


Effects of Different Altitudes on Coffea arabica Rhizospheric Soil Chemical Properties and Soil Microbiota
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Rhizosphere Soil Samples Collection
2.2. Measurement of Rhizospheric Soil Physical-Chemical Characteristics
2.3. DNA Extraction and MiSeq Sequencing
2.4. Bioinformatic Analyses
2.5. Statistical Analyses
3. Results
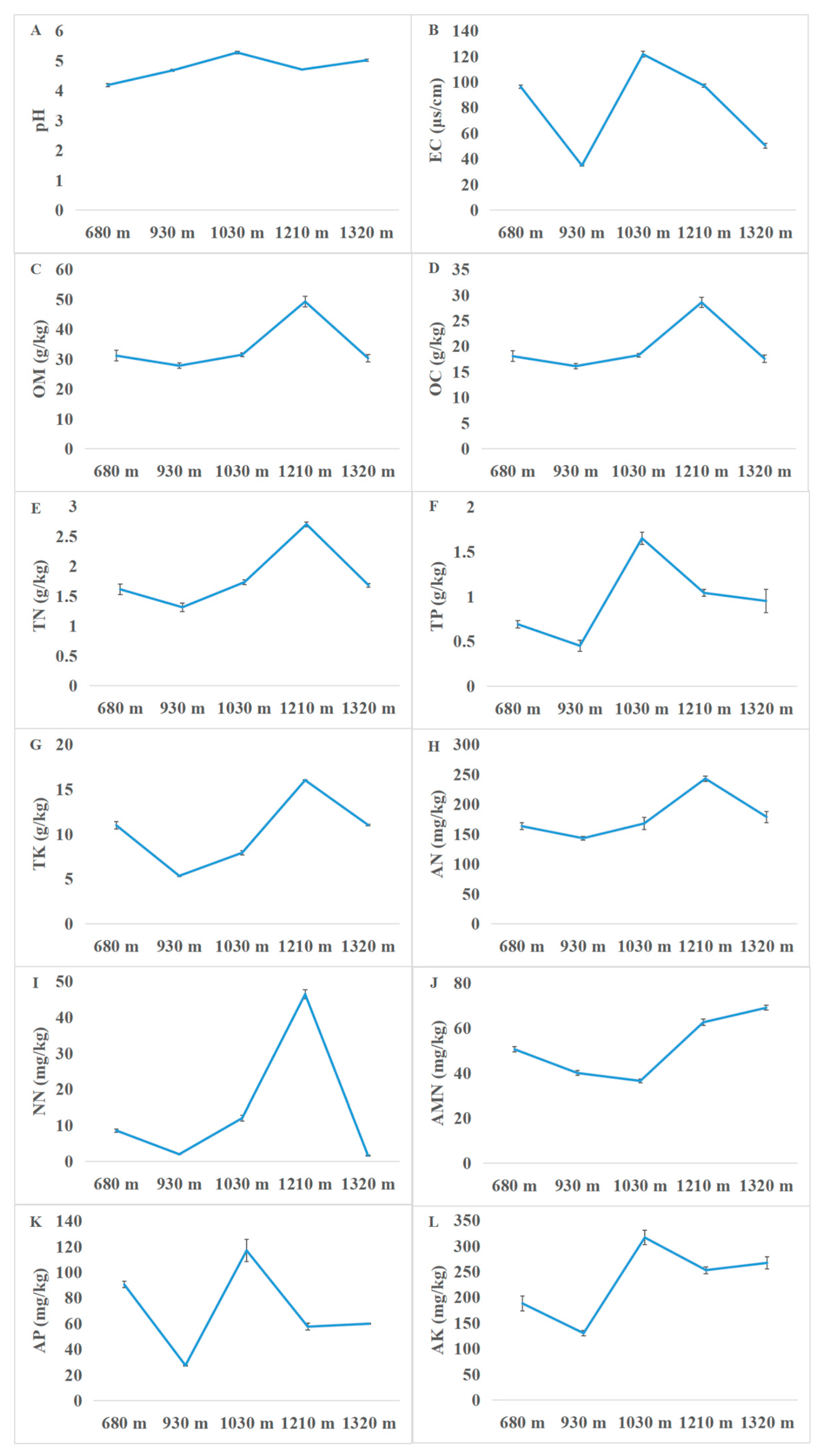
3.1. Soil Physicochemical Properties
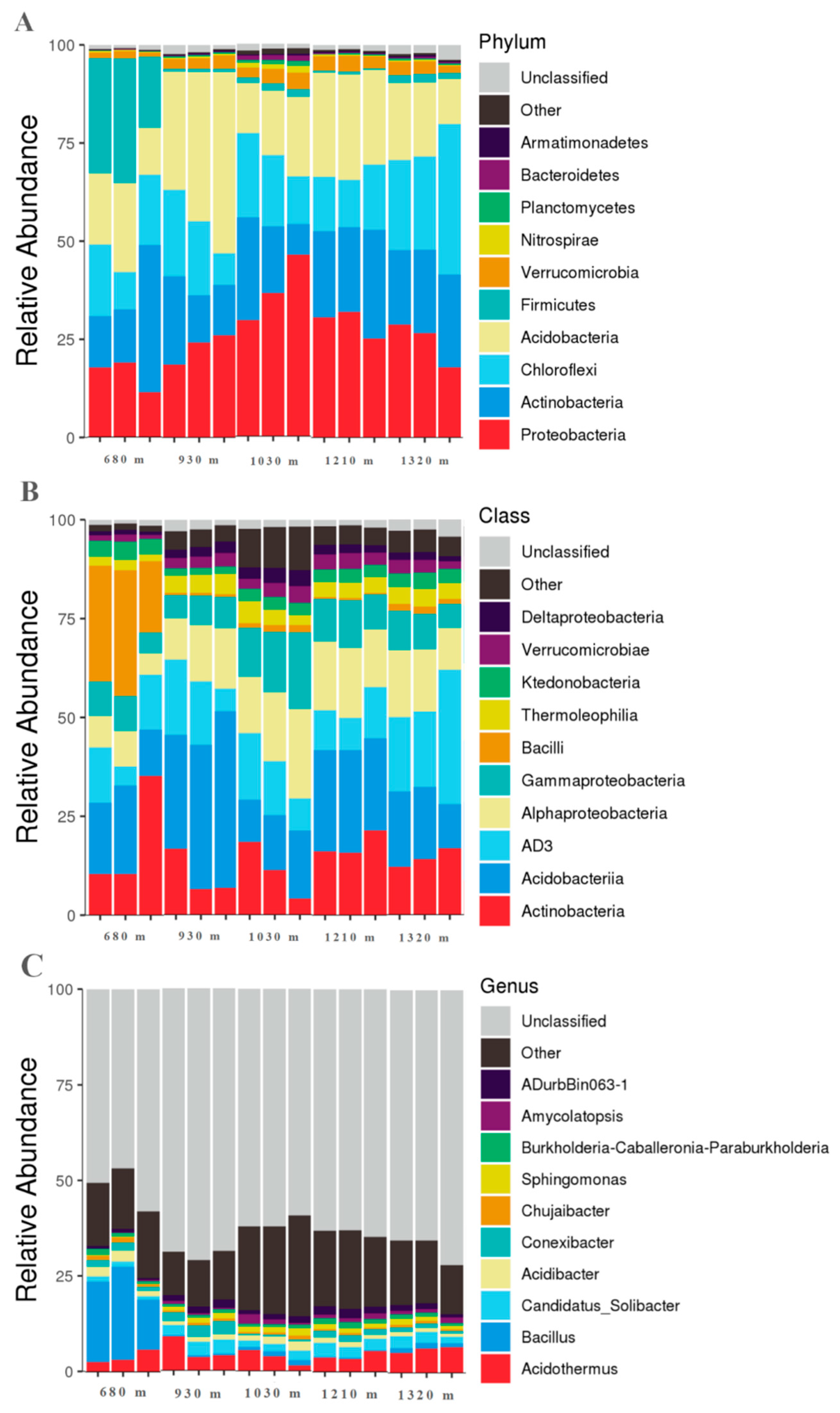
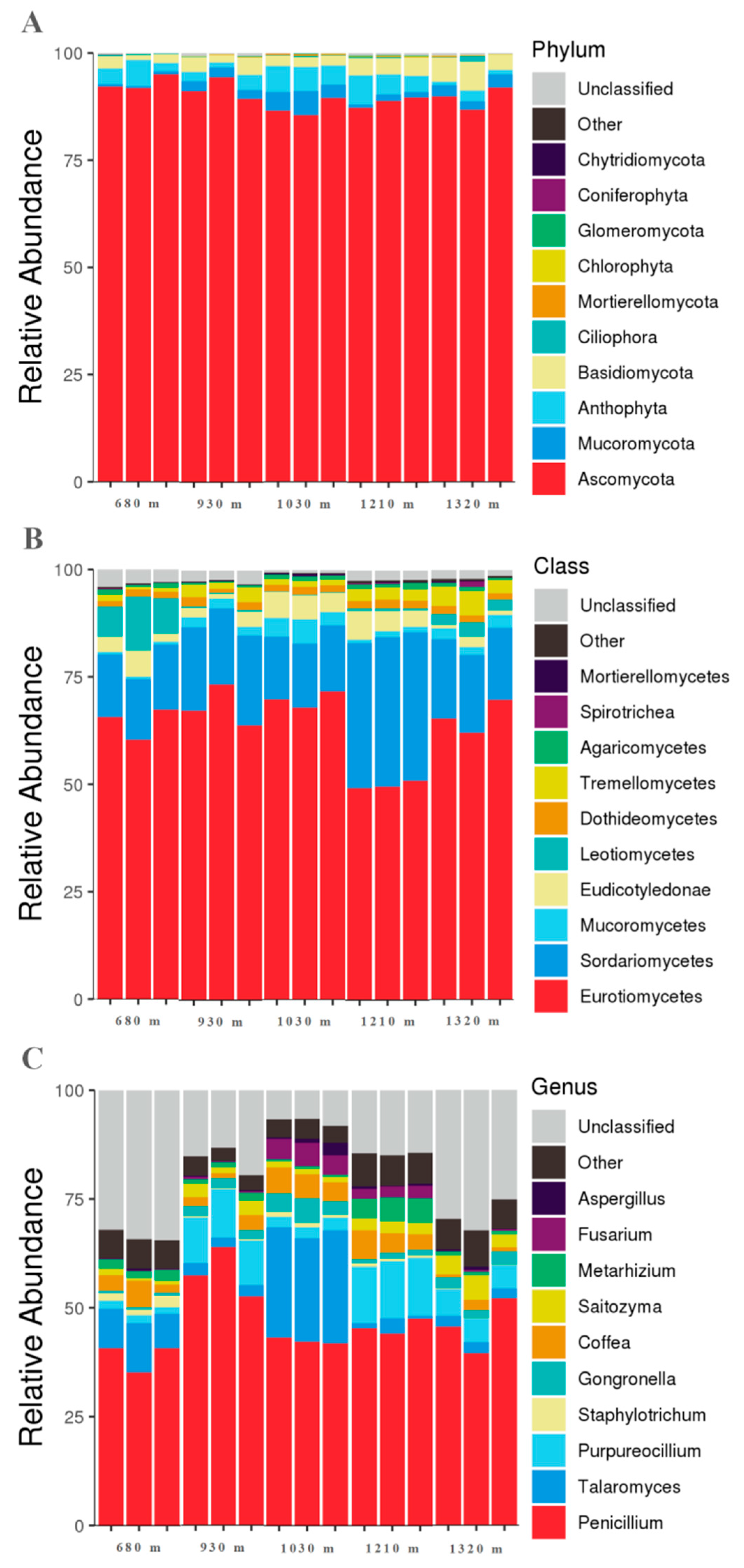
3.2. Microbial Community Characteristics
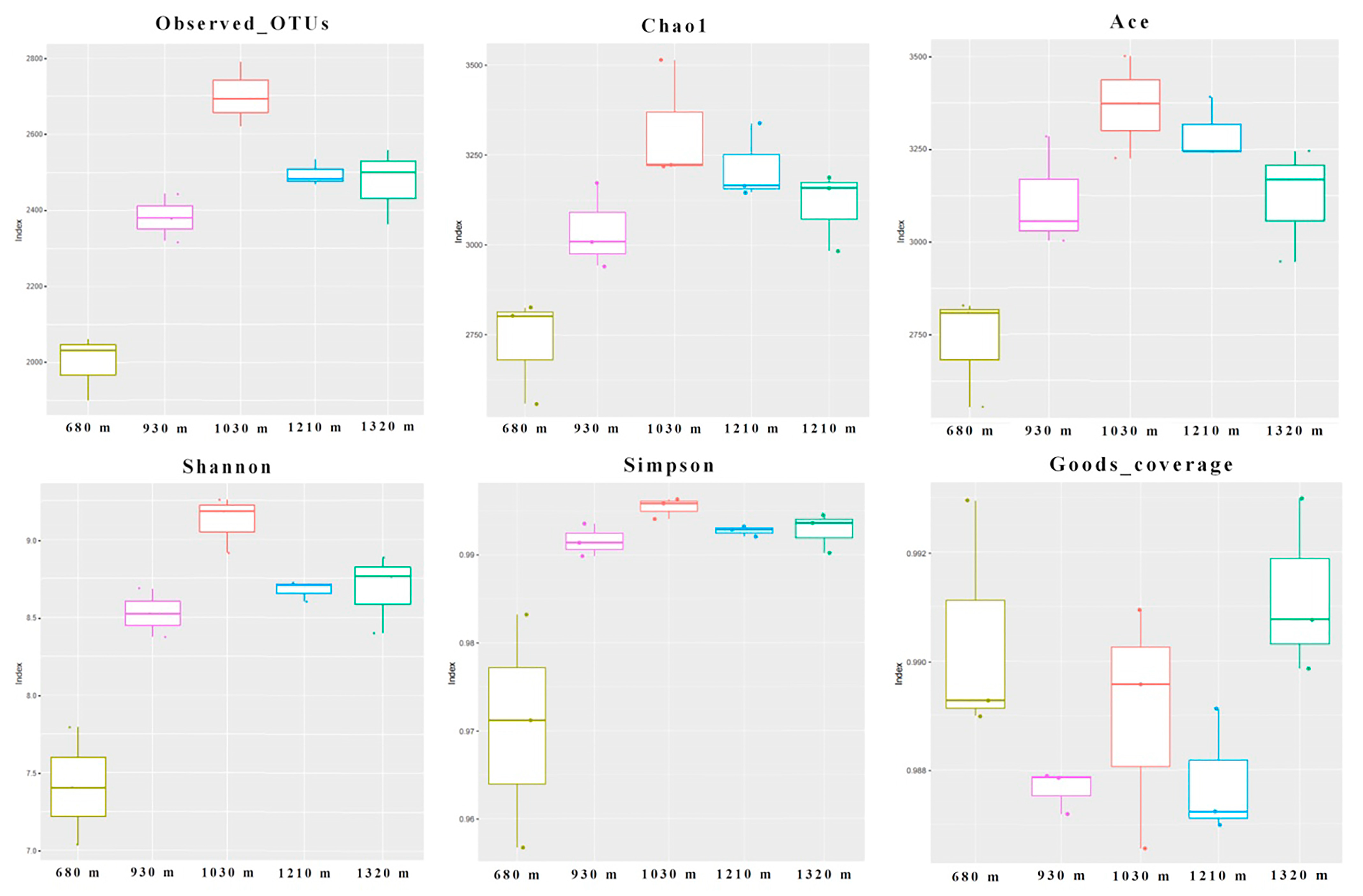
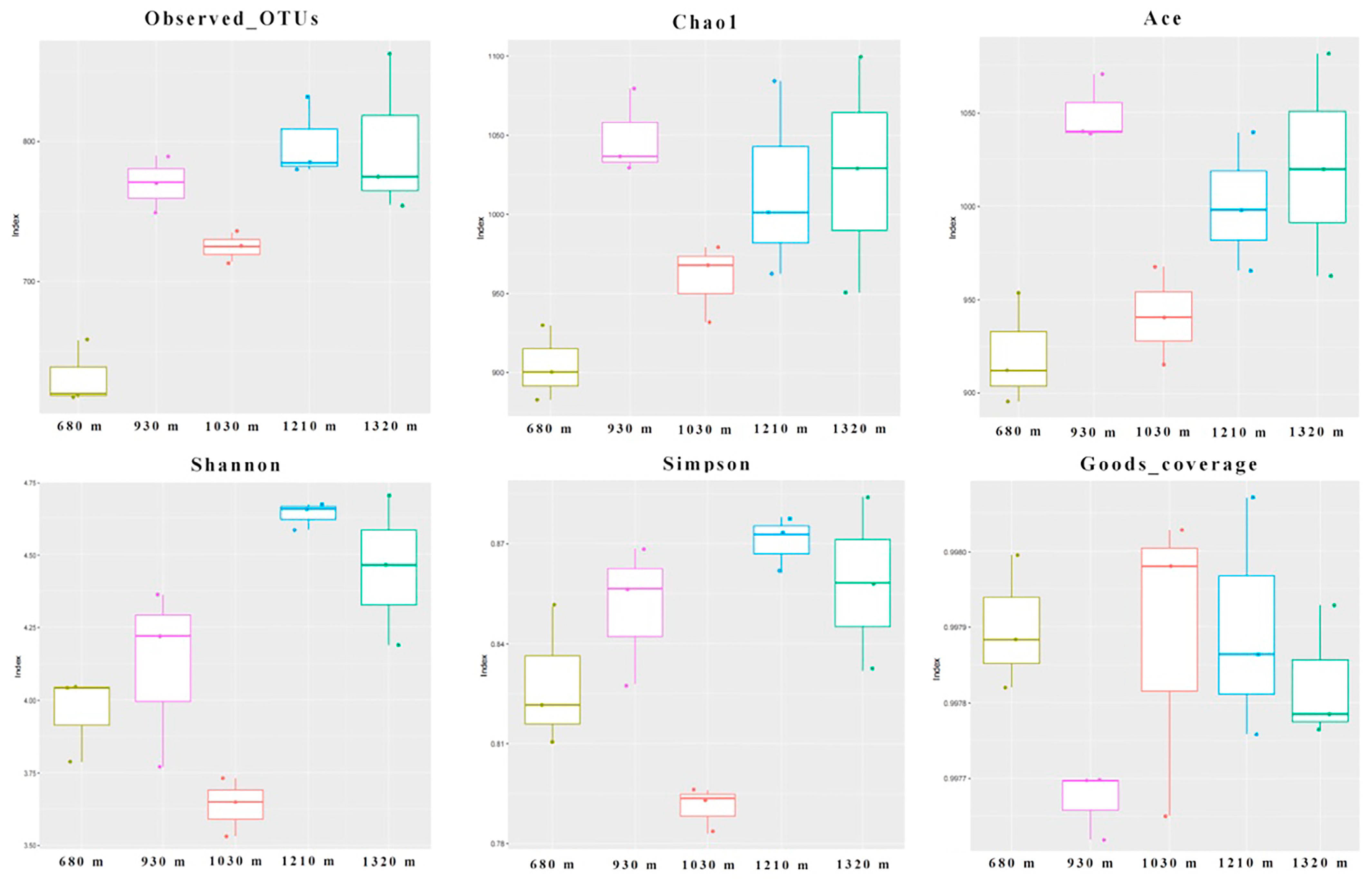
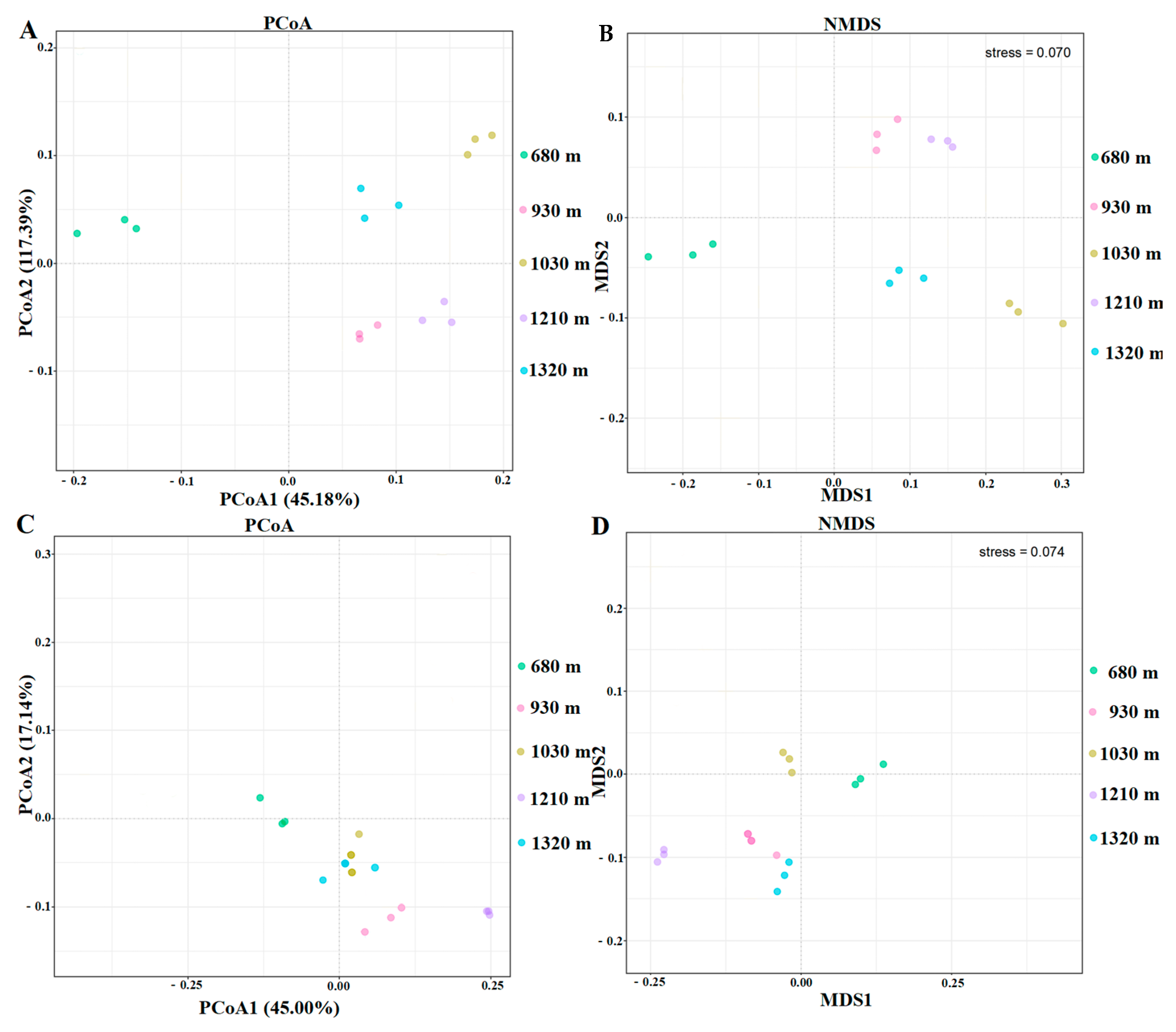
3.3. Microbial Diversity
3.3.1. Alpha Diversity Indices of the Microbial Communities
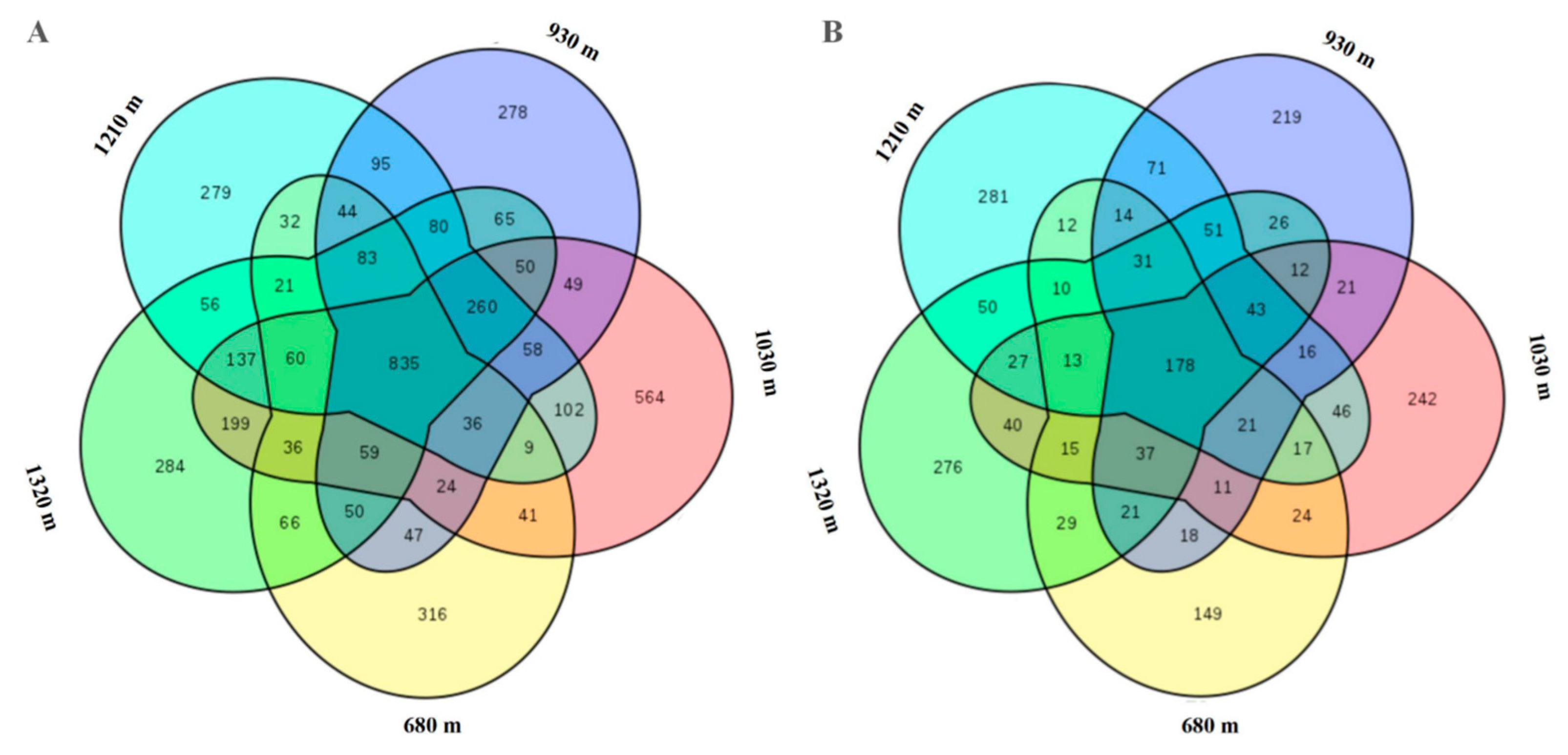
3.3.2. Beta Diversity Indices of the Microbial Communities
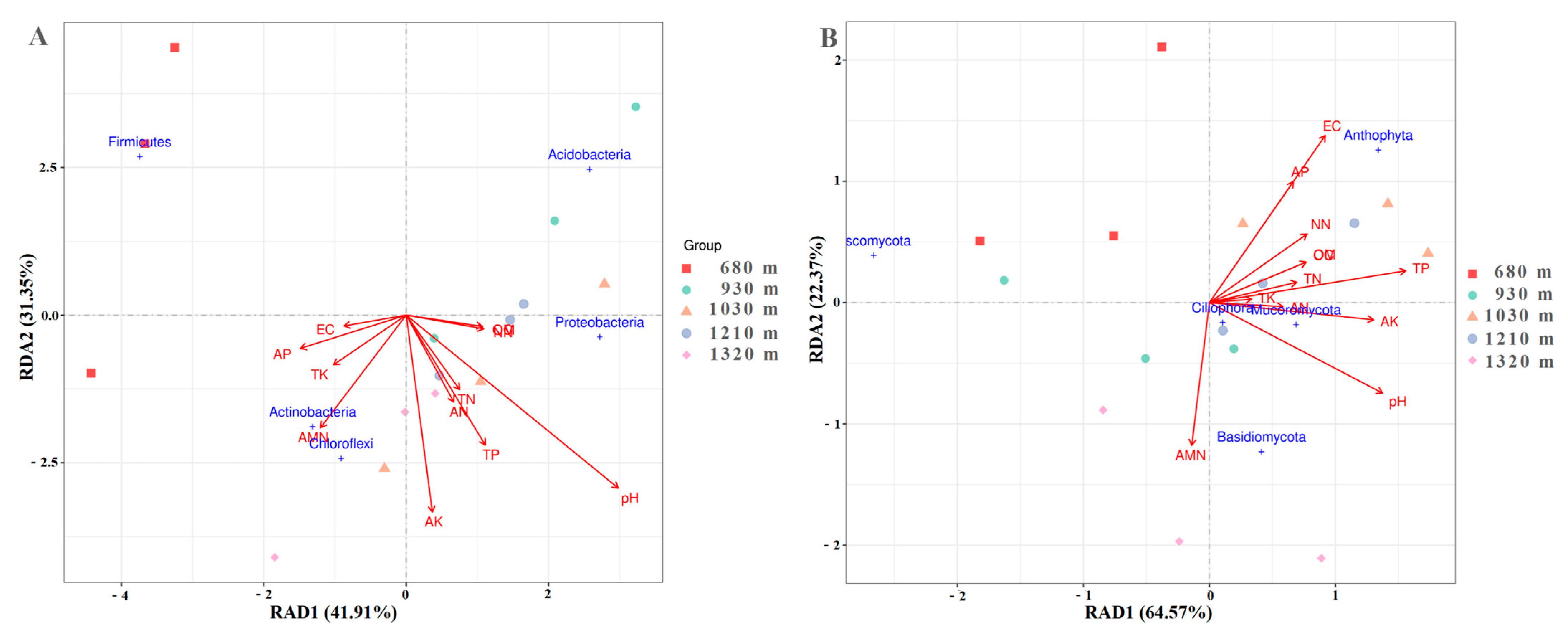
3.4. Effect of Environmental Factors on Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guimaraes, R.J.; Borem, F.M.; Shuler, J.; Farah, A.; Romero, J.C.P. Coffee growing and post-harvest processing. In Coffee Production, Quality and Chemistry; Farah, A., Ed.; The Royal Society of Chemistry: London, UK, 2019; pp. 150–178. [Google Scholar]
- DaMatta, F.M.; Ronchi, C.P.; Maestri, M.; Barros, R.S. Ecophysiology of coffee growth and production. Braz. J. Plant Physiol. 2007, 19, 485–510. [Google Scholar] [CrossRef]
- Yan, L.; Huang, L.F.; Wang, X.Y.; Zhou, H.; Cheng, J.H.; Li, J.H.; Long, Y.Z.; Dong, Y.P. Genetic diversity of coffee germplasms by ISSR markers. Chin. J. Trop. Crops 2019, 40, 300–307. (In Chinese) [Google Scholar]
- Martinez, S.J.; Bressani, A.P.P.; Simao, J.B.P.; Pylro, V.S.; Dias, D.R.; Schwan, R.F. Dominant microbial communities and biochemical profile of pulped natural fermented coffees growing in different altitudes. Food Res. Int. 2022, 159, 111605. [Google Scholar] [CrossRef]
- Mintesnot, A.; Dechassa, N. Effect of altitude, shade, and processing methods on the quality and biochemical composition of green coffee beans in Ethiopia. East Afr. J. Sci. 2018, 12, 87–100. [Google Scholar]
