
Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
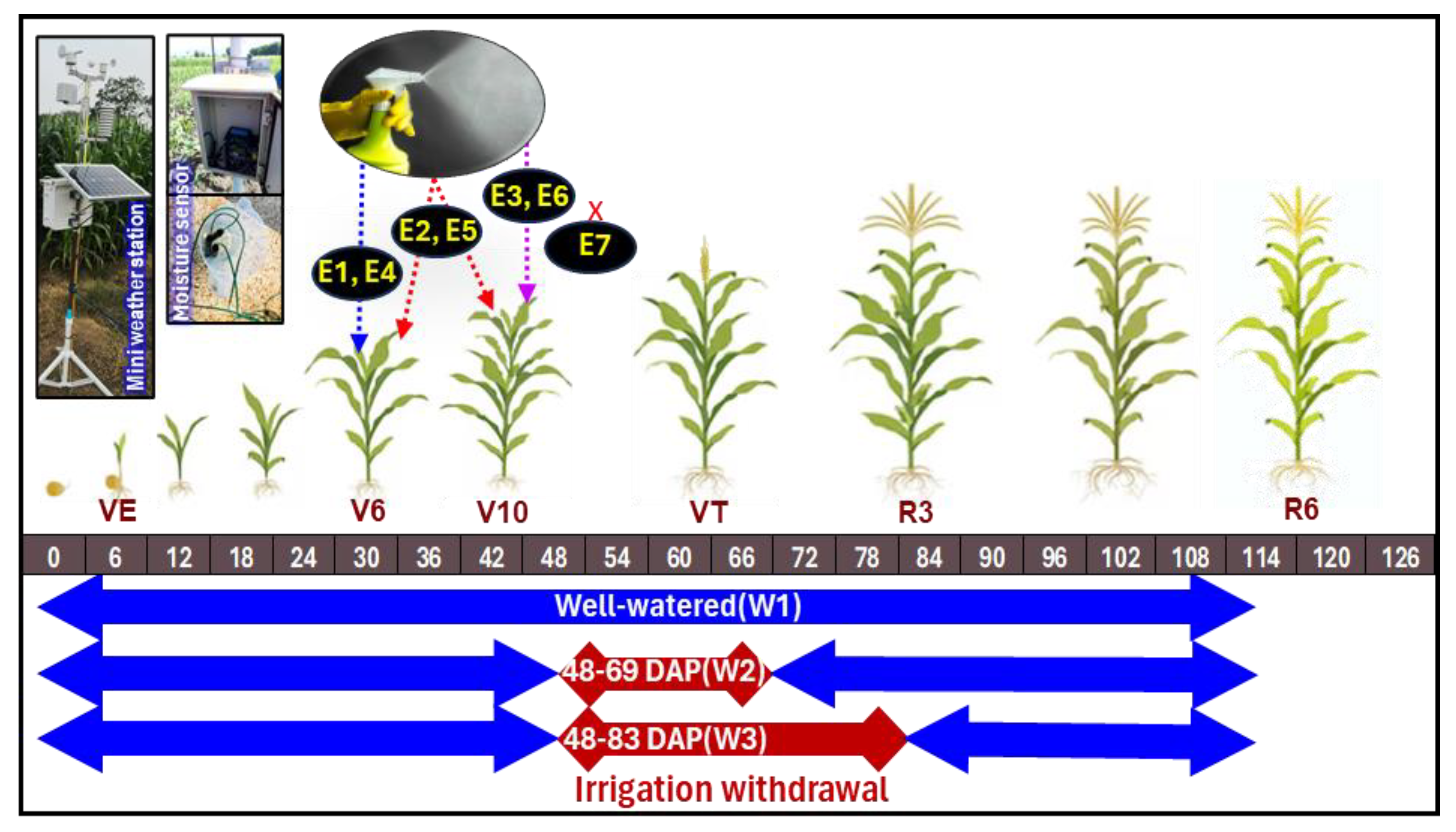
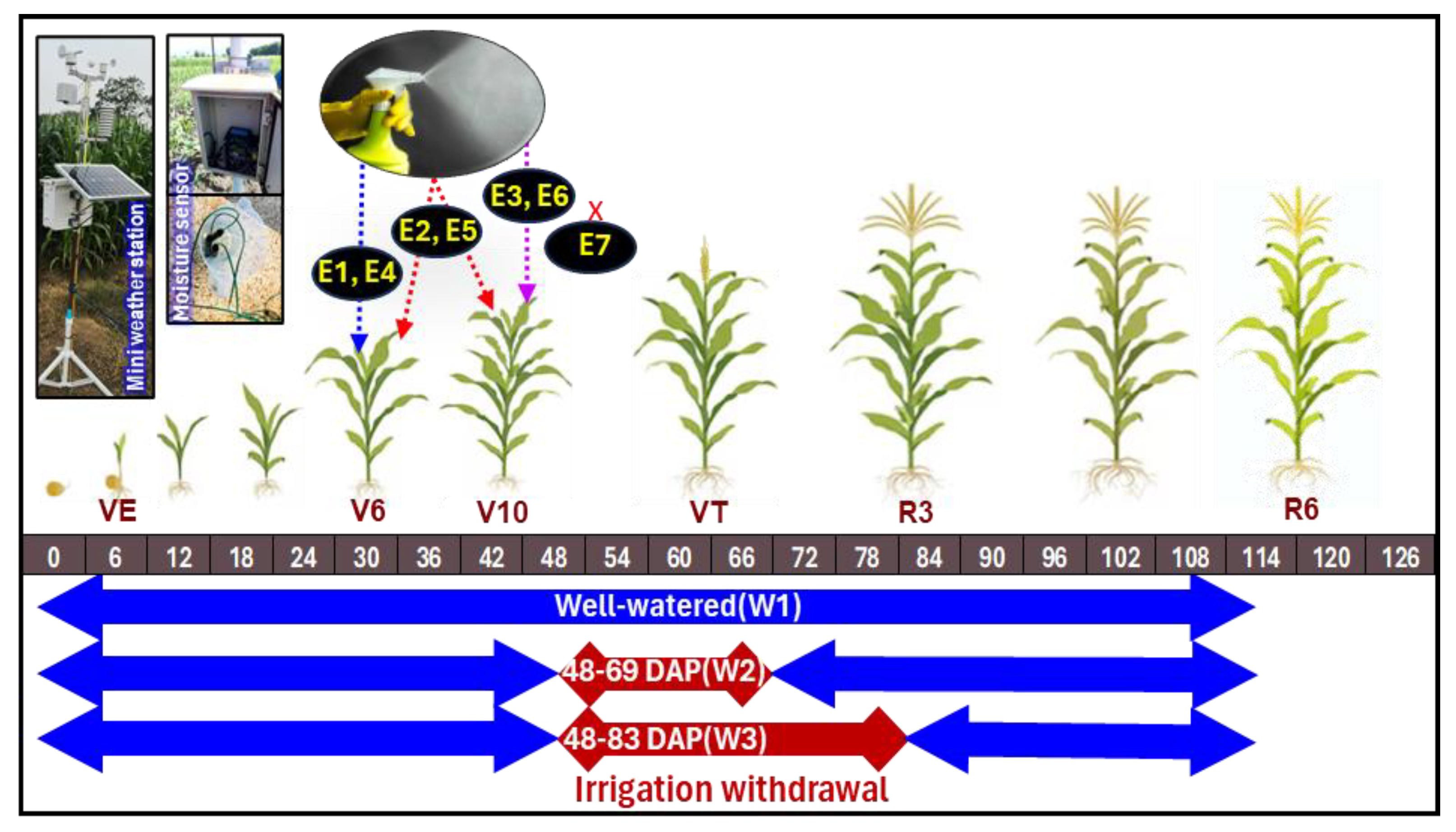
2.1. Experimental Location, Design, Treatments and Materials
2.2. Meteorological Information of the Experimental Location
2.3. Plant Sampling and Different Measurements
2.4. Statistical Analysis
3. Results
3.1. Environmental Conditions, Soil Moisture Tension and Soil Status
3.2. Treatment Effects on Morpho-Physio-Biochemical Traits of Maize
3.2.1. Morphological Traits of Maize
Plant Height (PH)
Stem Diameter (SD)
Fully Expanded Green Leaf Area (FEGLA) at V6, V10, and R3 and Leaf Area Index (LAI) at R3
Shoot Weight (SW)
Grain Number Per Plant (GNP)
Hundred-Grain Weight (100-GW)
Grain Yield (GY)
3.2.2. Physiological Traits of Maize
Relative Growth Rate (RGR)
Net Assimilation Rate (NAR)
Water Productivity (WP)
Accumulated Growing Degree Days (AGDD)
Heat Use Efficiency (HUE)
