP. CAP 2021-27: Proposals for Increasing Its Environmental and Climate Ambition. 2018. Available online: https://ieep.eu/wp-content/uploads/2022/12/NABU-CAP-Report-FINAL-.pdf (accessed on 8 July 2024).
- García-Tejero, I.F.; Gordillo, S.G.; Souza, L.; Cuadros-Tavira, S.; Zuazo, V.H.D. Fostering sustainable water use in almond (Prunus dulcis Mill.) orchards in a semiarid Mediterranean environment. Arch. Agron. Soil Sci. 2018, 65, 164–181. [Google Scholar] [CrossRef]
- Rodríguez, B.C.; Zuazo, V.H.D.; Rodríguez, M.S.; Ruiz, B.G.; García-Tejero, I.F. Soil Erosion and the Efficiency of the Conservation Measures in Mediterranean Hillslope Farming (SE Spain). Eurasian Soil Sci. 2021, 54, 792–806. [Google Scholar] [CrossRef]
- Ibarrola-Rivas, M.; Granados-Ramírez, R.; Nonhebel, S. Is the available cropland and water enough for food demand? A global perspective of the Land-Water-Food nexus. Adv. Water Resour. 2017, 110, 476–483. [Google Scholar] [CrossRef]
- Tójar-Hurtado, J.-C.; Mena-Rodríguez, E.; Fernández-Jiménez, M. Spanish Agriculture and Water: Educational Implications of Water Culture and Consumption from the Farmers’ Perspective. Water 2017, 9, 964. [Google Scholar] [CrossRef]
- EC. European Commission, Proposal for a Regulation of the European Parliament and the Council Establishing Rules on Support for Strategic Plans to be Drawn up by Member States under the Common Agricultural Policy (CAP Strategic Plans). COM/2018/392 Final. 2018. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=COM%3A2018%3A392%3AFIN (accessed on 8 July 2024).
- EC. European Commission. The European Green Deal Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions COM (2019) 640 Final. 2019. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=COM%3A2019%3A640%3AFIN (accessed on 8 July 2024).
- EC. European Commission 2023, Horizon Europe Work Programme (2023–24)—EU Missions European Commission Decision C (2023) 2178 of 31 March 2023. 2023. Available online: https://ec.europa.eu/info/funding-tenders/opportunities/docs/2021-2027/horizon/wp-call/2023-2024/wp-1-general-introduction_horizon-2023-2024_en.pdf (accessed on 9 July 2024).
- MAPA. Ministerio de Agricultura, Pesca y Alimentación, Plan Estratégico de la PAC de España 2023-2027. Resumen del Plan aprobado por la Comisión Europea. 2023. Available online: https://www.mapa.gob.es/es/pac/pac-2023-2027/divulgacion-del-plan.aspx (accessed on 5 June 2024).
- Durán, Z.V.H.; Rodríguez, P.C.R.; Francia, M.J.R.; Martínez, R.A.; Arroyo, P.L.; Cárceles, R.B.; Navarro, M.M.C. Benefits of plant strips for sustainable mountain agriculture. Agron. Sustain. Dev. 2008, 28, 497–505. [Google Scholar] [CrossRef]
- Repullo-Ruibérriz de Torres, M.A.; Moreno, G.M.; Ordóñez, F.R.; Rodríguez, L.A.; Cárceles, R.B.; García-Tejero, I.F.; Durán, Z.V.H.; Carbonell, B.R.M. Cover Crop Contributions to Improve the Soil Nitrogen and Carbon Sequestration in Almond Orchards (SW Spain). Agronomy 2021, 11, 387. [Google Scholar] [CrossRef]
- Durán, Z.V.H.; Cárceles, R.B.; García-Tejero, I.F.; Gálvez, R.B.; Cuadros, T.S. Benefits of organic olive rainfed systems to control soil erosion and runoff and improve soil health restoration. Agron. Sustain. Dev. 2020, 40, 41. [Google Scholar] [CrossRef]
