Abstract
Pleiotropic drug resistance (PDR) transporters, which are part of the ABCG subfamily of ATP-binding cassette (ABC) transporters, have been found to be involved in heavy metal tolerance. Salix species (willow) is widely regarded as a perfect candidate for phytoremediation of heavy metals because of its substantial biomass, strong tolerance, and remarkable capacity to accumulate heavy metals. However, the phylogeny and mechanisms underlying the response to heavy metals within the PDR family in willow have yet to be determined. In this study, we discovered and valuated a total of 21 PDR genes in the genome of Salix purpurea. The phylogenetic relationships of these genes were used to classify them into five major clades. The SpPDRs exhibited variations in exon-intron distribution patterns and gene lengths across different branches. Cis-acting elements linked to stress response, drought induction, low temperature, and defense response were discovered in the promoters of PDRs. Significant variations in the transcription levels of various PDR genes were observed across different tissues under heavy metal stress, with distinct heavy metals regulating different PDR members. In roots, PDR4 and PDR21 exhibited high expression levels. Meanwhile, PDR7 and PDR17 showed similar transcription patterns across all analyzed tissues. Furthermore, there was a significant and positive correlation between PDR5 and PDR16, whereas a significant and negative correlation was detected between PDR3 and PDR9, suggesting that the response of PDR members to heavy metals is complex and multifaceted. These findings will establish a vital basis for comprehending the biological functionalities of PDR genes, specifically their involvement in the regulation of willow’s tolerance to heavy metals.
1. Introduction
Over the past few decades, increased human activities like mining, improper disposal of waste, and the utilization of fertilizers have resulted in extensive contamination of heavy metals [1]. This contamination poses a significant danger to the safety of food and the environment and also has harmful impacts on the health of humans, animals, and plants. Phytoremediation, a method that utilizes plants, has gained popularity in many countries due to its low cost, effectiveness, and non-intrusiveness [2,3,4]. In this process, hyperaccumulators can accumulate larger quantities of heavy metals in their above-ground tissues compared to their below-ground tissues [5]. Woody plants possess several advantages over herbaceous plants, owing to their large biomass and deep-rooted root systems. Some woody plant species, like Phytolacca americana [6,7] for cadmium (Cd) and zinc (Zn) hyperaccumulation, and Schima superba [8] for manganese (Mn) hyperaccumulation, have been discovered. Additionally, willow and poplar have been recognized as preferred plants for phytoremediation because they have high resistance to heavy metals, can accumulate them, and produce significant above-ground biomass [9,10].
The transport systems within plants’ cells are vital for the absorption, movement, and compartmentalization of toxic metals. Currently, researchers have identified 17 transporter families with a total of 313 heavy metal transporters that facilitate the efficient movement of heavy metals [11]. ATP-binding cassette (ABC) transporters regulate a range of biological processes, such as nutrient uptake, metal tolerance, and phytohormone transport [12]. Integral membrane proteins known as ABC transporters are present in various organisms. They consist of two transmembrane domains (TMDs) and two nucleotide-binding domains (NBDs) arranged in tandem [13]. The NBDs, alternatively referred to as ATP-binding cassettes, contain highly conserved Walker A and Walker B sequences, a slightly less conserved H-ring Q ring, and ABC protein-specific sites. The TMDs, which have six membrane-spanning α-helices, utilize the energy released by NBD to regulate substrate recognition and specificity [14,15]. Plants have eight subfamilies (A, B, C, D, E, F, G, and I) of ABC transporters [16]. ABCA to ABCD have the TMD-NBD domain in the forward direction, while ABCG and ABCH subfamilies display reverse NBD-TMD motifs. Soluble proteins are identified as ABCE and ABCF subfamilies, each containing only two NBDs. The ABCI subfamily primarily contains a single domain, predominantly NBD, which is challenging to identify and classify [17]. Based on their molecular size, ABC transporters can be further categorized into two main categories. One category includes full-size proteins with two NBDs and two TMDs, while the other category comprises half-size proteins consisting of one set of these two domains. Among these, the ABCG subfamily is comprised of two types: white–brown complex (WBC), which is a half-size transporter, and pleiotropic drug resistance (PDR), which is a full-size transporter.
In plants, certain PDRs have been found to participate in heavy metal tolerance. For instance, AtABCG40/AtPDR12 in Arabidopsis showed a significant increase in expression under lead (Pb) stress. Overexpression of AtPDR12 in plants led to improved resistance against Pb(II) and decreased Pb levels in comparison to wild-type plants, suggesting that AtPDR12 facilitates the transport of Pb or Pb derivatives out of the cytoplasm, thereby improving Pb tolerance in Arabidopsis [18]. Another ABCG transporter, AtPDR8, functions as an efflux pump which is responsible for the transport of Cd2+ or Cd conjugates in Arabidopsis, thus conferring resistance against heavy metals. This pump functions by actively transporting Cd2+ from the cytosol to the apoplast [19]. Likewise, the rice OsABCG43 gene, which has Cd transport activity, demonstrates transcriptional expression in almost all tissues and organs under Cd stress. In yeast, the overexpression of OsABCG43 enhanced cellular tolerance and decreased Cd accumulation [20]. The overexpression of PtoABCG36 has shown the ability to decrease the accumulation of Cd in transgenic plants, indicating that PtoABCG36 improves Cd tolerance by reducing the Cd content in plants [21]. These studies indicate that the PDR family possesses the capacity to transport heavy metal ions and plays a crucial function in enhancing heavy metal tolerance in plants. However, our understanding of PDRs in response to heavy metal stress is limited to Arabidopsis, rice, and poplar. Hence, it is necessary to identify PDR genes on a genome-wide scale and explore their functions in response to various heavy metals.
The potential use of Salix species in phytoremediation has been realized due to their substantial biomass, strong tolerance to heavy metals, and remarkable ability for heavy metal accumulation. For instance, S. integra has demonstrated strong tolerance to Pb stress and is considered a potential candidate for the phytostalization of Pb-Zn mine tailings [22]. S. viminalis and S. caprea have been found to reduce the soil concentration of Cd, Zn, and copper (Cu) [23]. Additionally, S. matsudana var. matsudana f. umbraculifera Rehd, hereinafter referred to as S. matsudana, has been identified as an arbor willow species with high tolerance to and accumulation of Cd [24,25]. Based on our previous study, we found that several ABCG genes or PDRs exhibited a notable increase in expression levels in S. matsudana when exposed to Cd. This indicates that PDRs have a vital function in responding to Cd stress. Importantly, both S. purpurea and S. matsudana have demonstrated a remarkable ability to tolerate metals, as evidenced by their substantial biomass production and metal accumulation in their leaves [26]. The aim of this study was to identify the PDR family in the genome of S. purpurea (unpublished genome sequence of S. matsudana) and investigate their roles in responding to different heavy metal stresses in S. matsudana. Our study involved scientific bioinformatics analysis, including the examination of conserved motifs, phylogenetic analysis, and chromosomal localization. Additionally, we examined the expression patterns of homologous SmPDRs in different tissues of S. matsudana under heavy metal stress. These findings could improve our comprehension of the biological roles of PDR transporters and offer new perspectives into the molecular mechanisms of heavy metal response in willow.
2. Materials and Methods
2.1. Identification of SpPDR Genes in S. purpurea
To identify SpPDR proteins in the genome, we conducted BLAST searches against the S. purpurea database on the JGI Genome Portal database (https://genome.jgi.doe.gov/portal/, accessed on 10 February 2023). We utilized all PDR sequences of Arabidopsis and Populus trichocarpa (V 4.1) as queries for the searches. The gene sequences of AtPDR were retrieved from the Arabidopsis Information Resource (TAIR) database. The PDR domains were confirmed by the Conserved Domain Database (CDD) of NCBI and PROSITE (https://prosite.expasy.org/scanprosite/, accessed on 20 February 2023). To eliminate redundancy, we removed duplicate sequences and obtained a unique set of potential PDR transporter proteins.
We used the ClustalW program in MEGA X to compare SpPDR sequences and identify the characteristics of conserved domains in SpPDR proteins. The members of the SpPDR family were named based on their corresponding chromosome sequences and mapped to different chromosomes. The fundamental characteristics of the SpPDR proteins were predicted by utilizing the Expasy ProtParam tool (https://web.expasy.org/protparam/, accessed on 25 February 2023) [27], and their subcellular location was predicted using WoLF PSORT II (https://wolfpsort.hgc.jp/, accessed on 25 February 2023).
2.2. Phylogenetic Analysis and Location in the Chromosome of the SpPDR Genes
Using ClustalW in MEGA X, we generated a neighbor-joining (NJ) phylogenetic tree to align the full-length PDR amino acid sequences. The alignment parameters consisted of poisson correction, pairwise deletion, and bootstrap analysis with 1000 replicates. Internal branch support was estimated using a bootstrap consensus tree generated from 1000 replicates [28]. We visualized the resulting phylogenetic tree using ChiPlot (https://www.chiplot.online, accessed on 5 March 2023). To illustrate the positions of SpPDR genes in chromosomal sequences, we utilized the Gene Location Visualize and Gene Density Profile functions in the GTF/GFF module of TBtools software (version 1.109), along with gene density files and GFF annotation files of the genome [29].
2.3. Analysis of Gene Structure and Prediction of Conserved Motifs
Each PDR protein sequence in S. purpurea was submitted to the online program MEME (http://meme-suite.org/tools/meme, accessed on 10 March 2023) in order to identify conserved protein motifs. Additionally, we used the ProtParam tool to calculate the physicochemical properties of PDR proteins (http://web.expasy.org/protparam/, accessed on 10 March 2023) [30].
The exon-intron arrangement of each gene was analyzed by aligning the genomic and coding sequences (CDS) of SpPDR genes. For the analysis of PDR protein domains, we referred to the Conserved Domain Database (CDD) provided by NCBI (https://www.ncbi.nlm.nih.gov/cdd/, accessed on 15 March 2023). The gene structure and domains of PDR genes were visualized separately using the gene structure module and NCBI CDD domain pattern module of the TBtools software (https://github.com/CJ-Chen/TBtools, accessed on 15 March 2023).
2.4. Analysis of SpPDR Gene Promoters
The 2000 bp region located upstream of the translation initiation site was determined to be the promoter sequence of SpPDR genes. To predict the possible cis-acting regulatory elements within these sequences, the PlantCARE web server (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/, accessed on 25 March 2023) was utilized [31].