3.2.3. Physio-Biochemical Traits of Maize
SPAD Value for Leaf Greenness just after Water Stress Period (SPAD-JAWSP)
Relative Senescence Rate of Leaves at R3 (RSR-R3)
Relative Water Content of Leaves Just after Water Stress Period (RWC-JAWSP)
Electrolyte Leakage just after Water Stress Period (EL-JAWSP)
Proline Content in Leaf just after Water Stress Period (PrL-JAWSP)
Total Soluble Sugar in Leaf just after Water Stress Period (TSSL-JAWSP)
3.2.4. Correlation and Path Coefficient Analyses
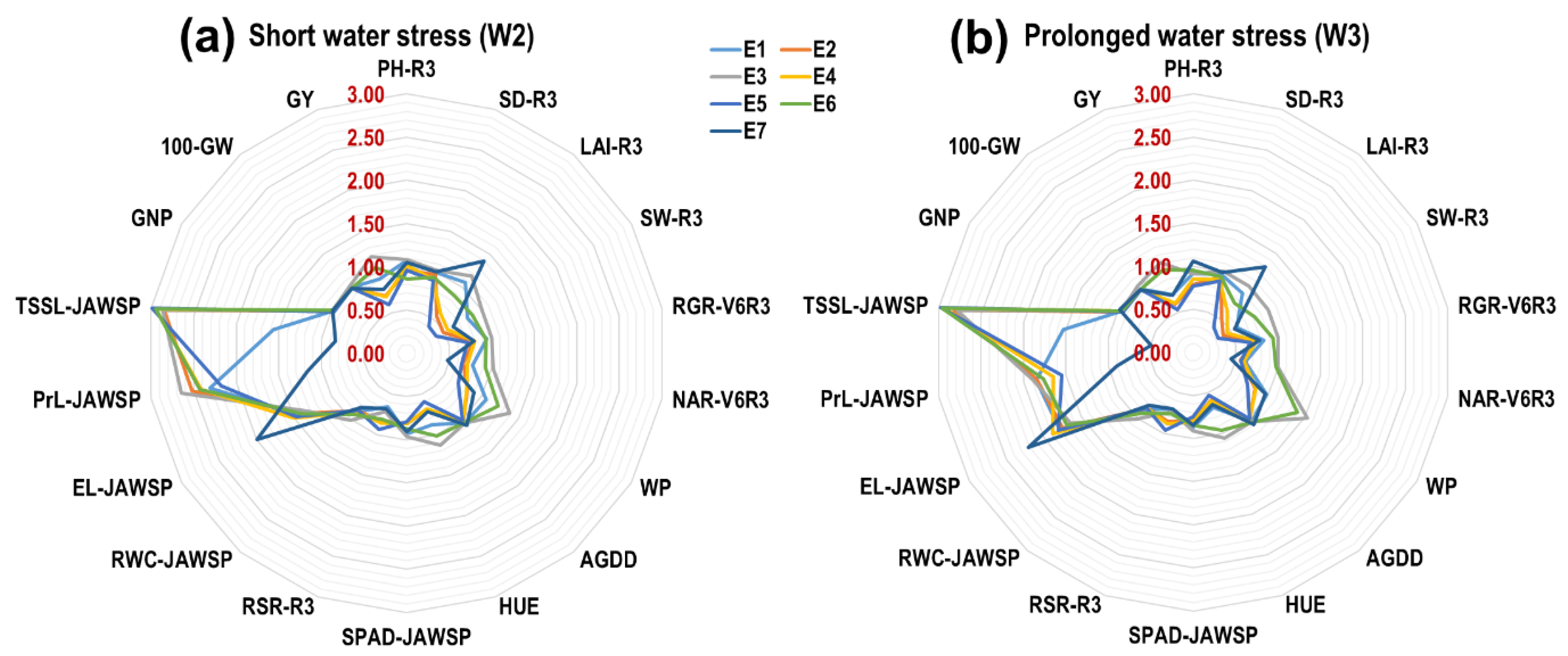
3.2.5. Drought Tolerance Index of Morpho-Physio-Biochemical Traits of Maize
3.3. Efficiency of Maize Production under Water Stress Using Ethephon
3.3.1. Energy Efficiency
3.3.2. Emission of CO2-eq
4. Discussion
4.1. Environmental Conditions and Soil Moisture Tension
4.2. Maize Morphological Performance under Water and Ethephon Applications
