Abstract
The world’s arable land has been contaminated by heavy metals to a large extent, which has led to the decreasing availability of farmland. Thus, the remediation of heavy metal pollution deserves due attention, and phytoremediation is preferred. Hemp has been proposed as an ideal alternative crop for remediating heavy-metal-contaminated soil, owing to its well-developed roots, large biomass, and tolerance to heavy metals. Chelators can activate heavy metals to enhance plant absorption, but they may cause stress to plant growth. Therefore, it is very important to optimize the combination of chelator and plant (cultivar) for efficacious phytoremediation. The effects of different concentrations (2, 5, 10, 15, and 25 mmol·L−1) of the chelators disodium ethylene diamine tetra-acetate (EDTA) and citric acid (CA) on the seed germination and plant growth of hemp cultivars were investigated. Triple application of increasing concentrations of EDTA or CA two days apart gradually reduced the germination potential, germination rate, radicle length, and embryonic shoot length of hemp seed, although 2 mmol·L−1 of CA could even promote seed germination. Distinct varietal differences were found in the response of hemp to chelator stress. Under the scheme of four-time uses one week apart, both chelators caused a concentration-dependent linear decrease in the plant height, stem diameter, and biomass of hemp plants, but the growth inhibition due to CA was relatively milder. This could be partially explained by the change in tested physiological indices in hemp leaf. In conclusion, 2 mmol·L−1 of CA helped with seed germination and was almost nontoxic to plant growth; cultivar BM was more tolerant to the chelators than cultivar Y1.
1. Introduction
More than ten million sites of soil pollution have been reported worldwide, and more than half of them are due to heavy metals [1]. An extensive investigation revealed that soil pollution in China is also quite serious [2] and has caused huge economic losses [3]. Soil heavy-metal contamination causes serious impacts on the living environment and human health, and therefore has grown to become a hot topic in modern society.
For heavy-metal-contaminated soil, various remediation strategies, including engineering remediation (replacement of contaminated soil, soil removal and soil isolation, electrokinetic remediation, soil leaching, adsorption, etc.) and bioremediation (remediation using plants, microbes, and animals), are being proposed, developed, and tested, but most of them are very costly and only applicable to small contaminated sites [4]. Phytoremediation is the preferred way to mitigate heavy metal pollution, that is, using green plants to extract, absorb, transfer, decompose, or fix heavy metals in soil [4,5,6], removing or reducing environmental pollutants with environmentally friendly, simple operations which are economic and have applicable characteristics [7]. However, the small biomass and slow growth rate of hyperaccumulator plants if they are used as alleviators of soil heavy metals and the low availability of heavy metals in the soil make them difficult to absorb and be used by plants, therefore reducing phytoremediation efficiency [8]. To this end, finding a plant that is fast-growing, has a large biomass, and increases the availability of soil heavy metals in order to improve phytoremediation efficiency is an urgent task.
A chelator is a compound with a multitooth ligand; its ligand can combine with metal ions to form extremely water-soluble chelate [9], which can be used to activate heavy metals in soil, promoting the absorption and accumulation of heavy metals by plants [10,11]. Meanwhile, a chelator can also be a stress factor that impedes seed germination and plant growth [12,13], and the obstruction varies by plant species, cultivar, and chelator type [14,15,16,17,18]. Therefore, investigation of the effect of chelators on plant growth would set the foundation for the selection of an optimal plant cultivar, which is very critical for improving the feasibility and efficiency of phytoremediation using the combination of chelators and plants.
Hemp (Cannabis sativa L.) is an annual herb of the genus Cannabis from the family Cannabinaceae and a multipurpose crop with a very long history of cultivation and utilization by human beings [19]. Hemp has well-developed roots, a large biomass, strong resistance to abiotic stress, a fast growth rate, and a short lifecycle and has strong tolerance to heavy metals and considerable potential for the enrichment of heavy metals. Therefore, it has been suggested as an ideal alternative crop for remediating soils contaminated by heavy metals [5,20,21,22,23,24], and the fibers produced from heavy metal (such as cadmium, lead, and zinc)-contaminated soil can be safely used in the textile industry [25].
The ability of the chelators disodium ethylene diamine tetra-acetate (EDTA) and citric acid (CA) to activate the heavy metal Pb in the substrate, resulting in a significant increase in the content of available Pb in the substrate and a significant increase in the uptake of Pb by hemp plants, was determined using a pot culture experiment [26], but the influences of chelators on hemp growth and the optimal concentration of chelators that can be used with hemp have not been reported up to now. In this paper, we used seven hemp cultivars originating from different parts of China as the materials, applying a paper culture for testing seed germination and a pot culture (two cultivars were selected from the above seven according to their germination performance in responding to chelator treatment) for testing the plant growth of hemp cultivars under chelator treatments of different concentrations, and investigated the impacts of the chelators EDTA and CA on seed germination, plant growth, and some physiological indices of hemp cultivars in order to screen out an optimized combination of “plant + chelator”, and, furthermore, provide reasonable reference and experimental data support for enhancing the remediation efficiency of soil heavy-metal pollution by using a chelator combined with a specific plant.
2. Materials and Methods
2.1. Materials
The materials were seven hemp cultivars from different production areas of China. Of them, ‘Yunma 1’ (Y1), ‘Yunma 7’ (Y7), and ‘Yunwan 6’ (YW6) were collected from Yunnan Province; ‘Bamahuoma’ (BM) from Guangxi Autonomous Region; ‘Jinma 1’ (J1) from Shanxi Province; ‘Wanma 1’ (W1) from Anhui Province; and ‘Qingma 1’ (Q1) from Heilongjiang Province. The analytical reagent chelators disodium ethylene diamine tetra-acetate (EDTA-2Na, simplified as EDTA) and citric acid (CA) were provided by XiLong Science Company Limited, Kunming, China (Lot No. 1609031). The substrate for pot culture was the commercial peat produced by JIFFY Company, Stange, Norway, with basic nutrient content (mg·kg−1) of total nitrogen 5200, hydrolyzed nitrogen 292, total phosphorus 760, Olsen phosphorus 338, total potassium 1780, available potassium 859, and pH 5.6.
2.2. Seed Germination Test
Hemp seeds were rinsed with distilled water and surface water was drained using filter paper. Thirty healthy seeds of uniform size were placed evenly into a 9 cm diameter Petri dish (with cover) laid with two layers of filter paper. To each dish, 5 mL EDTA or CA solution of different concentration, i.e., 0 (CK, no chelator), 2, 5, 10, 15, or 25 mmol·L−1, was added, and each treatment was repeated thrice. The chelator concentration levels in the study were set based on the results of a preparatory test, together with considerations of the execution feasibility of the experiment. The dishes with seeds were cultured in a light incubator with 12 h/d light of intensity 6800 lx, day/night temperature 24 °C/19 °C. During the culture, 2 mL of the designed chelator solution was added to each dish at day 3 and day 5 to maintain the water needed for seed germination and to keep the persistent stress of chelator.
2.3. Pot Culture of Plants
Depending on the results of the seed germination test, two cultivars, Y1 and BM, showing significant differences in germination rate were selected. Thirty carefully selected hemp seeds were sown into each plastic pot with tray of 40 cm diameter and 26 cm height; 3.5 kg peat and 15 g NPK (15-10-15) compound fertilizer were added in each pot and stirred well. The pots with seeds were cultured in a plastic greenhouse (Chenggong campus, Yunnan University) and quantitatively watered for moisturizing the culture substrate. At the stage of third-pair leaves, seedlings were thinned and eight plants with similar growth status in each pot were kept. EDTA or CA solution was applied starting from the third day after thinning at concentrations of 0 (CK), 2, 5, 10, and 15 mmol·L−1, with three replicates for each treatment. The 25 mmol·L−1 concentration used in seed germination test was omitted from this experiment, based on the observations in our preparatory test that the 25 mmol·L−1 chelator was too toxic to hemp growth and plant survival for the necessary tests. In total, 500 mL chelator solution (prepared using distilled water) was applied into each pot once a week for four times. Substrate moisture in the pots was maintained at 70–80% of field capacity throughout the plant growth period.
