Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leguminous and Non-Leguminous Litter Decomposition under Controlled Precipitation

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
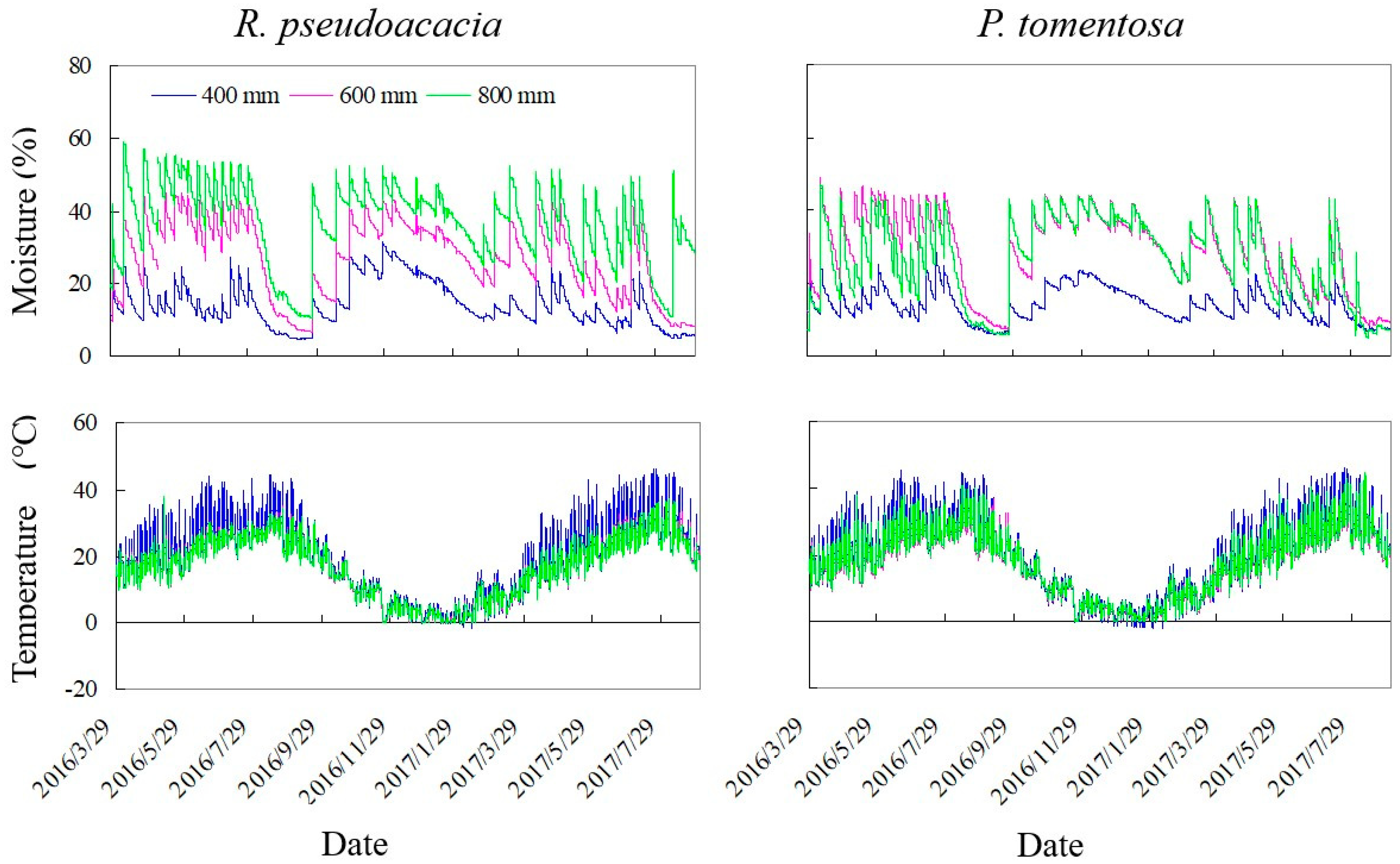
2.1. Study Site and Soil Properties
2.2. Experimental Design
2.3. Isotope Labeling and Leaf Sample Collection
2.4. Litter Collection and Soil Sampling
2.5. Chemical and Isotopic Analyses
2.6. Calculations and Statistical Analysis
3. Results
3.1. Remaining Mass of Litters
3.2. Leaf Litter Carbon and Nitrogen Dynamics
3.3. Leaf Litter δ13C and δ15N Dynamics
3.4. Soil Carbon and Nitrogen Dynamics
3.5. Soil δ13C and δ15N Dynamics
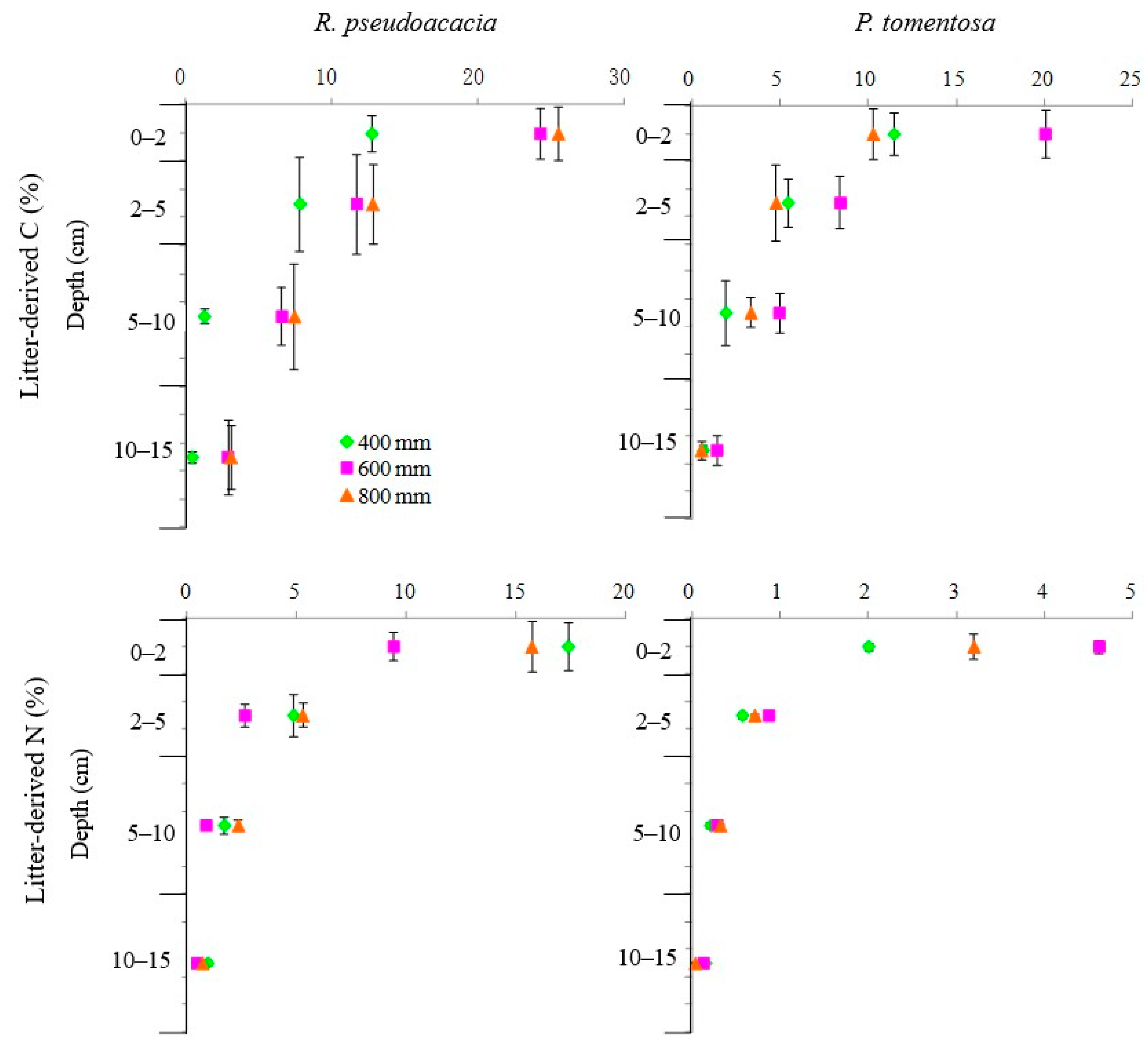
3.6. The Fractions of Litter-Derived Carbon and Nitrogen in Soil
3.7. Linear Carbon and Nitrogen Relationship
4. Discussion
4.1. Remaining Mass of Litters
4.2. Leaf Litter Carbon and Nitrogen Dynamics
4.3. Leaf Litter δ13C and δ15N Dynamics
4.4. Soil Carbon and Nitrogen Dynamics
4.5. Soil δ13C and δ15N Dynamics
4.6. The Fractions of Litter-Derived Carbon and Nitrogen in Soil
4.7. Linear Carbon and Nitrogen Relationship
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Bimüller, C.; Naumann, P.S.; Buegger, F.; Dannenmann, M.; Zeller, B.; von Lützow, M.; Kögel-Knabner, I. Rapid transfer of 15N from labeled beech leaf litter to functional soil organic matter fractions in a Rendzic Leptosol. Soil Biol. Biochem. 2013, 58, 323–331. [Google Scholar] [CrossRef]
- Cleveland, C.C.; Reed, S.C.; Keller, A.B.; Nemergut, D.R.; O’Neill, S.P.; Ostertag, R.; Vitousek, P.M. Litter quality versus soil microbial community controls over decomposition: A quantitative analysis. Oecologia 2014, 174, 283–294. [Google Scholar] [CrossRef]
- Qiu, L.; Wei, X.; Ma, T.; Wei, Y.; Horton, R.; Zhang, X.; Cheng, J. Effects of land-use change on soil organic carbon and nitrogen in density fractions and soil δ13C and δ15N in semiarid grasslands. Plant Soil 2015, 390, 419–430. [Google Scholar] [CrossRef]
- Pei, G.; Liu, J.; Peng, B.; Gao, D.; Wang, C.; Dai, W.; Jiang, P.; Bai, E. Nitrogen, lignin, C/N as important regulators of gross nitrogen release and immobilization during litter decomposition in a temperate forest ecosystem. For. Ecol. Manag. 2019, 440, 61–69. [Google Scholar] [CrossRef]
- Glaser, B.; Benesch, M.; Dippold, M.; Zech, W. In situ 15N and 13C labelling of indigenous and plantation tree species in a tropical mountain forest (Munessa, Ethiopia) for subsequent litter and soil organic matter turnover studies. Org. Geochem. 2012, 42, 1461–1469. [Google Scholar] [CrossRef]
- Gautam, M.K.; Lee, K.-S.; Song, B.-Y.; Lee, D.; Bong, Y.-S. Early-stage changes in natural 13C and 15N abundance and nutrient dynamics during different litter decomposition. J. Plant Res. 2016, 129, 463–476. [Google Scholar] [CrossRef]
- Rubino, M.; Dungait, J.; Evershed, R.; Bertolini, T.; De Angelis, P.; D’Onofrio, A.; Lagomarsino, A.; Lubritto, C.; Merola, A.; Terrasi, F. Carbon input belowground is the major C flux contributing to leaf litter mass loss: Evidences from a 13C labelled-leaf litter experiment. Soil Biol. Biochem. 2010, 42, 1009–1016. [Google Scholar] [CrossRef]
- Flessa, H.; Ludwig, B.; Heil, B.; Merbach, W. The origin of soil organic C, dissolved organic C and respiration in a long-term maize experiment in Halle, Germany, determined by 13C natural abundance. J. Plant Nutr. Soil Sci. 2000, 163, 157–163. [Google Scholar] [CrossRef]