- Paudel, M.; Parajuli, K.; Regmi, S.; Budhathoki, S. Effect of altitude and shade on production and physical attributes of coffee in Gulmi, Syangja and Palpa districts of Nepal. J. Agric. Nat. Resour. 2021, 4, 222–238. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Mangueze, A.V.J.; Leitão, A.E.; Pais, I.P.; Moreira, R.; Campa, C.; Chiulele, R.; Reis, F.O.; Marques, I.; Scotti-Campos, P.; et al. Shade and altitude implications on the physical and chemical attributes of green coffee beans from gorongosa mountain, Mozambique. Agronomy 2022, 12, 2540. [Google Scholar] [CrossRef]
- Ferreira, S.D.; Oliveira, E.S.M.; Ribeiro, R.W.; Filete, A.C.; Castanheira, T.D.; Rocha, C.P.B.; Moreli, P.A.; Oliveira, C.S.E.; Guarçoni, C.R.; Partelli, F.L.; et al. Association of altitude and solar radiation to understand coffee quality. Agronomy 2022, 12, 1885. [Google Scholar] [CrossRef]
- Worku, M.; de Meulenaer, B.; Duchateau, L.; Boeckx, P. Effect of altitude on biochemical composition and quality of green arabica coffee beans can be affected by shade and postharvest processing method. Food Res. Int. 2018, 105, 278–285. [Google Scholar] [CrossRef]
- Kamal, B.K.; Acharya, B.; Srivastava, A.; Pandey, M. Effect of different altitudes in qualitative and quantitative attributes of green coffee beans (Coffea arabica) in Nepal. Int. J. Hortic. Agric. Food Sci. 2021, 5, 1–7. [Google Scholar]
- Bertrand, B.; Vaast, P.; Alpizar, E.; Etienne, H.; Davrieux, F.; Charmetant, P. Comparison of bean biochemical composition and beverage quality of Arabica hybrids involving Sudanese-Ethiopian origins with traditional varieties at various elevations in Central America. Tree Physiol. 2006, 26, 1239–1248. [Google Scholar] [CrossRef]
- Colmenares, P.C.H.; Silva, P.A.; Mogollón, O.A.M. Impacts of different coffee systems on soil microbial populations at different altitudes in Villavicencio (Colombia). Agron. Colomb. 2016, 34, 285–291. [Google Scholar] [CrossRef]
- Zhang, H.B.; Zhou, H.; Li, J.H.; Lin, X.W.; Bai, X.H.; Guo, T.Y.; Xiao, B.; Xia, H.Y.; Yang, J.Z. Growing of Coffea arabica in high altitudes in China. Chin. J. Trop. Agric. 2014, 34, 21–32. (In Chinese) [Google Scholar]