2.5. Gene Duplication and Syntenic Analysis of SpPDR Genes
To identify tandem and segment replication events of the PDR genes, we obtained the CDS sequences of S. purpurea using Perl programs. These sequences were then compared pairwise using BLASTn procedures. The gene pairs of SpPDRs were analyzed using the Multiple Collinearity Scan Toolkit (MCScanX). Finally, the SpPDR gene pairs within S. purpurea were visualized by the Circos program.
We collected genomic sequences of four dicotyledons (A. thaliana, P. trichocarpa, Nico tiana tabacum, Gossypium herbaceum) and two monocotyledons (Oryza sativa and Zea mays) from various sources. The genomic data for N. tabacum was acquired from the Solanaceae Genomics Network (https://solgenomics.net, accessed on 25 March 2023), while the genomic information for G. herbaceum was obtained from Cotton MD (https://yanglab.hzau.edu.cn/CottonMD, accessed on 25 March 2023). Other species’ genomic information was obtained from Phytozome v.13 (https://phytozome-next.jgi.doe.gov/, accessed on 25 March 2023). The syntenic connections across the entire genome between S. purpurea and six additional plant species were analyzed using the MCScanX program. The obtained results were then visualized using the Circos program.
2.6. Protein–Protein Interaction Network Analysis of SpPDRs
We further identified homologous genes of SpPDRs in A. thaliana to study protein-protein interactions using ‘Proteins by sequences’ module in the String database (https://cn.string-db.org/, accessed on 25 August 2023). Cytoscape (version 3.8, https://cytoscape.org/, accessed on 25 August 2023) was used to visualize the interaction network.
2.7. Plant Materials and Different Heavy Metal Treatments
Shoot cuttings were collected from the same S. matsudana plant at the Research Institute of Subtropical Forestry, Chinese Academy of Forestry. The cuttings, approximately 20 cm in length, were propagated asexually in a half-strength modified Hoagland nutrient solution (Coolar, China) within a plant growth chamber. The growth chamber operated under a photoperiod of 14 h of light and 10 h of darkness, maintaining a temperature range of 22 °C to 25 °C. Following a 50 day pre-culturing period, uniform-height cuttings were exposed to various treatments: 200 μM ZnSO4 (Zn200), 0 μM ZnSO4 (Zn0), 100 μM CuSO4 (Cu100), 10 μM CdSO4 (Cd10), and 100 μM CdSO4 (Cd100). Samples of roots, stems, leaves, and tender shoots (axillary buds sprouted for 10 days) were collected at 0 (control), 1 day, 4 days, and 7 days after the treatments. Each treatment was replicated three times to ensure statistical validity. All collected samples were quickly frozen in liquid nitrogen and preserved at a temperature of −80 °C for future utilization.
2.8. Quantitative Real-Time RT-PCR (qRT-PCR) Analysis
Total RNA was extracted from all samples using the RNAprep Pure Plant Plus Kit (DP441, Tiangen Biotech Co., Ltd., Beijing, China). To evaluate the integrity of total RNAs, 1.0% (p/v) agarose gel electrophoresis was performed, and the concentration was determined using a NanoDrop2000 spectrophotometer (Thermo Scientific, Waltham, MA, USA). Subsequently, cDNA first strand was synthesized using the PrimeScriptTM RT Master Mix (RR036, TaKaRa, Dalian, China). For qRT-PCR analysis, the diluted cDNA was prepared by 10-fold dilution in deionized water. The CDS sequences of SpPDRs were obtained from S. purpurea, and the primer pairs for all genes in the CDS region were selected using the Primer3 program (https://bioinfo.ut.ee/primer3/, accessed on 15 April 2023). To normalize the expression levels, the endogenous gene DnaJ was employed as a reference gene [32]. All primers used in this study are comprehensively listed in Supplementary Table S1. The qRT-PCR reaction mixture (RR420, TaKaRa, Dalian, China) was prepared in a 20 µL volume, consisting of 10 µL of SYBR®Premix Ex TaqTM, 0.4 µL of both forward and reverse primers, 2 µL of the cDNA template, 0.4 µL of ROX Reference Dye, and 6.8 µL ddH2O. The qRT-PCR protocol started with a 30-second incubation at 95 °C, followed by 40 cycles at 95 °C for 5 s, and 60 °C for 31 s. The Ct values were determined using a 7300 Real-Time PCR system (Applied Biosystems, Foster, CA, USA). The 2−△△Ct method was utilized to calculate the relative expression levels [33]. The mRNA levels of the untreated control samples (CK) were arbitrarily assigned a value of 1 in this approach [34]. Heatmaps were generated through the ‘pheatmap’ function from the R package (version R4.3.1). Z-score normalization was used to normalize the expression values of the qRT-PCR data.
3. Results
3.1. Identification of SpPDR Sequences
The BLASTP algorithm was employed to search for S. purpurea protein sequences against NBD-TMD protein sequences from various plants. As a result, a total of 21 putative hits were identified, indicating the presence of both NBD and TMD domains. Utilizing bioinformatic tools such as CDD, SMART, and Pfam, we obtained 21 PDR sequences with two NBDs and two TMD domains. Additionally, we retrieved 132 full-size ABC protein sequences, including 21, 12, 23, 19, 15, 22, and 20 sequences from S. purpurea, A. thaliana, P. trichocarpa, N. tabacum, G. herbaceum, O. sativa, and Z. mays from genomics, respectively, and created a local sequence database.
As presented in Table 1, the range of amino acids varies from 1370 (SpPDR9) to 1501 (SpPDR6). The molecular weight ranges from 154.03 KDa to 169.79 KDa. Their pI (isoelectric point) values range from 5.95 (SpPDR9) to 9.07 (SpPDR15). Additionally, 21 SpPDRs were found to be localized on the plasma membrane (Table 1).

Table 1.
The basic information list of the identified pleiotropic drug resistance (PDR) in S. purpurea.
3.2. Phylogenetic Analysis of PDR Proteins
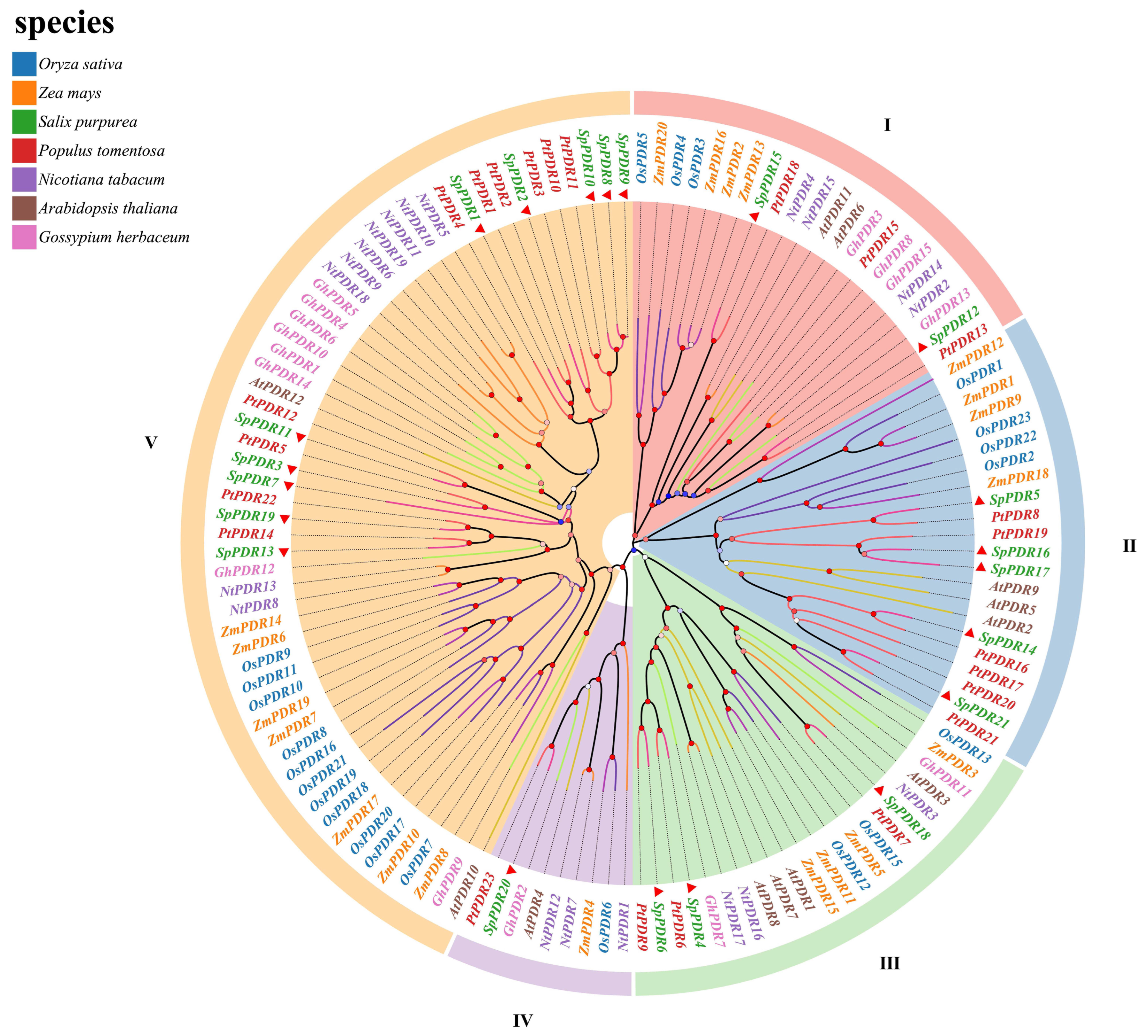
We constructed a phylogenetic tree using the NJ method implemented in MEGA X. The tree was created by aligning full-length PDR protein sequences from various species, including S. purpurea (21 proteins), A. thaliana (12 proteins), Z. mays (20 proteins), N. tabacum (19 proteins), P. tomentosa (23 proteins), O. sativa (20 proteins), and G. herbaceum (19 proteins).