- Berg, B.; McClaugherty, C. Plant Litter: Decomposition, Humus Formation, Carbon Sequestration; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Bai, X.; Huang, Y.; Wang, B.; Kuzyakov, Y.; An, S. Belowground allocation and fate of tree assimilates in plant–soil–microorganisms system: 13C labeling and tracing under field conditions. Geoderma 2021, 404, 115296. [Google Scholar] [CrossRef]
- Gong, C.; Tan, Q.; Liu, G.; Xu, M. Mixed-species plantations enhance soil carbon stocks on the loess plateau of China. Plant Soil 2021, 464, 13–28. [Google Scholar] [CrossRef]
- Rice, S.K.; Westerman, B.; Federici, R. Impacts of the exotic, nitrogen-fixing black locust (Robinia pseudoacacia) on nitrogen-cycling in a pine–oak ecosystem. Plant Ecol. 2004, 174, 97–107. [Google Scholar] [CrossRef]
- Wang, B.; Liu, G.; Xue, S. Effect of black locust (Robinia pseudoacacia) on soil chemical and microbiological properties in the eroded hilly area of China’s Loess Plateau. Environ. Earth Sci. 2012, 65, 597–607. [Google Scholar] [CrossRef]
- Ma, L.; Teng, Y.; Shangguan, Z. Ecohydrological responses to secondary natural Populus davidiana and plantation Pinus tabulaeformis woodlands on the Loess Plateau of China. Ecohydrology 2014, 7, 612–621. [Google Scholar] [CrossRef]
- Xia, L.; Song, X.; Fu, N.; Cui, S.; Li, L.; Li, H.; Li, Y. Effects of forest litter cover on hydrological response of hillslopes in the Loess Plateau of China. Catena 2019, 181, 104076. [Google Scholar] [CrossRef]
- Cornwell, W.K.; Cornelissen, J.H.; Amatangelo, K.; Dorrepaal, E.; Eviner, V.T.; Godoy, O.; Hobbie, S.E.; Hoorens, B.; Kurokawa, H.; Pérez-Harguindeguy, N. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecol. Lett. 2008, 11, 1065–1071. [Google Scholar] [CrossRef]
- Tateno, R.; Tokuchi, N.; Yamanaka, N.; Du, S.; Otsuki, K.; Shimamura, T.; Xue, Z.; Wang, S.; Hou, Q. Comparison of litterfall production and leaf litter decomposition between an exotic black locust plantation and an indigenous oak forest near Yan’an on the Loess Plateau, China. For. Ecol. Manag. 2007, 241, 84–90. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, Z.; Zhu, B.; Bing, Y.; Luc, N.T.; Du, L.; Zhu, Z. Impacts of mixed litter decomposition from Robinia pseudoacacia and other tree species on C loss and nutrient release in the Loess Plateau of China. J. For. Res. 2016, 27, 525–532. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Z.; Zha, T.; Luo, Z.; Zheng, J.; Sun, O.J. Predicting soil respiration using carbon stock in roots, litter and soil organic matter in forests of Loess Plateau in China. Soil Biol. Biochem. 2013, 57, 135–143. [Google Scholar] [CrossRef]
- Qian, L.; Liu, Z.; Du, L. Effects of mixed decomposition of Populus simonii and other tree species leaf litters on soil properties in Loess Plateau. Yingyong Shengtai Xuebao 2012, 23, 595–602. [Google Scholar] [CrossRef]
- Li, T.; Ren, B.; Wang, D.; Liu, G. Spatial variation in the storages and age-related dynamics of forest carbon sequestration in different climate zones—Evidence from black locust plantations on the Loess Plateau of China. PLoS ONE 2015, 10, e0121862. [Google Scholar] [CrossRef]
