
Leaf Trait Hyperspectral Characterization of Castanea sativa Miller Affected by Dryocosmus kuriphilus Yasumatsu

 ,
,  , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
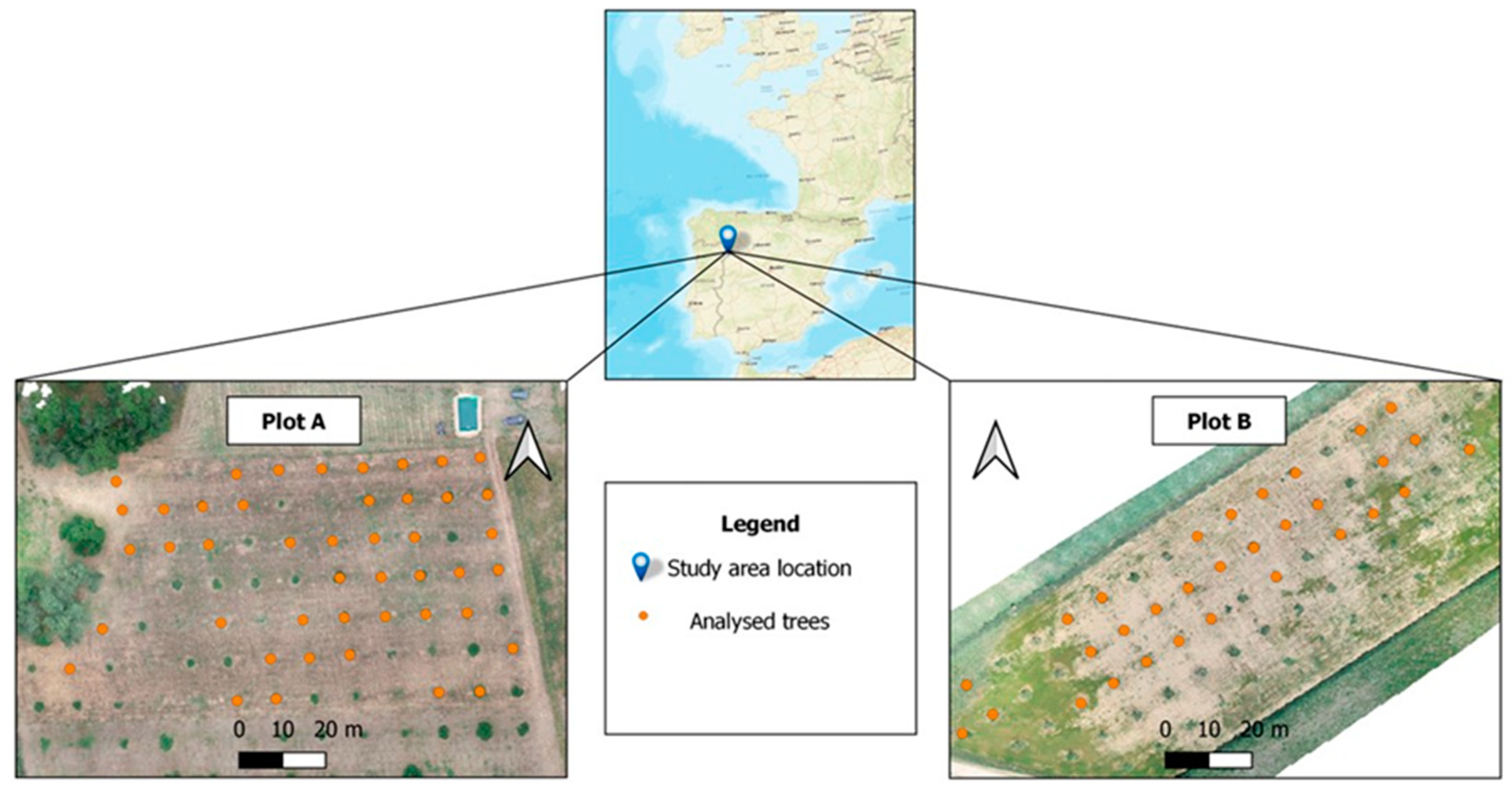
2.1. Study Area and Sampling Design
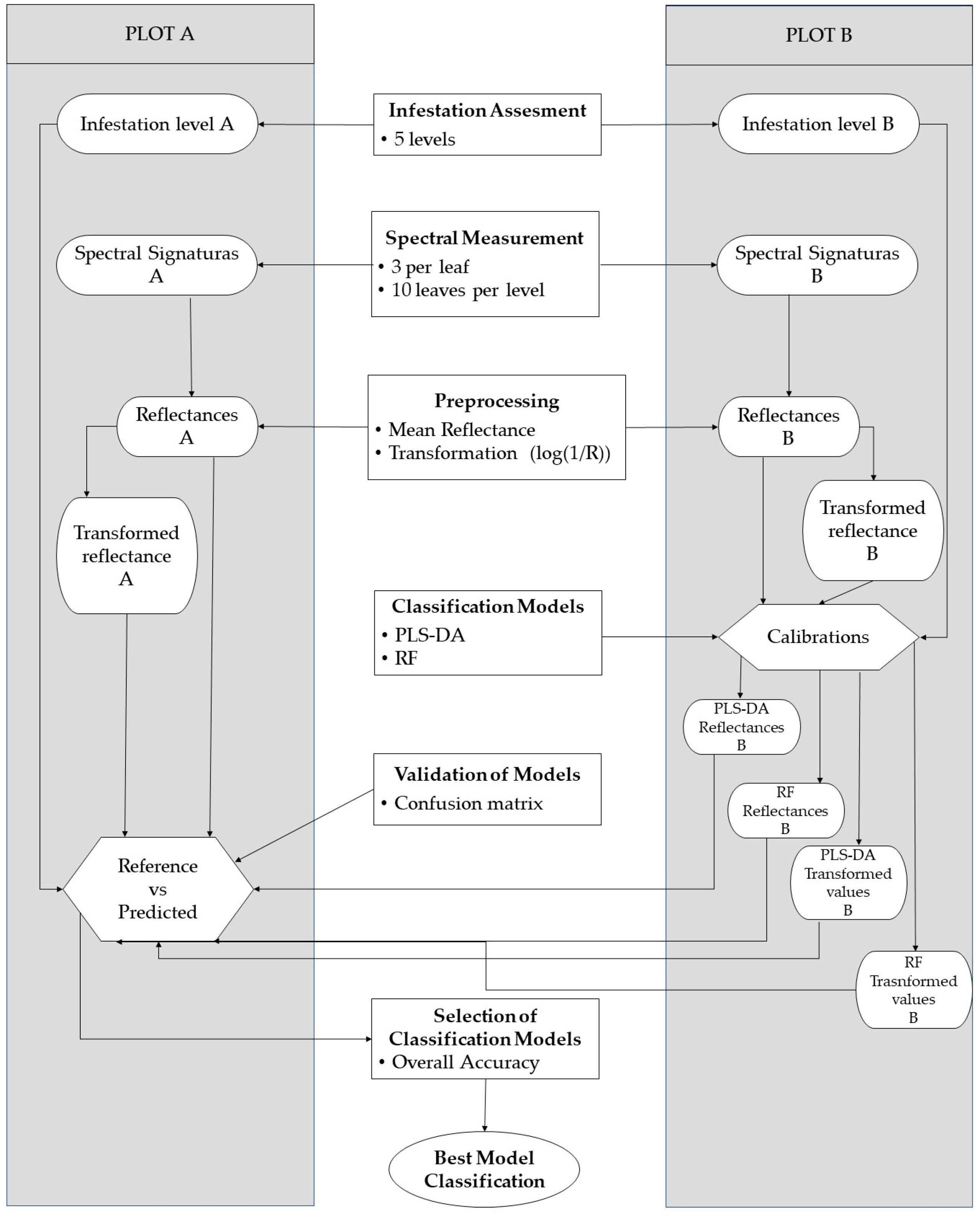
2.2. Experimental Workflow
2.3. Assessing Gall Wasp Infestation Levels
2.4. Field Spectroradiometer Measurements
2.5. Preprocessing
2.6. Classification (Training and Validation)
3. Results
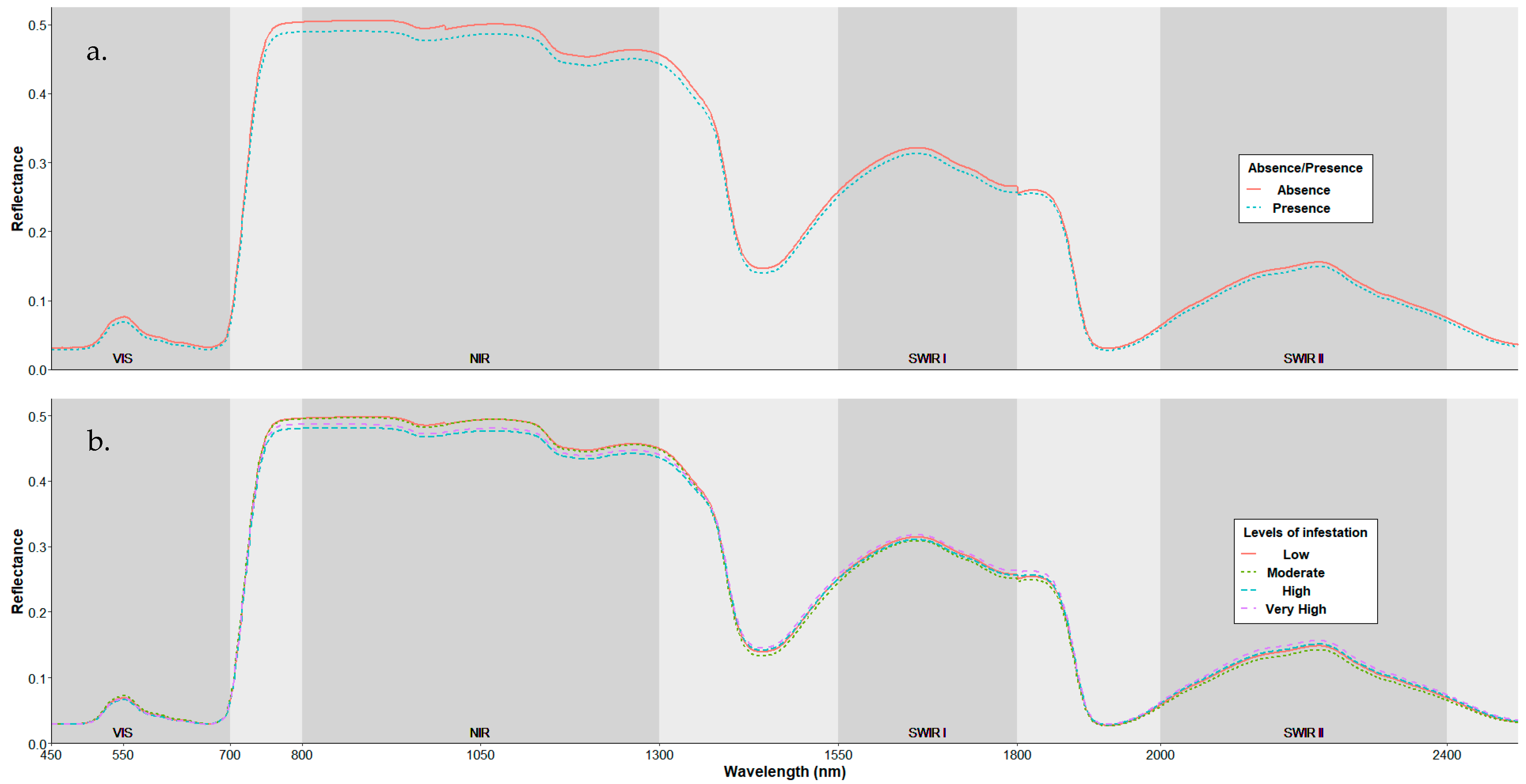
