
Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Biostimulants
2.3. Biostimulant and Water Stress Treatments
2.4. Growth Parameters
2.5. Proline Quantification
2.6. Antioxidant Enzymes
2.6.1. Superoxide Dismutase (SOD)
2.6.2. Ascorbate Peroxidase (APx)
2.6.3. Glutathione Reductase (GR)
2.7. Antioxidant Compounds Determination
2.7.1. Total Phenolic Compounds (TPC) and Flavonoids (TF)
2.7.2. Ascorbic Acid (AsA)
2.8. Statistical Analysis
3. Results
3.1. Effects of Water Stress and Biostimulant Pre-Treatments on Tomato Plants
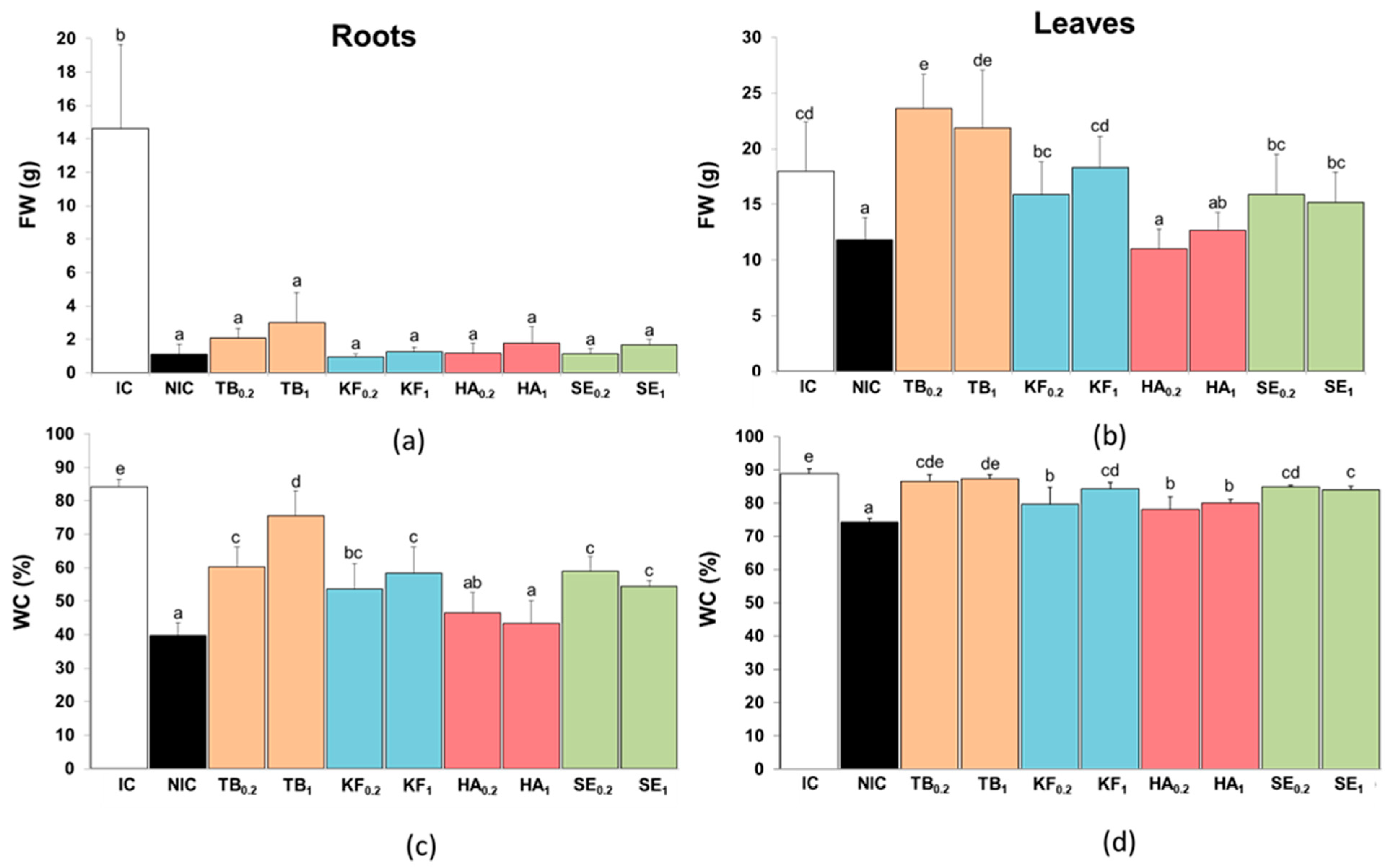
3.2. Growth Parameters
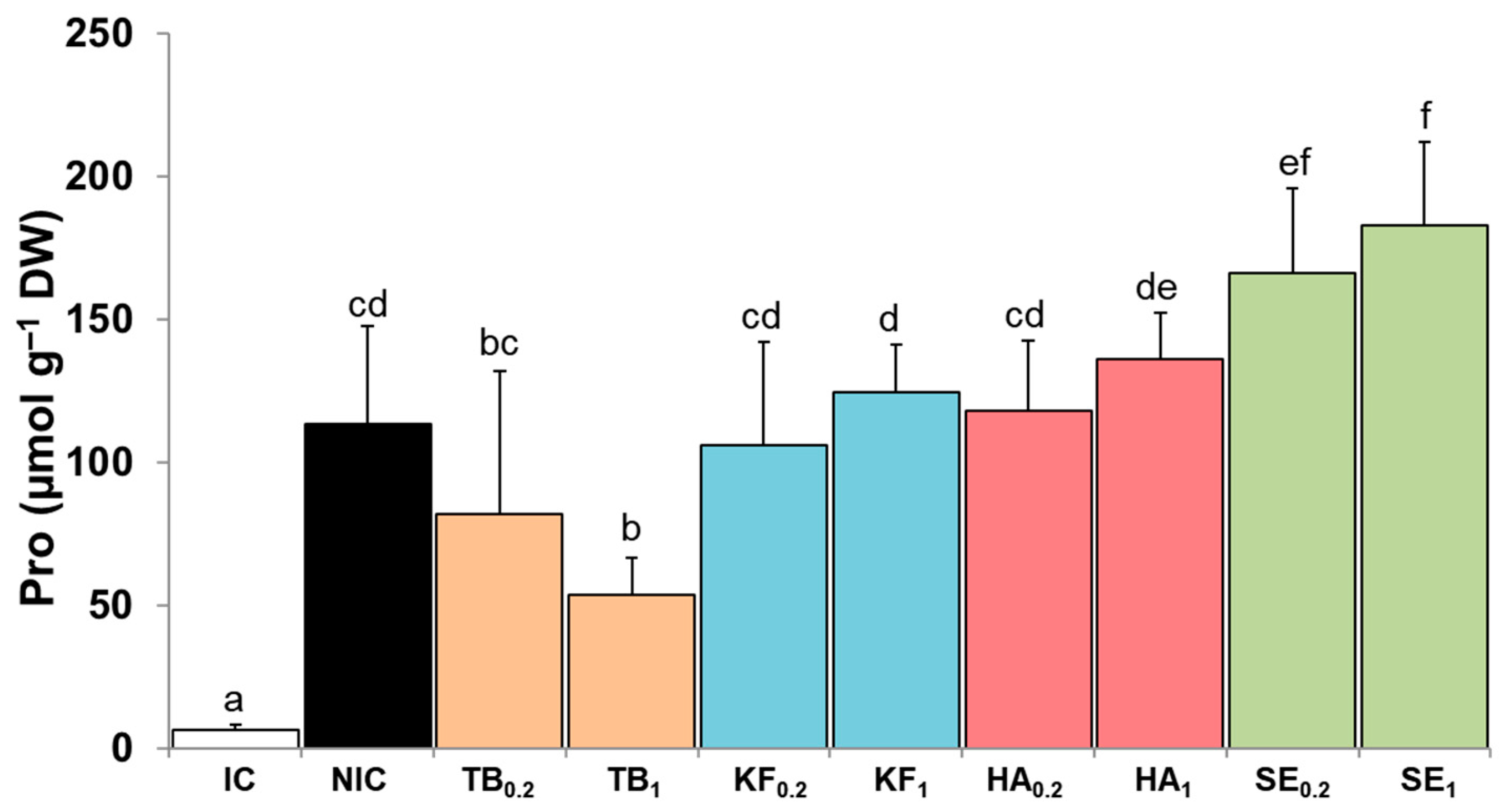
3.3. Proline Content
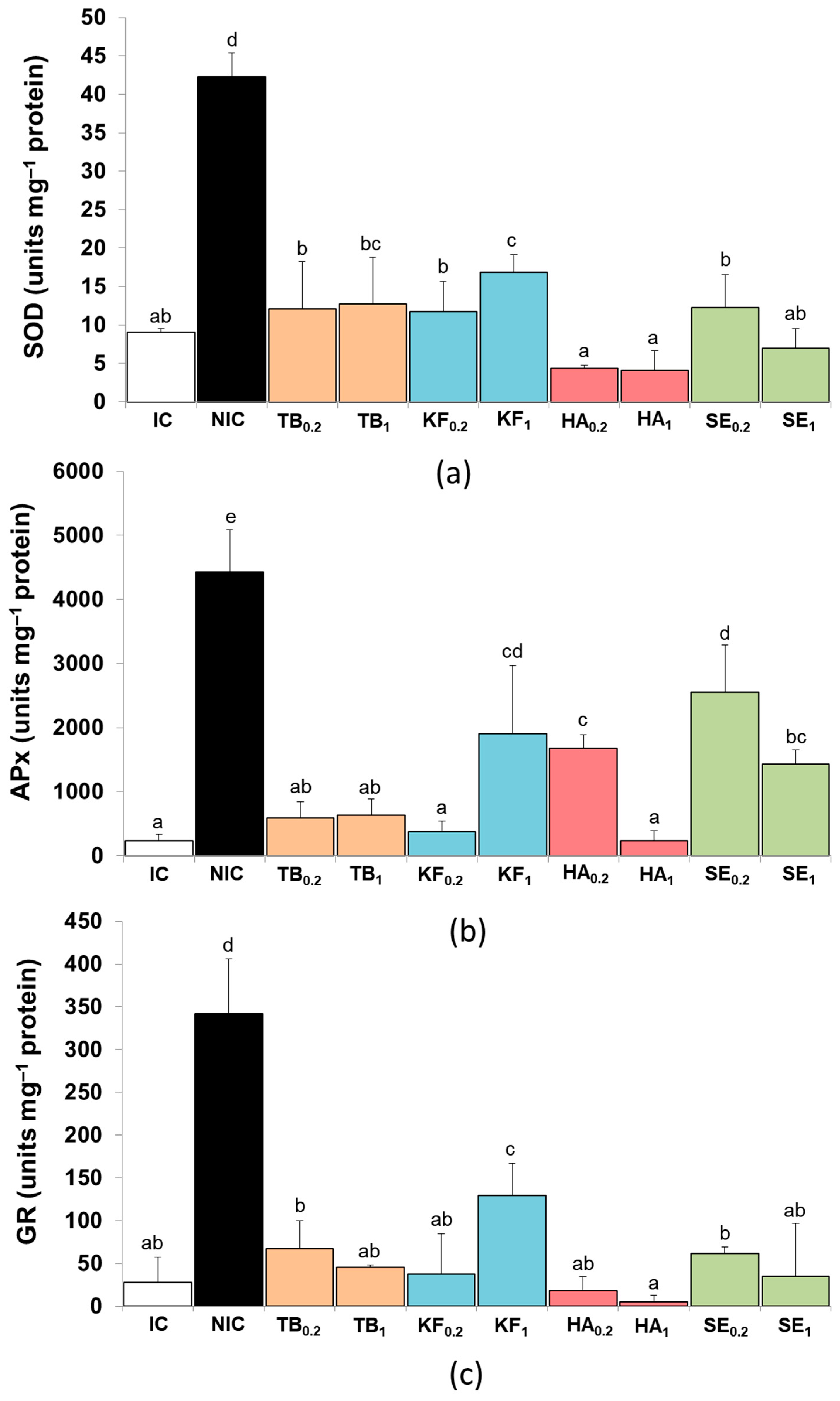
3.4. Antioxidant Enzymes
3.5. Antioxidant Compounds
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M. Mechanisms of drought and salt stress tolerance in plants. Sci. Bull. Ser. F Biotechnol. 2019, 23, 29–36. [Google Scholar]
- Kaur, G.; Asthir, B. Molecular responses to drought stress in plants. Biol. Plant. 2017, 61, 201–209. [Google Scholar] [CrossRef]
- Moradi, P. Key plant products and common mechanisms utilized by plants in water deficit stress responses. Bot. Sci. 2016, 94, 657–671. [Google Scholar] [CrossRef]
- Blum, A. Osmotic adjustment is a prime drought stress adaptive engine in support of plant production. Plant Cell Environ. 2017, 40, 4–10. [Google Scholar] [CrossRef]
- Houmani, H.; Corpas, F.J. Differential responses to salt-induced oxidative stress in three phylogenetically related plant species: Arabidopsis thaliana (glycophyte), Thellungiella salsuginea and Cakile maritima (halophytes). Involvement of ROS and NO in the control of K+/Na+ homeostasis. AIMS Biophys. 2016, 3, 380–397. [Google Scholar] [CrossRef]
- Liu, N.; Jin, Z.; Wang, S.; Gong, B.; Wen, D.; Wang, X.; Wei, M.; Shi, Q. Sodic alkaline stress mitigation with exogenous melatonin involves reactive oxygen metabolism and ion homeostasis in tomato. Sci. Hortic. 2015, 181, 18–25. [Google Scholar] [CrossRef]
- Gil, R.; Lull, C.; Boscaiu, M.; Bautista, I.; Lidón, A.L.; Vicente, O. Soluble carbohydrates as osmolytes in several halophytes from a Mediterranean salt marsh. Not. Bot. Horti Agrobot. 2011, 39, 9–17. [Google Scholar] [CrossRef]
- Kishor, P.B.K.; Kumari, P.H.; Sunita, M.S.; Sreenivasulu, N. Role of proline in cell wall synthesis and plant development and its implications in plant ontogeny. Front. Plant. Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Kumar, J.; Singh, S.; Singh, V.P.; Prasad, S.M. Roles of osmoprotectants in improving salinity and drought tolerance in plants: A review. Rev. Environ. Sci. Biotechnol. 2015, 14, 407–426. [Google Scholar] [CrossRef]