2.4. Parameters and Test Methods
2.4.1. Seed Germination Indices
Seed germination was observed and recorded every day for seven days. Seeds with radicle grown to more than half of the seed length were counted as germinated. Germination potential (GP) and germination rate (GR) were calculated following the formulas [27]: GP (%) = (number of seeds germinated on day 3/number of seeds tested) × 100; GR (%) = (number of seeds germinated on day 7/number of seeds tested) × 100. Radicle length (RL) and embryonic shoot length (SL) were measured for five germinated seeds randomly sampled from each dish when the germination was counted on day 7; all germinated seeds were used for this measurement if the germinated seeds were five or less.
2.4.2. Plant Growth and Physiological Indices
Seven weeks after the first application of chelator solution, leaf samples, the third pair of leaves counted from the first plant-top expanded leaf, were collected from five plants randomly selected from each pot during 9:00–10:00 a.m. Fresh leaves were cleaned and immediately stored in liquid nitrogen and then at −80 °C for further analysis. The relative chlorophyll contents (presented as SPAD reads) of intact functional leaves were determined using a TYS-A Chlorophyll Meter, provided by Zhejiang Top Instrument Co., Ltd. (Hangzhou, China) [28]. Malondialdehyde (MDA) content was measured via a method of thiobarbituric acid, soluble protein (SP) using Coomassie brilliant blue, superoxide dismutase (SOD) activity using nitroblue tetrazolium reduction [29], and peroxidase (POD) activity using guaiacol [30].
After collection of the samples for physiological analysis, the plant height and stem diameter of all plants in each pot were measured using a flexible rule and vernier caliper. The plants were harvested, then stems and leaves were separated and placed in different paper bags for drying in an oven at 108 °C for 20 min to inactivate enzymes and then dried at 80 °C. The roots were collected carefully and cleaned in running water, surface water was drained with filter paper, and dried at 80 °C. Dry weight of root, stem, and leaf (the leaf sample for physiological parameters was also counted) of the plants in each pot was quantified, and the total weight of root, stem, and leaf (plant biomass) was calculated accordingly for each treatment.
2.5. Data Process and Analysis
Every index of each treatment was achieved from three biological replicates. The data were sorted and graphed using Microsoft Excel 2010. ANOVA and LSD multiple comparisons (p < 0.05) were implemented using SPSS Statistics for Windows, version 25.0 (IBM Corp., Armonk, NY, USA).
3. Results
3.1. Effect of Chelators on Seed Germination of Seven Hemp Cultivars
3.1.1. Effect of Chelators on Hemp Seed Germination
Significant impacts on GP, GR, RL, and SL of a single factor, i.e., chelator type, chelator concentration, and hemp cultivar, and interactions of chelator type × chelator concentration and chelator type × hemp cultivar were observed. Moreover, the interaction of chelator concentration × hemp cultivar evidently influenced GP and RL, and the interaction of the three factors remarkably affected the RL and SL of germinated hemp seeds (Table 1).

Table 1.
Variance analysis for effects of chelators and their concentrations on hemp seed germination.
3.1.2. Effect of EDTA on Seed Germination
When treated with different concentrations of EDTA, except that BM increased its GP by a slight amount under a low EDTA concentration (2 mmol·L−1), the GPs of the other six cultivars were reduced to varied degrees, and the reductions were concentration-dependent. All the cultivars had decreased GPs along with the increase in EDTA concentration; of them, Y1 significantly decreased its GP by 47.91% compared with CK even under a treatment of 2 mmol·L−1. However, Y7, BM, W1, and J1 decreased GPs significantly in comparison with CK only under high concentrations (15~25 mmol·L−1), but YW6 under treatment of EDTA with the designed concentrations did not vary its GP of statistical importance. It is worth noting that J1 had the lowest GP under high concentrations of EDTA (15~25 mmol·L−1), with only 15.63~21.88% of CK (Table 2).
Table 2.
Effects of EDTA on germination potential (GP), germination rate (GR), radicle length (RL), and embryonic shoot length (SL) of hemp seed.
All seven cultivars decreased their GRs as EDTA concentration increased. Of them, Y1 and YW6 significantly decreased their GR by 40.39% and 32.61% compared with CK, even under the lowest EDTA concentration of 2 mmol·L−1, while the GR of BM was less affected by EDTA, maintaining 63% of CK even under a concentration of 25 mmol·L−1. The GRs of the seven cultivars were all remarkably lower than those of CK when EDTA concentration ≥ 10 mmol·L−1 (Table 2).
EDTA treatment significantly reduced both the RLs and SLs of the germinated seeds, with reductions in RLs and SLs, respectively, from 44.80% to 98.92% and 26.83% to 93.01%, showing evident concentration dependence. When EDTA concentration reached 25 mmol·L−1, the maximal RL and SL of the tested cultivars were just 0.16 cm and 0.60 cm, representing only 2.4% and 17.2% of CK (Table 2).
3.1.3. Effect of CA on Seed Germination
Except for YW6, the GPs of the other six cultivars increased to different extents when treated with CA at 2 mmol·L−1, while the GP of BM increased significantly by the greatest amount of 33.0%. However, as the concentration increased, the inhibition of CA to the GP in all the cultivars gradually augmented, although the inhibition was the greatest in W1, which reduced the GP to only 28.5% of CK under the treatment of CA at 25 mmol·L−1. But cultivar J1 did not show evident differences in GP when treated with CA of different concentrations (Table 3).
Table 3.
Effects of CA on germination potential (GP), germination rate (GR), radicle length (RL), and embryonic shoot length (SL) of hemp seed.
In general, as for GP, CA at low concentrations promoted the seed germination of hemp, but suppressed it at higher concentrations. All seven cultivars increased their GRs under treatment of low-concentration CA at 2 mmol·L−1. Among them, Y7, BM, and J1 persisted, with GRs higher than that of CK until the CA concentration exceeded 5 mmol·L−1, although the tested cultivars decreased their GRs by 31.03~51.67% over that of CK under CA at 25 mmol·L−1 (Table 3).
When treated with a low concentration of CA at 2 mmol·L−1, except for the mild reductions in Y1 and J1, the other five cultivars slightly increased their RLs, and W1 still had a longer radicle than CK even under treatment of CA at 5 mmol·L−1. As that for the GP and GR, hemp RLs reduced significantly under high concentrations of CA, and Y7 and Q1 reduced RLs to the greatest extents, retaining only 9.11~17.15% of CK under CA at 25 mmol·L−1 (Table 3).
Again, we observed that the hemp cultivars increased the SLs over CK under a low concentration CA at 2 mmol·L−1, except for cultivars BM and YW6, while Y7 and Q1 had the greatest increases in SLs achieving 1.37 and 1.22 times that of CK. But the inhibition of CA to the growth of the embryonic shoot was gradually augmented along with the increased concentration, and the SLs of all the cultivars under high concentrations of CA were remarkably reduced in comparison with CK. YW6 showed the greatest reduction in SL, with only 24.43% of CK remaining under CA at 25 mmol·L−1 (Table 3).
In general, comparison of the effect of the two chelators on hemp seed germination in 20 groups (five concentrations × four indices) resulted in only 2 groups (indicated in bold) showing the same stress effects of EDTA and CA, while the other 18 groups (labeled with letters a, b) showed a greater stress effect of EDTA than that of CA. Therefore, for hemp seed germination, the suppression of chelator CA was milder than that of EDTA (Table 4).
Table 4.
Comparison of the adverse effects of EDTA and CA on germination potential (GP), germination rate (GR), radicle length (RL), and embryonic shoot length (SL) of hemp seed.