4.3. Maize Physiological Performance under Water and Ethephon Applications
4.4. Maize Physio-Biochemical Performance under Water and Ethephon Applications
4.5. Correlation and Path Coefficient Analysis
4.6. Drought Tolerance Index of Morpho-Physio-Biochemical Traits of Maize
4.7. Efficiency of Maize Production under Water Stress Using Ethephon
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roy, R.; Núñez-Delgado, A.; Sultana, S.; Wang, J.; Battaglia, M.L.; Sarker, T.; Seleiman, M.F.; Barmon, M.; Zhang, R. Additions of optimum water, spent mushroom compost and wood biochar to improve the growth performance of Althaea rosea in drought-prone coal-mined spoils. J. Environ. Manag. 2021, 295, 113076. [Google Scholar] [CrossRef] [PubMed]
- Hafez, E.M.; Seleiman, M.F. Response of barley quality traits, yield and antioxidant enzymes to water-stress and chemical inducers. Int. J. Plant Prod. 2017, 11, 477–490. [Google Scholar]
- Drought. World Health Organization. Available online: https://www.who.int/health-topics/drought?gclid=Cj0KCQjwoK2mBhDzAIsADGbjer80j_jPw6nNlUgg0h46VgY3u1LGtbq9mQYeFl4Tw7bqmYAl1Lio4EaAra4EALw_wcB#tab=tab_1 (accessed on 25 September 2023).
- Duan, W.; He, B.; Nover, D.; Fan, J.; Yang, G.; Chen, W.; Meng, H.; Liu, C. Floods and Associated Socioeconomic Damages in China over the Last Century. Nat. Hazards 2016, 82, 401–413. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Tropics. WIKIPEDIA. Available online: https://en.wikipedia.org/wiki/Tropics (accessed on 25 September 2023).
- How Much Land Is in the Tropics? GDP (God Plays Dice). Available online: http://godplaysdice.blogspot.com/2007/12/how-much-land-is-in-tropics.html (accessed on 25 September 2023).
- Tropics. National Geographic Encyclopedia. Available online: https://education.nationalgeographic.org/resource/tropics/ (accessed on 25 September 2023).
