


Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Experimental Conditions
2.3. Data Collection
2.4. High-Performance Liquid Chromatography Analysis (HPLC) of Phenolic Compounds
2.5. Statistical Analysis
3. Results and Discussion
3.1. Adventitious Bulblet and Root Formation
3.2. Identification of Phenolic Acids in the Bulblets
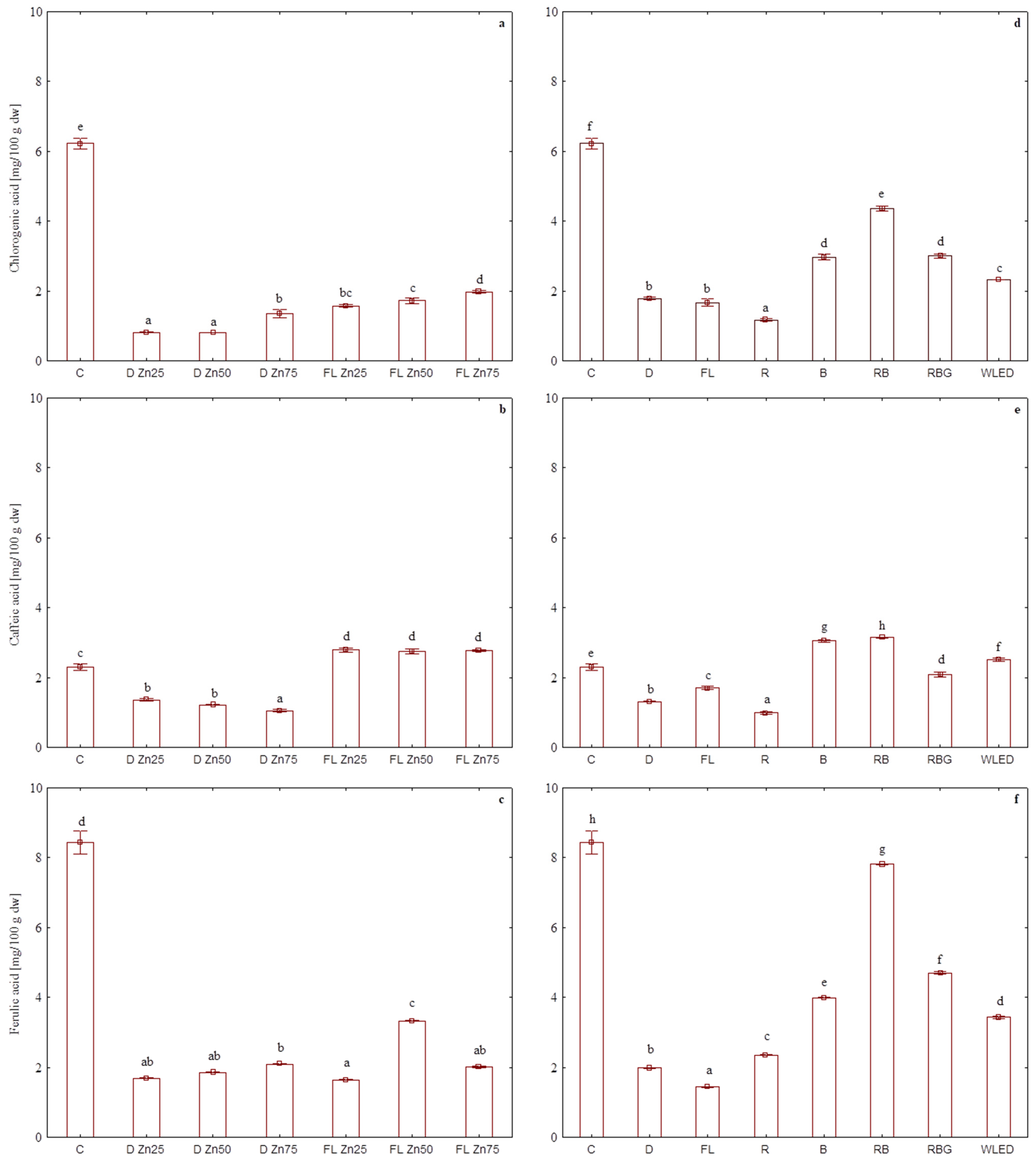
3.3. Effect of Zinc Oxide Nanoparticles on Phenolic Acid Content
3.4. Effect of LED Light on Phenolic Acid Content
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Özen, F.; Temeltaş, H.; Aksoy, Ö. The anatomy and morphology of the medicinal plant, Lilium candidum L. (Liliaceae) distributed in Marmara region of Turkey. Pak. J. Bot. 2012, 44, 1185–1192. [Google Scholar]
- Pieroni, A. Medicinal plants and food medicines in the folk traditions of the upper Lucca Province, Italy. J. Ethnopharmacol. 2000, 70, 235–273. [Google Scholar] [CrossRef] [PubMed]
- Patocka, J.; Navratilova, Z. Bioactivity of Lilium candidum L.: A mini review. Biomed. J. Sci. Technol. Res. 2019, 8, 13859–13862. [Google Scholar] [CrossRef]
- Zaccai, M.; Yarmolinsky, L.; Khalfin, B.; Budovsky, A.; Gorelick, J.; Dahan, A.; Ben-Shabat, S. Medicinal properties of Lilium candidum L. and its phytochemicals. Plants 2020, 9, 959. [Google Scholar] [CrossRef]
- Momtaz, S.; Dibaj, M.; Abdollahi, A.; Amin, G.; Bahramsoltani, R.; Abdollahi, M.; Mahdaviani, P.; Abdolghaffari, A.H. Wound healing activity of the flowers of Lilium candidum L. in burn wound model in rats. J. Med. Plants 2020, 19, 109–118. [Google Scholar] [CrossRef]
- Pałka, P.; Cioć, M.; Hura, K.; Szewczyk-Taranek, B.; Pawłowska, B. Adventitious organogenesis and phytochemical composition of Madonna lily (Lilium candidum L.) in vitro modeled by different light quality. Plant Cell Tissue Organ Cult. (PCTOC) 2023, 152, 99–114. [Google Scholar] [CrossRef]
- Khawar, K.M.; Cocu, S.; Parmaksiz, I.; Sarihan, E.O.; Özcan, S. Mass proliferation of Madonna Lily (Lilium candidum L.) under in vitro conditions. Pak. J. Bot. 2005, 37, 243–248. [Google Scholar]
- Sevimay, C.S.; Khawar, K.M.; Parmaksız, I.; Cocu, S.; Sancak, C.; Sarihan, E.; Özcan, S. Prolific in vitro bulblet formation from bulb scales of meadow lily (Lilium candidum L.). Period. Biol. 2005, 107, 107–111. [Google Scholar]
- Altan, F.; Bürün, B.; Şahin, N. Fungal contaminants observed during micropropagation of Lilium candidum L. and the effect of chemotherapeutic substances applied after sterilization. Afr. J. Biotechnol. 2010, 9, 991–995. [Google Scholar] [CrossRef]
- Burun, B.; Sahin, O. Micropropagation of Lilium candidum L.: A rare and native bulbous flower of Turkey. Bangladesh J. Bot. 2013, 42, 185–187. [Google Scholar] [CrossRef]
- Saadon, S.; Zaccai, M. Lilium candidum bulblet and meristem development. Vitr. Cell. Dev. Biol. Plant 2013, 49, 313–319. [Google Scholar] [CrossRef]
- Altan, F.; Bürün, B. The effect of some antibiotic and fungucide applications on the micropropagation of Lilium candidum L. Mugla J. Sci. Technol. 2017, 3, 86–91. [Google Scholar] [CrossRef]
- Daneshvar Royandazagh, S. Efficient approaches to in vitro multiplication of Lilium candidum L. with consistent and safe access throughout year and acclimatization of plant under hot-summer mediterranean (Csa Type) climate. Not. Bot. Horti Agrobot. 2019, 47, 734–742. [Google Scholar] [CrossRef]
- Tokgoz, H.B.; Altan, F. Callus induction and micropropagation of Lilium candidum L. using stem bulbils and confirmation of genetic stability via SSR-PCR. Int. J. Second. Metab. 2020, 7, 286–296. [Google Scholar] [CrossRef]