- Cárceles, R.B.; Durán, Z.V.H.; Herencia, G.J.F.; Lipan, L.; Soriano, M.; Hernández, F.; Sendra, E.; Carbonell, B.Á.A.; Gálvez, R.B.; García-Tejero, I.F. Soil Management Strategies in Organic Almond Orchards: Implications for Soil Rehabilitation and Nut Quality. Agronomy 2023, 13, 749. [Google Scholar] [CrossRef]
- Bossio, D.A.; Cook-Patton, S.C.; Ellis, P.W.; Fargione, J.; Sanderman, J.; Smith, P.; Wood, S.; Zomer, R.J.; von Unger, M.; Emmer, I.M.; et al. The role of soil carbon in natural climate solutions. Nat. Sustain. 2020, 3, 391–398. [Google Scholar] [CrossRef]
- Soares, P.R.; Harrison, M.T.; Kalantari, Z.; Zhao, W.; Ferreira, C.S.S. Drought effects on soil organic carbon under different agricultural systems. Environ. Res. Commun. 2023, 5, 112001. [Google Scholar] [CrossRef]
- Iglesias, A.; Garrote, L. Local and collective actions for adaptation to use less water for agriculture in the mediterranean region. In Water Scarcity and Sustainable Agriculture in Semiarid Environment; Elsevier: Amsterdam, The Netherlands, 2018; pp. 73–84. [Google Scholar]
- Tejero, I.F.G.; Duran-Zuazo, V.H. Optimizing Plant Water Use Efficiency for a Sustainable Environment; MDPI AG: Basel, Switzerland, 2022; p. 366. [Google Scholar] [CrossRef]
- ESYRCE. Encuesta Sobre Superficies y Rendimientos Cultivos (ESYRCE). 2023. Available online: https://www.mapa.gob.es/es/estadistica/temas/estadisticas-agrarias/agricultura/esyrce/ (accessed on 17 May 2024).
- García-Tejero, I.F.; Lipan, L.; Gutiérrez-Gordillo, S.; Zuazo, V.H.D.; Jančo, I.; Hernández, F.; Rodríguez, B.C.; Carbonell-Barrachina, A. Deficit Irrigation and Its Implications for HydroSOStainable Almond Production. Agronomy 2020, 10, 1632. [Google Scholar] [CrossRef]
- Gutiérrez-Gordillo, S. New Approaches to Hydrosustainable Almonds Production: Agronomical, Physiological and Quality Effects. Ph.D. Thesis, Universidad de Sevilla, Sevilla, Spain, 2022. [Google Scholar]
- Goldhamer, D.A.; Viveros, M.; Salinas, M. Regulated deficit irrigation in almonds: Effects of variations in applied water and stress timing on yield and yield components. Irrig. Sci. 2006, 24, 101–114. [Google Scholar] [CrossRef]
- Moldero, D.; López-Bernal, Á.; Testi, L.; Lorite, I.J.; Fereres, E.; Orgaz, F. Almond responses to a single season of severe irrigation water restrictions. Irrig. Sci. 2021, 40, 1–11. [Google Scholar] [CrossRef]
- López-López, M.; Espadafor, M.; Testi, L.; Lorite, I.J.; Orgaz, F.; Fereres, E. Water use of irrigated almond trees when subjected to water deficits. Agric. Water Manag. 2018, 195, 84–93. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Gonzalez-Dugo, V.; García-Tejero, I.F.; López-Urrea, R.; Intrigliolo, D.S.; Egea, G. Quantitative analysis of almond yield response to irrigation regimes in Mediterranean Spain. Agric. Water Manag. 2023, 279, 108208. [Google Scholar] [CrossRef]
- Egea, G.; Nortes, P.A.; Domingo, R.; Baille, A.; Pérez-Pastor, A.; González-Real, M.M. Almond agronomic response to long-term deficit irrigation applied since orchard establishment. Irrig. Sci. 2013, 31, 445–454. [Google Scholar] [CrossRef]