- Miller, G.; Shulaev, V.; Mittler, R. Reactive oxygen signaling and abiotic stress. Physiol. Plant. 2008, 133, 481–489. [Google Scholar] [CrossRef]
- Podgórska, A.; Burian, M.; Szal, B. Extra-cellular but extra-ordinarily important for cells: Apoplastic reactive oxygen species metabolism. Front. Plant Sci. 2017, 8, 1353. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Szabados, L.; Savouré, A. Proline: A multifunctional amino acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tomar, N.S.; Tittal, M.; Argal, S.; Agarwal, R.M. Plant growth under water/salt stress: ROS production; antioxidants and significance of added potassium under such conditions. Physiol. Mol. Biol. Plants 2017, 23, 731–744. [Google Scholar] [CrossRef] [PubMed]
- Dietz, K.J. Thiol-based peroxidases and ascorbate peroxidases: Why plants rely on multiple peroxidase systems in the photosynthesizing chloroplast? Mol. Cells 2016, 39, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Nianiou-Obeidat, I.; Madesis, P.; Kissoudis, C.; Voulgari, G.; Chronopoulou, E.; Tsaftaris, A.; Labrou, N.E. Plant glutathione transferase-mediated stress tolerance: Functions and biotechnological applications. Plant Cell Rep. 2017, 36, 791–805. [Google Scholar] [CrossRef]
- Parađiković, N.; Vinković, T.; Vinković Vrček, I.; Žuntar, I.; Bojić, M.; Medić-Šarić, M. Effect of natural biostimulants on yield and nutritional quality: An example of sweet yellow pepper (Capsicum annuum L.) plants. J. Sci. Food Agric. 2011, 91, 2146–2152. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in plant science: A global perspective. Front. Plant. Sci. 2016, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Battacharyya, D.; Babgohari, M.Z.; Rathor, P.; Prithiviraj, B. Seaweed extracts as biostimulants in horticulture. Sci. Hortic. 2015, 196, 39–48. [Google Scholar] [CrossRef]
- Canellas, L.P.; Olivares, F.L.; Aguiar, N.O.; Jones, D.L.; Nebbioso, A.; Mazzei, P.; Piccolo, A. Humic and fulvic acids as biostimulants in horticulture. Sci. Hortic. 2015, 196, 15–27. [Google Scholar] [CrossRef]
- Colla, G.; Nardi, S.; Cardarelli, M.; Ertani, A.; Lucini, L.; Canaguier, R.; Rouphael, Y. Protein hydrolysates as biostimulants in horticulture. Sci. Hortic. 2015, 196, 28–38. [Google Scholar] [CrossRef]
- Escaich, J.; Soler, F.; Juncosa, R.; Gomis, P. Fructificación en cultivos tratados con aminoácidos de hidrólisis enzimática. Horticultura 1991, 67, 47–53. [Google Scholar]
- Mire, G.L.; Nguyen, M.L.; Fassotte, B.; Jardin, P.D.; Verheggen, F.J.; Delaplace, P.; Jijakli, M.H. Implementing plant biostimulants and biocontrol strategies in the agroecological management of cultivated ecosystems. A review. Biotechnol. Agron. Soc. Environ. 2016, 20, 299–313. [Google Scholar] [CrossRef]