- Akshay, M.P.; Pooja, P.G.; Sonali, D. In vitro micropropagation of Lilium candidum bulb by application of multiple hormone concentrations using plant tissue culture technique. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 244–253. [Google Scholar] [CrossRef]
- Bornwaßer, T.; Tantau, H.J. Evaluation of LED lighting systems in in vitro cultures. Acta Hort. 2012, 956, 555–562. [Google Scholar] [CrossRef]
- Gupta, S.D.; Jatothu, B. Fundamentals and applications of lightemitting diodes (LEDs) in in vitro plant growth and morphogenesis. Plant Biotechnol. Rep. 2013, 7, 211–220. [Google Scholar] [CrossRef]
- Bantis, F.; Smirnakou, S.; Ouzounis, T.; Koukounaras, A.; Ntagkas, N.; Radoglou, K. Current status and recent achievements in the field of horticulture with the use of light-emitting diodes (LEDs). Sci. Hortic. 2018, 235, 437–451. [Google Scholar] [CrossRef]
- Lian, M.L.; Murthy, H.N.; Paek, K.Y. Effects of light emitting diodes (LED) on the in vitro induction and growth of bulblets of Lilium oriental hybrid ‘Pesaro’. Sci. Hortic. 2002, 94, 365–370. [Google Scholar] [CrossRef]
- Prokopiuk, B.; Cioć, M.; Maślanka, M.; Pawłowska, B. Effects of light spectra and benzyl adenine on in vitro adventitious bulb and shoot formation of Lilium regale E. H. Wilson. Propag. Ornam. Plants 2018, 18, 12–18. [Google Scholar]
- Rivero-Montejo, S.J.; Vargas-Hernandez, M.; Torres-Pacheco, I. Nanoparticles as novel elicitors to improve bioactive compounds in plants. Agriculture 2021, 11, 134. [Google Scholar] [CrossRef]
- Namdeo, A.G. Plant cell elicitation for production of secondary metabolites: A review. Pharmacogn. Rev. 2007, 1, 69–79. [Google Scholar]
- Shitan, N. Secondary metabolites in plants: Transport and self-tolerance mechanisms. Biosci. Biotechnol. Biochem. 2016, 80, 1283–1293. [Google Scholar] [CrossRef] [PubMed]
- Jafari, S.M.; McClements, D.J. Chapter One—Nanotechnology approaches for increasing nutrient bioavailability. In Advances in Food and Nutrition Research, 1st ed.; Toldrá, F., Ed.; Academic Press: London, UK, 2017; Volume 81, pp. 1–30. [Google Scholar] [CrossRef]
- Iranbakhsh, A.; Oraghi Ardebili, Z.; Oraghi Ardebili, N. Synthesis and characterization of zinc oxide nanoparticles and their impact on plants. In Plant Responses to Nanomaterials; Singh, V.P., Singh, S., Tripathi, D.K., Prasad, S.M., Chauhan, D.K., Eds.; Nanotechnology in the Life Sciences; Springer: Cham, Switzerland, 2021; pp. 33–93. [Google Scholar] [CrossRef]
- Misra, A.; Srivastava, A.K.; Srivastava, N.K.; Khan, A. Zn-acquisition and its role in growth, photosynthesis, photosynthetic pigments, and biochemical changes in essential monoterpene oil(s) of Pelargonium graveolens. Photosynthetica 2005, 43, 153–155. [Google Scholar] [CrossRef]
- Eisvand, H.R.; Kamaei, H.; Nazarian, F. Chlorophyll fluorescence, yield and yield components of bread wheat affected by phosphatebio-fertilizer, zinc and boron under late-season heat stress. Photosynthetica 2018, 56, 1287–1296. [Google Scholar] [CrossRef]
- Raliya, R.; Tarafdar, J.C. ZnO nanoparticle biosynthesis and its effect on phosphorus-mobilizing enzyme secretion and gum contents in clusterbean (Cyamopsis tetragonoloba L.). Agric. Res. 2013, 2, 48–57. [Google Scholar] [CrossRef]
- Sabir, S.; Arshad, M.; Chaudhari, S.K. Zinc oxide nanoparticles for revolutionizing agriculture: Synthesis and applications. Sci. World J. 2014, 2014, 925494. [Google Scholar] [CrossRef]