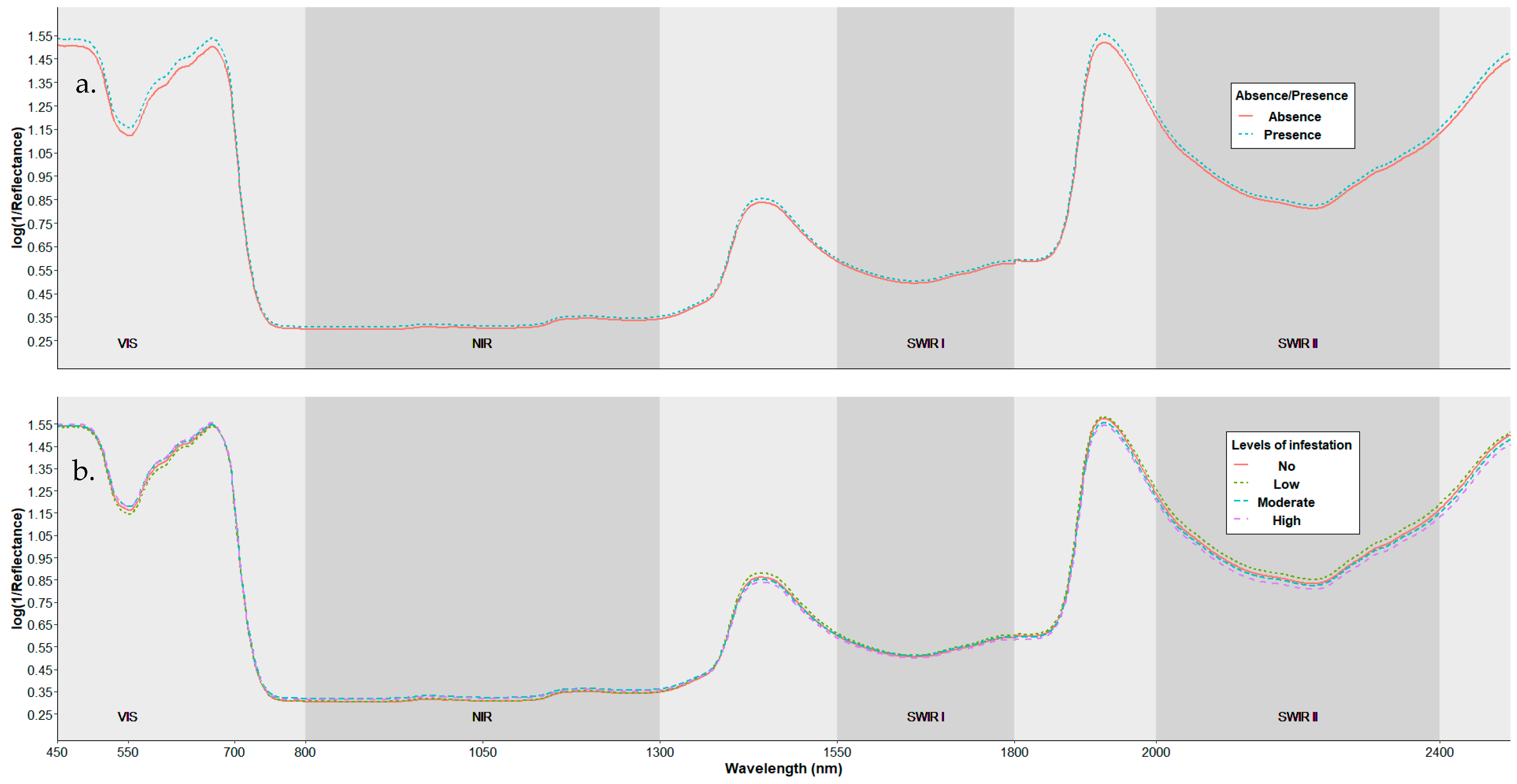
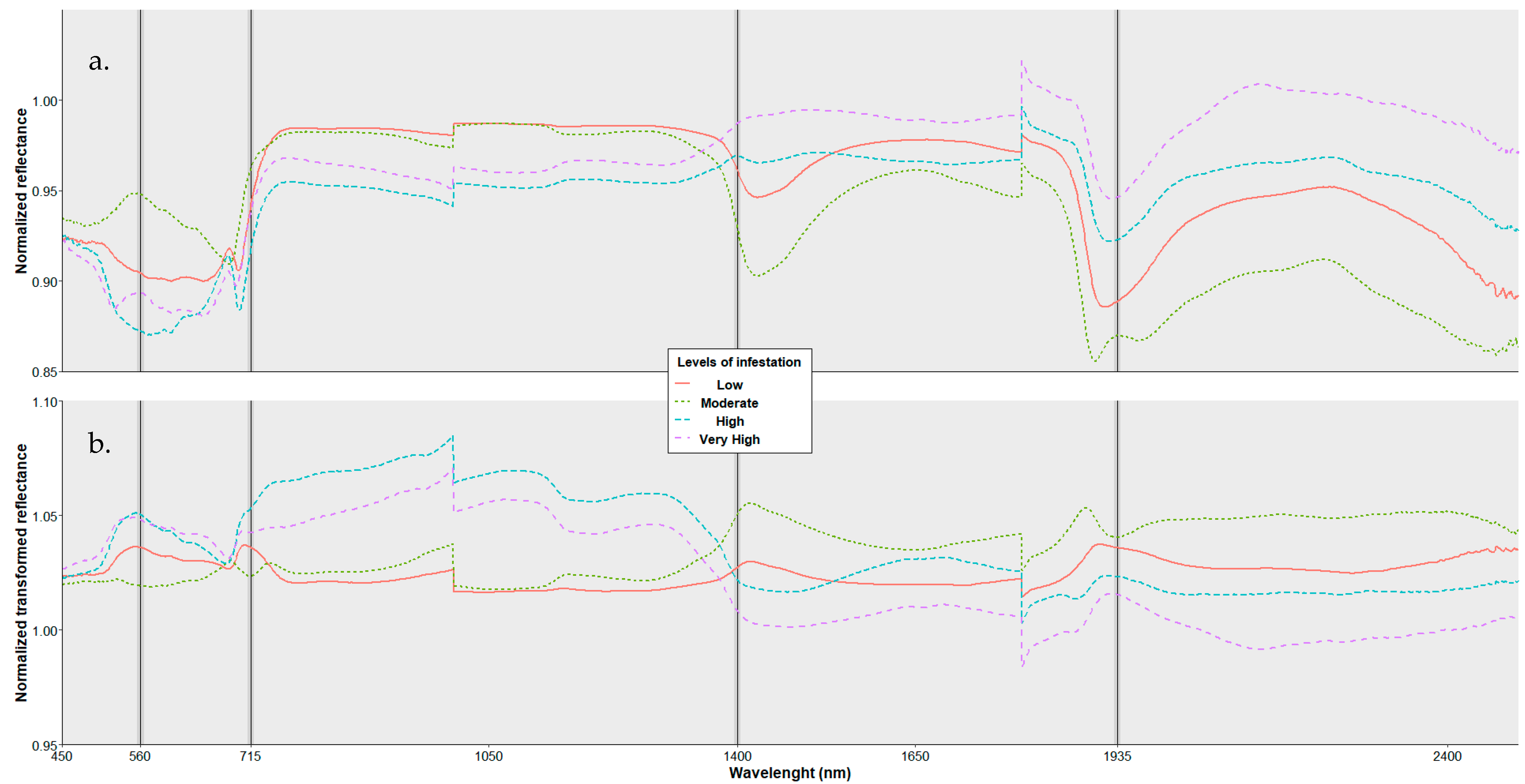
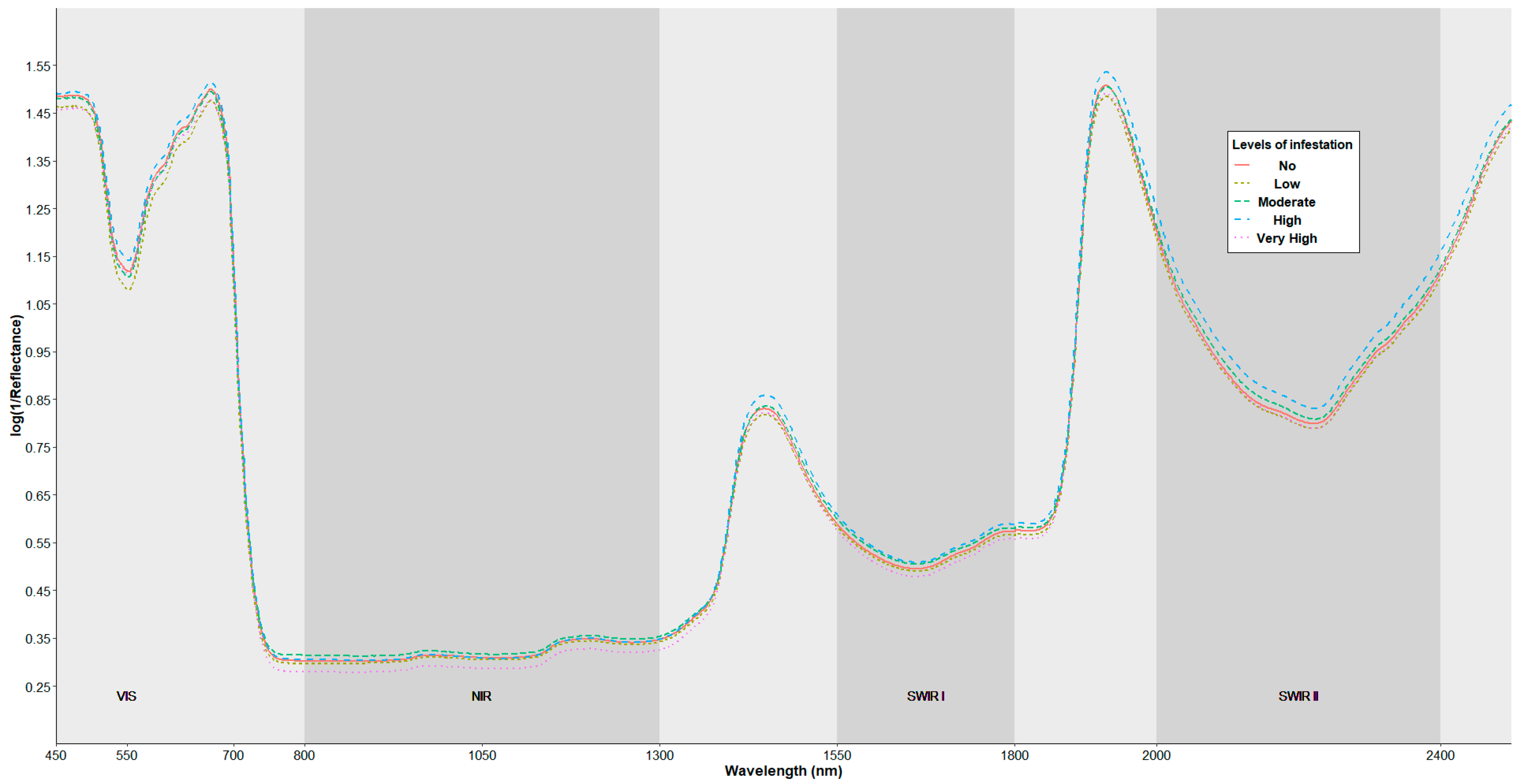
3.1. Spectral Characterization
3.2. Variable Selection
3.3. Classification Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conedera, M.; Manetti, M.C.; Giudici, F.; Amorini, E. Distribution and Economic Potential of the Sweet Chestnut (Castanea sativa Mill.) in Europe. Ecol. Mediterr. 2004, 30, 179–193. [Google Scholar] [CrossRef]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 30 January 2023).