- Lu, N.; Liski, J.; Chang, R.; Akujärvi, A.; Wu, X.; Jin, T.; Wang, Y.; Fu, B. Soil organic carbon dynamics of black locust plantations in the middle Loess Plateau area of China. Biogeosciences 2013, 10, 7053–7063. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Z. Nitrogen isotopic composition of plant-soil in the Loess Plateau and its responding to environmental change. Chin. Sci. Bull. 2009, 54, 272–279. [Google Scholar] [CrossRef]
- Santonja, M.; Milcu, A.; Fromin, N.; Rancon, A.; Shihan, A.; Fernandez, C.; Baldy, V.; Hättenschwiler, S. Temporal shifts in plant diversity effects on carbon and nitrogen dynamics during litter decomposition in a Mediterranean shrubland exposed to reduced precipitation. Ecosystems 2019, 22, 939–954. [Google Scholar] [CrossRef]
- Sanaullah, M.; Rumpel, C.; Charrier, X.; Chabbi, A. How does drought stress influence the decomposition of plant litter with contrasting quality in a grassland ecosystem? Plant Soil 2012, 352, 277–288. [Google Scholar] [CrossRef]
- Liu, E.; Wang, J.; Zhang, Y.; Angers, D.A.; Yan, C.; Oweis, T.; He, W.; Liu, Q.; Chen, B. Priming effect of C-13-labelled wheat straw in no-tillage soil under drying and wetting cycles in the Loess Plateau of China. Sci. Rep. 2015, 5, 13826. [Google Scholar] [CrossRef]
- Gray, C.M.; Fierer, N. Impacts of nitrogen fertilization on volatile organic compound emissions from decomposing plant litter. Glob. Change Biol. 2012, 18, 739–748. [Google Scholar] [CrossRef]
- Ma, J.-Y.; Sun, W.; Liu, X.-N.; Chen, F.-H. Variation in the stable carbon and nitrogen isotope composition of plants and soil along a precipitation gradient in northern China. PLoS ONE 2012, 7, e51894. [Google Scholar] [CrossRef]
- Salamanca, E.F.; Kaneko, N.; Katagiri, S. Rainfall manipulation effects on litter decomposition and the microbial biomass of the forest floor. Appl. Soil Ecol. 2003, 22, 271–281. [Google Scholar] [CrossRef]
- Paul, K.; Polglase, P.; O’connell, A.; Carlyle, J.; Smethurst, P.; Khanna, P. Defining the relation between soil water content and net nitrogen mineralization. Eur. J. Soil Sci. 2003, 54, 39–48. [Google Scholar] [CrossRef]
- Joly, F.X.; Kurupas, K.L.; Throop, H.L. Pulse frequency and soil-litter mixing alter the control of cumulative precipitation over litter decomposition. Ecology 2017, 98, 2255–2260. [Google Scholar] [CrossRef]
- Santonja, M.; Fernandez, C.; Gauquelin, T.; Baldy, V. Climate change effects on litter decomposition: Intensive drought leads to a strong decrease of litter mixture interactions. Plant Soil 2015, 393, 69–82. [Google Scholar] [CrossRef]
- Zhang, J.; Gao, M.; Wen, X.; Chen, Y.; Yang, S.; Li, L.; Liao, Y. Effects of different fertilizers on soil enzyme activities and CO2 emission in dry-land of maize. Acta Ecol. Sin. 2012, 32, 6147–6154. [Google Scholar] [CrossRef]
- FAO-UNESCO. Soil Map of the World, Revised Legend; World Soil Resources Report 60; FAO: Rome, Italy, 1988. [Google Scholar]
- Du, N.; Li, W.; Qiu, L.; Zhang, Y.; Wei, X.; Zhang, X. Mass loss and nutrient release during the decomposition of sixteen types of plant litter with contrasting quality under three precipitation regimes. Ecol. Evol. 2020, 10, 3367–3382. [Google Scholar] [CrossRef] [PubMed]
- Bernard, M.J.; Pitz, S.L.; Chang, C.-H.; Szlavecz, K. Continuous 13C and 15N labeling of tree litter using a climate-controlled chamber. Commun. Soil Sci. Plant Anal. 2015, 46, 2721–2733. [Google Scholar] [CrossRef]