- Singh, A.; Singh, N.B.; Afzal, S.; Singh, T.; Hussain, I. Zinc oxide nanoparticles: A review of their biological synthesis, antimicrobial activity, uptake, translocation and biotransformation in plants. J. Mater. Sci. 2018, 53, 185–201. [Google Scholar] [CrossRef]
- Wojnarowicz, J.; Chudoba, T.; Lojkowski, W. A Review of Microwave Synthesis of Zinc Oxide Nanomaterials: Reactants, Process Parameters and Morphologies. Nanomaterials 2020, 10, 1086. [Google Scholar] [CrossRef]
- Prasad, T.N.V.K.V.; Sudhakar, P.; Sreenivasulu, Y.; Latha, P.; Munaswamy, V.; Raja Reddy, K.; Sreeprasad, T.S.; Sajanlal, P.R.; Pradeep, T. Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 2012, 35, 905–927. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Metwally, M.A.; El-Hoseiny, H.; Omar, S.A.; El-Sheery, N.I. Effect of nanoparticles on biological contamination of in vitro cultures and organogenic regeneration of banana. Aust. J. Crop Sci. 2014, 8, 612–624. [Google Scholar]
- Raskar, S.V.; Laware, S.L. Effect of zinc oxide nanoparicles on cytology and seed germination in onion. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 467–473. [Google Scholar]
- Chamani, E.; Ghalehtaki, S.K.; Mohebodini, M.; Ghanbari, A. The effect of zinc oxide nano particles and humic acid on morphological characters and secondary metabolite production in Lilium ledebourii Bioss. Iran. J. Genet. Plant Breed. 2015, 4, 11–19. [Google Scholar]
- Mosavat, N.; Golkar, P.; Yousefifard, M.; Javed, R. Modulation of callus growth and secondary metabolites in different Thymus species and Zataria multiflora micropropagated under ZnO nanoparticles stress. Biotechnol. Appl. Biochem. 2019, 66, 316–322. [Google Scholar] [CrossRef]
- Ahmad, M.A.; Javed, R.; Adeel, M.; Rizwan, M.; Ao, Q.; Yang, Y. Engineered ZnO and CuO nanoparticles ameliorate morphological and biochemical response in tissue culture regenerants of candyleaf (Stevia rebaudiana). Molecules 2020, 25, 1356. [Google Scholar] [CrossRef] [PubMed]
- El-Mahdy, M.T.; Elazab, D. Impact of zinc oxide nanoparticles on pomegranate growth under in vitro conditions. Russ. J. Plant Physiol. 2020, 67, 162–167. [Google Scholar] [CrossRef]
- Mazaheri-Tirani, M.; Dayani, S. In vitro effect of zinc oxide nanoparticles on Nicotiana tabacum callus compared to ZnO micro particles and zinc sulfate (ZnSO4). Plant Cell Tissue Organ Cult. (PCTOC) 2020, 140, 279–289. [Google Scholar] [CrossRef]
- Tymoszuk, A.; Wojnarowicz, J. Zinc oxide and zinc oxide nanoparticles impact on in vitro germination and seedling growth in Allium cepa L. Materials 2020, 13, 2784. [Google Scholar] [CrossRef]
- Zaeem, A.; Drouet, S.; Anjum, S.; Khurshid, R.; Younas, M.; Blondeau, J.P.; Tungmunnithum, D.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Effects of biogenic zinc oxide nanoparticles on growth and oxidative stress response in flax seedlings vs. in vitro cultures: A comparative analysis. Biomolecules 2020, 10, 918. [Google Scholar] [CrossRef] [PubMed]
- Al-Mayahi, A.M.W. The effect of humic acid (HA) and zinc oxide nanoparticles (ZnO-NPS) on in vitro regeneration of date palm (Phoenix dactylifera L.) cv. Quntar. Plant Cell Tissue Organ Cult. 2021, 145, 445–456. [Google Scholar] [CrossRef]
- Hezaveh, T.A.; Rahmani, F.; Alipour, H.; Pourakbar, L. Effects of foliar application of ZnO nanoparticles on secondary metabolite and micro-elements of camelina (Camelina sativa L.) under salinity stress. J. Stress Physiol. Biochem. 2020, 16, 54–69. [Google Scholar]
- Sharifi-Rad, R.; Bahabadi, S.E.; Samzadeh-Kermani, A.; Gholami, M. The effect of non-biological elicitors on physiological and biochemical properties of medicinal plant Momordica charantia L. Iran. J. Sci. Technol. Trans. A Sci. 2020, 44, 1315–1326. [Google Scholar] [CrossRef]