- Aslam, M.; Travis, R.L.; Rains, D.W. Differential effect of amino acids on nitrate uptake and reduction systems in barley roots. Plant Sci. 2001, 160, 219–228. [Google Scholar] [CrossRef]
- Vasconcelos, A.; Zhang, X.; Ervin, E.; Kiehl, J. Enzymatic antioxidant responses to biostimulants in maize and soybean subjected to drought. Sci. Agric. 2009, 66, 395–402. [Google Scholar] [CrossRef]
- Torres-Rodríguez, J.A.; Reyes-Pérez, J.J.; González-Rodríguez, J.C. Effect of a natural biostimulant on some quality parameters of tomato seedlings (Solanum lycopersicum L.) under saline conditions. Biotecnia 2016, 18, 11–15. [Google Scholar] [CrossRef]
- Siwik-Ziomek, A.; Szczepanek, M. Soil enzyme activity and sulphur uptake by oilseed rape depending on fertilization and biostimulant application. Acta Agric. Scand. B Soil Plant Sci. 2018, 68, 50–56. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop production under drought and heat stress: Plant responses and management options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef]
- Maroto-Borrego, J.V. Horticultura Herbácea Especial, 5th ed.; Mundi-Prensa: Madrid, Spain, 2002; pp. 1–704. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Giannakoula, A.; Ilias, I. The effect of water stress and salinity on growth and physiology of tomato (Lycopersicon esculentum Mil.). Arch. Biol. Sci. 2013, 65, 611–620. [Google Scholar] [CrossRef]
- Sánchez-Rodríguez, E.; Moreno, D.A.; Ferreres, F.; Rubio-Wilhelmi, M.d.M.; Ruiz, J.M. Differential responses of five cherry tomato varieties to water stress: Changes on phenolic metabolites and related enzymes. Phytochemistry 2011, 72, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Zushi, K.; Matsuzoe, N. Seasonal and cultivar differences in salt-induced changes in antioxidant system in tomato. Sci. Hortic. 2009, 120, 181–187. [Google Scholar] [CrossRef]
- Wang, W.; Zheng, W.; Lv, H.; Liang, B.; Jin, S.; Li, J.; Zhou, W. Animal-derived plant biostimulant alleviates drought stress by regulating photosynthesis, osmotic adjustment, and antioxidant systems in tomato plants. Sci. Hortic. 2022, 305, 111365. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Agliassa, C.; Mannino, G.; Vigliante, I.; Contartese, V.; Secchi, F.; Bertea, C.M. A biostimulant based on seaweed (Ascophyllum Nodosum and Laminaria digitata) and yeast extracts mitigates water stress effects on tomato (Solanum lycopersicum L.). Agriculture 2021, 11, 557. [Google Scholar] [CrossRef]