- Olson, J.S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology 1963, 44, 322–331. [Google Scholar] [CrossRef]
- Foster, D.R.; Aber, J.D. Forests in Time: The Environmental Consequences of 1000 Years of Change in New England; Yale University Press: New Haven, CT, USA, 2006. [Google Scholar]
- Arai, H.; Tokuchi, N. Factors contributing to greater soil organic carbon accumulation after afforestation in a Japanese coniferous plantation as determined by stable and radioactive isotopes. Geoderma 2010, 157, 243–251. [Google Scholar] [CrossRef]
- Xu, X.; Hirata, E. Decomposition patterns of leaf litter of seven common canopy species in a subtropical forest: N and P dynamics. Plant Soil 2005, 273, 279–289. [Google Scholar] [CrossRef]
- Lensing, J.R.; Wise, D.H. Impact of changes in rainfall amounts predicted by climate-change models on decomposition in a deciduous forest. Appl. Soil Ecol. 2007, 35, 523–534. [Google Scholar] [CrossRef]
- Zhou, S.; Huang, C.; Xiang, Y.; Tie, L.; Han, B.; Scheu, S. Effects of reduced precipitation on litter decomposition in an evergreen broad-leaved forest in western China. For. Ecol. Manag. 2018, 430, 219–227. [Google Scholar] [CrossRef]
- Austin, A.T.; Vitousek, P.M. Precipitation, decomposition and litter decomposability of Metrosideros polymorpha in native forests on Hawai’i. J. Ecol. 2000, 88, 129–138. [Google Scholar] [CrossRef]
- Garcia-Pausas, J.; Casals, P.; Romanya, J. Litter decomposition and faunal activity in Mediterranean forest soils: Effects of N content and the moss layer. Soil Biol. Biochem. 2004, 36, 989–997. [Google Scholar] [CrossRef]
- Osono, T.; Azuma, J.-i.; Hirose, D. Plant species effect on the decomposition and chemical changes of leaf litter in grassland and pine and oak forest soils. Plant Soil 2014, 376, 411–421. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Easdale, T.A.; Pérez-Harguindeguy, N. Subtropical montane tree litter decomposition: Links with secondary forest types and species’ shade tolerance. Austral Ecol. 2003, 28, 666–673. [Google Scholar] [CrossRef]
- Drechsel, P.; Glaser, B.; Zech, W. Effect of four multipurpose tree species on soil amelioration during tree fallow in Central Togo. Agrofor. Syst. 1991, 16, 193–202. [Google Scholar] [CrossRef]
- Connin, S.; Feng, X.; Virginia, R. Isotopic discrimination during long-term decomposition in an arid land ecosystem. Soil Biol. Biochem. 2001, 33, 41–51. [Google Scholar] [CrossRef]
- Hart, S.C.; Nason, G.; Myrold, D.D.; Perry, D.A. Dynamics of gross nitrogen transformations in an old-growth forest: The carbon connection. Ecology 1994, 75, 880–891. [Google Scholar] [CrossRef]
- Parton, W.; Silver, W.L.; Burke, I.C.; Grassens, L.; Harmon, M.E.; Currie, W.S.; King, J.Y.; Adair, E.C.; Brandt, L.A.; Hart, S.C. Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 2007, 315, 361–364. [Google Scholar] [CrossRef]
- Wei, X.; Li, X.; Jia, X.; Shao, M. Accumulation of soil organic carbon in aggregates after afforestation on abandoned farmland. Biol. Fertil. Soils 2013, 49, 637–646. [Google Scholar] [CrossRef]
- Fahey, T.J.; Yavitt, J.B.; Sherman, R.E.; Groffman, P.M.; Fisk, M.C.; Maerz, J.C. Transport of carbon and nitrogen between litter and soil organic matter in a northern hardwood forest. Ecosystems 2011, 14, 326–340. [Google Scholar] [CrossRef]