- Rosa, L.A.; Moreno-Escamilla, J.O.; Rodrigo-García, J.; Alvarez-Parrilla1, E. Chapter 12 Phenolic compounds. In Postharvest Physiology and Biochemistry of Fruits and Vegetables; Yahia, E.M., Ed.; Woodhead Publishing: Sawton, UK, 2018; pp. 253–271. [Google Scholar] [CrossRef]
- Babenko, L.M.; Smirnov, O.E.; Romanenko, K.O.; Trunova, O.K.; Kosakivska, I.V. Phenolic compounds in plants: Biogenesis and functions. Ukr. Biochem. J. 2019, 91, 5–18. [Google Scholar] [CrossRef]
- Ghasemzadeh, A.; Jaafar, H.Z.E.; Rahmat, A.; Wahab, P.E.M.; Halim, M.R.A. Effect of different light intensities on total phenolics and flavonoids synthesis and anti-oxidant activities in young ginger varieties (Zingiber officinale Roscoe). Int. J. Mol. Sci. 2010, 11, 3885–3897. [Google Scholar] [CrossRef]
- Chang, H.P.; Kim, N.S.; Park, J.S.; Lee, S.Y.; Lee, J.W.; Park, S.U. Effects of light-emitting diodes on the accumulation of glucosinolates and phenolic compounds in sprouting canola (Brassica napus L.). Foods 2019, 8, 76. [Google Scholar] [CrossRef]
- Hsie, B.S.; Bueno, A.I.S.; Bertolucci, S.K.V.; Carvalho, A.A.; Cunha, S.H.B.; Martins, E.R.; Pinto, J.E.B.P. Study of the influence of wavelengths and intensities of LEDs on the growth, photosynthetic pigment, and volatile compounds production of Lippia rotundifolia Cham in vitro. J. Photochem. Photobiol. B Biol. 2019, 198, 111577. [Google Scholar] [CrossRef]
- Upadhyay, R.; Rao, L.J.M. An outlook on chlorogenic acids—Occurrence, chemistry, technology, and biological activities. Crit. Rev. Food Sci. Nutr. 2013, 53, 968–984. [Google Scholar] [CrossRef]
- Marques, V.; Farah, A. Chlorogenic acids and related compounds in medicinal plants and infusions. Food Chem. 2009, 113, 1370–1376. [Google Scholar] [CrossRef]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; Xia, F.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Martínez, G.; Regente, M.; Jacobi, S.; Del Rio, M.; Pinedo, M.; de la Canal, L. Chlorogenic acid is a fungicide active against phytopathogenic fungi. Pestic. Biochem. Physiol. 2017, 140, 30–35. [Google Scholar] [CrossRef]
- Kundu, A.; Vadassery, J. Chlorogenic acid-mediated chemical defence of plants against insect herbivores. Plant Biol. 2019, 21, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Magnani, C.; Isaac, V.L.B.; Correa, M.A.; Salgado, H.R.N. Caffeic Acid: A review of its potential use for medications and cosmetics. Anal. Methods 2014, 6, 3203–3210. [Google Scholar] [CrossRef]
- Greenwald, P. Clinical trials in cancer prevention: Current results and perspectives for the future. J. Nutr. 2004, 134, 3507S–3512S. [Google Scholar] [CrossRef] [PubMed]
- Bouzaiene, N.N.; Jaziri, S.K.; Kovacic, H.; Chekir-Ghedira, L.; Ghedira, K.; Luis, J. The effects of caffeic, coumaric and ferulic acids on proliferation, superoxide production, adhesion and migration of human tumor cells in vitro. Eur. J. Pharmacol. 2015, 766, 99–105. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Jimenez-Escrig, A.; Saura-Calixto, F. Study of low-density lipoprotein oxidizability indexes to measure the antioxidant activity of dietary polyphenol. Nutr. Res. 2000, 20, 941–953. [Google Scholar] [CrossRef]
- Vinson, J.A.; Teufel, K.; Wu, N. Red wine, dealcoholized red wine, and especially grape juice, inhibit atherosclerosis in a hamster model. Atherosclerosis 2001, 156, 67–72. [Google Scholar] [CrossRef]