- Zhang, Y.; Cong, J.; Lu, H.; Li, G.; Qu, Y.; Su, X.; Zhou, J.; Li, D. Community structure and elevational diversity patterns of soil Acidobacteria. J. Environ. Sci. 2014, 26, 1717–1724. [Google Scholar] [CrossRef]
- Zhao, M.Z.; Wu, T.; Tang, J.; Ma, G.R.; Guo, T.Y.; Xiao, Z.W.; Su, L.L.; Zhou, H.; Bai, X.H. Status quo and variation of soil fertility in different altitude of Coffea arabica L. Chin. J. Trop. Crops 2019, 40, 629–637. (In Chinese) [Google Scholar]
- Wang, B.; Lu, Y.; Li, W.; He, S.; Lin, R.; Qu, P.; Chen, H.; Zhang, F.; Zhao, M.; Shi, X.; et al. Effects of the continuous cropping of Amomum villosum on rhizosphere soil physicochemical properties, enzyme activities, and microbial communities. Agronomy 2022, 12, 2548. [Google Scholar] [CrossRef]
- Pandey, A.; Palni, L.M.S. The rhizosphere effect in trees of the Indian central himalaya with special reference to altitude. Appl. Ecol. Environ. Res. 2007, 5, 93–102. [Google Scholar] [CrossRef]
- Evizal, R.; Tohari, I.D.; Prijambada, J.W.; Widianto, D. Soil bacterial diversity and productivity of coffee shade tree agro-ecosystem. J. Trop. Soils 2012, 17, 181–187. [Google Scholar] [CrossRef]
- Nigussie, A.; Kissi, E. The contribution of coffee agroecosystem to soil fertility in Southwestern Ethiopia. Afr. J. Agric. Res. 2012, 7, 74–81. [Google Scholar] [CrossRef]
- Haggar, J.P.; Barrios, M.; Bolanos, M.; Merlo, M.; Moraga, P.; Munguia, R.; Ponce, A.; Romero, S.; Soto, G.; Staver, C.; et al. Coffee agroecosystem performance under full sun, shade, conventional and organic management regimes in Central America. Agrofor. Syst. 2011, 82, 285–301. [Google Scholar] [CrossRef]
- Bagyaraj, D.J.; Thilagar, G.; Ravisha, C.; Kushalappa, C.G.; Krishnamurthy, K.N.; Vaast, P. Below ground microbial diversity as influenced by coffee agroforestry systems in the Western Ghats, India. Agric. Ecosyst. Environ. 2015, 202, 198–202. [Google Scholar] [CrossRef]
- Fierer, N.; McCain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.M.; Silman, M.R. Microbes do not follow the elevational diversity patterns of plants and animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Xiong, J.; Zhang, H.; Feng, Y.; Lina, X.; Li, X.; Liang, W.; Chu, H. Soil pH drives the spatial distribution of bacterial communities along elevation on Changbai Mountain. Soil Biol. Biochem. 2013, 57, 204–211. [Google Scholar] [CrossRef]