- de Oliveira, A.F.; Mameli, M.G.; De Pau, L.; Satta, D. Almond Tree Adaptation to Water Stress: Differences in Physiological Performance and Yield Responses among Four Cultivar Grown in Mediterranean Environment. Plants 2023, 12, 1131. [Google Scholar] [CrossRef]
- Egea, G.; González-Real, M.M.; Baille, A.; Nortes, P.A.; Sánchez-Bel, P.; Domingo, R. The effects of contrasted deficit irrigation strategies on the fruit growth and kernel quality of mature almond trees. Agric. Water Manag. 2009, 96, 1605–1614. [Google Scholar] [CrossRef]
- Alcon, F.; Egea, G.; Nortes, P.A. Financial feasibility of implementing regulated and sustained deficit irrigation in almond orchards. Irrig. Sci. 2013, 31, 931–941. [Google Scholar] [CrossRef]
- Mañas, F.; López-Fuster, P.; López-Urrea, R. Effects of Different Regulated and Sustained Deficit Irrigation Strategies in Almond Production. Acta Hortic. 2014, 1028, 391–394. [Google Scholar] [CrossRef]
- Rubio-Asensio, J.S.; Abbatantuono, F.; Ramírez-Cuesta, J.M.; Hortelano, D.; Ruíz, J.L.; Parra, M.; Martínez-Meroño, R.M.; Intrigliolo, D.S.; Buesa, I. Effects of Cover Crops and Drip Fertigation Regime in a Young Almond Agroecosystem. Agronomy 2022, 12, 2606. [Google Scholar] [CrossRef]
- Ramos, M.E.; Benítez, E.; García, P.A.; Robles, A.B. Cover crops under different managements vs. frequent tillage in almond orchards in semiarid conditions: Effects on soil quality. Appl. Soil Ecol. 2010, 44, 6–14. [Google Scholar] [CrossRef]
- FAO. World Reference Base for Soil Resources; World Soil Resources; Report 84; FAO: Rome, Italy, 1998. [Google Scholar]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration. FAO Irrigation and Drainage Paper; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 1998; Volume 56, p. 156. [Google Scholar]
- García-Tejero, I.F.; Hernandez, A.; Rodriguez, V.M.; Ponce, J.R.; Ramos, V.; Muriel, J.L.; Durán, Z.V.H. Estimating almond crop coefficients and physiological response to water stress in semiarid environments (SW Spain). J. Agric. Sci. Technol. 2015, 17, 1255–1266. [Google Scholar]
- MAPA. Ministerio De Agricultura Pesca y Alimentación. Métodos Oficiales De Análisis; Ministerio de Agricultura Pesca y Alimentación: Madrid, Spain, 1994. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining organic carbon in soils: Effect of variations in digestion conditions and of inorganic soil constituents. Soil Sci. 1934, 63, 251–263. [Google Scholar] [CrossRef]
- Adam, G.; Duncan, H. Development of a sensitive and rapid method for the measurement of total microbial activity using fluorescein diacetate (FDA) in a range of soils. Soil Biol. Biochem. 2001, 33, 943–951. [Google Scholar] [CrossRef]
- Brookes, P.; Landman, A.; Pruden, G.; Jenkinson, D. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol. Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol. Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Thalmann, A. Methodology for determining the dehydrogenase activity in soil using triphenyltetrazolium chloride (TTC). Landwirtsch. Forsch. 1968, 21, 249–258. (In German) [Google Scholar]