- Pei, K.; Ou, J.; Huang, J.; Ou, S. p-Coumaric acid and its conjugates: Dietary sources, pharmacokinetic properties and biological activities. J. Sci. Food Agric. 2016, 96, 2952–2962. [Google Scholar] [CrossRef]
- Zang, L.Y.; Cosma, G.; Gardner, H.; Shi, X.; Castranova, V.; Vallyathan, V. Effect of antioxidant protection by p-coumaric acid on low-density lipoprotein cholesterol oxidation. Am. J. Physiol. Cell Physiol. 2000, 279, C954–C960. [Google Scholar] [CrossRef]
- Kiliç, I.; Yeşiloğlu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Lou, Z.; Wang, H.; Rao, S.; Sun, J.; Ma, C.; Li, J. p-Coumaric acid kills bacteria through dual damage mechanisms. Food Control 2012, 25, 550–554. [Google Scholar] [CrossRef]
- Boz, H. p-Coumaric acid in cereals: Presence, antioxidant and antimicrobial effects. Int. J. Food Sci. Technol. 2015, 50, 2323–2328. [Google Scholar] [CrossRef]
- Janicke, B.; Hegardt, C.; Krogh, M.; Onning, G.; Akesson, B.; Cirenajwis, H.M.; Oredsson, S.M. The antiproliferative effect of dietary fiber phenolic compounds ferulic acid and p-coumaric acid on the cell cycle of Caco-2 cells. Nutr. Cancer. 2011, 63, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K.; Supriyanto, E.; Mandal, M. Events associated with apoptotic effect of p-Coumaric acid in HCT-15 colon cancer cells. World J. Gastroenterol. 2013, 19, 7726–7734. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.; Narayanankutty, A.; Nazeem, P.A.; Valsalan, R.; Babu, T.D.; Mathew, D. Plant phenolics ferulic acid and p-coumaric acid inhibit colorectal cancer cell proliferation through EGFR down-regulation. Asian Pac. J. Cancer Prev. 2016, 17, 4019–4023. [Google Scholar] [PubMed]
- Contardi, M.; Alfaro-Pulido, A.; Picone, P.; Guzman-Puyol, S.; Goldoni, L.; Benítez, J.J.; Heredia, A.; Barthel, M.J.; Ceseracciu, L.; Cusimano, G.; et al. Low molecular weight epsilon-caprolactone-p-coumaric acid copolymers as potential biomaterials for skin regeneration applications. PLoS ONE 2019, 14, e0214956. [Google Scholar] [CrossRef]
- Contardi, M.; Heredia-Guerrero, J.A.; Guzman-Puyol, S.; Summa, M.; Benítez, J.J.; Goldoni, L.; Caputo, G.; Cusimano, G.; Picone, P.; Di Carlo, M.; et al. Combining dietary phenolic antioxidants with polyvinylpyrrolidone: Transparent biopolymer films based on p-coumaric acid for controlled release. J. Mater. Chem. B 2019, 7, 1384–1396. [Google Scholar] [CrossRef]
- Boo, Y.C. p-Coumaric acid as an active ingredient in cosmetics: A review focusing on its antimelanogenic effects. Antioxidants 2019, 8, 275. [Google Scholar] [CrossRef]
- Ou, S.; Kwok, K. Ferulic acid: Pharmaceutical functions, preparation and applications in foods. J. Sci. Food Agric. 2004, 84, 1261–1269. [Google Scholar] [CrossRef]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic acid: Therapeutic potential through its antioxidant property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef]
- Li, D.; Rui, Y.; Guo, S.; Luan, F.; Liu, R.; Zeng, N. Ferulic acid: A review of its pharmacology, pharmacokinetics and derivatives. Life Sci. 2021, 284, 119921. [Google Scholar] [CrossRef]
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant properties of ferulic acid and its possible application. Skin Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Sakai, S.; Ochiai, H.; Nakajima, K.; Terasawa, K. Inhibitory effect of ferulic acid on macrophage inflammatory protein-2 production in a murine macrophage cell line, RAW264.7. Cytokine 1997, 9, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Ou, L.; Kong, L.; Zhang, X.; Niwa, M. Oxidation of ferulic acid by Momordica charantia peroxidase and related anti-inflammation activity changes. Biol. Pharm. Bull. 2003, 26, 1511–1516. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Yamamoto, T.; Hara, A.; Shimizu, M.; Yamada, Y.; Matsunaga, K.; Tanaka, T.; Mori, H. Modifying effects of ferulic acid on azoxymethane-induced colon carcinogenesis in F344 rats. Cancer Lett. 2000, 157, 15–21. [Google Scholar] [CrossRef]