- Peripolli, M.; Dornelles, S.; Lopes, S.; Tabaldi, L.; Trivisiol, V.; Rubert, J. Application of biostimulants in tomato subjected to water deficit: Physiological, enzymatic and production responses. Rev. Bras. Eng. Agrícola Ambient. 2021, 25, 90–95. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil, 2nd ed.; Circular 347; University of California Agricultural Experiment Station: Berkeley, CA, USA, 1950; pp. 1–39. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Vicente, O.; Boscaiu, M.; Naranjo, M.Á.; Estrelles, E.; Bellés, J.M.; Soriano, P. Responses to salt stress in the halophyte Plantago crassifolia (Plantaginaceae). J. Arid Environ. 2004, 58, 463–481. [Google Scholar] [CrossRef]
- Gil, R.; Bautista, I.; Boscaiu, M.; Lidón, A.; Wankhade, S.; Sánchez, H.; Llinares, J.; Vicente, O. Responses of five Mediterranean halophytes to seasonal changes in environmental conditions. AoB Plants 2014, 6, plu049. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell. Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Connell, J.P.; Mullet, J.E. Pea chloroplast glutathione reductase: Purification and characterization. Plant Physiol. 1986, 82, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Law, M.Y.; Charles, S.A.; Halliwell, B. Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of Paraquat. Biochem. J. 1983, 210, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef]
- Giordano, M.; Petropoulos, S.A.; Rouphael, Y. Response and defence mechanisms of vegetable crops against drought, heat and salinity stress. Agriculture 2021, 11, 463. [Google Scholar] [CrossRef]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed]
- Bodner, G.; Nakhforoosh, A.; Kaul, H.-P. Management of crop water under drought: A review. Agron. Sustain. Dev. 2015, 35, 401–442. [Google Scholar] [CrossRef]
- Pandey, P.; Irulappan, V.; Bagavathiannan, M.V.; Senthil-Kumar, M. Impact of combined abiotic and biotic stresses on plant growth and avenues for crop improvement by exploiting physio-morphological traits. Front. Plant Sci. 2017, 8, 537. [Google Scholar] [CrossRef]
- Petrozza, A.; Santaniello, A.; Summerer, S.; Di Tommaso, G.; Di Tommaso, D.; Paparelli, E.; Piaggesi, A.; Perata, P.; Cellini, F. Physiological responses to Megafol® treatments in tomato plants under drought stress: A phenomic and molecular approach. Sci. Hortic. 2014, 174, 185–192. [Google Scholar] [CrossRef]
- Chen, S.; Heuer, B. Effect of genotype and exogenous application of glycinebetaine on antioxidant enzyme activity in native gels of 7-day-old salt-stressed tomato (Solanum lycopersicum) seedlings. Sci. Hortic. 2013, 162, 106–116. [Google Scholar] [CrossRef]
- Claussen, W. Proline as a measure of stress in tomato plants. Plant Sci. 2005, 168, 241–248. [Google Scholar] [CrossRef]