- Ladd, J.; Butler, J. Short-term assays of soil proteolytic enzyme activities using proteins and dipeptide derivatives as substrates. Soil Biol. Biochem. 1972, 4, 19–30. [Google Scholar] [CrossRef]
- Tabatabai, M.A.; Bremner, J.M. Use of p-nitrophenyl phosphate for assay of soil phosphatase activity. Soil Biol. Biochem. 1969, 1, 301–307. [Google Scholar] [CrossRef]
- Eivazi, F.; Tabatabai, M. Glucosidases and galactosidases in soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Sterk, G.; Stein, A. Mapping Wind-Blown Mass Transport by Modeling Variability in Space and Time. Soil Sci. Soc. Am. J. 1997, 61, 232–239. [Google Scholar] [CrossRef]
- Sperling, O.; Gardi, I.; Ben-Gal, A.; Kamai, T. Deficit irrigation limits almond trees’ photosynthetic productivity and compromises yields. Agric. Water Manag. 2023, 289, 108562. [Google Scholar] [CrossRef]
- Álvarez-Maldini, C.; Acevedo, M.; Estay, D.; Aros, F.; Dumroese, R.K.; Sandoval, S.; Pinto, M. Examining physiological, water relations, and hydraulic vulnerability traits to determine anisohydric and isohydric behavior in almond (Prunus dulcis) cultivars: Implications for selecting agronomic cultivars under changing climate. Front. Plant Sci. 2022, 13, 974050. [Google Scholar] [CrossRef]
- Torrús-Castillo, M.; Domouso, P.; Herrera-Rodríguez, J.M.; Calero, J.; García-Ruiz, R. Aboveground Carbon Fixation and Nutrient Retention in Temporary Spontaneous Cover Crops in Olive Groves of Andalusia. Front. Environ. Sci. 2022, 10, 868410. [Google Scholar] [CrossRef]
- Madejón, E.; Murillo, J.; Moreno, F.; López, M.; Arrue, J.; Alvaro-Fuentes, J.; Cantero, C. Effect of long-term conservation tillage on soil biochemical properties in Mediterranean Spanish areas. Soil Tillage Res. 2009, 105, 55–62. [Google Scholar] [CrossRef]
- Rawls, W.; Pachepsky, Y.; Ritchie, J.; Sobecki, T.; Bloodworth, H. Effect of soil organic carbon on soil water retention. Geoderma 2003, 116, 61–76. [Google Scholar] [CrossRef]
- Núñez, A.; Schipanski, M. Changes in soil organic matter after conversion from irrigated to dryland cropping systems. Agric. Ecosyst. Environ. 2023, 347, 108392. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Melero, S.; Madejón, E.; Ruiz, J.C.; Herencia, J.F. Chemical and biochemical properties of a clay soil under dryland agriculture system as affected by organic fertilization. Eur. J. Agron. 2007, 26, 327–334. [Google Scholar] [CrossRef]
- Lepcha, N.T.; Devi, N.B. Effect of land use, season, and soil depth on soil microbial biomass carbon of Eastern Himalayas. Ecol. Process. 2020, 9, 1–14. [Google Scholar] [CrossRef]
- Wang, Q.R.; Li, Y.C.; Klassen, W. Changes of Soil Microbial Biomass Carbon and Nitrogen with Cover Crops and Irrigation in a Tomato Field. J. Plant Nutr. 2007, 30, 623–639. [Google Scholar] [CrossRef]
- Naylor, D.; Coleman-Derr, D. Drought Stress and Root-Associated Bacterial Communities. Front. Plant Sci. 2018, 8, 2223. [Google Scholar] [CrossRef]
- Schimel, J.P. Life in Dry Soils: Effects of Drought on Soil Microbial Communities and Processes. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 409–432. [Google Scholar] [CrossRef]