Based on the division of the phylogenetic tree (Figure 1), the SpPDRs were categorized into five clusters, which were consistent with N. tabacum and A. thaliana as well [35,36]. Notably, the class III subfamily exhibited the highest number of members, with 57 genes accounting for 43.2% of the total. Conversely, the class IV subfamily had a smaller number of members, with only nine genes accounting for 6.8%. The number of SpPDR proteins in each subfamily ranged from 9 to 57, indicating distinct evolutionary trajectories. A fascinating discovery was the dispersed arrangement of SpPDRs in every subgroup, indicating gene amplification prior to the divergence of S. purpurea and A. thaliana. Furthermore, the presence of OsPDRs and SpPDRs in each branch indicated the conservation of PDR gene evolution between dicots and monocots. Additionally, the clustering of SpPDRs and PtPDRs in separate branches indicated a strong similarity between S. purpurea and P. trichocarpa (Figure 1).
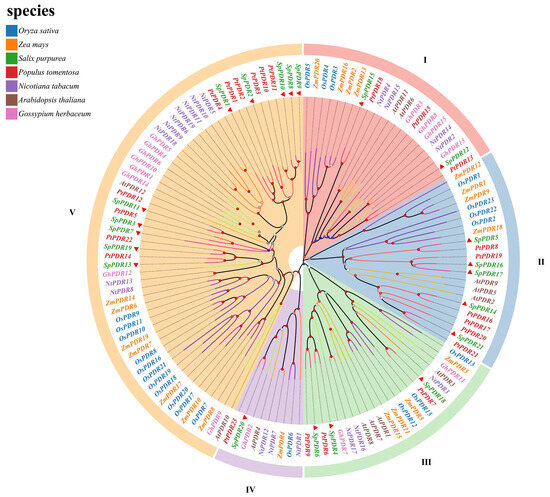
Figure 1.
Phylogenetic tree of the PDR family of seven species. The PDR sequences from five dicotyledons (A. thaliana, S. purpurea, P. trichocarpa, N. tabacum, and G. herbaceum) and two monocotyledons (O. sativa and Z. mays) are incorporated in the phylogenetic tree. The phylogenetic tree was built using the maximum-likelihood approach, and 1000 bootstrap replicates were employed. The tree is divided into five unique clades, with each clade being depicted by different colors. The figure displays bootstrap values ranging from 0% to 100%, with the SpPDRs indicated by a red triangle.
3.3. Gene Structure and Motif Analysis of SpPDRs
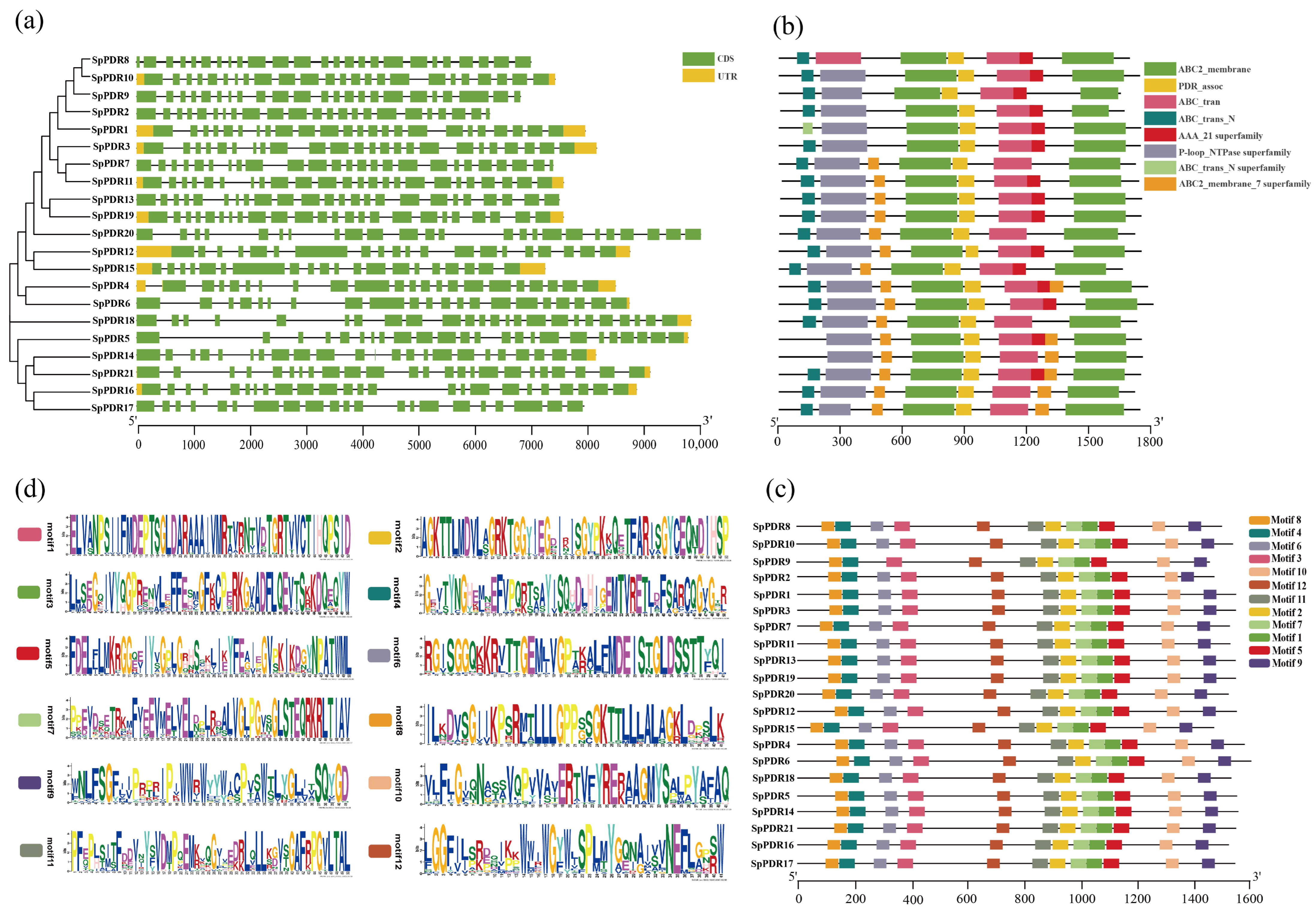
The distribution patterns of exons and introns, as well as the lengths of genes in SpPDRs, are illustrated in Figure 2a. Among the SpPDR genes, SpPDR7, SpPDR8, and SpPDR16 exhibited the highest number of exons. These genes differ in terms of the number and location of introns. Due to the close proximity of the exon-intron associations, the SpPDR family experienced gene duplication throughout its evolutionary history.
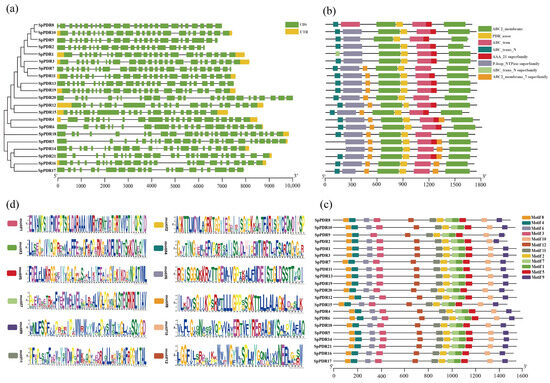
Figure 2.
Phylogenetic relationship, gene structure analysis, domain, and conserved motif analysis. (a) The NJ phylogenetic tree of SpPDR genes and the exon-intron arrangement of SpPDR genes; (b) the domains of SpPDRs; (c) all conserved motifs in the SpPDR proteins. Different motifs are highlighted by a differently colored box (number 1–12); (d) the sequences of twelve motifs in SpPDR proteins.
The domains of the SpPDR proteins were identified using the bioinformatic tool CDD (Figure 2b). All members of the PDR family were found to have the PDR association domain. Furthermore, most SpPDRs had seven or eight domains, indicating the highly conserved nature of PDR proteins throughout evolution and suggesting similar functions among them.
A total of 12 motifs were identified in each PDR protein (except for PDR9) according to the SpPDR structure map. The lengths and best-matching sequences of these 12 motifs were depicted in Figure 2c, and the corresponding motif logos were generated using WebLogo (Figure 2d). These motifs exhibited the highest level of conservation. The order, type, and quantity of motifs in PDR protein sequences were generally consistent within the same subfamily, but they differed between the class I subfamily and other subfamilies. This phenomenon implies that some family members may have undergone the loss of certain motifs during the course of evolution, resulting in the development of new functions.