- Wang, J.T.; Cao, P.; Hu, H.W.; Li, J.; Han, L.L.; Zhang, L.M.; Zheng, Y.M.; He, J.Z. Altitudinal distribution patterns of soil bacterial and archaeal communities along Mt. Shegyla on the Tibetan Plateau. Microb. Ecol. 2015, 69, 135–145. [Google Scholar] [CrossRef] [PubMed]
- De Armas-Ricard, M.; Orlando, J.; Bustamante, R.; Carú, M. Microbial communities of bulk and Eschscholzia californica rhizosphere soils at two altitudes in Central Chile. J. Soil. Sci. Plant Nutr. 2016, 16, 1–13. [Google Scholar] [CrossRef]
- Cui, Y.X.; Bing, H.J.; Fang, L.C.; Wu, Y.H.; Yu, J.L.; Shen, J.T.; Jiang, M.; Wang, X.; Zhang, X.C. Diversity patterns of the rhizosphere and bulk soil microbial communities along an attitudinal gradient in an alpine ecosystem of the eastern Tibetan Plateau. Geoderma 2019, 338, 118–127. [Google Scholar] [CrossRef]
- Praeg, N.; Pauli, H.; Illmer, P. Microbial diversity in bulk and rhizosphere soil of Ranunculus glacialis along a high-alpine attitudinal gradient. Front. Microbiol. 2019, 10, 1429. [Google Scholar] [CrossRef]
- Wei, N.; Yue, X. Distribution of core root microbiota of Tibetan hulless barley along an attitudinal and geographical gradient in the Tibetan plateau. Microorganisms 2022, 10, 1737. [Google Scholar] [CrossRef]
- Bao, S.D. Analysis Method of Soil and Agricultural Chemistry, 3rd ed.; China Agricultural Press: Beijing, China, 2000; pp. 25–108. [Google Scholar]
- Logue, J.B.; Stedmon, C.A.; Kellerman, A.M.; Nielsen, N.J.; Andersson, A.F.; Laudon, H.; Lindstrom, E.S.; Kritzberg, E.S. Experimental insights into the importance of aquatic bacterial community composition to the degradation of dissolved organic matter. ISME J. 2016, 10, 533–545. [Google Scholar] [CrossRef]
- Karlsson, I.; Friberg, H.; Steinberg, C.; Persson, P. Fungicide effects on fungal community composition in the wheat phyllosphere. PloS ONE 2014, 9, e111786. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Matthew, R.D.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Author correction: Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 1091. [Google Scholar] [CrossRef]