- Goel, D.; Singh, A.K.; Yadav, V.; Babbar, S.B.; Murata, N.; Bansal, K.C. Transformation of tomato with a bacterial codA gene enhances tolerance to salt and water stresses. J. Plant Physiol. 2011, 168, 1286–1294. [Google Scholar] [CrossRef]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Barone, A.; Rigano, M.M. A novel protein hydrolysate-based biostimulant improves tomato performances under drought stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef] [PubMed]
- Casadesús, A.; Polo, J.; Munné-Bosch, S. Hormonal effects of an enzymatically hydrolyzed animal protein-based biostimulant (pepton) in water-stressed tomato plants. Front. Plant Sci. 2019, 10, 758. [Google Scholar] [CrossRef]
- Ahmed, M.; Ullah, H.; Attia, A.; Tisarum, R.; Cha-um, S.; Datta, A. Interactive effects of Ascophyllum nodosum seaweed extract and silicon on growth, fruit yield and quality, and water productivity of tomato under water stress. Silicon 2023, 15, 2263–2278. [Google Scholar] [CrossRef]
- Domingo, G.; Marsoni, M.; Álvarez-Viñas, M.; Torres, M.D.; Domínguez, H.; Vannini, C. The role of protein-rich extracts from Chondrus crispus as biostimulant and in enhancing tolerance to drought stress in tomato plants. Plants 2023, 12, 845. [Google Scholar] [CrossRef]
- Qin, K.; Leskovar, D.I. Humic substances improve vegetable seedling quality and post-transplant yield performance under stress conditions. Agriculture 2020, 10, 254. [Google Scholar] [CrossRef]
- Turan, M.; Yildirim, E.; Ekinci, M.; Argin, S. Effect of biostimulants on yield and quality of cherry tomatoes grown in fertile and stressed soils. HortScience 2021, 56, 414–423. [Google Scholar] [CrossRef]
- Xu, X.; Lei, X.; Liao, S.; Li, Y.; Sun, Y. Foliar application of potassium silicate, potassium fulvate and betaine improve summer-time tomato yield by promoting plant nitrogen and potassium uptake. Folia Hortic. 2022, 34, 125–138. [Google Scholar] [CrossRef]
- Top, S.; Vandoorne, B.; Pauwels, E.; Perneel, M.; Van Labeke, M.C.; Steppe, K. Plant sensors untangle the water-use and growth effects of selected seaweed-derived biostimulants on drought-stressed tomato plants (Solanum lycopersicum). J. Plant Growth Regul. 2023, 42, 5615–5627. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G.; Giordano, M.; El-Nakhel, C.; Kyriacou, M.C.; De Pascale, S. Foliar applications of a legume-derived protein hydrolysate elicit dose-dependent increases of growth, leaf mineral composition, yield and fruit quality in two greenhouse tomato cultivars. Sci. Hortic. 2017, 226, 353–360. [Google Scholar] [CrossRef]