3.4. Promotor Analysis of SpPDRs
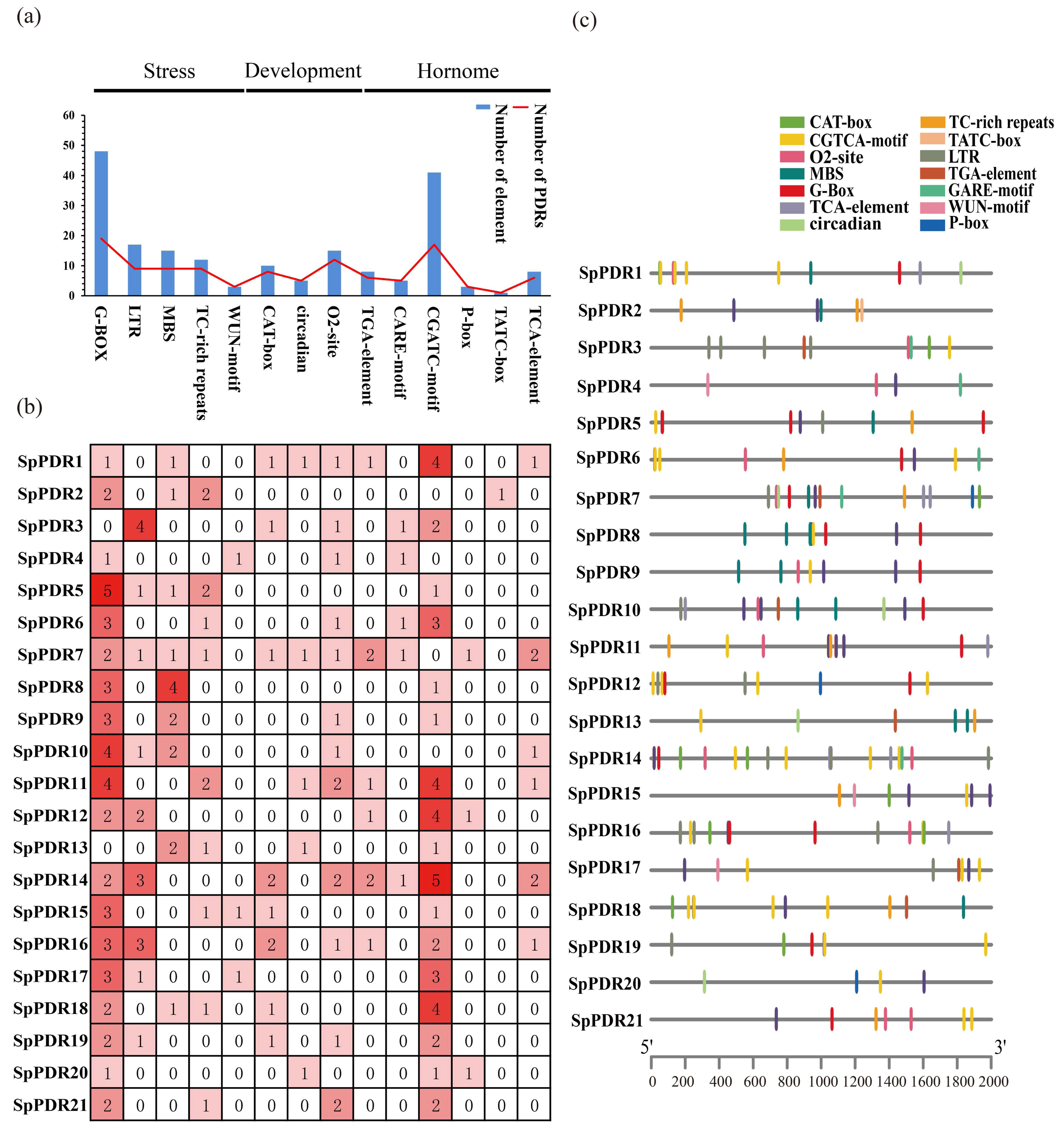
A total of 14 cis-acting elements were displayed in Figure 3a. Here, we discovered cis-acting elements associated with stress (G-box, LTR, MBS, TC-rich repeats, WUN-motif), growth and development (CAT-box, circadian, and O2-site), and hormones (TGA-element, CGTCA-motif, TCA-element, p-box, and TATC-box) [37]. Significantly, the G-box and CGATC-motif displayed the highest occurrences, indicating their potential involvement in stress and hormone responses under certain conditions (Figure 3b). However, the analysis of cis-acting elements revealed that SpPDRs are capable of responding to various abiotic stresses. The potential regulatory roles of G-box and MBS binding sites in PDR expression are also suggested (Figure 3c).
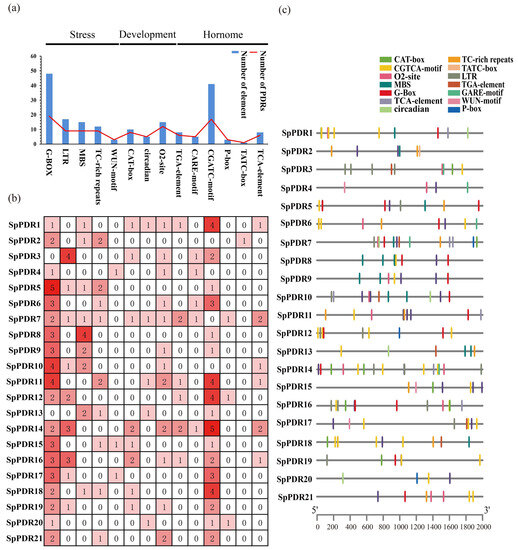
Figure 3.
The cis-acting elements of the SpPDR genes in S. purpurea. (a) The total count of cis-acting elements in response to various factors; (b) the number of each cis-acting element within the promoter region (2 kb upstream of the translation start site). The intensity of the color indicates the abundance of cis-acting elements; and (c) the distribution of cis-acting elements in SpPDR promoters.
3.5. Chromosomal Distribution of SpPDRs
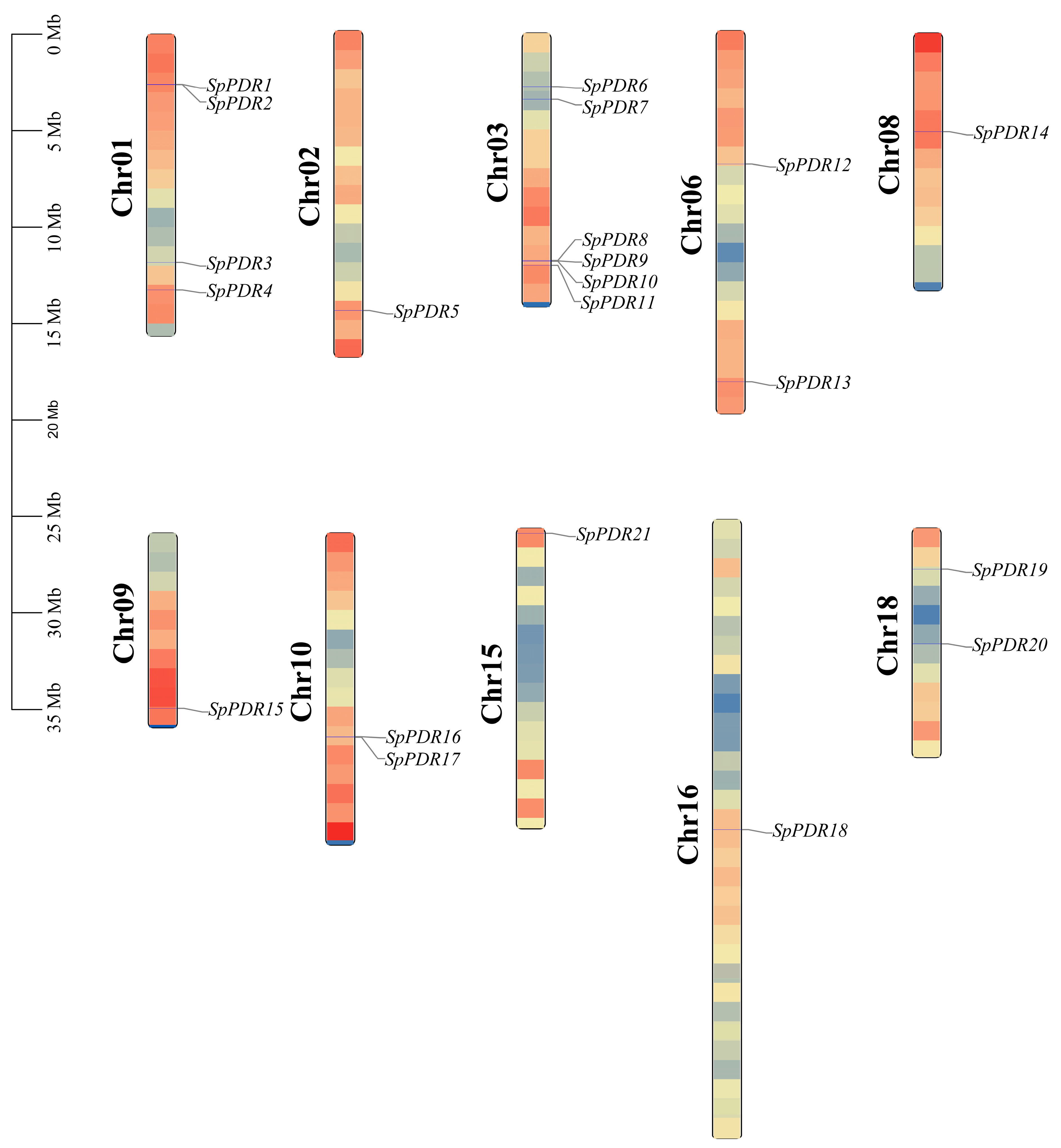
The chromosomal distributions of the 21 SpPDR genes were investigated, and it was found that they are present on 10 out of the 19 chromosomes (Figure 4). The number of SpPDR genes varied on every chromosome, with a range of 1 to 6. Notably, chr-03 harbored the highest number of SpPDR genes, totaling 6 (28.57% of the genes examined). In contrast, chr-02, chr-08, chr-09, chr-15, and chr-16 contained only one gene each, respectively.
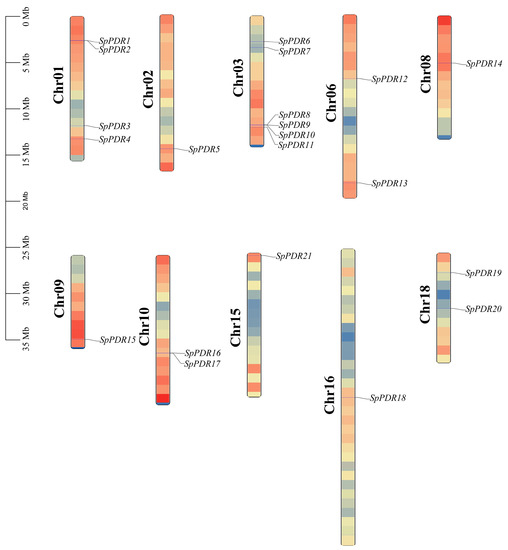
Figure 4.
The distribution of SpPDR genes on the chromosomes of S. purpurea. The genetic distance of 10 chromosomes is depicted on the left scale in megabases (Mb). The black lines represent the location of the gene on each chromosome.