- Joët, T.; Lafargue, A.; Descroix, F.; Doulbeau, S.; Bertrandkochko, B.A.; Dussert, S. Infuence of environmental factors, wet processing and their interactions on the biochemical composition of green Arabica cofee beans. Food Chem. 2010, 118, 693–701. [Google Scholar] [CrossRef]
- Ferreira, W.P.M.; Queiroz, D.M.; Silvac, S.A.; Tomaz, R.S.; Corrêa, P.C. Effects of the orientation of the mountainside, altitude and varieties on the quality of the coffee beverage from the “Matas de Minas” Region, Brazilian Southeast Retrieved from. Am. J. Plant Sci. 2016, 7, 1291–1303. [Google Scholar] [CrossRef]
- Tsegay, G.; Redi-Abshiro, M.; Chandravanshi, B.S.; Ele, E.; Mohammed, A.M.; Mamo, H. Effect of altitude of coffee plants on the composition of fatty acids of green coffee beans. BMC Chem. 2020, 14, 36. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, B.; Boulanger, R.; Dussert, S.; Ribeyre, F.; Berthiot, L.; Descroix, F.; Joët, T. Climatic factors directly impact the volatile organic compound fingerprint in green Arabica coffee bean as well as coffee beverage quality. Food Chem. 2012, 135, 2575–2583. [Google Scholar] [CrossRef]
- Davis, A.P.; Gole, T.W.; Baena, S.; Moat, J. The impact of climate change on indigenous Arabica coffee (Coffea arabica): Predicting future trends and identifying priorities. PLoS ONE 2012, 7, 10–14. [Google Scholar] [CrossRef]
- Schroth, G.; Läderach, P.; Blackburn Cuero, D.S.; Neilson, J.; Bunn, C. Winner or loser of climate change? A modeling study of current and future climatic suitability of Arabica coffee in Indonesia. Reg. Environ. Chang. 2015, 15, 1473–1482. [Google Scholar] [CrossRef]
- Tolessa, K.; D’heer, J.; Duchateau, L.; Boeckx, P. Influence of growing altitude, shade and harvest period on quality and biochemical composition of Ethiopian specialty coffee. J. Sci. Food Agric. 2017, 97, 2849–2857. [Google Scholar] [CrossRef]
- Rengel, Z. Soil pH, Soil Health and Climate Change. In Soil Health and Climate Change. Soil Biology; Singh, B., Cowie, A., Chan, K., Eds.; Springer: Berlin, Germany, 2011; pp. 50–58. [Google Scholar]