3.2. Effects of Chelators on Plant Growth and Physiological Attributes of Two Hemp Cultivars
3.2.1. Effect of Chelators on Plant Growth of Hemp Cultivars
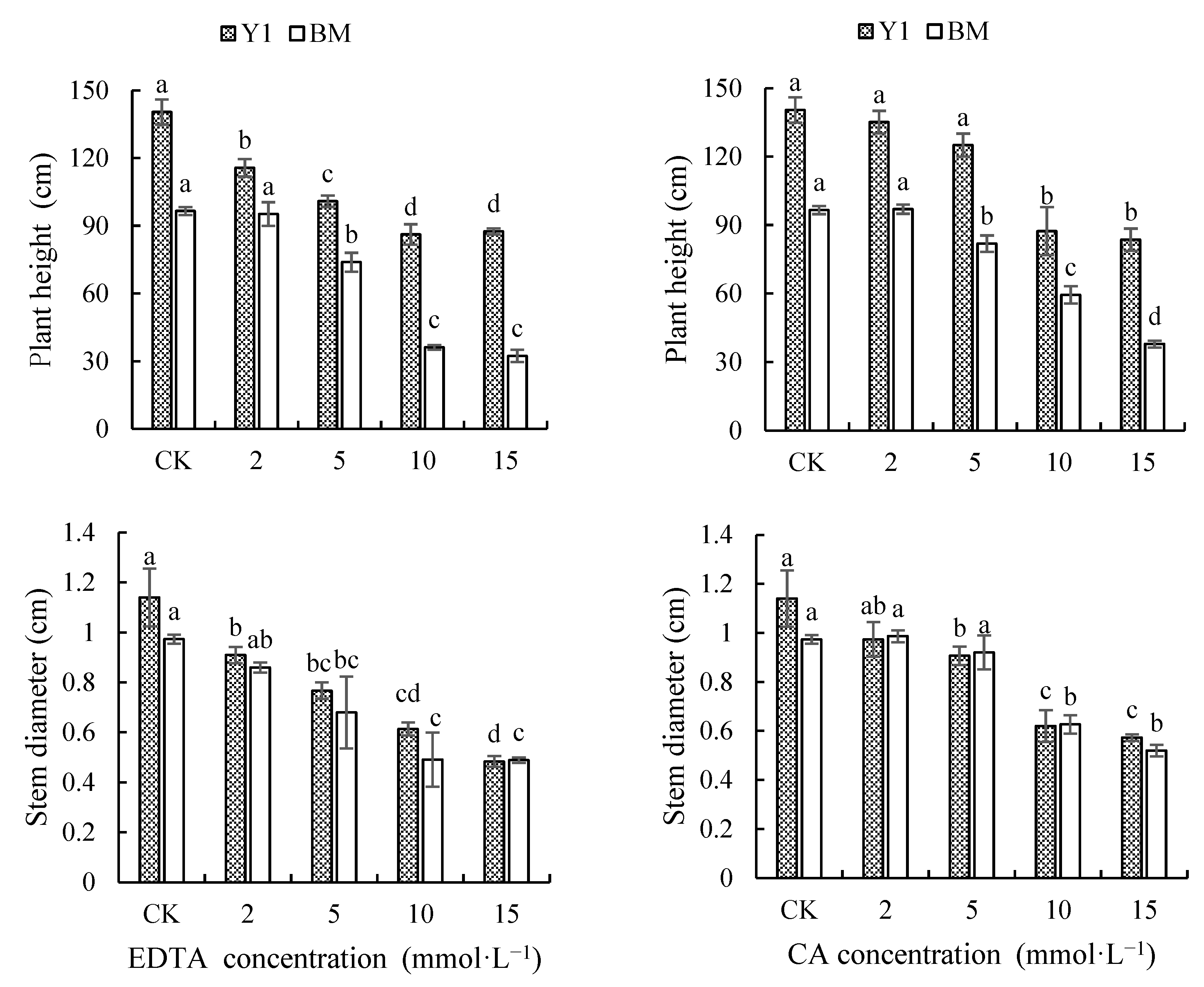
As shown in Figure 1, the application of different types and concentrations of chelators reduced the plant height and stem diameter of cultivar Y1, but this was observed in BM only when chelators of higher concentration (5 mmol·L−1 or more) were applied. However, chelators caused a greater reduction in plant height in BM than in Y1; average reductions of 38.48% and 28.49% in BM and 30.53% and 23.26% in Y1 for plant height were recorded, respectively, by EDTA and CA of the four concentrations together. On the contrary, a greater reduction in stem diameter in Y1 was caused by chelators compared with that in BM; average reductions of 39.18% and 32.60% in Y1 and 35.27% and 21.58% in BM for stem diameter were recorded, respectively, for EDTA and CA treatment.
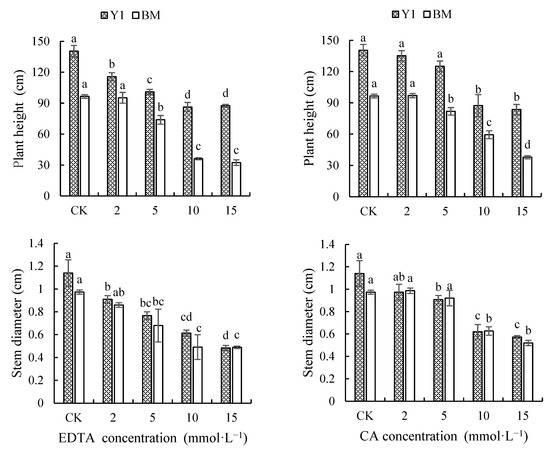
Figure 1.
Effects of chelators and their concentrations on plant height and stem diameter of two hemp cultivars. Note: Different letters present the significant effect of chelators at different concentrations within cultivar (p < 0.05, LSD). EDTA—disodium ethylene diamine tetra-acetate; CA—citric acid; Y1—hemp cultivar ‘Yunma 1’; BM—hemp cultivar ‘Bamahuoma’.
The biomass of root, stem, and leaf of both cultivars was reduced by the chelator treatment, and only the biomass of BM showed a slight increase at low concentrations of CA (2 mmol·L−1). The reduction in hemp biomass caused by chelator treatment showed a concentration effect; for example, the greatest reduction in biomass was observed when the concentration of chelator EDTA increased to 15 mmol·L−1, leaving only 27.99% and 31.25% of CK for Y1 and BM, respectively (Table 5).
Table 5.
Effects of chelators and their concentrations on biomass of hemp in different parts (g·plant−1).
In addition, CA reduced the three growth indices of both cultivars less than that caused by EDTA when treated with the same concentration of chelator (labeled with letters a, b). Of the 12 combinations comparing the adverse effects of CA and EDTA, only 3 did not show significant differences (marked by bold numbers), implying that CA caused less inhibition of hemp plant growth in general than EDTA (Table 6).
Table 6.
Comparison of the adverse effects of EDTA and CA on plant growth of two hemp cultivars.