- Fernandez, I.; Mahieu, N.; Cadisch, G. Carbon isotopic fractionation during decomposition of plant materials of different quality. Glob. Biogeochem. Cycles 2003, 17, 1075. [Google Scholar] [CrossRef]
- Ngao, J.J.; Cotrufo, M.-F.M.-F. Carbon isotope discrimination during litter decomposition can be explained by selective use of substrate with differing δ13C. Biogeosci. Discuss. 2011, 8, 51–82. [Google Scholar] [CrossRef]
- Xu, S.; Liu, Y.; Cui, Y.; Pei, Z. Litter decomposition in a subtropical plantation in Qianyanzhou, China. J. For. Res. 2011, 16, 8–15. [Google Scholar] [CrossRef]
- Ono, K.; Hiradate, S.; Morita, S.; Ohse, K.; Hirai, K. Humification processes of needle litters on forest floors in Japanese cedar (Cryptomeria japonica) and Hinoki cypress (Chamaecyparis obtusa) plantations in Japan. Plant Soil 2011, 338, 171–181. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Hobbie, J.E. Natural abundance of 15N in nitrogen-limited forests and tundra can estimate nitrogen cycling through mycorrhizal fungi: A review. Ecosystems 2008, 11, 815–830. [Google Scholar] [CrossRef]
- Bragazza, L.; Iacumin, P.; Siffi, C.; Gerdol, R. Seasonal variation in nitrogen isotopic composition of bog plant litter during 3 years of field decomposition. Biol. Fertil. Soils 2010, 46, 877–881. [Google Scholar] [CrossRef]
- Perakis, S.S.; Sinkhorn, E.R. Biogeochemistry of a temperate forest nitrogen gradient. Ecology 2011, 92, 1481–1491. [Google Scholar] [CrossRef]
- Dijkstra, P.; LaViolette, C.M.; Coyle, J.S.; Doucett, R.R.; Schwartz, E.; Hart, S.C.; Hungate, B.A. 15N enrichment as an integrator of the effects of C and N on microbial metabolism and ecosystem function. Ecol. Lett. 2008, 11, 389–397. [Google Scholar] [CrossRef]
- Murphy, B.P.; Bowman, D.M. The carbon and nitrogen isotope composition of Australian grasses in relation to climate. Funct. Ecol. 2009, 23, 1040–1049. [Google Scholar] [CrossRef]
- Handley, L.; Odee, D.; Scrimgeour, C. δ15N and δ13C patterns in savanna vegetation: Dependence on water availability and disturbance. Funct. Ecol. 1994, 8, 306–314. [Google Scholar] [CrossRef]
- Welker, J.M.; Jónsdóttir, I.S.; Fahnestock, J.T. Leaf isotopic (δ13C and δ15N) and nitrogen contents of Carex plants along the Eurasian Coastal Arctic: Results from the Northeast Passage expedition. Polar Biol. 2003, 27, 29–37. [Google Scholar] [CrossRef]
- Schmidt, M.W.; Torn, M.S.; Abiven, S.; Dittmar, T.; Guggenberger, G.; Janssens, I.A.; Kleber, M.; Kögel-Knabner, I.; Lehmann, J.; Manning, D.A. Persistence of soil organic matter as an ecosystem property. Nature 2011, 478, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Luo, Y.; Xu, X.; Sherry, R.; Zhang, Q. Soil organic matter dynamics in a North America tallgrass prairie after 9 yr of experimental warming. Biogeosciences 2011, 8, 1487–1498. [Google Scholar] [CrossRef]
- Cao, J.; He, X.; Chen, Y.; Chen, Y.; Zhang, Y.; Yu, S.; Zhou, L.; Liu, Z.; Zhang, C.; Fu, S. Leaf litter contributes more to soil organic carbon than fine roots in two 10-year-old subtropical plantations. Sci. Total Environ. 2020, 704, 135341. [Google Scholar] [CrossRef] [PubMed]