- Biaggi, M.D.; Beccaro, G.; Casey, J.; Riqué, P.H.; Conedera, M.; Gomes-Laranjo, J.; Fulbright, D.W.; Nishio, S.; Serdar, Ü.; Zou, F.; et al. Distribution, Marketing, and Trade. In The Chestnut Handbook; CRC Press: Boca Raton, FL, USA, 2019; ISBN 978-0-429-44560-6. [Google Scholar]
- Fernandes, P.; Colavolpe, M.B.; Serrazina, S.; Costa, R.L. European and American Chestnuts: An Overview of the Main Threats and Control Efforts. Front. Plant Sci. 2022, 13, 951844. [Google Scholar] [CrossRef] [PubMed]
- Gil-Tapetado, D.; Gomez, J.F.; Cabrero-Sanudo, F.J.; Nieves-Aldrey, J.L. Distribution and Dispersal of the Invasive Asian Chestnut Gall Wasp, Dryocosmus kuriphilus (Hymenoptera: Cynipidae), across the Heterogeneous Landscape of the Iberian Peninsula. Eur. J. Entomol. 2018, 115, 575–586. [Google Scholar] [CrossRef]
- Sartor, C.; Dini, F.; Torello Marinoni, D.; Mellano, M.G.; Beccaro, G.L.; Alma, A.; Quacchia, A.; Botta, R. Impact of the Asian Wasp Dryocosmus kuriphilus (Yasumatsu) on Cultivated Chestnut: Yield Loss and Cultivar Susceptibility. Sci. Hortic. 2015, 197, 454–460. [Google Scholar] [CrossRef]
- Míguez-Soto, B.; Chamorro, E.M.; López, J.F. TOLERANCIA A LA AVISPA DEL CASTAÑO (DRYOCOSMUS KURIPHILUS) EN VARIEDADES TRADICIONALES DE FRUTO E HÍBRIDOS INTERESPECÍFICOS. 2018. Available online: https://lourizan.xunta.gal/es/transferencias/tolerancia-la-avispa-del-castano-dryocosmus-kuriphilus-envariedades-tradicionales-de (accessed on 21 July 2022).