3.2.2. Effect of Chelators on MDA Content in Hemp Leaves
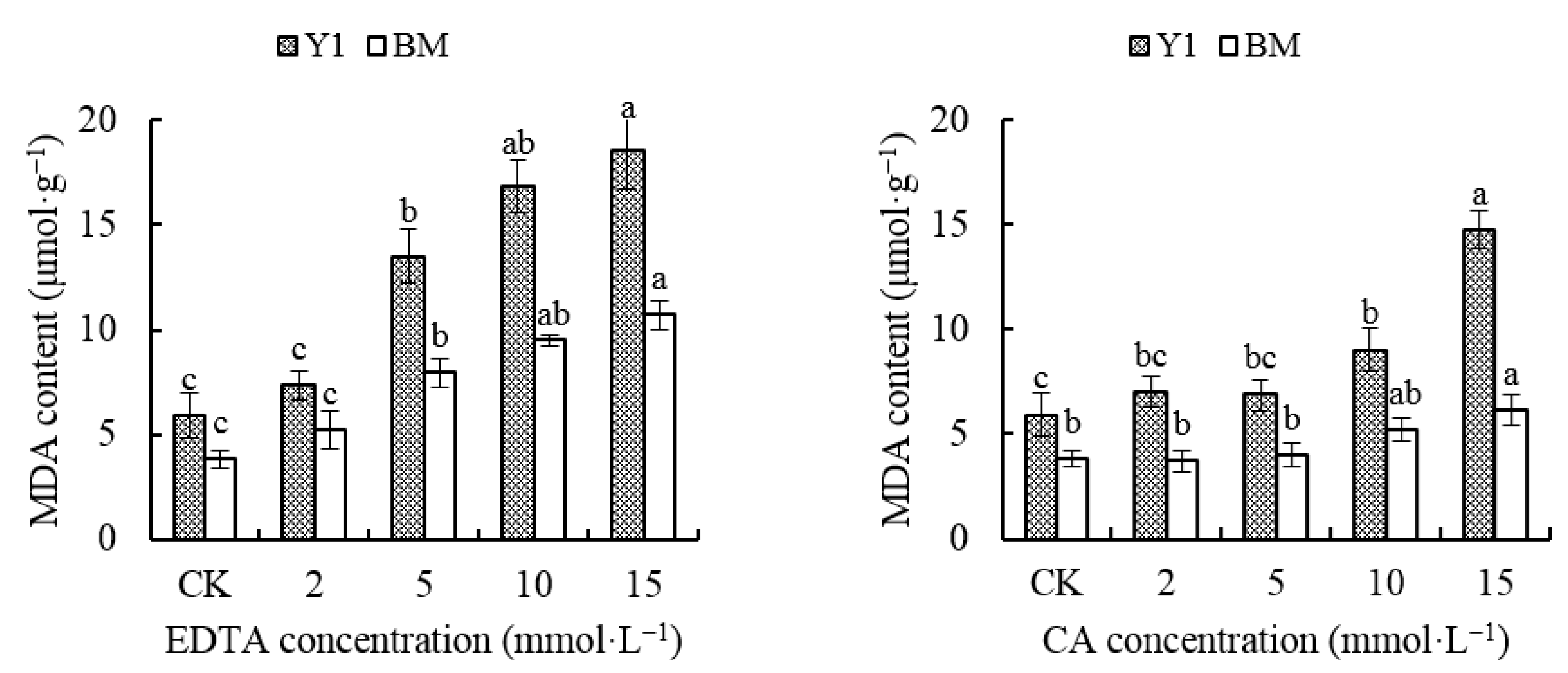
The MDA content in the hemp leaves gradually increased as the chelator concentration increased. The MDA content in BM leaves was less than that in Y1 under the same chelator concentration, while for the same cultivar, the MDA content of CA-treated leaves was less than that of EDTA-treated leaves (Figure 2). When EDTA was applied, the MDA content increased significantly over CK in both cultivars, Y1 and BM, starting from the concentration of 5 mmol·L−1, but until the concentration of CA rose up to 10 mmol·L−1, the MDA content in BM leaves did not rise obviously compared with that of CK. The MDA content of Y1 leaves increased to 3.12 times and 2.49 times over the content in CK, while that of BM was 2.53 times and 1.60 times the content in CK, under treatment of EDTA or CA at 15 mmol·L−1 (Figure 2).
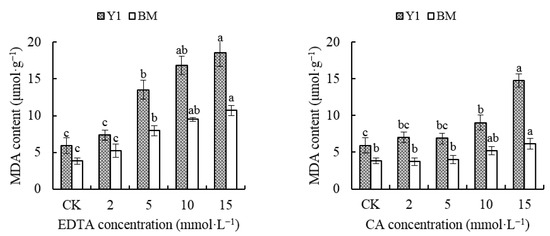
Figure 2.
Effects of chelators and their concentrations on malondialdehyde (MDA) content in the leaves of two hemp cultivars. Note: Different letters present the significant effect of chelators at different concentrations within the cultivar (p < 0.05, LSD). EDTA—disodium ethylene diamine tetra-acetate; CA—citric acid; Y1—hemp cultivar ‘Yunma 1’; BM—hemp cultivar ‘Bamahuoma’.
3.2.3. Effect of Chelators on Activities of SOD and POD in Hemp Leaves
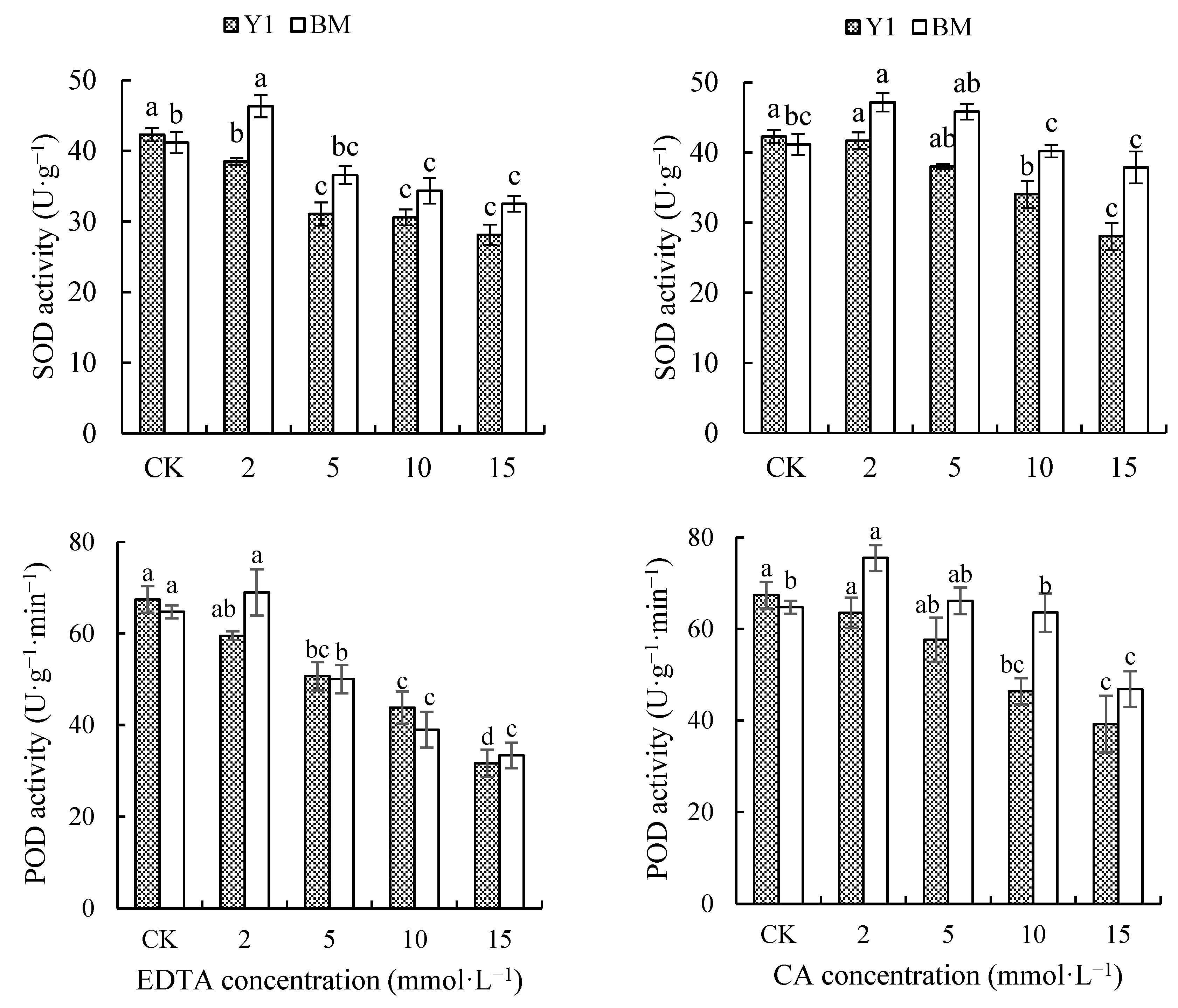
As shown in Figure 3, the application of different types and concentrations of chelators all resulted in a gradual decrease in the leaf SOD activity of Y1, while that in BM even showed a slight increase under chelator treatments at a low concentration (2 mmol·L−1). Under the treatments of EDTA or CA at 15 mmol·L−1, the leaf SOD activity of Y1 was decreased by 33.50% and 33.62%, but that of BM only decreased by 25.93% and 7.99%, compared with CK.