- Garten, C.T., Jr.; Cooper, L.W.; Post, W., III; Hanson, P.J. Climate controls on forest soil C isotope ratios in the southern Appalachian Mountains. Ecology 2000, 81, 1108–1119. [Google Scholar] [CrossRef]
- Hatton, P.-J.; Kleber, M.; Zeller, B.; Moni, C.; Plante, A.F.; Townsend, K.; Gelhaye, L.; Lajtha, K.; Derrien, D. Transfer of litter-derived N to soil mineral–organic associations: Evidence from decadal 15N tracer experiments. Org. Geochem. 2012, 42, 1489–1501. [Google Scholar] [CrossRef]
- Bowden, R.D.; Deem, L.; Plante, A.F.; Peltre, C.; Nadelhoffer, K.; Lajtha, K. Litter input controls on soil carbon in a temperate deciduous forest. Soil Sci. Soc. Am. J. 2014, 78, S66–S75. [Google Scholar] [CrossRef]
- Lajtha, K.; Bowden, R.D.; Nadelhoffer, K. Litter and root manipulations provide insights into soil organic matter dynamics and stability. Soil Sci. Soc. Am. J. 2014, 78, S261–S269. [Google Scholar] [CrossRef]
- Xu, S.; Liu, L.; Sayer, E.J. Variability of above-ground litter inputs alters soil physicochemical and biological processes: A meta-analysis of litterfall-manipulation experiments. Biogeosciences 2013, 10, 7423–7433. [Google Scholar] [CrossRef]
- Tuo, D.; Gao, G.; Chang, R.; Li, Z.; Ma, Y.; Wang, S.; Wang, C.; Fu, B. Effects of revegetation and precipitation gradient on soil carbon and nitrogen variations in deep profiles on the Loess Plateau of China. Sci. Total Environ. 2018, 626, 399–411. [Google Scholar] [CrossRef]
- Díaz-Raviña, M.; Bueno, J.; González-Prieto, S.J.; Carballas, T. Cultivation effects on biochemical properties, C storage and 15N natural abundance in the 0–5 cm layer of an acidic soil from temperate humid zone. Soil Tillage Res. 2005, 84, 216–221. [Google Scholar] [CrossRef]
- Hagedorn, F.; Kammer, A.; Schmidt, M.W.; Goodale, C.L. Nitrogen addition alters mineralization dynamics of 13C-depleted leaf and twig litter and reduces leaching of older DOC from mineral soil. Glob. Chang. Biol. 2012, 18, 1412–1427. [Google Scholar] [CrossRef]
- Rubino, M.; Lubritto, C.; D’Onofrio, A.; Terrasi, F.; Gleixner, G.; Cotrufo, M. An isotopic method for testing the influence of leaf litter quality on carbon fluxes during decomposition. Oecologia 2007, 154, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Zeller, B.; Dambrine, E. Coarse particulate organic matter is the primary source of mineral N in the topsoil of three beech forests. Soil Biol. Biochem. 2011, 43, 542–550. [Google Scholar] [CrossRef]
- Berg, B. Litter decomposition and organic matter turnover in northern forest soils. For. Ecol. Manag. 2000, 133, 13–22. [Google Scholar] [CrossRef]
- García-Palacios, P.; Prieto, I.; Ourcival, J.-M.; Hättenschwiler, S. Disentangling the litter quality and soil microbial contribution to leaf and fine root litter decomposition responses to reduced rainfall. Ecosystems 2016, 19, 490–503. [Google Scholar] [CrossRef]
- Horodecki, P.; Jagodziński, A.M. Tree species effects on litter decomposition in pure stands on afforested post-mining sites. For. Ecol. Manag. 2017, 406, 1–11. [Google Scholar] [CrossRef]
- Marichal, R.; Mathieu, J.; Couteaux, M.-M.; Mora, P.; Roy, J.; Lavelle, P. Earthworm and microbe response to litter and soils of tropical forest plantations with contrasting C:N:P stoichiometric ratios. Soil Biol. Biochem. 2011, 43, 1528–1535. [Google Scholar] [CrossRef]