- Sun, W.; Zhao, X.; Ling, Q.; Li, H.; Gao, X. Revegetation modifies patterns of temporal soil respiration responses to extreme-drying-and-rewetting in a semiarid ecosystem. Plant Soil 2018, 433, 227–241. [Google Scholar] [CrossRef]
- Zhao, M.; Guo, S.; Wang, R. Diverse soil respiration responses to extreme precipitation patterns in arid and semiarid ecosystems. Appl. Soil Ecol. 2021, 163, 103928. [Google Scholar] [CrossRef]
- Bhandari, K.B.; Acosta-Martínez, V.; Pérez-Guzmán, L.; West, C.P. Soil health within transitions from irrigation to limited irrigation and dryland management. Agric. Environ. Lett. 2022, 7, e20077. [Google Scholar] [CrossRef]
- Chavarría, D.N.; Verdenelli, R.A.; Muñoz, E.J.; Conforto, C.; Restovich, S.B.; Andriulo, A.E.; Meriles, J.M.; Vargas-Gil, S. Soil microbial functionality in response to the inclusion of cover crop mixtures in agricultural systems. Span. J. Agric. Res. 2016, 14, e0304. [Google Scholar] [CrossRef]
- Mikanová, O.; Javůrek, M.; Šimon, T.; Friedlová, M.; Vach, M. The effect of tillage systems on some microbial characteristics. Soil Tillage Res. 2009, 105, 72–76. [Google Scholar] [CrossRef]
- Brzezińska, M.; Stępniewska, Z.; Stępniewski, W. Soil oxygen status and dehydrogenase activity. Soil Biol. Biochem. 1998, 30, 1783–1790. [Google Scholar] [CrossRef]
- Brzezinska, M.; Stepniewska, Z.; Stepniewski, W. Dehydrogenase and catalase activity of soil irrigated with municipal wastewater. Pol. J. Environ. Stud. 2001, 10, 307–311. [Google Scholar]
- Nivelle, E.; Verzeaux, J.; Habbib, H.; Kuzyakov, Y.; Decocq, G.; Roger, D.; Lacoux, J.; Duclercq, J.; Spicher, F.; Nava-Saucedo, J.-E.; et al. Functional response of soil microbial communities to tillage, cover crops and nitrogen fertilization. Appl. Soil Ecol. 2016, 108, 147–155. [Google Scholar] [CrossRef]
- Kaur, J.; Kaur, G. Dehydrogenase activity as a biological indicator of soil health. Chem. Sci. Rev. Lett. 2021, 10, 326–329. [Google Scholar] [CrossRef]
- Roldán, A.; Caravaca, F.; Hernández, M.; Garcıa, C.; Sánchez-Brito, C.; Velásquez, M.; Tiscareño, M. No-tillage, crop residue additions, and legume cover cropping effects on soil quality characteristics under maize in Patzcuaro watershed (Mexico). Soil Tillage Res. 2003, 72, 65–73. [Google Scholar] [CrossRef]
- Bausenwein, U.; Gattinger, A.; Langer, U.; Embacher, A.; Hartmann, H.-P.; Sommer, M.; Munch, J.; Schloter, M. Exploring soil microbial communities and soil organic matter: Variability and interactions in arable soils under minimum tillage practice. Appl. Soil Ecol. 2008, 40, 67–77. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L.; Wu, Z.; Sun, C. Kinetic parameters of soil β-glucosidase response to environmental temperature and moisture regimes. Rev. Bras. Cienc. Solo 2011, 35, 1285–1291. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Drought decreases soil enzyme activity in a Mediterranean Quercus ilex L. Forest. Soil Biol. Biochem. 2005, 37, 455–461. [Google Scholar] [CrossRef]
- Acosta-Martínez, V.; Klose, S.; Zobeck, T.M. Enzyme activities in semiarid soils under conservation reserve program, native rangeland, and cropland. J. Plant Nutr. Soil Sci. 2003, 166, 699–707. [Google Scholar] [CrossRef]