- Iglesias, A.; Garrote, L. Adaptation strategies for agricultural water management under climate change in Europe. Agric. Water Manag. 2015, 155, 113–124. [Google Scholar] [CrossRef]
- Caesar, J.; Alexander, L.V.; Trewin, B.; Tse-ring, K.; Sorany, L.; Vuniyayawa, V.; Keosavang, N.; Shimana, A.; Htay, M.M.; Karmacharya, J.; et al. Changes in Temperature and Precipitation Extremes over the Indo-Pacific Region from 1971 to 2005: Changes in Climate Extremes over the Indo-Pacific Region. Int. J. Climatol. 2011, 31, 791–801. [Google Scholar] [CrossRef]
- Villafuerte, M.Q.; Matsumoto, J. Significant Influences of Global Mean Temperature and ENSO on Extreme Rainfall in Southeast Asia. J. Clim. 2015, 28, 1905–1919. [Google Scholar] [CrossRef]
- Economic and Social Commission for Asia and the Pacific (ESCAP), United Nation. Ready for the Dry Years: Building Resilience to Drought in South-East Asia; United Nations Publication: Bangkok, Thailand, 2021; pp. 1–64. ISBN 978-92-1-120787-3. [Google Scholar]
- Zaki, M.K.; Noda, K.A. Systematic Review of Drought Indices in Tropical Southeast Asia. Atmosphere 2022, 13, 833. [Google Scholar] [CrossRef]
- Habib-ur-Rahman, M.; Ahmad, A.; Raza, A.; Hasnain, M.U.; Alharby, H.F.; Alzahrani, Y.M.; Bamagoos, A.A.; Hakeem, K.R.; Ahmad, S.; Nasim, W.; et al. Impact of climate change on agricultural production; Issues, challenges, and opportunities in Asia. Front. Plant Sci. 2022, 13, 925548. [Google Scholar] [CrossRef]
- Climate Risk Country Profile: Thailand. The World Bank Group and the Asian Development Bank. 2021. Available online: https://www.adb.org/sites/default/files/publication/722251/climate-risk-country-profile-thailand.pdf (accessed on 25 September 2023).
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Office of Agricultural Economics. Economic Data, Agricultural Product Forecasts. Available online: https://dl.parliament.go.th/backoffice/viewer2300/web/previewer.php (accessed on 14 November 2016).
- Hirich, A.; Fatnassi, H.; Ragab, R.; Choukr-Allah, R. Prediction of climate change impact on corn grown in the south of Morocco using the saltmed model. Irrig. Drain. 2016, 65, 9–18. [Google Scholar] [CrossRef]
- Khongdee, N.; Hilger, T.; Pansak, W.; Cadisch, G. Early planting and relay cropping: Pathways to cope with heat and drought. JARTS 2021, 122, 61–71. [Google Scholar] [CrossRef]
- Pradawet, C.; Khongdee, N.; Pansak, W.; Spreer, W.; Hilger, T.; Cadisch, G. Thermal imaging for assessment of maize water stress and yield prediction under drought conditions. J. Agron. Crop Sci. 2023, 209, 56–70. [Google Scholar] [CrossRef]
- Qi, Y.; Zhang, Q.; Hu, S.; Wang, R.; Wang, H.; Zhang, K.; Zhao, H.; Ren, S.; Yang, Y.; Zhao, F. Effects of High Temperature and Drought Stresses on Growth and Yield of Summer Maize during Grain Filling in North China. Agriculture 2022, 12, 1948. [Google Scholar] [CrossRef]
- Hama, B.M.; Mohammed, A.A. Physiological performance of maize (Zea mays L.) under stress conditions of water deficit and high temperature. Appl. Ecol. Environ. Res. 2019, 17, 1261–1278. [Google Scholar] [CrossRef]
- Saini, H.S.; Aspinall, D. Abnormal sporogenesis in wheat (Triticum aestivum L.) induced by short periods of high temperature. Ann. Bot. 1982, 49, 835–846. [Google Scholar] [CrossRef]
- Porter, J.R.; Gawith, M. Temperature and growth and development of wheat: A review. Eur. J. Agron. 1999, 10, 23–36. [Google Scholar] [CrossRef]