3.6. Gene Duplication and Syntenic Analysis of SpPDRs
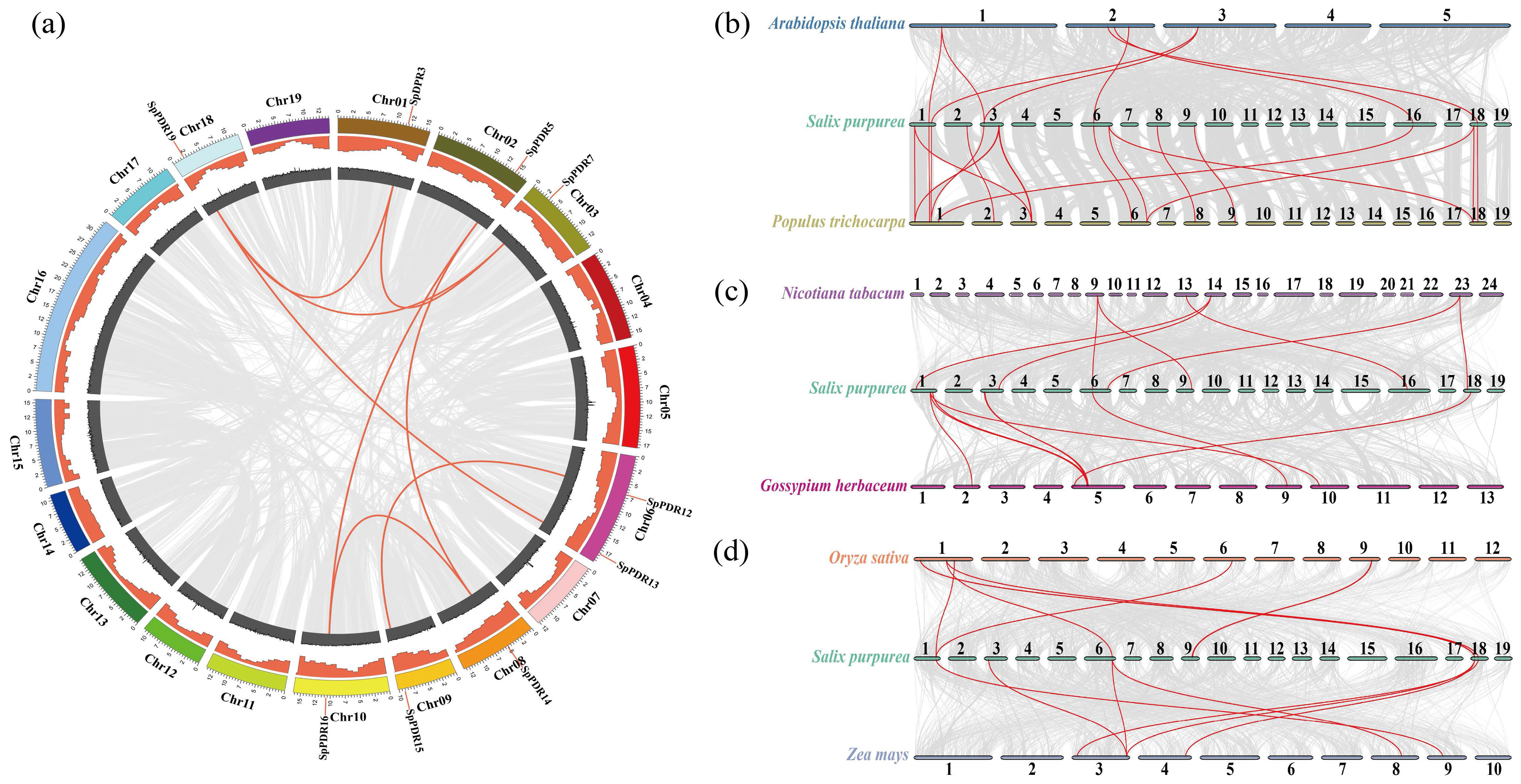
Gene duplication may illustrate the evolutionary process of the PDR family in genomes, as it played a significant role in driving genome evolution [38]. To illustrate genomic evolution, the syntenic analysis of PDR genes can be conducted by identifying conserved gene pairs among them [39]. We performed a genome-level collinearity analysis of SpPDR genes, and three comparative syntenic maps of S. purpurea were created using the MCScanX and Circos programs. The comparison included six representative species, encompassing both herbaceous and woody plants, as well as dicotyledons and monocotyledons (Figure 5a–d). Among the 21 genes examined, nine genes (including eight gene pairs with tandem duplication) exhibited matching gene pairs, and the assessment of selective pressure indicated that all collinearity pairs had a Ka/Ks ratio below one. This result suggests that SpPDR genes are conserved within the PDR family (Table S2). Moreover, seven homologous gene pairs were identified between S. purpurea and other species such as A. thaliana, N. tabacum, and G. herbaceum. Similarly, six homologous gene pairs were discovered between S. purpurea and O. sativa or Z. mays, whereas 17 homologous gene pairs were identified between S. purpurea and P. trichocarpa, suggesting resemblances between S. purpurea and dicotyledons or monocotyledons. However, there were notable differences in gene collinearity between S. purpurea and herbs or woody plants. These findings showed a closer relationship between S. purpurea and P. trichocarpa compared to other species.
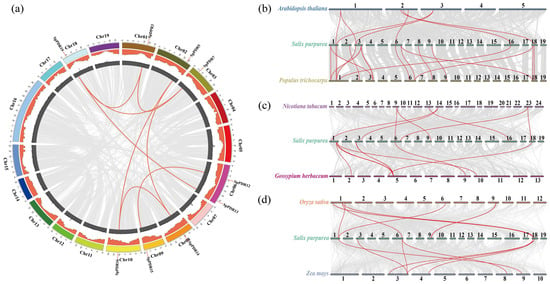
Figure 5.
Gene duplication of SpPDR genes on 19 chromosomes within the S. purpurea genome and collinearity analysis of PDR genes of S. purpurea with other species. (a) Genome location and synteny of SpPDR genes. The density of genes on the chromosomes is represented by the orange circles, while the GC content is indicated by the gray circles. Tandem duplication gene pairs of other genes are depicted with gray solid lines, whereas the tandem duplication gene pairs of SpPDRs are represented by red solid lines; (b) collinearity analysis of PDR genes of A. thaliana, S. purpurea, and P. trichocarpa; (c) collinearity analysis in S. purpurea with N. tabacum and G. herbaceum; and (d) collinearity analysis in S. purpurea with O. sativa and Z. mays. The red lines highlight the collinear gene pairs of PDR genes in various species.
3.7. Protein-Protein Interaction Network Analysis of SpPDRs
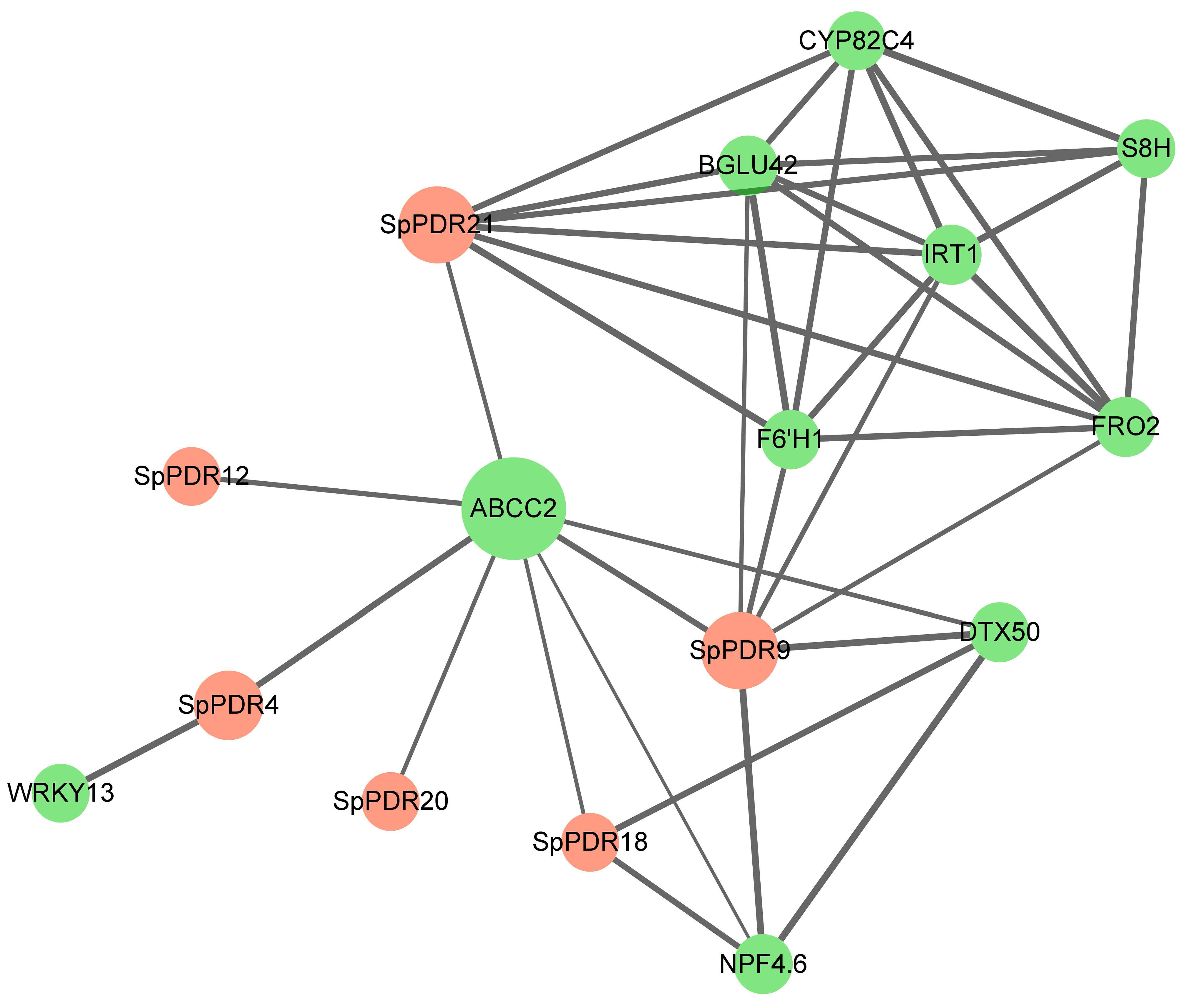
A protein-protein interaction map of SpPDRs is shown in Figure 6. Two SpPDRs (PDR21 and PDR9) exhibit associations with two target transporters, including ABCC2 and iron-regulated transport protein 1 (IRT1), which have been previously implicated in Cd tolerance [40,41]. Other target proteins, such as CYP82C4 (Xanthotoxin 5-hydroxylase) and S8H (Scopoletin 8-hydroxylase), play an important role in the response to iron deficiency. Curiously, no protein-protein interactions were discovered between different SpPDR members, suggesting independent functions among SpPDR proteins.
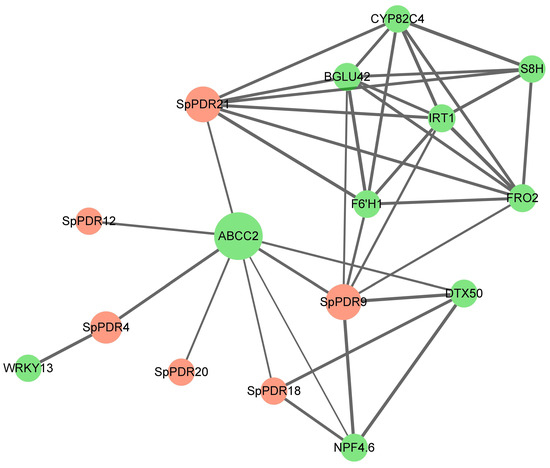
Figure 6.