- Balasubashini, M.S.; Rukkumani, R.; Viswanathan, P.; Menon, V.P. Ferulic acid alleviates lipid peroxidation in diabetic rats. Phytother. Res. 2004, 18, 310–314. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Pawłowska, B.; Żupnik, M.; Szewczyk-Taranek, B.; Cioć, M. Impact of LED light sources on morphogenesis and levels of photosynthetic pigments in Gerbera jamesonii grown in vitro. Hortic. Environ. Biotechnol. 2018, 59, 115–123. [Google Scholar] [CrossRef]
- Li, H.; Xu, Z.; Tang, C. Effect of light-emitting diodes on growth and morphogenesis of upland cotton (Gossypium hirsutum L.) plantlets in vitro. Plant Cell Tissue Organ Cult. 2010, 103, 155–163. [Google Scholar] [CrossRef]
- Fan, X.X.; Zang, J.; Xu, Z.G.; Guo, S.R.; Jiao, X.L.; Liu, X.Y.; Gao, Y. Effects of different light quality on growth, chlorophyll concentration and chlorophyll biosynthesis precursors of non-heading Chinese cabbage (Brassica campestris L.). Acta Physiol. Plant. 2013, 35, 2721–2726. [Google Scholar] [CrossRef]
- Habiba, A.U.; Kazuhiko, S.; Ahasan, M.; Alam, M. Effects of different light quality on growth and development of protocorm-like bodies (PLBs) in Dendrobium kingianum cultured in vitro. Bangladesh Res. Publ. J. 2014, 10, 223–227. [Google Scholar]
- Bello-Bello, J.J.; Martínez-Estrada, E.; Caamal-Velázquez, J.H.; Morales-Ramos, V. Effect of LED light quality on in vitro shoot proliferation and growth of vanilla (Vanilla planifolia Andrews). Afr. J. Biotechnol. 2016, 15, 272–277. [Google Scholar] [CrossRef]
- Coelho, A.D.; Souza, C.K.; Bertolucci, S.K.V.; Carvalho, A.A.; Santos, G.C.; Oliveira, T.; Marques, E.A.; Salimena, J.P.; Pinto, J.E.B.P. Wavelength and light intensity enhance growth, phytochemical contents and antioxidant activity in micropropagated plantlets of Urtica dioica L. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 145, 59–74. [Google Scholar] [CrossRef]
- Klimek-Szczykutowicz, M.; Prokopiuk, B.; Dziurka, K.; Pawłowska, B.; Ekiert, H.; Szopa, A. The influence of different wavelengths of LED light on the production of glucosinolates and phenolic compounds and the antioxidant potential in in vitro cultures of Nasturtium ofcinale (watercress). Plant Cell Tissue Organ Cult. (PCTOC) 2022, 149, 113–122. [Google Scholar] [CrossRef]
- García-López, J.I.; Zavala-García, F.; Olivares-Sáenz, E.; Lira-Saldívar, R.H.; Barriga-Castro, E.D.; Ruiz-Torres, N.A.; Ramos-Cortez, E.; Vázquez-Alvarado, R.; Niño-Medina, G. Zinc oxide nanoparticles boosts phenolic compounds and antioxidant activity of Capsicum annuum L. during germination. Agronomy 2018, 8, 215. [Google Scholar] [CrossRef]
- Kim, D.H.; Gopal, J.; Sivanesan, I. Nanomaterials in plant tissue culture: The disclosed and undisclosed. RSC Adv. 2017, 7, 6492–36505. [Google Scholar] [CrossRef]
- Garcia-Lópes, J.; Nino-Medina, G.; Olivares-Sàenz, E.; LiraSaldivar, R.; Barriga-Costro, E.; Vàzques-Alvarado, R.; Rodriguez-Salinas, P.; Zavala-Garcia, F. Foliar application of zinc oxide nanoparticles and zinc sulfate boosts the content of bioactive compound in Habanero peppers. Plants 2019, 8, 254. [Google Scholar] [CrossRef]