- Smit, E.; Leeflflang, P.; Gommans, S.; Broek, J.V.D.; van Mil, S.; Wernars, K. Diversity and seasonal fluctuations of the dominant members of the bacterial soil community in a wheat field as determined by cultivation and molecular methods. Appl. Environ. Microbiol. 2001, 67, 2284–2291. [Google Scholar] [CrossRef]
- Gao, Z.; Hu, Y.; Han, M.; Xu, J.; Wang, X.; Liu, L.; Tang, Z.; Jiao, W.; Jin, R.; Liu, M.; et al. Effects of continuous cropping of sweet potatoes on the bacterial community structure in rhizospheric soil. BMC Microbiol. 2021, 21, 102. [Google Scholar] [CrossRef]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Hu, C.; Yuan, Z. Soil metagenomics reveals effects of continuous sugarcane cropping on the structure and functional pathway of rhizospheric microbial community. Front. Microbiol. 2021, 12, 369. [Google Scholar] [CrossRef]
- Sikorski, J.; Baumgartner, V.; Birkhofer, K.; Boeddinghaus, R.S.; Bunk, B.; Fischer, M.; Fösel, B.U.; Friedrich, M.W.; Göker, M.; Hölzel, N.; et al. The evolution of ecological diversity in Acidobacteria. Front. Microbiol. 2022, 13, 715637. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.Z.; Li, L.; Wang1, X.L.; You, J.; Li, J.N.; Chen, X. Elevational is the main factor controlling the soil microbial community structure in alpine tundra of the Changbai Mountain. Sci. Rep. 2020, 10, 12442. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Cheng, J.; Jin, H.; Xu, Z.; Yang, X.; Min, D.; Xu, X.; Shao, X.; Lu, D.; Qin, B. Characterization of rhizosphere and endophytic microbial communities associated with Stipa purpurea and their correlation with soil environmental factors. Plants 2022, 11, 363. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, P.; Rodríguez–Echeverría, S.; González, L.; Freitas, H. Effect of invasive Acacia dealbata Link on soil microorganisms as determined by PCR-DGGE. Appl. Soil Ecol. 2010, 44, 245–251. [Google Scholar] [CrossRef]