- Vaghela, P.; Trivedi, K.; Anand, K.G.V.; Brahmbhatt, H.; Nayak, J.; Khandhediya, K.; Prasad, K.; Moradiya, K.; Kubavat, D.; Konwar, L.J.; et al. Scientific basis for the use of minimally processed homogenates of Kappaphycus alvarezii (red) and Sargassum wightii (brown) seaweeds as crop biostimulants. Algal Res. 2023, 70, 102969. [Google Scholar] [CrossRef]
- Villa e Vila, V.; Marques, P.A.A.; Rezende, R.; Wenneck, G.S.; Terassi, D.D.S.; Andrean, A.F.B.A.; Nocchi, R.C.D.F.; Matumoto-Pintro, P.T. Deficit Irrigation with Ascophyllum nodosum Extract Application as a Strategy to Increase Tomato Yield and Quality. Agronomy 2023, 13, 1853. [Google Scholar] [CrossRef]
- Koleška, I.; Hasanagić, D.; Todorović, V.; Murtić, S.; Klokić, I.; Parađiković, N.; Kukavica, B. Biostimulant prevents yield loss and reduces oxidative damage in tomato plants grown on reduced NPK nutrition. J. Plant Interact. 2017, 12, 209–218. [Google Scholar] [CrossRef]
- Rady, M.M.; Rehman, H.u. Supplementing organic biostimulants into growing media enhances growth and nutrient uptake of tomato transplants. Sci. Hortic. 2016, 203, 192–198. [Google Scholar] [CrossRef]
- Zodape, S.T.; Gupta, A.R.; Bhandari, S.; Rawat, U.S.; Chaudhary, D.; Eswaran, K.; Chikara, J. Foliar application of seaweed sap as biostimulant for enhancement of yield and quality of tomato (Lycopersicon esculentum Mill.). J. Sci. Ind. Res. 2011, 70, 215–219. [Google Scholar]
- Vinoth, S.; Gurusaravanan, P.; Jayabalan, N. Effect of seaweed extracts and plant growth regulators on high-frequency in vitro mass propagation of Lycopersicon esculentum L (tomato) through double cotyledonary nodal explant. J. Appl. Phycol. 2012, 24, 1329–1337. [Google Scholar] [CrossRef]
- Kalozoumis, P.; Vourdas, C.; Ntatsi, G.; Savvas, D. Can biostimulants increase resilience of hydroponically-grown tomato to combined water and nutrient stress? Horticulturae 2021, 7, 297. [Google Scholar] [CrossRef]
- Cristofano, F.; El-Nakhel, C.; Rouphael, Y. Biostimulant substances for sustainable agriculture: Origin, operating mechanisms and effects on cucurbits, leafy greens, and nightshade vegetables species. Biomolecules 2021, 11, 1103. [Google Scholar] [CrossRef]
- Ali, S.; Rab, A.; Khan, D.N.; Nawab, K. Enhanced proline synthesis may determine resistance to salt stress in tomato cultivars. Pak. J. Bot. 2011, 43, 2707–2710. [Google Scholar]
- Amini, F.; Ehsanpour, A. Soluble Proteins, Proline, Carbohydrates and Na+/K+ Changes in two tomato (Lycopersicon esculentum Mill.) cultivars under in vitro salt stress. Am. J. Biochem. Biotechnol. 2005, 1, 212–216. [Google Scholar] [CrossRef]