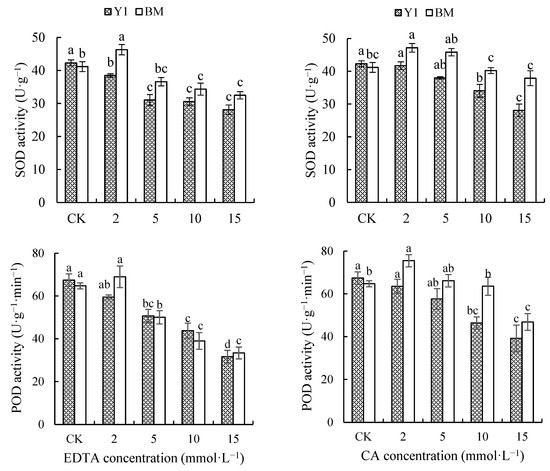
Figure 3.
Effects of chelators and their concentrations on the activity of superoxide dismutase (SOD) and peroxidase (POD) in the leaves of two hemp cultivars. Note: Different letters present the significant effect of chelators at different concentrations within the cultivar (p < 0.05, LSD). EDTA—disodium ethylene diamine tetra-acetate; CA—citric acid; Y1—hemp cultivar ‘Yunma 1’; BM—hemp cultivar ‘Bamahuoma’.
The application of different types and concentrations of chelators also caused a decrease in leaf POD activity in Y1, while that in BM again showed a slight increase at a low concentration (2 mmol·L−1) of chelators. Under treatments of EDTA or CA at 15 mmol·L−1, the leaf POD activity of Y1 was decreased by 53.08% and 41.83%, and that of BM decreased by 48.48% and 27.61%, compared with CK (Figure 3).
3.2.4. Effect of Chelators on Contents of Chlorophyll and SP in Hemp Leaves
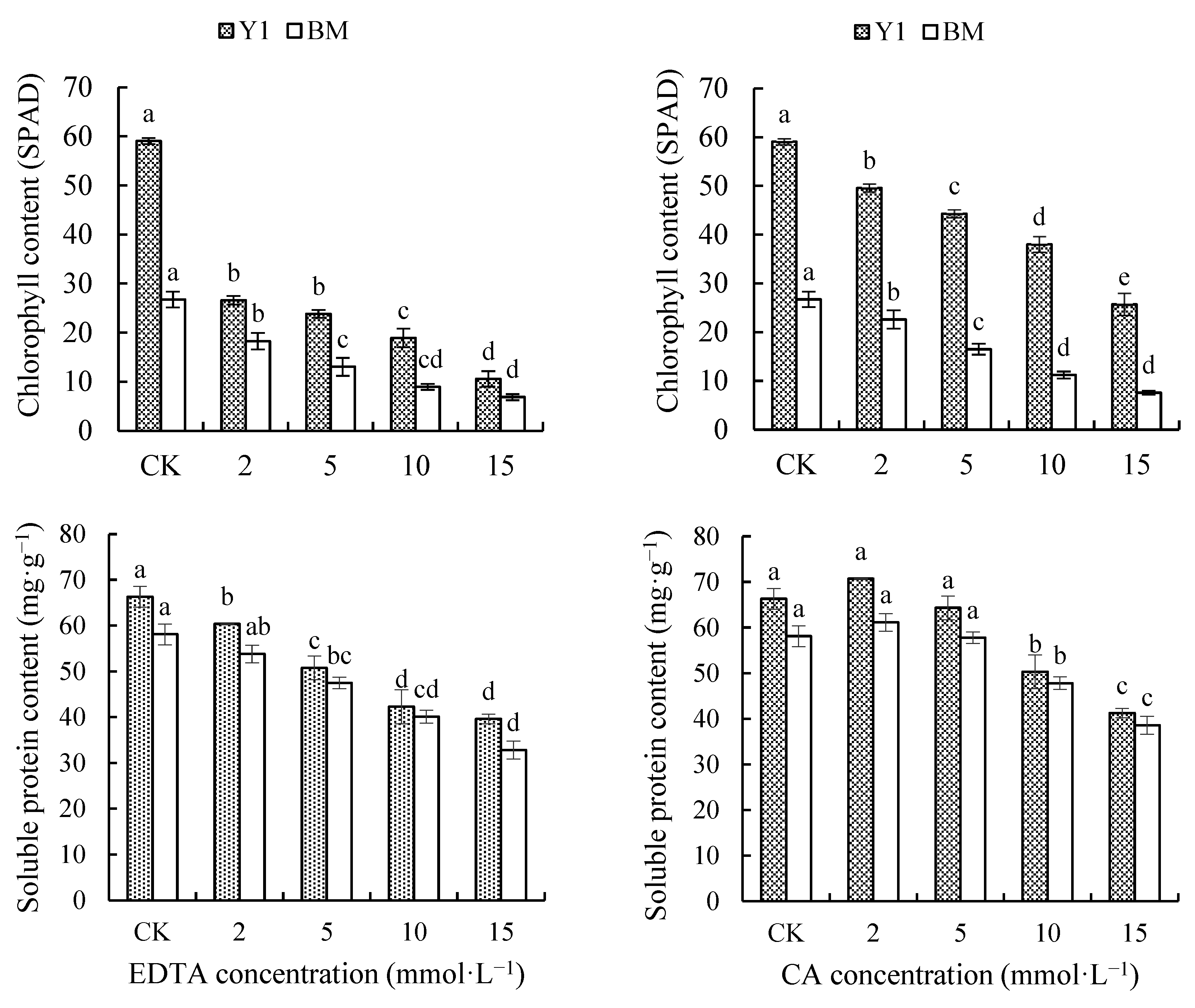
As chelator concentration increased, the total relative chlorophyll content (presented as SPAD reads) in both Y1 and BM leaves was decreased persistently and significantly (Figure 4). The total chlorophyll content of Y1 treated with 15 mmol·L−1 EDTA or CA remained at only 17.91% and 43.50% that of CK, and that of BM remained at 25.74% and 28.26% of CK. Here, a greater impact of EDTA on chlorophyll content in hemp leaves could be viewed, supported by the fact that EDTA caused more reduction in chlorophyll content, compared with CK, than CA caused (Figure 4).
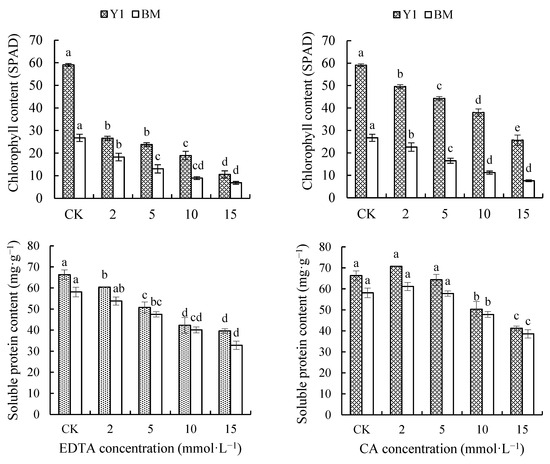
Figure 4.