- Min, L.; Mei, X.X.; Ying, P.; Juan, J.C.; Na, C. Characteristics of soil microbial community functional and structure diversity with coverage of Solidago canadensis L. J. Cent. South Univ. T. 2013, 20, 749–756. [Google Scholar]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Wang, J.; Soininen, J.; Zhang, Y.; Wang, B.; Yang, X.; Shen, J. Contrasting patterns in elevational diversity between microorganisms and macroorganisms. J. Biogeogr. 2011, 38, 595–603. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, H.; Cheng, H.; Chang, F.; Wan, Y.; She, X. Niche differentiation in the rhizosphere and endosphere fungal microbiome of wild Paris polyphylla Sm. Peer J. 2020, 8, e8510. [Google Scholar] [CrossRef]
- Chen, L.F.; Kong, J.Q.; He, Z.B.; Zhao, W.Z.; Song, M.D.; Li, Y.M.; Gao, Y.; Yang, S.P. A fundamental role of slope aspect and elevation in controlling diversity patterns of soil bacterial communities: Insights from an arid-montane ecosystem in China. J. Soil Sci. Plant Nut. 2022, 22, 3996–4007. [Google Scholar] [CrossRef]
- Singh, D.; Lee-Cruz, L.; Kim, W.S.; Kerfahi, D.; Chun, J.H.; Adams, J.M. Strong elevational trends in soil bacterial community composition on Mt. Halla. South Korea Soil Biol. Biochem. 2014, 68, 140–149. [Google Scholar] [CrossRef]
- Liu, D.; Wu, X.; Shi, S.; Liu, H.; Liu, G. A hollow bacterial diversity pattern with elevation in Wolong Nature Reserve, Western Sichuan Plateau. J. Soil Sediment 2016, 16, 2365–2374. [Google Scholar] [CrossRef]
- Wang, J.; Soininen, J.; He, J.; Shen, J. Phylogenetic clustering increases with elevation for microbes. Environ. Microbiol. Rep. 2012, 4, 217–226. [Google Scholar] [CrossRef]
- Körner, C. The use of “altitude” in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Donhauser, J.; Frey, B. Alpine soil microbial ecology in a changing world. FEMS Microbiol. Ecol. 2018, 94, fiy099. [Google Scholar] [CrossRef]
- Yuan, Y.; Si, G.; Wang, J.; Luo, T.; Zhang, G. Bacterial community in alpine grasslands along an altitudinal gradient on the Tibetan Plateau. FEMS Microbiol. Ecol. 2014, 87, 121–132. [Google Scholar] [CrossRef]
- Fierer, N.; Jackson, R. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–630. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef]
- Malard, L.A.; Anwar, M.Z.; Jacobsen, C.S.; Pearce, D.A. Biogeographical patterns in soil bacterial communities across the Arctic region. FEMS Microbiol. Ecol. 2019, 95, fz128. [Google Scholar] [CrossRef]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef]