The diagram of the protein–protein interaction network of SpPDRs. The pink dots represent SpPDRs; the green dots show other genes.
3.8. Expression Patterns of PDR Genes in Different Tissues
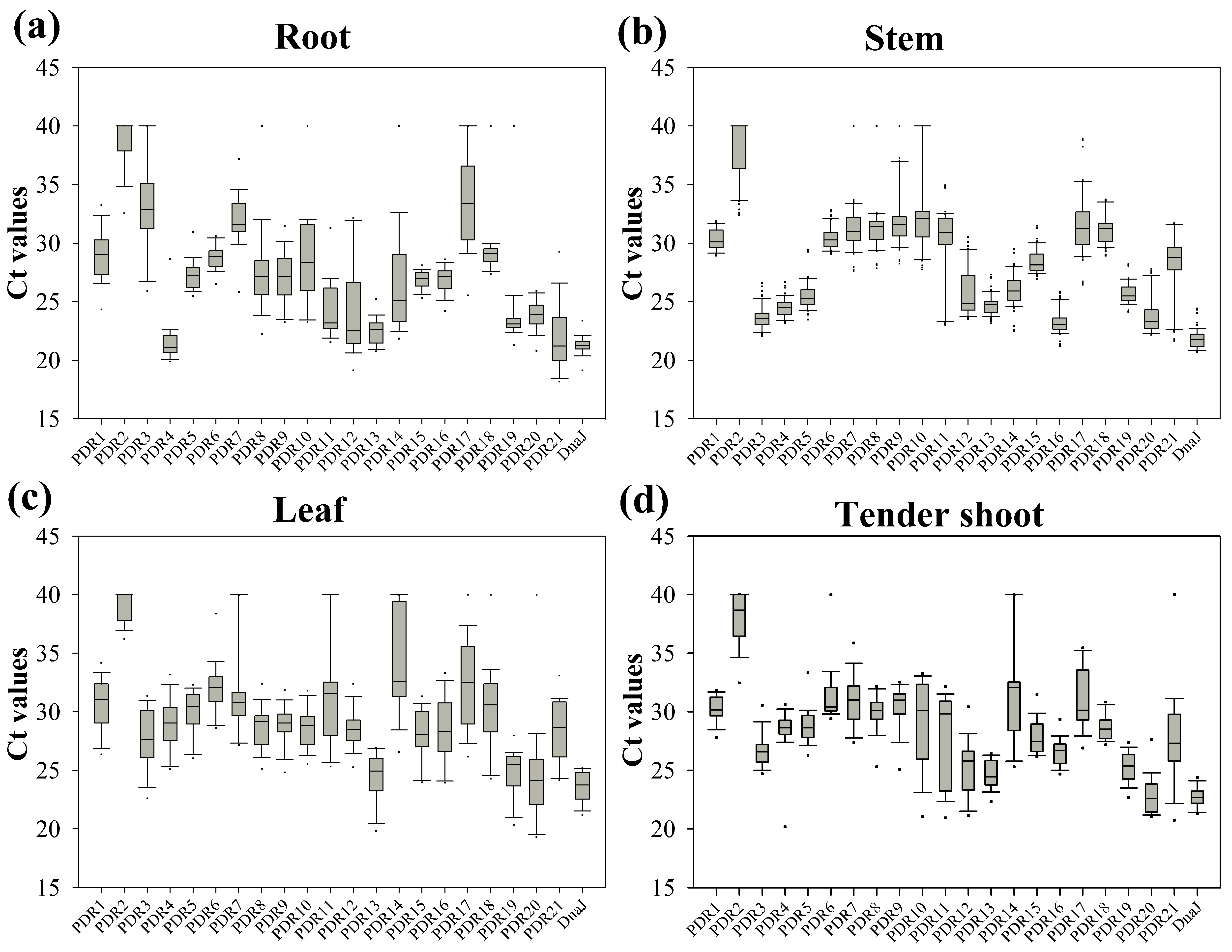
The expression patterns of PDRs in different tissues of S. matsudana were evaluated, as both S. matsudana and S. purpurea displayed strong tolerance to heavy metals and similar heavy metal distributions in their leaves [26]. The Ct range was used to assess the variations in transcript levels among these 21 PDR members in response to heavy metals. As expected, significant variations in average Ct values were observed among different members across various tissues (Figure 7). All SmPDRs exhibited narrower Ct value variations in stems compared to other tissues, suggesting that these genes are less responsive to heavy metal stress. However, more than 10 genes specifically responded to heavy metals in roots (Figure 7). Among all tissues, SmPDR2 displayed the lowest abundance, with Ct values approximately at 38. In roots, SmPDR4 and SmPDR21 exhibited higher transcript abundance with a Ct value around 21. These results demonstrated the tissue-specific responses of different PDR members to heavy metal stress.
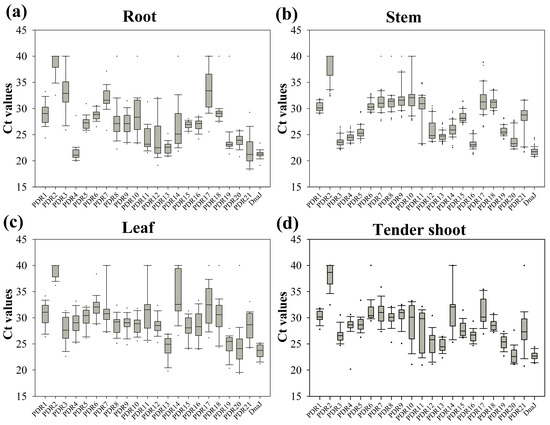
Figure 7.
Ct values for PDR genes by qRT-PCR. Expression data is displayed as Ct values for each gene in all samples. (a) Root; (b) stem; (c) leaf; and (d) tender shoot. A line across the box shows the median. The box indicates the 25th and 75th percentiles; whisker caps represent the maximum and minimum values. Outliers are symbolized by dots.
3.9. Effects of Different Heavy Metal Treatments on SpPDR Gene Expression Levels
The 21 PDR genes exhibited diverse transcription patterns under heavy metal stress, and no clear tissue-specific expression was observed in the control samples (Figure 8a). Interestingly, two PDR members, PDR7 and PDR17, displayed similar transcription patterns across all the analyzed tissues. The expression levels of these two genes were significantly increased in roots and stems treated with Zn0 after 1 day of treatment, in leaves after 7 days of Cu100 treatment, and in tender shoots after 4 days of Cu100 treatment (Figure 8b–d). Additionally, five genes (PDR2, PDR3, PDR16, PDR18, and PDR20) in roots and three genes (PDR4, PDR5, and PDR14) in leaves were significantly up-regulated after 4 days of treatment with Cd100 (Figure 8b,d). In addition, previous transcriptome data indicated that the expression levels for 14 PDR genes in roots were significantly enhanced by treatment with 100 μM CdCl2 (Table S3) [24]. In addition, three members (PDR7, PDR17, and PDR21) in roots, seven members (PDR1, PDR2, PDR7, PDR8, PDR9, PDR17, and PDR18) in stems, and only one member (PDR14) were strongly induced after 1 day of Zn0 treatment, respectively (Figure 8b,c,e). Therefore, different PDR members were regulated by different heavy metals.
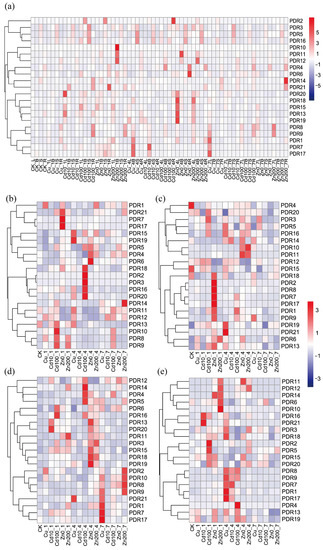
Figure 8.
The expression levels of all PDR genes are in response to different heavy metal treatments in all tissues. (a) All tissues; (b) root; (c) stem; (d) leaf; and (e) tender shoot. The highest values are shown as the most intense red; the lowest values are shown as the most intense blue; and the values in between are a lighter shade of either blue or red.
Furthermore, PDR5 and PDR16 showed the highest positive correlation, while PDR3 and PDR9 exhibited the strongest negative correlation across all examined tissues (Figure 9). This result suggests that the expression patterns of PDR members do not significantly correlate with their homology. To summarize the expression patterns of PDR genes in four tissues, we paid close attention to the genes with expression changes over twofold under different heavy metal stresses (Table S4). In roots, Cu100 enhanced the expression levels of PDR8, PDR9, PDR10, and PDR1. Additionally, Cd100, Zn0, and Zn200 treatments increased the expression levels of PDR3, whereas Zn0 and Zn200 repressed its expression. In the leaves, all heavy metals upregulated PDR4, and Cd100, Zn0, and Zn200 treatments also induced the expression of PDR6. Additionally, both PDR10 and PDR11 were upregulated by Zn0 and Zn200. Only Cd10 decreased the expression levels of PDR17. In the tender shoots, all heavy metals elevated the expression levels of PDR9. However, Cu100 and Zn0 inhibited the expression levels of PDR6 and PDR2, respectively. In the stems, PDR4 was downregulated by all heavy metals, while PDR6 was also induced by Cu100, Cd100, and Zn0 treatments (Table S4). These results highlight the complex and multifaceted response of PDR members to heavy metals.
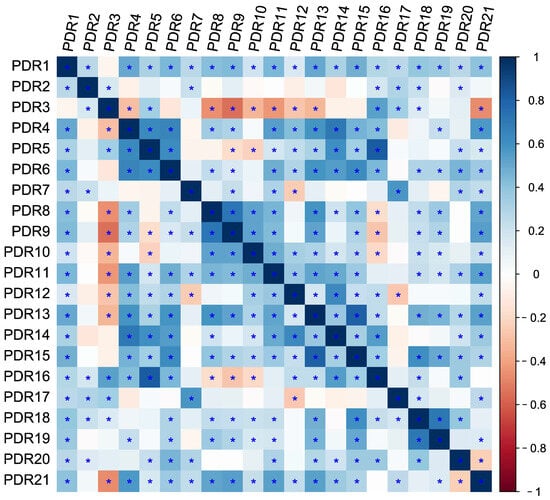
Figure 9.