- Khan, A.U.; Khan, T.; Khan, M.A.; Nadhman, A.; Aasim, M.; Khan, N.Z.; Ali, W.; Nazir, N.; Zahoor, M. Iron-doped zinc oxide nanoparticles-triggered elicitation of important phenolic compounds in cell cultures of Fagonia indica. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 147, 287–296. [Google Scholar] [CrossRef]
- Zafar, H.; Alli, A.; Ali, J.S.; Haq, I.U.; Zia, M. Effect of ZnO nanoparticles on Brassica nigra seedlings and stem explants: Growth dynamics and antioxidative response. Front. Plant Sci. 2016, 7, 535. [Google Scholar] [CrossRef]
- Raigond, P.; Raigond, B.; Kaundal, B.; Singh, B.; Joshi, A.; Dutt, S. Effect of zinc nanoparticles on antioxidative system of potato plants. J. Environ. Biol. 2017, 38, 435–439. [Google Scholar] [CrossRef]
- Marslin, G.; Sheeba, C.J.; Franklin, G. Nanoparticles alter secondary metabolism in plants via ROS burst. Front. Plant Sci. 2017, 8, 832. [Google Scholar] [CrossRef]
- Michalak, A. Phenolic compounds and their antioxidant activity in plants growing under heavy metal stress. Pol. J. Environ. 2006, 15, 523–530. [Google Scholar]
- Chen, C.; Agrawal, D.C.; Lee, M.; Lee, R.; Kuo, C.; Wu, C.; Tsay, H.; Chang, H. Influence of LED light spectra on in vitro somatic embryogenesis and LC–MS analysis of chlorogenic acid and rutin in Peucedanum japonicum Thunb.: A medicinal herb. Bot. Stud. 2016, 57, 9. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Lin, C. Red light-emitting diode light irradiation improves root and leaf formation in difficult-to-propagate Protea cynaroides L. plantlets in vitro. HortScience 2012, 47, 1490–1494. [Google Scholar] [CrossRef]
- Guo, B.; Liu, Y.; Yan, Q.; Liu, C. Spectral composition of irradiation regulates the cell growth and flavonoids biosynthesis in callus cultures of Saussurea medusa Maxim. Plant Growth Regulat. 2007, 52, 259–263. [Google Scholar] [CrossRef]
- Urbonaviciute, A.; Samuoliene, G.; Brazaityte, A.; Duchovskis, P.; Ruzgas, V.; Zukauskas, A. The effect of variety and lighting quality on wheatgrass antioxidant properties. ZemdirbysteAgriculture 2009, 96, 119–128. [Google Scholar]
- Leal-Costa, M.V.; dos Santos Nascimento, L.B.; dos Santos Moreira, N.; Reinert, F.; Costa, S.; Lage, C.L.S.; Tavares, E.S. Influence of blue light on the leaf morphoanatomy of in vitro Kalanchoe pinnata (Lamarck) Persson (Crassulaceae). Microsc. Microanal. 2010, 16, 576–582. [Google Scholar] [CrossRef]
- Szopa, A.; Ekiert, H. The importance of applied light quality on the production of lignans and phenolic acids in Schisandra chinensis (Turcz.) Baill. cultures in vitro. Plant Cell Tissue Organ Cult. 2016, 127, 115–121. [Google Scholar] [CrossRef]
- Kawka, B.; Kwiecień, I.; Ekiert, H. Infuence of culture medium composition and light conditions on the accumulation of bioactive compounds in shoot cultures of Scutellaria laterifora L. (American Skullcap) grown in vitro. Appl. Biochem. Biotechnol. 2017, 183, 1414–1425. [Google Scholar] [CrossRef]
- Kubica, P.; Szopa, A.; Ekiert, H. Production of verbascoside and phenolic acids in biomass of Verbena ofcinalis L. (vervain) cultured under diferent in vitro conditions. Nat. Prod. Res. 2017, 31, 1663–1668. [Google Scholar] [CrossRef]
- Szopa, A.; Kokotkiewicz, A.; Bednarz, M.; Luczkiewicz, M.; Ekiert, H. Studies on the accumulation of phenolic acids and favonoids in diferent in vitro culture systems of Schisandra chinensis (Turcz.) Baill. using a DAD-HPLC method. Phytochem. Lett. 2017, 20, 462–469. [Google Scholar] [CrossRef]