- Wang, X.-C.; Lu, Q. Beta-Glucosidase Activity in Paddy Soils of the Taihu Lake Region, China. Pedosphere 2006, 16, 118–124. [Google Scholar] [CrossRef]
- Sinsabaugh, R.L.; Lauber, C.L.; Weintraub, M.N.; Ahmed, B.; Allison, S.D.; Crenshaw, C.; Contosta, A.R.; Cusack, D.; Frey, S.; Gallo, M.E.; et al. Stoichiometry of soil enzyme activity at global scale. Ecol. Lett. 2008, 11, 1252–1264. [Google Scholar] [CrossRef]
- Makoi, J.H.J.R.; Chimphango, S.B.M.; Dakora, F.D. Elevated levels of acid and alkaline phosphatase activity in roots and rhizosphere of cowpea (Vigna unguiculata L. Walp.) genotypes grown in mixed culture and at different densities with sorghum (Sorghum bicolor L.). Crop. Pasture Sci. 2010, 61, 279–286. [Google Scholar] [CrossRef]
- Kandeler, E.; Palli, S.; Stemmer, M.; Gerzabek, M.H. Tillage changes microbial biomass and enzyme activities in particle-size fractions of a Haplic Chernozem. Soil Biol. Biochem. 1999, 31, 1253–1264. [Google Scholar] [CrossRef]
- Green, V.; Stott, D.; Cruz, J.; Curi, N. Tillage impacts on soil biological activity and aggregation in a Brazilian Cerrado Oxisol. Soil Tillage Res. 2007, 92, 114–121. [Google Scholar] [CrossRef]
- Geisseler, D.; Horwath, W.R.; Scow, K.M. Soil moisture and plant residue addition interact in their effect on extracellular enzyme activity. Pedobiologia 2011, 54, 71–78. [Google Scholar] [CrossRef]
- Stewart, W.L.; Fulton, A.E.; Krueger, W.H.; Lampinen, B.D.; Shackel, K.A. Regulated deficit irrigation reduces water use of almonds without affecting yield. Calif. Agric. 2011, 65, 90–95. [Google Scholar] [CrossRef]
- Barreales, D.; Capitão, S.; Bento, A.A.; Casquero, P.A.; Ribeiro, A.C. Adapting Almond Production to Climate Change through Deficit Irrigation and Foliar Kaolin Application in a Mediterranean Climate. Atmosphere 2023, 14, 1593. [Google Scholar] [CrossRef]
- Moldero, D.; López-Bernal, Á.; Testi, L.; Lorite, I.J.; Fereres, E.; Orgaz, F. Long-term almond yield response to deficit irrigation. Irrig. Sci. 2021, 39, 409–420. [Google Scholar] [CrossRef]
- Girona, J.; Mata, M.; Marsal, J. Regulated deficit irrigation during the kernel-filling period and optimal irrigation rates in almond. Agric. Water Manag. 2005, 75, 152–167. [Google Scholar] [CrossRef]
- García, J.; Romero, P.; Botía, P.; García, F. Cost-benefit analysis of almond orchard under regulated deficit irrigation (RDI) in SE Spain. Span. J. Agric. Res. 2004, 2, 157–165. [Google Scholar] [CrossRef]
- Koumanov, K.S.; Hopmans, J.W.; Schwankl, L.W. Spatial and temporal distribution of root water uptake of an almond tree under microsprinkler irrigation. Irrig. Sci. 2006, 24, 267–278. [Google Scholar] [CrossRef]
- Durán-Zuazo, V.H.; Pleguezuelo, C.R.; Panadero, L.A.; Raya, A.M.; Martínez, J.F.; Rodríguez, B.C. Soil Conservation Measures in Rainfed Olive Orchards in South-Eastern Spain: Impacts of Plant Strips on Soil Water Dynamics. Pedosphere 2009, 19, 453–464. [Google Scholar] [CrossRef]
- Palese, A.; Vignozzi, N.; Celano, G.; Agnelli, A.; Pagliai, M.; Xiloyannis, C. Influence of soil management on soil physical characteristics and water storage in a mature rainfed olive orchard. Soil Tillage Res. 2014, 144, 96–109. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Resource | Strategy and Management Systems | |