Effects of chelators and their concentrations on the content of relative chlorophyll (SPAD reads) and soluble protein (SP) in the leaves of two hemp cultivars. Note: Different letters present the significant effect of chelators at different concentrations within the cultivar (p < 0.05, LSD). EDTA—disodium ethylene diamine tetra-acetate; CA—citric acid; Y1—hemp cultivar ‘Yunma 1’; BM—hemp cultivar ‘Bamahuoma’.
SP content gradually decreased with increasing EDTA concentration, while it even slightly increased under a low concentration (2 mmol·L−1) of CA treatment. When 15 mmol·L−1 EDTA or CA was applied, the SP content of Y1 leaves was reduced by 40.24% and 37.77%, and that of BM leaves decreased by 43.50% and 33.59%, compared with CK (Figure 4). Moreover, at the same concentration, the SP content of EDTA-treated plants was reduced more than that of CA-treated plants, indicating that EDTA caused worse damage to hemp.
4. Discussion
The application of different concentrations of EDTA caused gradual declination in the seed GP and GR of the seven cultivars, while CA even promoted the germination of hemp seeds at low concentrations. In another studies, the GR of maize seed was significantly reduced after prolonged treatment with EDTA, and the germination index decreased rapidly with increasing EDTA concentration [31,32]. This may be explained, as CA is a natural organic acid of small molecule that can be used as a reaction substrate to participate in plant metabolism [33] or as a growth regulator [34] to improve seed germination, while EDTA is a synthetic amino polycarboxylic acid with biological toxicity [35].
As for the obvious variation in the seed germination of the tested cultivars under zero chelator treatment, this may be due to differences in cultivar characteristics, harvesting, processing, and storage conditions that affected seed quality, as seeds of these cultivars were collected from research institutions in different regions of China. Fortunately, we were not interested in the absolute difference of seed germination, but rather “the reduction or increase of seed germination” in comparison with the germination at zero level of chelator treatment (CK). It was noted that the GPs of cultivar YW6 treated with EDTA and the GPs of J1 treated with CA were gradually reduced along with the concentration increment, but the reduction showed no statistical significance over the designed concentrations. This might be due to the relatively low germination rate, or the fluctuation of GPs among replicates of a treatment, but the actual biological mechanism needs to be explored.
From the effect of chelators on the growth of radicles and embryonic shoots of hemp cultivars, the RL decreased more than the SL, which was especially obvious when treated with EDTA; EDTA treatment shortened the RLs of all seven cultivars significantly in this experiment, with decreases of 76.16~94.82% over CK. The inhibition of chelators on radicle growth was greater than that on embryonic shoots, probably because roots are important primary nutrient absorbers of plants, and all the substances required for plant growth need to be absorbed through roots and transported to other organs, so that radicle growth was worse hindered due to more direct poisoning to root cells caused by chelators [36].
The growth of plants, especially the seedlings, provides a more visual response to the surrounding environmental conditions, and growth delays are the most important response of plants to adverse circumstances [37]. The application of EDTA caused a significant decrease in the plant height, stem diameter, and total biomass of hemp plants, while CA at low concentrations (2 mmol·L−1) even promoted the growth of hemp plants, and CA only at high concentrations showed a toxic effect, which can be partially explained by the reduction pattern of chlorophyll content in hemp leaves outlined in Figure 2 after the treatment of chelators. Similar phenomena were observed in previous studies on plants such as wheat [38], Beta vulgaris var. cicla [39], and Hemerocallis fulva [40]. The plant height, stem diameter, and biomass of hemp under CA treatment all were higher than those of EDTA treatment at same concentration. This can be interpreted with how organic acid CA with low concentrations can moderately decrease soil pH and then improve the soil environment and enhance microbial activity, thus favoring plant growth [41,42], while EDTA exerts a toxic effect on plants and inhibits their growth [43].
Reactive oxygen species (ROS) produced by plants during senescence or encountering biotic and abiotic stress would oxidize the structural backbone phospholipid molecules of cell membranes, thus damaging the membranes, while MDA is one of the main products in this process, and its content can reflect the degree of damage to the cell membrane system [44]. Simultaneously, a large amount of ROS will prompt the activation of the antioxidant enzyme system in plants as compensation to remove ROS from plants by increasing the activity of antioxidant enzymes such as SOD and POD to eradicate or alleviate the damage caused by ROS to plant cell membranes [45]. In our experiment, the leaf MDA content of the two hemp cultivars increased with the increasing chelator concentration. This indicates that chelator treatment induced excessive ROS production in the plants and caused damage to the cytoplasmic membrane, as evidenced by the significant increase in MDA content and the stunted growth of hemp plants.
The antioxidant enzyme activities varied between hemp cultivars. SOD and POD activities in Y1 gradually decreased with increasing chelator concentration, while BM showed a slight increase in leaf antioxidant enzyme activities under low chelator concentrations (2 mmol·L−1). The reason for the inconsistent changes in Y1 and BM antioxidant enzyme activities may be attributable to differences in the cultivars, with Y1 being less tolerant of chelator stress. Under the present experimental conditions, the antioxidant enzyme activity in Y1 started to decrease when a low concentration (2 mmol·L−1) of chelators was applied, probably because the stress of this level of chelators exceeded the regulatory capacity of Y1’s own antioxidant enzyme system, and excessive accumulation of ROS in the plants caused damage to the cells, decreasing enzyme activity [46]. However, whether the antioxidant enzyme activity in hemp would increase if chelators were applied at a concentration less than 2 mmol·L−1 has yet to be studied. In contrast, BM was quite tolerant of the chelators; even under treatment of CA with a range of concentrations, BM still could increase the activity of antioxidant enzymes in vivo to maintain the balance of ROS metabolism, which was similar to previous findings in seagrass Cymodocea serrulata [44] and Alternanthera bettzickiana [47].
Photosynthetic pigment is essential for the absorption, transmission and conversion of light energy in plants, while soluble proteins are important enzymes or active molecules involved in physiological metabolic processes in plants, and the levels of the both can reflect the degree of damage to plants caused by adverse conditions [48]. In this experiment, the total relative chlorophyll content (SPAD reads) of Y1 and BM decreased gradually with increasing chelator concentration, showing that chelators had a remarkable destruction effect on photosynthetic pigments, because chelators inhibit chlorophyll synthesis and accelerate its degradation [49]. Soluble protein content decreased persistently with increasing EDTA concentration, but it increased in both Y1 and BM leaves under low-concentration CA treatment, indicating that CA facilitates protein synthesis and maintains the stability of the cellular antioxidant system, whereas EDTA causes damage to plant cells such that it decreases the ability to synthesize soluble protein.
It was demonstrated that chelators (EDTA and CA) activated the heavy metal Pb, causing a significant increase in the content of available Pb in the substrate and, therefore, a significant increase in the uptake of Pb by hemp plants, and the effect of chelators was closely related to the concentration used [26]. In detail, although hemp showed the best remediation efficacy when 5 mmol·L−1 EDTA or 10 mmol·L−1 CA was added in Pb-contaminated substrate, the growth of hemp, such as plant height, stem diameter, and biomass, increased when 2 mmol·L−1 EDTA and 2~5 mmol·L−1 CA were added, respectively, and the growth indices of hemp decreased in varying degrees under other chelator concentrations [26]. As the leaching of heavy metals due to enhanced mobility using EDTA-assisted phytoextraction would be one of the potential hazards [50], in the light of our study results that EDTA was more harmful than CA to plant growth, and the experimental results of Chen [27] that BM had a Pb tolerance coefficient (1.1–1.4) greater than Y1 (0.8–1.1), “cultivar BM + 2 mmol·L−1 of chelator CA” was recommended for the phytoremediation of soil heavy-metal contamination.