Heatmap of the correlation coefficient between PDR genes. “*” represents p < 0.01.
4. Discussion
ABC transporters are a widely distributed group of transmembrane transporters that can be observed in various organisms. Among these transporters, ABCG proteins are a subgroup that belongs to the ABC transporter family. They possess two conserved structural domains: TMDs and NBDs in plants and microbiology [42,43]. The PDR subfamily shows a diverse array of substrates and is widely present in plants, microorganisms, and humans. These transporters have shown the potential to enhance the tolerance and accumulation capacity of organisms to heavy metals, making them valuable tools for bioremediation in heavy metal-contaminated soils [19]. Therefore, they are considered important candidates for plant improvement. Currently, only a few PDR genes have been recognized for their involvement in heavy metal tolerance in plants [18,44,45]. Thus, conducting a genome-wide analysis is necessary to clarify the biological roles of the PDR family in response to heavy metal stress. The accessibility of the whole genome of S. purpurea enables the comprehensive characterization of PDR family genes.
We identified a total of 21 SpPDR genes in the genome of S. purpurea. The structural domains of the TMDs and NBDs in these 21 SpPDR protein sequences exhibited a high degree of conservation and similarity with the PDR sequences from other species, such as Cucumis sativus [46]. It is important to mention that the number of genes specific to the PDR families is relatively stable across different species, implying that they evolve at a slower rate and exhibit less variation in gene copy numbers [19,44,47].
A comprehensive phylogenetic analysis of the PDR gene families was built from seven plant species using the neighbor-joining method. According to the phylogenetic analysis, 21 SpPDRs were classified into five groups, which align with the classification of known PDRs [35,36]. PDR proteins in Arabidopsis, N. tabacum, and S. purpurea share a common subcellular localization on the plasma membrane, aligning with their transport functions [35,48]. Early studies in this field have preliminarily shown that some members of the ABCG subfamily, namely AtABCG11, AtABCG12, AtABCG13, and AtABCG32, located on the cell membrane, have a function in pathways related to lipid metabolism. Specifically, these members are involved in regulating the transport of wax and keratin in Arabidopsis. Furthermore, increased wax and keratin content has been found to enhance resilience to abiotic stress in Arabidopsis [49,50]. Nearly half of the PDR genes had LTR elements, indicating that they could respond to low temperature stress [51]. TCA-element and P-box were representative of hormone-related elements [52]. Overall, the promoters of SpPDRs contain cis-acting elements associated with stress, growth and development, and hormone responses, suggesting that the transcriptional regulation of SpPDR genes is controlled by multiple stimuli.
Tandem gene duplications were observed on chromosomes one, three, and ten based on the analysis of gene location (Figure 4). The distribution of tandem gene duplications was found to be similar to that of cucumber’s PDR gene [46]. Consistently, collinearity analysis further supported the relatively conserved evolution of SpPDR genes, revealing divergence between herbaceous and woody plants. The evolution process in plants has the potential to impact the quantity and arrangement of introns [53]. Regarding the analysis of introns and exons, most SpPDR gene families had minimal difference in the number of introns and exons. Additionally, SpPDR possessed a splicing pattern of 21–25 exons, which aligns with previous findings in other plants [54].
Plants display gene expression patterns that are related to their responses to abiotic stress under dynamic growing conditions [55]. The expressions of ZmABC genes had different responses to Pb stress conditions [47]. The AtPDR8 gene is known to regulate Cd uptake via transit, leading to reduced Cd accumulation and improved tolerance to heavy metals. WRKY13 functions upstream of AtPDR8 to enhance Cd tolerance by directly binding to its promoter and activating its transcription [56]. Meanwhile, AtPDR8 also interacted with calmodulin 7 (CaM7) to enhance nonhost resistance [57]. In N. tabacum, most PDR genes were induced by jasmonate or salicylic acid and were also responsive to abiotic stresses [35]. In our study, all PDR genes were expressed in every tissue, with most of them being stimulated by at least two metal ions under heavy metal stresses. Moreover, PDR5 and PDR16 showed the highest positive correlation, while PDR3 and PDR9 exhibited the strongest negative correlation (Figure 9). The important function of PDR genes in the stress responses of plant-specific tissues is attributed to their varied expression patterns. These results will make a significant contribution to further investigation of the biological roles of PDR genes, specifically their involvement in the regulation of heavy metal tolerance in willow trees.
5. Conclusions
In this study, a total of 21 SpPDR proteins were identified in S. purpurea and further classified into five subfamilies. Our results revealed that these genes exhibited highly conserved intron architectures and motif compositions throughout their evolution process. Furthermore, the synteny analysis and Ka/Ks ratios indicated that SpPDRs experienced tandem repeat events and underwent purification selection during their evolutionary history. Most PDR genes highly responded to the heavy metal stresses and displayed different expression levels in the four different tissues. Overall, these findings aid in enhancing comprehension of the functions of the PDR genes in response to heavy metal stresses. The provided results will act as a base for additional research focused on uncovering the specific functions of the PDR genes and developing Salix variations with improved tolerance to heavy metals.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13092330/s1, Table S1: Primer sequences for qRT-PCR; Table S2: Segmentally duplicated PDR gene pairs in S. purpurea; Table S3: The expression of the PDR gene in S. matsudana under Cd; Table S4: Overview of PDR genes in response to different heavy metal stresses.
Author Contributions
Designed and conducted the experiments, S.C. and X.H.; performed the experiments and analyzed the data, S.C., J.C., Z.L., Y.J., Y.Y. and X.H.; conducted the data and manuscript—writing, S.C.; contributed to the discussion in the writing process, X.H., R.Z. and J.C.;manuscript—revision, X.H. and J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (No. 2021YFD2200201) to R.Z. and the National Natural Science Foundation of China (No. 31872168) to X.H.
Data Availability Statement
The S. purpurea genome (v5.1) sequence and protein sequence information files were downloaded from the JGI Genome Portal database (https://genome.jgi.doe.gov/portal/).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sharma, P.; Kumar, S. Bioremediation of heavy metals from industrial effluents by endophytes and their metabolic activity: Recent advances. Bioresour. Technol. 2021, 339, 125589. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Sun, H.F.; Fan, J.H.; Li, Y.Y.; Ma, L.J.; Wang, L.L.; Li, X.M. Transcriptome modulation by endophyte drives rice seedlings response to Pb stress. Ecotoxicol. Environ. Saf. 2023, 254, 114740. [Google Scholar] [CrossRef] [PubMed]
- Ihtisham, M.; Noori, A.; Yadav, S.; Sarraf, M.; Kumari, P.; Brestic, M.; Imran, M.; Jiang, F.; Yan, X.; Rastogi, A. Silver nanoparticle′s toxicological effects and phytoremediation. Nanomaterials 2021, 11, 2164. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Rastogi, A.; Shukla, A.; Srivastava, S.; Yadav, S. Prospects of genetic engineering utilizing potential genes for regulating arsenic accumulation in plants. Chemosphere 2018, 211, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Li, J.T.; Gurajala, H.K.; Wu, L.H.; van der Ent, A.; Qiu, R.L.; Baker, A.J.M.; Tang, Y.T.; Yang, X.E.; Shu, W.S. Hyperaccumulator plants from China: A synthesis of the current state of knowledge. Environ. Sci. Technol. 2018, 52, 11980–11994. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Qiu, R.; Zeng, X.; Fang, X.; Yu, F.; Zhou, X.; Wu, Y. Zn and Cd hyperaccumulating characteristics of Picris divaricata Vant. Int. J. Environ. Pollut. 2009, 38, 26–38. [Google Scholar] [CrossRef]
- Chen, C.; Zhang, H.; Wang, A.; Lu, M.; Shen, Z.; Lian, C. Phenotypic plasticity accounts for most of the variation in leaf manganese concentrations in Phytolacca americana growing in manganese-contaminated environments. Plant Soil 2015, 396, 215–227. [Google Scholar] [CrossRef]
- Yang, S.X.; Deng, H.; Li, M.S. Manganese uptake and accumulation in a woody hyperaccumulator, schima superba. Plant Soil Environ. 2008, 54, 441–446. [Google Scholar] [CrossRef]
- Pilipović, A.; Zalesny, R.S., Jr.; Rončević, S.; Nikolić, N.; Orlović, S.; Beljin, J.; Katanić, M. Growth, physiology, and phytoextraction potential of poplar and willow established in soils amended with heavy-metal contaminated, dredged river sediments. J. Environ. Manag. 2019, 239, 352–365. [Google Scholar] [CrossRef]
- Palm, E.; Guidi Nissim, W.; Mancuso, S.; Azzarello, E. Split-root investigation of the physiological response to heterogeneous elevated Zn exposure in poplar and willow. Environ. Exp. Bot. 2021, 183, 104347. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, F.; Liu, J.L.; Wu, H.T.; Yang, H.; Shi, Y.; Liu, J.; Zhang, Y.F.; Luo, Y.R.; Chen, K.M. Heavy metal transporters: Functional mechanisms, regulation, and application in phytoremediation. Sci. Total Environ. 2022, 809, 151099. [Google Scholar] [CrossRef] [PubMed]