- Nugnes, F.; Gualtieri, L.; Bonsignore, C.P.; Parillo, R.; Annarumma, R.; Griffo, R.; Bernardo, U. Resistance of a Local Ecotype of Castanea sativa to Dryocosmus kuriphilus (Hymenoptera: Cynipidae) in Southern Italy. Forests 2018, 9, 94. [Google Scholar] [CrossRef]
- Bombi, P.; Fedi, C.; Zapparoli, M.; Cammarano, M.; Guidolotti, G.; Pallozzi, E.; Gaudet, M.; Mattioni, C.; Cherubini, M.; Beritognolo, I.; et al. Infestation Potential of Dryocosmus kuriphilus Yasumatsu, 1951 (Hymenoptera: Cynipidae) in Different Natural Populations of Castanea sativa Miller: An Experimental Ex Situ Test. Int. J. Pest Manag. 2019, 65, 147–153. [Google Scholar] [CrossRef]
- Maltoni, A.; Mariotti, B.; Jacobs, D.; Tani, A. Pruning Methods to Restore Castanea sativa Stands Attacked by Dryocosmus kuriphilus. New For. 2012, 43, 869–885. [Google Scholar] [CrossRef]
- Gehring, E.; Bellosi, B.; Quacchia, A.; Conedera, M. Assessing the Impact of Dryocosmus kuriphilus on the Chestnut Tree: Branch Architecture Matters. J. Pest Sci. 2018, 91, 189–202. [Google Scholar] [CrossRef]
- Kato, K.; Hijii, N. Effects of Gall Formation by Dryocosmus kuriphilus Yasumatsu (Hym., Cynipidae) on the Growth of Chestnut Trees. J. Appl. Entomol. 1997, 121, 9–15. [Google Scholar] [CrossRef]
- Battisti, A.; Benvegnù, I.; Colombari, F.; Haack, R.A. Invasion by the Chestnut Gall Wasp in Italy Causes Significant Yield Loss in Castanea sativa Nut Production. Agric. For. Entomol. 2014, 16, 75–79. [Google Scholar] [CrossRef]
- Gehring, E.; Kast, C.; Kilchenmann, V.; Bieri, K.; Gehrig, R.; Pezzatti, G.B.; Conedera, M. Impact of the Asian Chestnut Gall Wasp, Dryocosmus kuriphilus (Hymenoptera, Cynipidae), on the Chestnut Component of Honey in the Southern Swiss Alps. J. Econ. Entomol. 2018, 111, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Marcolin, E.; Pividori, M.; Colombari, F.; Manetti, M.C.; Pelleri, F.; Conedera, M.; Gehring, E. Impact of the Asian Gall Wasp Dryocosmus kuriphilus on the Radial Growth of the European Chestnut Castanea sativa. J. Appl. Ecol. 2021, 58, 1212–1224. [Google Scholar] [CrossRef]
- Nieves-Aldrey, J.L.; Gil-Tapetado, D.; Gavira, O.; Boyero, J.R.; Polidori, C.; Lombardero, M.J.; Blanco, D.; Rey del castillo, C.; Rodriguez Rojo, P.; Vela, J.M.; et al. Torymus Sinensis Kamijo, a Biocontrol Agent against the Invasive Chestnut Gall Wasp Dryocosmus kuriphilus Yasumatsu in Spain: Its Natural Dispersal from France and the First Data on Establishment after Experimental Releases. For. Syst. 2019, 28, e001. [Google Scholar] [CrossRef]
- Cheshkova, A.F. A Review of Hyperspectral Image Analysis Techniques for Plant Disease Detection and Identif Ication. Vavilov J. Genet. Breed. 2022, 26, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Terentev, A.; Dolzhenko, V.; Fedotov, A.; Eremenko, D. Current State of Hyperspectral Remote Sensing for Early Plant Disease Detection: A Review. Sensors 2022, 22, 757. [Google Scholar] [CrossRef]
- Junges, A.H.; Almança, M.A.K.; Fajardo, T.V.M.; Ducati, J.R. Leaf Hyperspectral Reflectance as a Potential Tool to Detect Diseases Associated with Vineyard Decline. Trop. Plant Pathol. 2020, 45, 522–533. [Google Scholar] [CrossRef]
- Nguyen, C.; Sagan, V.; Maimaitiyiming, M.; Maimaitijiang, M.; Bhadra, S.; Kwasniewski, M.T. Early Detection of Plant Viral Disease Using Hyperspectral Imaging and Deep Learning. Sensors 2021, 21, 742. [Google Scholar] [CrossRef]
- Cen, Y.; Huang, Y.; Hu, S.; Zhang, L.; Zhang, J. Early Detection of Bacterial Wilt in Tomato with Portable Hyperspectral Spectrometer. Remote Sens. 2022, 14, 2882. [Google Scholar] [CrossRef]
- Pádua, L.; Marques, P.; Martins, L.; Sousa, A.; Peres, E.; Sousa, J.J. Monitoring of Chestnut Trees Using Machine Learning Techniques Applied to UAV-Based Multispectral Data. Remote Sens. 2020, 12, 3032. [Google Scholar] [CrossRef]
- Castedo-Dorado, F.; Álvarez-Álvarez, P.; Cuenca Valera, B.; Lombardero, M.J. Local-Scale Dispersal Patterns and Susceptibility to Dryocosmus kuriphilus in Different Castanea Species and Hybrid Clones: Insights from a Field Trial. New For. 2021, 54, 9–28. [Google Scholar] [CrossRef]