Although repeated applications of chelator (three times for seed germination and four times for seedling culture) to compensate for lost moisture may increase the overall chelator concentration, so that the results obtained may be due to a higher chelator concentration than the expressed, the results reported here still provide a useful reference for the phytoremediation of soil heavy-metal contamination.
5. Conclusions
Results showed triple application of chelators two days apart produced significant inhibition on the germination of hemp seeds. The GR, GP, RL, and SL of the seven hemp cultivars gradually decreased with increasing EDTA concentrations, but a low concentration of CA promoted seed germination. Of the cultivars, BM showed the strongest tolerance in seed germination to the chelators used. With the use of chelators four times one week apart, both EDTA and CA inhibited the growth of Y1 plants, disrupted the cytoplasmic membrane structure (MDA evidently increased), and continuously decreased the antioxidant enzyme activity, total chlorophyll content, and soluble protein content due to the increasing concentration. However, low concentrations of CA treatment even promoted BM plant growth by increasing antioxidant enzyme activity and soluble protein content in the leaves. Under the same concentration of chelator treatment, BM had a significantly lower MDA content and higher antioxidant enzyme activity than that of Y1. It was feasible to conclude that BM was more tolerant of chelators EDTA and CA than Y1, and CA had fewer adverse effects on the seed germination, plant growth, and physiological process of hemp than EDTA. Consequently, chelator CA (2 mmol·L−1) + cultivar BM might be a promising combination for reconciling plant growth and heavy-metal remediation efficiency.
Author Contributions
F.L.: Conceptualization, Funding acquisition, Supervision, Project administration, Methodology, Writing—review and editing; J.H.: Investigation, Formal analysis, Validation, Writing—original draft; Y.Z.: Investigation, Data curation, Formal analysis; X.L.: Investigation, Formal analysis; Y.Y.: Methodology, Writing—review and editing; G.D.: Writing—review and editing; K.T.: Writing—review and editing, Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the China Agriculture Research System of MOF and MARA under grant number CARS-16-E15, and the National Natural Science Foundation of China under grant number 31871671. The APC was funded by CARS-16-E15.
Data Availability Statement
Data available on request due to restrictions of privacy.
Acknowledgments
This study was kindly supported by China Agriculture Research System of MOF and MARA (CARS-16-E15) and National Natural Science Foundation of China “Comparative study of the adaptation molecular mechanisms of industrial hemp responding to salt and alkali stresses (31871671)”. The authors are grateful to Muzammal Rehman for reading and English editing our manuscript.
Conflicts of Interest
The authors declare that they have no known competing financial interest or personal relationships that could have appeared to influence the work reported in this paper.
Abbreviations
GP—germination potential; GR—germination rate; RL—radicle length; SL—embryonic shoot length; EDTA-2Na (simplified as EDTA)—disodium ethylene diamine tetra-acetate; CA—citric acid; MDA—malondialdehyde; SOD—superoxide dismutase; POD—peroxidase; SP—soluble protein.
References
- He, Z.; Shentu, J.; Yang, X.; Baligar, V.C.; Zhang, T.; Stoffella, P.J. Heavy metal contamination of soils sources, indicators, and assessment. J. Environ. Indic. 2015, 9, 17–18. [Google Scholar]
- Ministry of Environmental Protection and Ministry of Land and Resources of China. National Survey Report of Soil Contamination Status of China; Department of Environment and Natural Resources: Beijing, China, 2014; pp. 26–27. [Google Scholar]
- Yang, S.C.; Nan, Z.R.; Zeng, J.J. Current situation of soil contaminated by heavy metals and research advances on the remediation techniques. J. Anhui Agric. Sci. 2006, 34, 549–552. [Google Scholar]
- Su, C.; Jiang, L.Q.; Zhang, W.J. A review on heavy metal contamination in the soil worldwide: Situation, impact and remediation techniques. Environ. Skept. Crit. 2014, 3, 24–38. [Google Scholar]
- Rheay, H.T.; Omondi, E.C.; Brewer, C.E. Potential of hemp (Cannabis sativa L.) for paired phytoremediation and bioenergy production. GCB Bioenergy 2021, 13, 525–536. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, Y.X.; Qin, Y.; Deng, S.H. Progress of phytoremediation on soil polluted by heavy metal. Hubei For. Sci. Technol. 2016, 45, 40–43+63. [Google Scholar]
- Yang, P.; Chen, X.Y.; Tian, L.L.; Zhang, K.K.; Xu, L.L.; Guan, P. Effects of EDTA mixed cadmium stress on the seed germination an physiological resistance characters of initial growth in Astragalus sinicus. J. Mt. Agric. Biol. 2016, 35, 26–30+80. [Google Scholar]
- Udawatta, R.P.; Motavalli, P.P.; Garrett, H.E.; Krstansky, J.J. Nitrogen losses in runoff from three adjacent agricultural watersheds with claypan soils. Agric. Ecosyst. Environ. 2006, 117, 39–48. [Google Scholar] [CrossRef]
- Wuana, R.A.; Okieimen, F.E.; Imborvungu, J.A. Removal of heavy metals from a contaminated soil using organic chelating acids. Int. J. Environ. Sci. Technol. 2010, 7, 485–496. [Google Scholar] [CrossRef]
- Jalali, M.; Khanboluki, G. Redistribution of fractions of zinc, cadmium and lead in calcareous soils treated with EDTA. Arch. Agron. Soil Sci. 2007, 53, 147–160. [Google Scholar] [CrossRef]
- Meers, E.; Tack, F.M.G.; Verloo, M.G. Degradability of ethylenediaminedisuccinic acid (EDDS) in metal contaminated soils: Implications for its use soil remediation. Chemosphere 2008, 70, 358–363. [Google Scholar] [CrossRef]
- RöMkens, P.; Bouwman, L.; Japenga, J.; Draaisma, C. Potentials and drawbacks of chelate-enhanced phytoremediation of soils. Environ. Pollut. 2002, 116, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.Y.; Chen, L.S.; Zhu, Y. Effects of EDTA and EM composite agent on seed germination, seedling growth and lead uptake of ryegrass under lead stress. Guangdong Seric. 2020, 54, 22–25. [Google Scholar]
- Lee, J.; Sung, K. Effects of chelates on soil microbial properties, plant growth and heavy metal accumulation in plants. Ecol. Eng. 2014, 73, 386–394. [Google Scholar] [CrossRef]
- Wang, Y.K.; Chen, M.; Zhang, J.Q.; Yuan, B. Research progress of chelate- induced phytoremediation of soil contaminated by heavy metals. J. Hubei Polytech. Univ. 2014, 30, 30–32+40. [Google Scholar]
- Shi, Q.L. Leaching Characteristics od Pb and Cd in the Contaminated Soils by Organic Chelating Agents and Biosurfactant. Master’s Thesis, Southwest University, Chongqing, China, 2015. [Google Scholar]
- Wang, S.Y.; Duo, L.A.; Zhao, S.L. Effects of biodegradable chelator on growth and physiology of Festuca arundinacea seedlings. Acta Hortic. Sin. 2017, 44, 2186–2194. [Google Scholar]