- Fan, F.; Yin, C.; Tang, Y.; Li, Z.; Song, A.; Wakelin, S.A.; Zou, J.; Liang, Y. Probing potential microbial coupling of carbon and nitrogen cycling during decomposition of maize residue by 13C-DNA-SIP. Soil Biol. Biochem. 2014, 70, 12–21. [Google Scholar] [CrossRef]
- Prescott, C.E. Litter decomposition: What controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry 2010, 101, 133–149. [Google Scholar] [CrossRef]
- Zhang, K.; Cheng, X.; Dang, H.; Ye, C.; Zhang, Y.; Zhang, Q. Linking litter production, quality and decomposition to vegetation succession following agricultural abandonment. Soil Biol. Biochem. 2013, 57, 803–813. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Months | 800 mm | 600 mm | 400 mm | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Week 1 | Week 2 | Week 3 | Week 4 | Sub-Sum | Week 1 | Week 2 | Week 3 | Week 4 | Sub-Sum | Week 1 | Week 2 | Week 3 | Week 4 | Sub-Sum | |
| November | 15.9 | 0.0 | 15.9 | 0.0 | 31.8 | 12.7 | 0.0 | 12.7 | 0.0 | 25.4 | 8.0 | 0.0 | 8.0 | 0.0 | 16.0 |
| December | 6.4 | 0.0 | 6.4 | 0.0 | 12.8 | 4.8 | 0.0 | 4.8 | 0.0 | 9.6 | 3.2 | 0.0 | 3.2 | 0.0 | 6.4 |
| January | 6.4 | 0.0 | 6.4 | 0.0 | 12.8 | 4.8 | 0.0 | 4.8 | 0.0 | 9.6 | 3.2 | 0.0 | 3.2 | 0.0 | 6.4 |
| February | 9.6 | 0.0 | 9.6 | 0.0 | 19.2 | 8.0 | 0.0 | 8.0 | 0.0 | 16.0 | 4.8 | 0.0 | 4.8 | 0.0 | 9.6 |
| March | 19.1 | 0.0 | 19.1 | 0.0 | 38.2 | 14.3 | 0.0 | 14.3 | 0.0 | 28.6 | 9.6 | 0.0 | 9.6 | 0.0 | 19.2 |
| April | 38.2 | 0.0 | 38.2 | 0.0 | 76.4 | 28.7 | 0.0 | 28.7 | 0.0 | 57.4 | 19.1 | 0.0 | 19.1 | 0.0 | 38.2 |
| May | 25.5 | 25.5 | 25.5 | 25.5 | 102.0 | 19.1 | 19.1 | 19.1 | 19.1 | 76.4 | 12.7 | 12.7 | 12.7 | 12.7 | 50.8 |
| June | 19.1 | 19.1 | 19.1 | 19.1 | 76.4 | 14.3 | 14.3 | 14.3 | 14.3 | 57.2 | 9.6 | 9.6 | 9.6 | 9.6 | 38.4 |
| July | 38.2 | 38.2 | 38.2 | 38.2 | 152.8 | 28.7 | 28.7 | 28.7 | 28.7 | 114.8 | 19.1 | 19.1 | 19.1 | 19.1 | 76.4 |
| August | 5.1 | 0.0 | 5.1 | 0.0 | 10.2 | 3.8 | 0.0 | 3.8 | 0.0 | 7.6 | 2.5 | 0.0 | 2.5 | 0.0 | 5.0 |
| September | 41.4 | 41.4 | 41.4 | 41.4 | 165.6 | 30.3 | 30.3 | 30.3 | 30.3 | 121.2 | 20.7 | 20.7 | 20.7 | 20.7 | 82.8 |
| October | 25.5 | 25.5 | 25.5 | 25.5 | 102.0 | 19.1 | 19.1 | 19.1 | 19.1 | 76.4 | 12.7 | 12.7 | 12.7 | 12.7 | 50.8 |
| R. pseudoacacia | P. tomentosa | |||||
|---|---|---|---|---|---|---|
| Precipitation | 400 mm | 600 mm | 800 mm | 400 mm | 600 mm | 800 mm |
| RM | 72.73 a | 63.44 b | 60.00 b | 65.53 a | 50.76 b | 61.03 a |
| k | 0.37 a | 0.41 a | 0.45 a | 0.33 b | 0.56 a | 0.58 a |
| Initial C | 492.0 ± 3.0 | 492.0 ± 3.0 | 492.0 ± 3.0 | 517.0 ± 2.0 | 517.0 ± 2.0 | 517.0 ± 2.0 |
| Final C | 385.2 ± 8.0 a | 399.0 ± 10.0 a | 384.4 ± 8.0 a | 385.1 ± 5.0 a | 364.7 ± 3.0 a | 375.6 ± 6.0 a |
| Initial N | 44.2 ± 0.6 | 44.2 ± 0.6 | 44.2 ± 0.6 | 36.6 ± 1.4 | 36.6 ± 1.4 | 36.6 ± 1.4 |
| Final N | 37.6 ± 0.6 a | 37.6 ± 0.8 a | 35.9 ± 1.0 a | 23.7 ± 2.2 a | 25.4 ± 1.8 a | 29.6 ± 1.9 a |
| Initial C/N | 11.1 ± 0.6 | 11.1 ± 0.6 | 11.1 ± 0.6 | 14.14 ± 1.28 | 14.14 ± 1.28 | 14.14 ± 1.28 |
| Final C/N | 10.2 ± 0.6 a | 10.6 ± 0.8 a | 10.7 ± 1.0 a | 16.2 ± 1.2 a | 14.4 ± 1.8 ab | 12.7 ± 1.2 b |
| r | R2 | p | RMSE | a | b | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Whole | 0.396 ** | 0.149 | <0.001 | 5.775 | 0.070 | 4.557 | ||||||
| RP | PT | RP | PT | RP | PT | RP | PT | RP | PT | RP | PT | |
| Precipitation regimes | ||||||||||||