- Gil-Tapetado, D.; Castedo-Dorado, F.; Lombardero, M.J.; Martel, J.; Álvarez-Álvarez, P. Spatial Propagation and Patterns of Abundance of Dryocosmus kuriphilus throughout an Invaded Region. J. Appl. Entomol. 2021, 145, 10–25. [Google Scholar] [CrossRef]
- Bertoldi, D.; Miorelli, P.; Pedrazzoli, F.; Delugan, S.; Deromedi, M.; Maresi, G. Investigations on Yellowing of Chestnut Crowns in Trentino (Alps, Northern Italy). IForest 2020, 13, 466–472. [Google Scholar] [CrossRef]
- Gehring, E.; Bellosi, B.; Reynaud, N.; Conedera, M. Chestnut Tree Damage Evolution Due to Dryocosmus kuriphilus Attacks. J. Pest Sci. 2020, 93, 103–115. [Google Scholar] [CrossRef]
- Barreto, A.; Paulus, S.; Varrelmann, M.; Mahlein, A.-K. Hyperspectral Imaging of Symptoms Induced by Rhizoctonia Solani in Sugar Beet: Comparison of Input Data and Different Machine Learning Algorithms. J. Plant Dis. Prot. 2020, 127, 441–451. [Google Scholar] [CrossRef]
- Fallon, B.; Yang, A.; Lapadat, C.; Armour, I.; Juzwik, J.; Montgomery, R.A.; Cavender-Bares, J. Spectral Differentiation of Oak Wilt from Foliar Fungal Disease and Drought Is Correlated with Physiological Changes. Tree Physiol. 2020, 40, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Pithan, P.A.; Ducati, J.R.; Garrido, L.R.; Arruda, D.C.; Thum, A.B.; Hoff, R. Spectral Characterization of Fungal Diseases Downy Mildew, Powdery Mildew, Black-Foot and Petri Disease on Vitis Vinifera Leaves. Int. J. Remote Sens. 2021, 42, 5680–5697. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, H.; Ruan, C.; Huang, W.; Hu, T.; Zhao, J. Detection of Scab in Wheat Ears Using in Situ Hyperspectral Data and Support Vector Machine Optimized by Genetic Algorithm. Int. J. Agric. Biol. Eng. 2020, 13, 182–188. [Google Scholar] [CrossRef]
- Kotobuki, K.; Mori, K.; Sato, Y. Two Methods to Estimate the Tree Damage by Chestnut Gall Wasp, Dryocosmus kuriphilus Yasumatsu. Bull. Fruit Tree Res. Stn. Ser. A. Hiratsuka 1985, 12, 29–36. [Google Scholar]
- ASD Inc. FieldSpec 4 User Guide. Available online: https://www.malvernpanalytical.com/en/learn/knowledge-center/user-manuals/fieldspec-4-user-guide (accessed on 27 November 2022).
- Zahir, S.A.D.M.; Omar, A.F.; Jamlos, M.F.; Azmi, M.A.M.; Muncan, J. A Review of Visible and Near-Infrared (Vis-NIR) Spectroscopy Application in Plant Stress Detection. Sens. Actuators Phys. 2022, 338, 113468. [Google Scholar] [CrossRef]
- Gitelson, A.; Solovchenko, A. Non-Invasive Quantification of Foliar Pigments: Possibilities and Limitations of Reflectance- and Absorbance-Based Approaches. J. Photochem. Photobiol. B 2018, 178, 537–544. [Google Scholar] [CrossRef]
- Lee, L.C.; Liong, C.-Y.; Jemain, A.A. Partial Least Squares-Discriminant Analysis (PLS-DA) for Classification of High-Dimensional (HD) Data: A Review of Contemporary Practice Strategies and Knowledge Gaps. Analyst 2018, 143, 3526–3539. [Google Scholar] [CrossRef] [PubMed]
- Appeltans, S.; Pieters, J.G.; Mouazen, A.M. Potential of Laboratory Hyperspectral Data for In-Field Detection of Phytophthora Infestans on Potato. Precis. Agric. 2022, 23, 876–893. [Google Scholar] [CrossRef]
- Ramos, A.P.M.; Gomes, F.D.G.; Pinheiro, M.M.F.; Furuya, D.E.G.; Gonçalvez, W.N.; Junior, J.M.; Michereff, M.F.F.; Blassioli-Moraes, M.C.; Borges, M.; Alaumann, R.A.; et al. Detecting the Attack of the Fall Armyworm (Spodoptera Frugiperda) in Cotton Plants with Machine Learning and Spectral Measurements. Precis. Agric. 2022, 23, 470–491. [Google Scholar] [CrossRef]
- Hornero, A.; Hernández-Clemente, R.; North, P.R.J.; Beck, P.S.A.; Boscia, D.; Navas-Cortes, J.A.; Zarco-Tejada, P.J. Monitoring the Incidence of Xylella Fastidiosa Infection in Olive Orchards Using Ground-Based Evaluations, Airborne Imaging Spectroscopy and Sentinel-2 Time Series through 3-D Radiative Transfer Modelling. Remote Sens. Environ. 2020, 236, 111480. [Google Scholar] [CrossRef]