- Machado, J.; Vasconcelos, M.W.; Soares, C.; Fidalgo, F.; Heuvelink, E.; Carvalho, S.M.P. Young tomato plants respond differently under single or combined mild nitrogen and water deficit: An insight into morphophysiological responses and primary metabolism. Plants 2023, 12, 1181. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, S.; Al Hassan, M.; Bandara, W.M.C.; Yabor, L.; Llinares, J.V.; Boscaiu, M.; Vicente, O. Screening for salt tolerance in four local varieties of Phaseolus lunatus from Spain. Agriculture 2018, 8, 201. [Google Scholar] [CrossRef]
- Plazas, M.; Nguyen, H.T.; González-Orenga, S.; Fita, A.; Vicente, O.; Prohens, J.; Boscaiu, M. Comparative analysis of the responses to water stress in eggplant (Solanum melongena) cultivars. Plant Physiol. Biochem. 2019, 143, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Arteaga, S.; Yabor, L.; Díez, M.J.; Prohens, J.; Boscaiu, M.; Vicente, O. The use of proline in screening for tolerance to drought and salinity in common bean (Phaseolus vulgaris L.) genotypes. Agronomy 2020, 10, 817. [Google Scholar] [CrossRef]
- Goñi, O.; Quille, P.; O'Connell, S. Ascophyllum nodosum extract biostimulants and their role in enhancing tolerance to drought stress in tomato plants. Plant Physiol. Biochem. 2018, 126, 63–73. [Google Scholar] [CrossRef]
- Ali, O.; Ramsubhag, A.; Jayaraman, J. Biostimulant properties of seaweed extracts in plants: Implications towards sustainable crop production. Plants 2021, 10, 531. [Google Scholar] [CrossRef]
- González-Orenga, S.; Al Hassan, M.; Llinares, J.V.; Lisón, P.; López-Gresa, M.P.; Verdeguer, M.; Vicente, O.; Boscaiu, M. Qualitative and quantitative differences in osmolytes accumulation and antioxidant activities in response to water deficit in four Mediterranean Limonium species. Plants 2019, 8, 506. [Google Scholar] [CrossRef] [PubMed]
- Zuzunaga-Rosas, J.; González-Orenga, S.; Calone, R.; Rodríguez-Heredia, R.; Asaff-Torres, A.; Boscaiu, M.; Ibáñez-Asensio, S.; Moreno-Ramón, H.; Vicente, O. Use of a biostimulant to mitigate the effects of excess salinity in soil and irrigation water in tomato plants. Plants 2023, 12, 1190. [Google Scholar] [CrossRef]
- Liu, R.; Jiang, X.; Guan, H.; Li, X.; Du, Y.; Wang, P.; Mou, H. Promotive effects of alginate-derived oligosaccharides on the inducing drought resistance of tomato. J. Ocean. Univ. China 2009, 8, 303–311. [Google Scholar] [CrossRef]
- Zhang, X.; Ervin, E.; Schmidt, R. Physiological effects of liquid applications of a seaweed extract and a humic acid on creeping bentgrass. J. Am. Soc. Hortic. Sci. 2003, 128, 492–496. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Free Amino Acids | % (w/v) | Free Amino Acids | % (w/v) |
|---|---|---|---|