|---|---|---|
| Water | Full irrigation (100% II.RR) (FI) | |
| Regulated deficit irrigation (RDI) | ||
| Soil | Bare soil without cover crops (BS) | |
| Cover crop (legume + cereal) (CC) | ||
| Water and soil treatments | FI-BS | RDI-BS |
| FI-CC | RDI-CC | |
| Samples | Timing | Measurements | |
|---|---|---|---|
| Soil | Spring (Mid-April) | Autumn (October) | SOC, MBC, FDA, DH, PROT, AP, GLUC |
| Almond leaves | Summer (Mid-June) | Mineral status (N, K, P, Na, Ca, Mg, Cu, Zn, Fe, Mn) | |
| Almond tree | Fortnightly (from April to September) | Ψstem, yield and its components (mid-August) | |
| Cover crop | Before sowing (April) | Dry matter | |
| Strategy | II.RR. | Rainfall | ETC | Phenological Stage | Season | |
|---|---|---|---|---|---|---|
| RDI | FI | |||||
| (mm) | ||||||
| 73 | 116 | 85 | 198 | 224 | P-I | 2021 |
| 156 | 397 | 406 | 3 | 408 | P-II | |
| 79 | 118 | 85 | 79 | 140 | P-III | |
| 308 | 631 | 576 | 280 | 772 | Total | |
| 67 | 96 | 75 | 164 | 190 | P-I | 2022 |
| 142 | 355 | 412 | 2 | 413 | P-II | |
| 93 | 155 | 164 | 73 | 215 | P-III | |
| 302 | 606 | 651 | 239 | 818 | Total | |
| 144 | 184 | 203 | 107 | 278 | P-I | 2023 |
| 124 | 313 | 358 | 25 | 375 | P-II | |
| 104 | 128 | 141 | 37 | 167 | P-III | |
| 362 | 625 | 702 | 169 | 820 | Total | |
| Mn (ppm) | Cu (ppm) | Zn (ppm) | Fe (ppm) | Na (ppm) | %Ca | % Mg | %K | %P | %N | Treatment | Cultivar |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 39.1 | 8.6 a | 35.3 a | 358.8 a | 230.8 a | 5.2 a | 0.7 a | 2.2 a | 0.14 a | 2.8 a | FI CC | Guara |
| 37.9 a | 8.2 a | 34.7 a | 335.9 a | 210.2 a | 4.8 a | 0.6 a | 2.7 a | 0.14 a | 2.9 a | FI BS | |
| 33.7 a | 9.8 a | 35.0 a | 397.5 a | 320.4 b | 7.2 b | 0.8 a | 2.7 a | 0.13 a | 2.4 a | DI CC | |
| 32.4 a | 9.9 a | 33.7 a | 246.9 a | 232.8 a | 7.0 b | 0.8 a | 2.6 a | 0.13 a | 2.1 a | DI BS | |
| 39.7 a | 11.2 a | 31.6 a | 326.9 a | 273.4 a | 4.8 a | 0.5 a | 2.5 a | 0.14 a | 2.5 a | FI CC | Marta |
| 38.5 a | 8.7 a | 33.9 a | 298.6 a | 254.8 a | 5.0 a | 0.6 a | 2.8 a | 0.14 a | 2.6 a | FI BS | |
| 35.6 a | 9.4 a | 29.6 a | 324.5 a | 287.8 a | 6.7 b | 0.8 a | 2.5 a | 0.12 a | 2.3 a | DI CC | |
| 39.1 a | 12.1 a | 39.0 a | 386.6 a | 246.0 a | 5.8 b | 0.9 a | 2.7 a | 0.12 a | 2.3 a | DI BS | |
| 34.0 a | 9.4 a | 34.3 a | 384.4 a | 305.3 a | 4.8 a | 0.6 a | 2.8 a | 0.15 a | 2.7 a | FI CC | Lauranne |
| 35.0 a | 11.2 a | 33.9 a | 272.3 a | 260.2 a | 4.9 a | 0.6 a | 2.7 a | 0.14 a | 2.8 a | FI BS | |
| 33.1 a | 8.7 a | 34.1 a | 340.1 a | 332.8 a | 8.2 b | 0.9 b | 2.7 a | 0.13 a | 2.3 a | DI CC | |
| 36.7 a | 11.3 a | 37.3 a | 341.4 a | 290.8 a | 6.8 b | 0.9 b | 2.8 a | 0.13 a | 2.4 a | DI BS |
| Year | cv. Guara | cv. Marta | cv. Lauranne | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FI-BS | FI-CC | RDI-BS | RDI-CC | FI-BS | FI-CC | RDI-BS | RDI-CC | FI-BS | FI-CC | RDI-BS | RDI-CC | |
| Yield (kg ha−1) | ||||||||||||
| 2021 | 792 b | 1071 a | 487 c | 500 c | 764 c | 1001 ab | 932 b | 1175 a | 1180 b | 1418 a | 1050 b | 843 c |
| 2022 | 1631 a | 1776 a | 1433 ab | 10,340 b | 1993 a | 2143 a | 2160 a | 2091 a | 2249 a | 2522 a | 1940 b | 1826 b |