- Martins, P.M.M.; Batista, N.N.; Miguel, M.G.; Simão, J.B.P.; Soares, J.R.; Schwan, R.F. Coffee growing altitude influences the microbiota, chemical compounds and the quality of fermented coffees. Food Res. Int. 2020, 129, 108872. [Google Scholar] [CrossRef] [PubMed]
- Galland, J.C.; Avelino, J.; Larraín, A. Origin coffees: Are appellations of origin on the horizon? In Coffee: Terroirs and Qualities, CIRAD; Montagnon, C., Ed.; Editions Quae: Versailles, Switzerland, 2006; p. 49. [Google Scholar]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of soil properties and microbial communities to agriculture: Implications for primary productivity and soil health indicators. Front. Plant Sci. 2016, 7, 990. [Google Scholar] [CrossRef] [PubMed]
- Franzluebbers, A.J. Stratification of soil porosity and organic matter. In Encyclopedia of Agrophysics; Horabik, J., Lipiec, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 858–861. [Google Scholar]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, Y.; Zhang, F.; Xie, C.; Qu, P.; Jiang, K.; Du, H.; Zhao, M.; Lu, Y.; Wang, B.; Shi, X.; et al. Effects of Different Altitudes on Coffea arabica Rhizospheric Soil Chemical Properties and Soil Microbiota. Agronomy 2023, 13, 471. https://doi.org/10.3390/agronomy13020471
Ge Y, Zhang F, Xie C, Qu P, Jiang K, Du H, Zhao M, Lu Y, Wang B, Shi X, et al. Effects of Different Altitudes on Coffea arabica Rhizospheric Soil Chemical Properties and Soil Microbiota. Agronomy. 2023; 13(2):471. https://doi.org/10.3390/agronomy13020471
Chicago/Turabian StyleGe, Yu, Fengying Zhang, Chun Xie, Peng Qu, Kuaile Jiang, Huabo Du, Meng Zhao, Yunfeng Lu, Butian Wang, Xuedong Shi, and et al. 2023. "Effects of Different Altitudes on Coffea arabica Rhizospheric Soil Chemical Properties and Soil Microbiota" Agronomy 13, no. 2: 471. https://doi.org/10.3390/agronomy13020471
APA StyleGe, Y., Zhang, F., Xie, C., Qu, P., Jiang, K., Du, H., Zhao, M., Lu, Y., Wang, B., Shi, X., Li, X., & Zhang, C. (2023). Effects of Different Altitudes on Coffea arabica Rhizospheric Soil Chemical Properties and Soil Microbiota. Agronomy, 13(2), 471. https://doi.org/10.3390/agronomy13020471