- Szopa, A.; Starzec, A.; Ekiert, H. The importance of monochromatic lights in the production of phenolic acids and favonoids in shoot cultures of Aronia melanocarpa, Aronia arbutifolia and Aronia × prunifolia. J. Photochem. Photobiol. B 2018, 179, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Samuoliene, G.; Brazaityte, A.; Vaštakaitė-Kairienė, V. Light-emitting diodes (LEDs) for improved nutritional quality. In Light-Emitting Diodes for Agriculture; Dutta Gupta, S., Ed.; Springer: New York, NY, USA, 2017; pp. 149–190. [Google Scholar] [CrossRef]
- Zielińska, S.; Piątczak, E.; Kozłowska, W.; Bohater, A.; Jezierska-Domaradzka, A.; Kolniak-Ostek, J.; Matkowski, A. LED illumination and plant growth regulators’ efects on growth and phenolic acids accumulation in Moluccella laevis L. in vitro cultures. Acta Physiol. Plant. 2020, 42, 72. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Culture Condition | Bulblets per Regenerating Bulbsacale | Bulblet Diameter (mm) | Bulblet Weight (g) | Root per Bulblet |
|---|---|---|---|---|
| D a | 1.70 ± 0.1 a–c b | 4.21 ± 0.3 a | 0.14 ± 0.0 ab | 0.92 ± 0.1 ab |
| FL | 1.97 ± 0.1 bc | 4.23 ± 0.3 a | 0.14 ± 0.0 ab | 0.59 ± 0.1 a |
| D Zn25 | 2.07 ± 0.2 c | 4.29 ± 0.2 a | 0.22 ± 0.0 c | 0.60 ± 0.1 a |
| D Zn50 | 1.33 ± 0.1 a | 4.26 ± 0.3 a | 0.18 ± 0.0 bc | 0.84 ± 0.1 ab |
| D Zn75 | 1.59 ± 0.1 ab | 4.32 ± 0.2 a | 0.20 ± 0.0 c | 0.91 ± 0.1 ab |
| FL Zn25 | 2.01 ± 0.1 c | 4.18 ± 0.3 a | 0.10 ± 0.0 a | 0.51 ± 0.1 a |
| FL Zn50 | 1.74 ± 0.1 bc | 5.28 ± 0.4 a | 0.19 ± 0.0 bc | 1.02 ± 0.2 b |
| FL Zn75 | 1.78 ± 0.2 bc | 4.68 ± 0.4 a | 0.22 ± 0.0 c | 0.88 ± 0.1 ab |
| Source of variation | ||||
| Culture condition | *** | n.s. | *** | n.s. |
| Light | Bulblets per Regenerating Bulbsacale | Bulblet Diameter (mm) | Bulblet Weight (g) | Root per Bulblet |
|---|---|---|---|---|
| D a | 1.70 ± 0.1 ab b | 4.21 ± 0.3 a | 0.14 ± 0.0 ab | 0.92 ± 0.1 bc |
| FL | 1.97 ± 0.1 bc | 4.23 ± 0.3 a | 0.14 ± 0.0 ab | 0.59 ± 0.1 ab |
| R | 1.34 ± 0.1 a | 4.20 ± 0.4 a | 0.16 ± 0.0 bc | 0.81 ± 0.1 bc |
| B | 2.17 ± 0.1 bc | 4.06 ± 0.2 a | 0.11 ± 0.0 a | 0.39 ± 0.1 a |
| RB | 2.14 ± 0.2 bc | 4.63 ± 0.3 a | 0.24 ± 0.0 d | 0.98 ± 0.1 c |
| RBG | 1.99 ± 0.1 bc | 5.71 ± 0.2 b | 0.19 ± 0.0 c | 0.89 ± 0.1 bc |
| WLED | 2.34 ± 0.3 c | 4.20 ± 0.2 a | 0.11 ± 0.0 a | 0.67 ± 0.1 a-c |
| Source of variation | ||||
| Light | *** | *** | *** | *** |
| Culture Condition | Chlorogenic Acid | Caffeic Acid | p-Coumaric Acid | Ferulic Acid |
|---|---|---|---|---|
| D a | 1.79 ± 0.1 b b | 1.31 ± 0.0 a | 15.40 ± 0.2 b | 1.98 ± 0.0 ab |
| FL | 1.67 ± 0.2 b | 1.70 ± 0.1 b | 16.85 ± 0.1 b | 1.44 ± 0.0 a |
| D Zn | 0.99 ± 0.3 a | 1.25 ± 0.1 a | 6.82 ± 1.4 a | 1.88 ± 0.2 ab |
| FL Zn | 1.75 ± 0.2 b | 2.77 ± 0.1 c | 22.19 ± 2.3 c | 2.33 ± 0.8 b |
| Source of variation | ||||
| Culture condition | *** | n.s. | *** | n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pałka, P.; Muszyńska, B.; Szewczyk, A.; Pawłowska, B. Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro. Agronomy 2023, 13, 1437. https://doi.org/10.3390/agronomy13061437
Pałka P, Muszyńska B, Szewczyk A, Pawłowska B. Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro. Agronomy. 2023; 13(6):1437. https://doi.org/10.3390/agronomy13061437
Chicago/Turabian StylePałka, Piotr, Bożena Muszyńska, Agnieszka Szewczyk, and Bożena Pawłowska. 2023. "Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro" Agronomy 13, no. 6: 1437. https://doi.org/10.3390/agronomy13061437
APA StylePałka, P., Muszyńska, B., Szewczyk, A., & Pawłowska, B. (2023). Elicitation and Enhancement of Phenolics Synthesis with Zinc Oxide Nanoparticles and LED Light in Lilium candidum L. Cultures In Vitro. Agronomy, 13(6), 1437. https://doi.org/10.3390/agronomy13061437