- Lin, Q.; Huang, H.; Wang, J.; Huang, K.; Liu, Y. Detection of Pine Shoot Beetle (PSB) Stress on Pine Forests at Individual Tree Level Using UAV-Based Hyperspectral Imagery and Lidar. Remote Sens. 2019, 11, 2540. [Google Scholar] [CrossRef]
- Wu, W.; Zhang, Z.; Zheng, L.; Han, C.; Wang, X.; Xu, J.; Wang, X. Research Progress on the Early Monitoring of Pine Wilt Disease Using Hyperspectral Techniques. Sensors 2020, 20, 3729. [Google Scholar] [CrossRef]
- Xi, G.; Huang, X.; Xie, Y.; Gang, B.; Bao, Y.; Dashzebeg, G.; Nanzad, T.; Dorjsuren, A.; Enkhnasan, D.; Ariunaa, M. Detection of Larch Forest Stress from Jas’s Larch Inchworm (Erannis Jacobsoni Djak) Attack Using Hyperspectral Remote Sensing. Remote Sens. 2022, 14, 124. [Google Scholar] [CrossRef]
- Ahamed, T. Big Data Scheme from Remote Sensing Applications: Concluding Notes for Agriculture and Forestry Applications. In Remote Sensing Application: Regional Perspectives in Agriculture and Forestry; Ahamed, T., Ed.; New Frontiers in Regional Science: Asian Perspectives; Springer Nature: Singapore, 2022; pp. 351–361. ISBN 978-981-19021-3-0. [Google Scholar]
- Duarte, A.; Borralho, N.; Cabral, P.; Caetano, M. Recent Advances in Forest Insect Pests and Diseases Monitoring Using UAV-Based Data: A Systematic Review. Forests 2022, 13, 911. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Coordinates | Castanea sativa Variety | Irrigation System | Spacing | Trees, n |
|---|---|---|---|---|---|
| A | 42°42′27.65″ N 6°26′13.54″ W | Parede roja | Drip | 8.5 m × 8.5 m | 52 |
| B | 42°42′35.5″ N 6°26′5.28″ W | Manual | 9 m × 9 m | 31 |
| Infestation Levels | Plot A (n = 211) | Plot B (n = 123) |
|---|---|---|
| None (0%) | 17 | 3 |
| Low (≤30) | 30 | 25 |
| Moderate (>30 and ≤40) | 23 | 23 |
| High (>40 and ≤60) | 79 | 39 |
| Very high (>60) | 62 | 33 |
| Infestation Levels | Plot A Validation Set | Plot B Training Set |
|---|---|---|
| None (0%) | 11 | 10 |
| Low (≤30) | 10 | 9 |
| Moderate (>30 and ≤40) | 10 | 10 |
| High (>40 and ≤60) | 10 | 10 |
| Very high (>60) | 11 | 10 |
| Spectral Dataset | Wavelengths Included |
|---|---|
| All spectra | [450:2500 nm] |
| ±10 nm intervals | [555:565 nm], [665:685 nm], [1395:1405 nm], [1930:1940 nm] |
| Four wavelengths | (560 nm, 680 nm, 1400 nm, 1935 nm) |
| Method | Set of Variables | Reflectance Overall Accuracy (%) | Transformed Values Overall Accuracy (%) |
|---|---|---|---|
| PLS-DA | all the spectra | 59.62 | 57.69 |
| ±10 nm intervals | 53.85 | 59.62 | |
| 4 wavelengths | 53.85 | 59.62 | |
| RF | all the spectra | 51.92 | 50 |
| ±10 nm intervals | 67.31 | 69.23 | |
| 4 wavelengths | 67.31 | 69.23 |
| Reference Values | ||||||
| N | L | M | H | VH | ||
| Predicted values | N | 4 | 1 | 1 | 0 | 5 |
| L | 4 | 1 | 1 | 1 | 3 | |
| M | 5 | 1 | 2 | 0 | 2 | |
| H | 3 | 3 | 1 | 1 | 2 | |
| VH | 3 | 0 | 0 | 2 | 6 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira-Obaya, D.; Castedo-Dorado, F.; Sanz-Ablanedo, E.; Mejía-Correal, K.B.; Rodríguez-Pérez, J.R. Leaf Trait Hyperspectral Characterization of Castanea sativa Miller Affected by Dryocosmus kuriphilus Yasumatsu. Agronomy 2023, 13, 923. https://doi.org/10.3390/agronomy13030923
Pereira-Obaya D, Castedo-Dorado F, Sanz-Ablanedo E, Mejía-Correal KB, Rodríguez-Pérez JR. Leaf Trait Hyperspectral Characterization of Castanea sativa Miller Affected by Dryocosmus kuriphilus Yasumatsu. Agronomy. 2023; 13(3):923. https://doi.org/10.3390/agronomy13030923
Chicago/Turabian StylePereira-Obaya, Dimas, Fernando Castedo-Dorado, Enoc Sanz-Ablanedo, Karen Brigitte Mejía-Correal, and José Ramón Rodríguez-Pérez. 2023. "Leaf Trait Hyperspectral Characterization of Castanea sativa Miller Affected by Dryocosmus kuriphilus Yasumatsu" Agronomy 13, no. 3: 923. https://doi.org/10.3390/agronomy13030923
APA StylePereira-Obaya, D., Castedo-Dorado, F., Sanz-Ablanedo, E., Mejía-Correal, K. B., & Rodríguez-Pérez, J. R. (2023). Leaf Trait Hyperspectral Characterization of Castanea sativa Miller Affected by Dryocosmus kuriphilus Yasumatsu. Agronomy, 13(3), 923. https://doi.org/10.3390/agronomy13030923