| Aspartic acid | 0.26 | Isoleucine | 0.09 |
| Glutamic acid | 2.68 | Leucine | 0.13 |
| Alanine | 0.28 | Lysine | 0.67 |
| Arginine | 0.56 | Methionine | 0.07 |
| Cysteine | 0.1 | Serine | 0.2 |
| Phenylalanine | 0.1 | Tyrosine | 0.11 |
| Glycine | 1.21 | Threonine | 0.1 |
| Histidine | 0.16 | Valine | 0.09 |
| Treatment | Stem Length (cm) | Leaf Number | Flower Number | Leaf Area (cm2) |
|---|---|---|---|---|
| IC | 43 ± 1.2 f | 20 ± 0.7 d | 33.8 ± 5.3 c | 1004.8 ± 45.6 c |
| NIC | 30 ± 1 abc | 14.2 ± 0.5 bc | 9.4 ± 1.5 a | 768.4 ± 116.2 ab |
| TB0.2 | 33.3 ± 2.3 e | 14 ± 0.7 abc | 9.2 ± 3.6 a | 681.1 ± 185.9 ab |
| TB1 | 32.2 ± 1.6 cde | 14 ± 0.7 abc | 8.4 ± 2.6 a | 640.4 ± 83.3 a |
| KF0.2 | 32.6 ± 2.4 de | 13.8 ± 0.5 abc | 8.8 ± 4.2 a | 831.8 ± 190.4 b |
| KF1 | 31.6 ± 2.2 bcde | 13.6 ± 1.1 abc | 7.2 ± 4.7 a | 714.4 ± 182.4 ab |
| HA0.2 | 30.2 ± 1.3 bcd | 13.4 ± 0.6 ab | 11.4 ± 2.7 ab | 719.6 ± 69.3 ab |
| HA1 | 29.6 ± 1.8 ab | 13.2 ± 0.5 a | 15.6 ± 8.1 b | 686.6 ± 64.4 ab |
| SE0.2 | 27.6 ± 3.5 a | 14.2 ± 0.8 bc | 9.6 ± 2.1 a | 641.6 ± 73.6 a |
| SE1 | 30.4 ± 1.1 bcd | 14.4 ± 0.6 c | 11.2 ± 2.3 ab | 686 ± 67.1 ab |
| Treatment | TPC (mg eq. GA g−1 DW) | TF (mg eq. C g−1 DW) | Ascorbic Acid (mg g−1 DW) |
|---|---|---|---|
| IC | 15.7 ± 2.9 de | 12.6 ± 2.1 f | 0.71 ± 0.16 a |
| NIC | 16.4 ± 0.7 e | 7.4 ± 2.2 bcde | 1.23 ± 0.18 e |
| TB0.2 | 13.8 ± 0.8 abc | 8.0 ± 2 de | 0.80 ± 0.26 abc |
| TB1 | 13.4 ± 0.8 ab | 5.9 ± 0.7 abc | 0.69 ± 0.14 ab |
| KF0.2 | 12.7 ± 0.7 a | 7.6 ± 1.7 cde | 0.80 ± 0.17 cd |
| KF1 | 14.8 ± 1.1 bcde | 9.0 ± 1.4 e | 0.85 ± 0.13 bc |
| HA0.2 | 14.4 ± 1.8 abcd | 5.0 ± 1.6 a | 1.29 ± 0.16 e |
| HA1 | 15.5 ± 1.3 cde | 5.3 ± 1.6 bc | 1.55 ± 0.12 e |
| SE0.2 | 13.7 ± 0.6 abc | 5.5 ± 0.7 ab | 1.01 ± 0.16 cd |
| SE1 | 14.4 ± 1.7 abcd | 6.2 ± 1 abcd | 1.24 ± 0.31 e |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Ortiz, R.; Naranjo, M.Á.; Atares, S.; Vicente, O. Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants. Agronomy 2023, 13, 2314. https://doi.org/10.3390/agronomy13092314
Gil-Ortiz R, Naranjo MÁ, Atares S, Vicente O. Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants. Agronomy. 2023; 13(9):2314. https://doi.org/10.3390/agronomy13092314
Chicago/Turabian StyleGil-Ortiz, Ricardo, Miguel Ángel Naranjo, Sergio Atares, and Oscar Vicente. 2023. "Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants" Agronomy 13, no. 9: 2314. https://doi.org/10.3390/agronomy13092314
APA StyleGil-Ortiz, R., Naranjo, M. Á., Atares, S., & Vicente, O. (2023). Antioxidant Responses of Water-Stressed Cherry Tomato Plants to Natural Biostimulants. Agronomy, 13(9), 2314. https://doi.org/10.3390/agronomy13092314