| 400 mm | 0.740 ** | 0.018 | 0.515 | –0.071 | 0.001 | 0.948 | 1.746 | 4.498 | 0.050 | 0.002 | 17.177 | 28.872 |
| 600 mm | 0.853 ** | 0.391 | 0.710 | 0.096 | <0.001 | 0.121 | 1.607 | 2.585 | 0.082 | 0.030 | 4.121 | 15.318 |
| 800 mm | 0.843 ** | 0.314 | 0.711 | 0.046 | <0.001 | 0.190 | 1.766 | 4.539 | 0.083 | 0.050 | 3.993 | 9.341 |
| Decomposition times | ||||||||||||
| 2 months | 0.661 | –0.390 | 0.356 | 0.031 | 0.053 | 0.299 | 1.803 | 5.494 | 0.137 | –0.129 | –22.231 | 90.290 |
| 4 months | 0.331 | 0.416 | 0.039 | 0.035 | 0.423 | 0.306 | 1.485 | 3.688 | 0.039 | 0.049 | 24.010 | 6.288 |
| 6 months | 0.285 | 0.186 | 0.072 | 0.126 | 0.494 | 0.659 | 1.332 | 3.938 | 0.018 | 0.037 | 32.438 | 14.839 |
| 8 months | 0.467 | 0.704 | 0.106 | 0.411 | 0.205 | 0.051 | 1.543 | 3.561 | 0.026 | 0.119 | 28.009 | –20.271 |
| 12 months | 0.962 ** | 0.131 | 0.909 | 0.147 | 0.001 | 0.757 | 0.663 | 3.101 | 0.073 | 0.023 | 5.696 | 18.576 |
| 16 months | 0.573 | –0.095 | 0.245 | –0.101 | 0.083 | 0.782 | 1.420 | 3.994 | 0.048 | –0.030 | 18.149 | 39.741 |
| r | R2 | p | RMSE | a | b | |
|---|---|---|---|---|---|---|
| Layers | ||||||
| 0–2 cm | 0.604 ** | 0.347 | <0.001 | 0.096 | 0.171 | –0.065 |
| 2–5 cm | 0.188 | 0.007 | 0.272 | 0.101 | 0.090 | 0.239 |
| 5–10 cm | 0.502 ** | 0.230 | 0.002 | 0.089 | 0.170 | –0.047 |
| 10–15 cm | 0.557 ** | 0.289 | <0.001 | 0.084 | 0.169 | –0.035 |
| Precipitation regimes | ||||||
| 400 mm | 0.614 ** | 0.363 | <0.001 | 0.050 | 0.102 | 0.182 |
| 600 mm | 0.586 ** | 0.329 | <0.001 | 0.104 | 0.229 | –0.249 |
| 800 mm | 0.457 ** | 0.192 | 0.001 | 0.107 | 0.145 | 0.048 |
| Decomposition times | ||||||
| Control soil | 0.533 ** | 0.251 | 0.007 | 0.134 | 0.299 | –0.509 |
| 2 months | 0.882 ** | 0.767 | <0.001 | 0.048 | 0.276 | –0.560 |
| 4 months | 0.731 ** | 0.514 | <0.001 | 0.042 | 0.183 | –0.041 |
| 6 months | 0.723 ** | 0.501 | <0.001 | 0.077 | 0.198 | –0.047 |
| 8 months | 0.675 ** | 0.431 | <0.001 | 0.057 | 0.095 | 0.243 |
| 12 months | 0.857 ** | 0.723 | <0.001 | 0.020 | 0.104 | 0.165 |
| 16 months | 0.753 ** | 0.547 | <0.001 | 0.022 | 0.107 | 0.147 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, N.; Zheng, K.; Zhang, J.; Qiu, L.; Zhang, Y.; Wei, X.; Zhang, X. Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leguminous and Non-Leguminous Litter Decomposition under Controlled Precipitation. Agronomy 2023, 13, 1205. https://doi.org/10.3390/agronomy13051205
Du N, Zheng K, Zhang J, Qiu L, Zhang Y, Wei X, Zhang X. Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leguminous and Non-Leguminous Litter Decomposition under Controlled Precipitation. Agronomy. 2023; 13(5):1205. https://doi.org/10.3390/agronomy13051205
Chicago/Turabian StyleDu, Ningning, Ke Zheng, Jingshun Zhang, Liping Qiu, Yanjiang Zhang, Xiaorong Wei, and Xingchang Zhang. 2023. "Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leguminous and Non-Leguminous Litter Decomposition under Controlled Precipitation" Agronomy 13, no. 5: 1205. https://doi.org/10.3390/agronomy13051205
APA StyleDu, N., Zheng, K., Zhang, J., Qiu, L., Zhang, Y., Wei, X., & Zhang, X. (2023). Dual Isotopes Tracing Carbon and Nitrogen Dynamics during Leguminous and Non-Leguminous Litter Decomposition under Controlled Precipitation. Agronomy, 13(5), 1205. https://doi.org/10.3390/agronomy13051205