| 2023 | 1431 ab | 1562 a | 1372 b | 1594 a | 1921 a | 1709 b | 2016 a | 1790 ab | 1383 a | 1479 a | 1447 a | 1328 a |
| 2021–23 | 1285 ab | 1470 a | 1097 b | 1145 b | 1559 a | 1618 a | 1703 a | 1685 a | 1604 ab | 1806 a | 1479 b | 1332 c |
| Kernel weight (g) | ||||||||||||
| 2021 | 1.31 a | 1.25 b | 1.26 b | 1.23 b | 1.32 a | 1.35 a | 1.32 a | 1.31 a | 1.18 b | 1.20 a | 1.17 b | 1.17 b |
| 2022 | 1.41 a | 1.42 a | 1.44 a | 1.42 a | 1.51 a | 1.47 a | 1.46 a | 1.38 a | 1.22 a | 1.28 a | 1.25 a | 1.19 b |
| 2023 | 1.18 a | 1.06 b | 1.03 b | 0.96 b | 1.15 b | 1.31 a | 1.10 b | 1.15 b | 1.09 b | 1.21 a | 1.10 b | 1.04 c |
| 2021–23 | 1.30 a | 1.24 b | 1.24 b | 1.20 c | 1.33 b | 1.37 a | 1.29 c | 1.28 c | 1.16 b | 1.23 a | 1.17 b | 1.13 c |
| Ratio | ||||||||||||
| 2021 | 0.35 b | 0.35 b | 0.37 a | 0.37 a | 0.31 a | 0.31 a | 0.3 a | 0.3 a | 0.31 a | 0.31 a | 0.31 a | 0.31 a |
| 2022 | 0.40 a | 0.40 a | 0.39 a | 0.39 a | 0.37 a | 0.37 a | 0.36 a | 0.36 a | 0.36 a | 0.36 a | 0.36 a | 0.36 a |
| 2023 | 0.32 b | 0.33 b | 0.34 ab | 0.35 a | 0.32 b | 0.32 b | 0.34 a | 0.32 b | 0.31 a | 0.31 a | 0.30 a | 0.32 a |
| 2021–23 | 0.36 a | 0.36 a | 0.37 a | 0.37 a | 0.33 a | 0.33 a | 0.24 b | 0.24 b | 0.33 a | 0.33 a | 0.32 a | 0.33 a |
| IWP (kg m−3) | ||||||||||||
| 2021 | 0.13 a | 0.17 a | 0.16 a | 0.16 a | 0.12 b | 0.16 b | 0.30 a | 0.38 a | 0.19 b | 0.22 b | 0.34 a | 0.27 a |
| 2022 | 0.27 b | 0.29 b | 0.48 a | 0.44 a | 0.33 b | 0.35 b | 0.72 a | 0.70 a | 0.37 b | 0.42 b | 0.64 a | 0.60 a |
| 2023 | 0.23 b | 0.25 b | 0.38 a | 0.44 a | 0.31 b | 0.27 b | 0.56 a | 0.50 a | 0.22 b | 0.24 b | 0.40 a | 0.37 a |
| 2021–23 | 0.21 b | 0.24 b | 0.34 a | 0.35 a | 0.25 b | 0.25 b | 0.53 a | 0.53 a | 0.26 b | 0.29 b | 0.46 a | 0.41 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Tejero, I.F.; Herencia-Galán, J.F.; Cárceles Rodríguez, B.; Calderón-Pavón, A.; Aldana Navarro, J.; Rubio-Casal, A.E.; Durán Zuazo, V.H. Integrating Deficit Irrigation Strategies and Soil-Management Systems in Almond Orchards for Resilient Agriculture. Agronomy 2024, 14, 2288. https://doi.org/10.3390/agronomy14102288
García-Tejero IF, Herencia-Galán JF, Cárceles Rodríguez B, Calderón-Pavón A, Aldana Navarro J, Rubio-Casal AE, Durán Zuazo VH. Integrating Deficit Irrigation Strategies and Soil-Management Systems in Almond Orchards for Resilient Agriculture. Agronomy. 2024; 14(10):2288. https://doi.org/10.3390/agronomy14102288
Chicago/Turabian StyleGarcía-Tejero, Iván Francisco, Juan Francisco Herencia-Galán, Belén Cárceles Rodríguez, Abel Calderón-Pavón, Javier Aldana Navarro, Alfredo Emilio Rubio-Casal, and Víctor Hugo Durán Zuazo. 2024. "Integrating Deficit Irrigation Strategies and Soil-Management Systems in Almond Orchards for Resilient Agriculture" Agronomy 14, no. 10: 2288. https://doi.org/10.3390/agronomy14102288
APA StyleGarcía-Tejero, I. F., Herencia-Galán, J. F., Cárceles Rodríguez, B., Calderón-Pavón, A., Aldana Navarro, J., Rubio-Casal, A. E., & Durán Zuazo, V. H. (2024). Integrating Deficit Irrigation Strategies and Soil-Management Systems in Almond Orchards for Resilient Agriculture. Agronomy, 14(10), 2288. https://doi.org/10.3390/agronomy14102288