- Diarra, I.; Kotra, K.K.; Prasad, S. Assessment of biodegradable chelating agents in the phytoextraction of heavy metals from multi-metal contaminated soil. Chemosphere 2021, 273, 128483. [Google Scholar] [CrossRef]
- Liu, F.H.; Hu, H.R.; Du, G.H.; Deng, G.; Yang, Y. Ethnobotanical research on origin, cultivation, distribution and utilization of hemp (Cannabis sativa L.) in China. Indian J. Tradit. Knowl. 2017, 16, 235–242. [Google Scholar]
- Galić, M.; Perčin, A.; Zgorelec, Ž.; Kisić, I. Evaluation of heavy metals accumulation potential of hemp (Cannabis sativa L.). J. Cent. Eur. Agric. 2019, 20, 700–711. [Google Scholar]
- Xu, Y.P.; Lü, P.; Zhang, Q.Y.; Guo, R.; Deng, G.; Guo, H.Y.; Yang, M. Comparison of the absorption and accumulation characteristics of five heavy metals among different industrial hemp varieties. J. Agric. Resour. Environ. 2020, 37, 106–114. [Google Scholar]
- Placido, D.F.; Lee, C.C. Potential of Industrial Hemp for Phytoremediation of Heavy Metals. Plants 2022, 11, 595. [Google Scholar] [CrossRef]
- Golia, E.E.; Bethanis, J.; Ntinopoulos, N.; Kaffe, G.-G.; Komnou, A.A.; Vasilou, C. Investigating the potential of heavy metal accumulation from hemp. The use of industrial hemp (Cannabis Sativa L.) for phytoremediation of heavily and moderated polluted soils. Sustain. Chem. Pharm. 2023, 31, 100961. [Google Scholar] [CrossRef]
- Wu, Y.; Trejo, H.X.; Chen, G.; Li, S. Phytoremediation of contaminants of emerging concern from soil with industrial hemp (Cannabis sativa L.): A review. Environ. Dev. Sustain. 2021, 23, 14405–14435. [Google Scholar] [CrossRef]
- De Vos, B.; Souza, M.F.; Michels, E.; Meers, E. Industrial hemp (Cannabis sativa L.) in a phytoattenuation strategy: Remediation potential of a Cd, Pb and Zn contaminated soil and valorization potential of the fibers for textile production. Ind. Crops Prod. 2022, 178, 114592. [Google Scholar] [CrossRef]
- Hu, J.M.; Zhang, Y.T.; Zhou, Y.; Liu, H.; Li, X.; Du, G.H.; Liu, F.H. Chelators could enhance hemp efficacy in remediation of Pb contamination in culture substrate. J. Yunnan Univ. Nat. Sci. Ed. 2022, 44, 1297–1304. [Google Scholar]
- Chen, Z.Y. Research of Seed Germination, Plant Growth and Physiology of Hemp Variety under Heavy Metal Pb Stress. Master’s Thesis, Yunnan University, Kunming, China, 2016. [Google Scholar]
- Hu, H.R. Effect of Salinity on Seed Germination and Seeding Growth of Hemp. Master’s Thesis, Yunnan University, Kunming, China, 2015. [Google Scholar]
- Li, H.S.; Sun, Q.; Zhao, S.J.; Zhang, W.H. Plant Physiological Biochemical Experiment; Higher Education Press: Beijing, China, 2006; pp. 260–261, 167–169, 184–185. [Google Scholar]
- Zhang, Z.L.; Zhai, W.J. Plant Physiological Experiment Instruction; Higher Education Press: Beijing, China, 2004. [Google Scholar]
- Liu, H. Research on Pollution Effects of Chelating Agents in Chelate-Induced Phytoremediation Technology. Master’s Thesis, Northeast Forestry University, Harbin, China, 2010. [Google Scholar]
- Zhang, Y.; Deng, B.; Li, Z. Inhibition of NADPH oxidase increases defense enzyme activities and improves maize seed germination under Pb stress. Ecotoxicol. Envion. Saf. 2018, 158, 187–192. [Google Scholar] [CrossRef]
- Li, H.Y. The Effects of Exogenous Citric and Oxalic Acid on Cadmium Induced Physiological Response in Boehmeria nivea (L.) Gaud. Master’s Thesis, Hunan University, Changsha, China, 2014. [Google Scholar]
- Wang, X.Y.; Peng, W.B. Effect of organic acids and boron and zinc on metabolism of active oxygen during filling period and seed weight of wheat. Sci. Agric. Sin. 1995, 28, 69–74. [Google Scholar]
- Sillanp, M.; Oikari, A. Assessing the impact of complexation by EDTA and DTPA on heavy metal toxicity using microtox bioassay. Chemosphere 1996, 32, 1485–1497. [Google Scholar] [CrossRef]
- Xu, W.J.; Guo, J.; Zhao, M.; Wang, R.Y.; Hou, S.Z.; Yang, Y.; Zhong, B.; Guo, H.; Liu, C.; Shen, Y.; et al. Research progress of soil plant root exudates in heavy metal contaminated soil. J. Zhejiang A F Univ. 2017, 34, 1137–1148. [Google Scholar]
- Liu, L. The Regulation and Mechanism of Phytohormone on Rice Seed Germination and Seeding Root Growth under Salinity. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2018. [Google Scholar]
- Ding, Z.H.; Hu, X.; Zhang, Y.F. Metal uptake of wheat seedlings and metal distribution after the addition of EDTA, [S, S]-EDDS and DTPA. Ecol. Environ. Sci. 2010, 19, 97–101. [Google Scholar]
- Ma, Y.; Zhao, G.L.; Wang, X.F.; Cheng, J.M. Remediation of lead and cadmium contaminated soil with chelate induced Beta vulgaris Var. Cicla L. Chin. J. Soil Sci. 2021, 52, 416–424. [Google Scholar]
- Guo, H.; Zhuang, J.J. Effect of citric acid amendment on cadmium uptake and translocation by three ornamental plants. J. Anhui Agric. Univ. 2021, 48, 121–127. [Google Scholar]
- Chen, Y.X.; Lin, Q.; Luo, Y.M.; He, Y.F.; Zhen, S.J.; Yu, Y.L.; Tian, G.M.; Wong, M.H. The role of citric acid on the phytoremediation of heavy metal contaminated soil. Chemosphere 2003, 50, 807–811. [Google Scholar] [CrossRef]
- Najeeb, U.; Xu, L.; Ali, S.; Jilani, G.; Gong, H.J.; Shen, W.Q.; Zhou, W.J. Citric acid enhances the phytoextraction of manganese and plant growth by alleviating the ultrastructural damages in Juncus effusus L. J. Hazard. Mater. 2009, 170, 1156–1163. [Google Scholar] [CrossRef]
- Aderholt, M.; Vogelien, D.L.; Koether, M.; Greipsson, S. Phytoextraction of contaminated urban soils by Panicum virgatum L. enhanced with application of a plant growth regulator (BAP) and citric acid. Chemosphere 2017, 175, 85–96. [Google Scholar] [CrossRef]
- Aljahdali, M.O.; Alhassan, A.B. Heavy metal accumulation and anti-oxidative feedback as a biomarker in Seagrass Cymodocea serrulata. Sustainability 2020, 12, 2841. [Google Scholar] [CrossRef]
- Abedi, T.; Paknyat, H. Antioxidant enzyme changes in response to drought stress in ten cultivars of oilseed rape (Brassica napus L.). Czech J. Genet. Plant Breed. 2010, 46, 27–34. [Google Scholar] [CrossRef]
- Zhu, S.J.; Gui, S.B.; Xu, W.; Xiang, P.; Meng, W.; Zhu, J.; Li, K.K. Experimental research on the tolerance of four emergent plants under high nitrogen and phosphorus pollution conditions. China Rural. Water Hydropower 2022, 2, 34–38. [Google Scholar]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Araújo, R.P.d.; Almeida, A.-A.F.d.; Pereira, L.S.; Mangabeira, P.A.O.; Souza, J.O.; Pirovani, C.P.; Ahnert, D.; Baligar, V.C. Photosynthetic, antioxidative, molecular and ultrastructural responses of young cacao plants to Cd toxicity in the soil. Ecotoxicol. Environ. Saf. 2017, 144, 148–157. [Google Scholar] [CrossRef]
- Li, F.M.; Xiong, Z.T.; Hu, H.Y. Effects of chelating agents on toxicity of copper to Elsholtzia splendens. Environ. Sci. 2003, 24, 96–100. [Google Scholar]
- Meers, E.; Qadir, M.; De Caritat, P.; Tack, F.M.; Du Laing, G.; Zia, M.H. EDTA-assisted Pb phytoextraction. Chemosphere 2009, 74, 1279–1291. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).