- Dahuja, A.; Kumar, R.R.; Sakhare, A.; Watts, A.; Singh, B.; Goswami, S.; Sachdev, A.; Praveen, S. Role of ATP-binding cassette transporters in maintaining plant homeostasis under abiotic and biotic stresses. Physiol. Plant 2021, 171, 785–801. [Google Scholar] [CrossRef] [PubMed]
- Rees, D.C.; Johnson, E.; Lewinson, O. ABC transporters: The power to change. Nat. Rev. Mol. Cell Biol. 2009, 10, 218–227. [Google Scholar] [CrossRef] [PubMed]
- Dean, M.; Hamon, Y.; Chimini, G. The human ATP-binding cassette (ABC) transporter superfamily. J. Lipid Res. 2001, 42, 1007–1017. [Google Scholar] [CrossRef] [PubMed]
- Hollenstein, K.; Dawson, R.J.; Locher, K.P. Structure and mechanism of ABC transporter proteins. Curr. Opin. Struct. Biol. 2007, 17, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Lane, T.S.; Rempe, C.S.; Davitt, J.; Staton, M.E.; Peng, Y.; Soltis, D.E.; Melkonian, M.; Deyholos, M.; Leebens-Mack, J.H.; Chase, M.; et al. Diversity of ABC transporter genes across the plant kingdom and their potential utility in biotechnology. BMC Biotechnol. 2016, 16, 47. [Google Scholar] [CrossRef] [PubMed]
- Mishra, A.K.; Choi, J.; Rabbee, M.F.; Baek, K.H. In silico genome-wide analysis of the ATP-binding cassette transporter gene family in Soybean (Glycine max L.) and their expression profiling. Biomed. Res. Int. 2019, 2019, 8150523. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Lee, K.; Lee, J.; Noh, E.W.; Lee, Y. AtPDR12 contributes to lead resistance in Arabidopsis. Plant Physiol. 2005, 138, 827–836. [Google Scholar] [CrossRef]
- Kim, D.Y.; Bovet, L.; Maeshima, M.; Martinoia, E.; Lee, Y. The ABC transporter AtPDR8 is a cadmium extrusion pump conferring heavy metal resistance. Plant J. 2007, 50, 207–218. [Google Scholar] [CrossRef]
- Oda, K.; Otani, M.; Uraguchi, S.; Akihiro, T.; Fujiwara, T. Rice ABCG43 is Cd inducible and confers Cd tolerance on yeast. Biosci. Biotechnol. Biochem. 2011, 75, 1211–1213. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Peng, Z.; Li, J.; Huang, W.; Liu, Y.; Wang, X.; Xie, S.; Sun, L.; Han, E.; et al. Ectopic expression of poplar ABC transporter PtoABCG36 confers Cd tolerance in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 3293. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Wang, S.; Sun, H.; Chen, Y.; Wang, D.; Pan, H.; Zou, Y.; Liu, J.; Zheng, L.; Zhao, X.; et al. Comparative of Quercus spp. and Salix spp. for phytoremediation of Pb/Zn mine tailings. Environ. Sci. Pollut. Res. Int. 2017, 24, 3400–3411. [Google Scholar] [CrossRef] [PubMed]
- Landberg, T.; Greger, M. Phytoremediation using willow in industrial contaminated soil. Sustainability 2022, 14, 8449. [Google Scholar] [CrossRef]
- Han, X.; Zhang, Y.; Yu, M.; Zhang, J.; Xu, D.; Lu, Z.; Qiao, G.; Qiu, W.; Zhuo, R. Transporters and ascorbate-glutathione metabolism for differential cadmium accumulation and tolerance in two contrasting willow genotypes. Tree Physiol. 2020, 40, 1126–1142. [Google Scholar] [CrossRef]
- Yu, M.; Zhuo, R.; Lu, Z.; Li, S.; Chen, J.; Wang, Y.; Li, J.; Han, X. Molecular insights into lignin biosynthesis on cadmium tolerance: Morphology, transcriptome and proteome profiling in Salix matsudana. J. Hazard. Mater. 2023, 441, 129909. [Google Scholar] [CrossRef]
- Dos Santos Utmazian, M.N.; Wieshammer, G.; Vega, R.; Wenzel, W.W. Hydroponic screening for metal resistance and accumulation of cadmium and zinc in twenty clones of willows and poplars. Environ. Pollut. 2007, 148, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Kushwaha, H.R.; Kumar, K.; Verma, P.K. Comparative structural modeling of a monothiol GRX from chickpea: Insight in iron-sulfur cluster assembly. Int. J. Biol. Macromol. 2012, 51, 266–273. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Han, X.; Chen, S.; Zheng, L.; He, X.; Liu, M.; Qiao, G.; Wang, Y.; Zhuo, R. Selection of suitable reference genes for quantitative real-time PCR gene expression analysis in Salix matsudana under different abiotic stresses. Sci. Rep. 2017, 7, 40290. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Jiang, J.; Han, X.J.; Zhang, Y.X.; Zhuo, R.Y. Identification, expression analysis of the Hsf family, and characterization of class A4 in Sedum Alfredii Hance under cadmium stress. Int. J. Mol. Sci. 2018, 19, 1216. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Cao, P.; Wang, Z.; Gao, J.; Wu, M.; Li, X.; Zhang, J.; Wang, Y.; Gong, D.; Yang, J. Genome-wide characterization and expression profiling of the PDR gene family in tobacco (Nicotiana tabacum). Gene 2021, 788, 145637. [Google Scholar] [CrossRef] [PubMed]
- Moons, A. Transcriptional profiling of the PDR gene family in rice roots in response to plant growth regulators, redox perturbations and weak organic acid stresses. Planta 2008, 229, 53–71. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Song, X.; He, H.; Chu, L.; Zhou, H.; Zhao, Y.; Xu, Y.; Zeng, W.; Lin, X.; Lu, M.-Z. Genome-wide identification of plasma membrane aquaporin gene family in Populus and functional identification of PIP1;1 involved in osmotic stress. Environ. Exp. Bot. 2020, 179, 104200. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef]
- Wang, J.; Wang, Z.; Jia, C.; Miao, H.; Zhang, J.; Liu, J.; Xu, B.; Jin, Z. Genome-wide identification and transcript analysis of TCP gene family in banana (Musa acuminata L.). Biochem. Genet. 2022, 60, 204–222. [Google Scholar] [CrossRef]
- Zhang, C.; Tong, C.; Cao, L.; Zheng, P.; Tang, X.; Wang, L.; Miao, M.; Liu, Y.; Cao, S. Regulatory module WRKY33-ATL31-IRT1 mediates cadmium tolerance in Arabidopsis. Plant Cell Environ. 2023, 46, 1653–1670. [Google Scholar] [CrossRef]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, S. Structure and mechanism of ABC transporters. F1000Prime Rep. 2015, 7, 14. [Google Scholar] [CrossRef] [PubMed]
- Bienert, M.D.; Baijot, A.; Boutry, M. ABCG transporters and their role in the biotic stress response. In Plant ABC Transporters; Springer: Cham, Switzerland, 2014; pp. 137–162. [Google Scholar] [CrossRef]
- Moons, A. Ospdr9, which encodes a PDR-type ABC transporter, is induced by heavy metals, hypoxic stress and redox perturbations in rice roots. FEBS Lett. 2003, 553, 370–376. [Google Scholar] [CrossRef]
- Kobae, Y.; Sekino, T.; Yoshioka, H.; Nakagawa, T.; Martinoia, E.; Maeshima, M. Loss of AtPDR8, a plasma membrane ABC transporter of Arabidopsis thaliana, causes hypersensitive cell death upon pathogen infection. Plant Cell Physiol. 2006, 47, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Q.; Zhang, J.; Zhang, W.; Nie, J. Genome-wide characterization, phylogenetic and expression analysis of ABCG gene subfamily in cucumber (Cucumis sativus L.). Front. Plant Sci. 2023, 14, 1178710. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Yuan, X.; Li, L.; Zeng, M.; Yang, J.; Tang, H.; Duan, C. Genome-wide analysis of the ATP-binding cassette (ABC) transporter family in Zea mays L. and its response to heavy metal stresses. Int. J. Mol. Sci. 2022, 23, 2109. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.; Hwang, J.U.; Lee, M.; Kim, Y.Y.; Assmann, S.M.; Martinoia, E.; Lee, Y. PDR-type ABC transporter mediates cellular uptake of the phytohormone abscisic acid. Proc. Natl. Acad. Sci. USA 2010, 107, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Pighin, J.A.; Zheng, H.; Balakshin, L.J.; Goodman, I.P.; Western, T.L.; Jetter, R.; Kunst, L.; Samuels, A.L. Plant cuticular lipid export requires an ABC transporter. Science 2004, 306, 702–704. [Google Scholar] [CrossRef]
- Panikashvili, D.; Savaldi-Goldstein, S.; Mandel, T.; Yifhar, T.; Franke, R.B.; Höfer, R.; Schreiber, L.; Chory, J.; Aharoni, A. The Arabidopsis DESPERADO/AtWBC11 transporter is required for cutin and wax secretion. Plant Physiol. 2007, 145, 1345–1360. [Google Scholar] [CrossRef]
- Wu, W.; Zhu, S.; Xu, L.; Zhu, L.; Wang, D.; Liu, Y.; Liu, S.; Hao, Z.; Lu, Y.; Yang, L.; et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022, 22, 25. [Google Scholar] [CrossRef]
- Yu, J.; Yuan, Y.; Dong, L.; Cui, G. Genome-wide investigation of NLP gene family members in alfalfa (Medicago sativa L.): Evolution and expression profiles during development and stress. BMC Genom. 2023, 24, 320. [Google Scholar] [CrossRef]
- Rogozin, I.B.; Sverdlov, A.V.; Babenko, V.N.; Koonin, E.V. Analysis of evolution of exon-intron structure of eukaryotic genes. Brief Bioinform. 2005, 6, 118–134. [Google Scholar] [CrossRef]
- Chen, P.; Li, Y.; Zhao, L.; Hou, Z.; Yan, M.; Hu, B.; Liu, Y.; Azam, S.M.; Zhang, Z.; Rahman, Z.U.; et al. Genome-wide identification and expression profiling of ATP-binding cassette (ABC) transporter gene family in pineapple (Ananas comosus (L.) Merr.) reveal the role of AcABCG38 in pollen development. Front. Plant Sci. 2017, 8, 2150. [Google Scholar] [CrossRef] [PubMed]
- Zarattini, M.; Farjad, M.; Launay, A.; Cannella, D.; Soulié, M.C.; Bernacchia, G.; Fagard, M. Every cloud has a silver lining: How abiotic stresses affect gene expression in plant-pathogen interactions. J. Exp. Bot. 2021, 72, 1020–1033. [Google Scholar] [CrossRef]
- Sheng, Y.; Yan, X.; Huang, Y.; Han, Y.; Zhang, C.; Ren, Y.; Fan, T.; Xiao, F.; Liu, Y.; Cao, S. The WRKY transcription factor, WRKY13, activates PDR8 expression to positively regulate cadmium tolerance in Arabidopsis. Plant Cell Environ. 2019, 42, 891–903. [Google Scholar] [CrossRef]
- Campe, R.; Langenbach, C.; Leissing, F.; Popescu, G.V.; Popescu, S.C.; Goellner, K.; Beckers, G.J.; Conrath, U. ABC transporter PEN3/PDR8/ABCG36 interacts with calmodulin that, like PEN3, is required for Arabidopsis nonhost resistance. New Phytol. 2016, 209, 294–306. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).