- Boyer, J.S.; Westgate, M.E. Grain yield with limited water. J. Exp. Bot. 2004, 55, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Britz, S.J.; Prasad, P.V.V.; Moreau, R.A.; Allen, L.H.D., Jr.; Kremer, F.; Boote, K.J. Influence of growth temperature on amounts of tocopherols, tocotrienols and gamma-ozyzanol in brown rice. J. Agric. Food Chem. 2007, 55, 7559–7565. [Google Scholar] [CrossRef]
- Sah, R.P.; Chakraborty, M.; Prasad, K.; Pandit, M.; Tudu, V.K.; Chakravarty, M.K.; Narayan, S.C.; Rana, M.; Moharana, D. Impact of water deficit stress in maize: Phenology and yield components. Sci. Rep. 2020, 10, 2944. [Google Scholar] [CrossRef]
- He, J.; Wen, R.; Tian, S.; Su, Y.; He, X.; Su, Y.; Cheng, W.; Huang, K.; Zhang, S. Effects of drought stress and re-watering on growth and yield of various maize varieties at tasseling stage. Agric. Sci. Technol. 2017, 18, 1145–1151. [Google Scholar]
- Ge, T.; Sui, F.; Bai, L.; Tong, C.; Sun, N. Effects of water stress on growth, biomass partitioning, and water use efficiency in summer maize (Zea mays L.) throughout the growth cycle. Acta Physiol. Plant. 2012, 34, 1043–1053. [Google Scholar] [CrossRef]
- Cakir, R. Effect of water stress at different development stages on vegetative and reproductive growth of corn. Field Crop. Res. 2004, 89, 1–16. [Google Scholar] [CrossRef]
- Chaves, M.M.; Marôco, J.P.; Pereira, J.S.; Chaves, M.M. Understanding plant responses to drought from genes to the whole plant. Funct. Plant Boil. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Zhang, Z.P.; Hua, Q.I.; Zhang, Y.; Sun, S.X.; Yang, G.H. Effects of water stress on photosynthetic rate and water use efficiency of maize. Acta Agric. Boreali Sin. 2009, 24, 155–158. [Google Scholar]
- Ge, T.; Sui, F.; Bai, L.; Lu, Y.-Y.; Zhou, G. Effects of Different Soil Water Content on the Photosynthetic Character and Pod Yields of Summer Maize. J. Shanghai Jiaotong Univ. 2005, 23, 143–147. [Google Scholar]
- Bu, L.; Zhang, R.; Chang, Y.; Xue, J.; Han, M. Response of photosynthetic characteristics to water stress of maize leaf in seeding. Acta Ecol. Sin. 2010, 30, 1184–1191. [Google Scholar]
- Chaves, M.M.; Pereira, J.S.; Maroco, J.; Rodrigues, M.L.; Ricardo, C.P.P.; Osório, M.L.; Carvalho, I.; Faria, T.; Pinheiro, C. How Plants Cope with Water Stress in the Field? Photosynthesis and Growth. Ann. Bot. 2002, 89, 907–916. [Google Scholar] [CrossRef]
- Chang, J.; Yang, D.; Tan, W.; Yueshang, L.U. Effects of water stress on maize leaf photosynthesis. J. Northeast Agric. Univ. 2008, 39, 1–5. [Google Scholar]
- Kumdee, O.; Molla, M.S.H.; Kanavittaya, K.; Romkaew, J.; Sarobol, E.; Nakasathien, S. Morpho-Physiological and Biochemical Responses of Maize Hybrids under Recurrent Water Stress at Early Vegetative Stage. Agriculture 2023, 13, 1795. [Google Scholar] [CrossRef]
- Cheng, M.; Wang, H.; Fan, J.; Zhang, F.; Wang, X. Effects of soil water deficit at different growth stages on maize growth, yield, and water use efficiency under alternate partial root-zone irrigation. Water 2021, 13, 148. [Google Scholar] [CrossRef]
- Sinay, H.; Karuwal, R.L. Proline and total soluble sugar content at the vegetative phase of six corn cultivars from Kisar Island Maluku, grown under drought stress conditions. Int. J. Adv. Agric. Res. 2014, 2, 77–82. [Google Scholar]
- Lei, L.; Shi, J.; Chen, J.; Zhang, M.; Sun, S.; Xie, S.; Li, X.; Zeng, B.; Peng, L.; Hauck, A. Ribosome profiling reveals dynamic translational landscape in maize seedlings under drought stress. Plant J. 2015, 84, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Bray, E.A. Plant responses to water deficit. Trends Plant Sci. 1997, 97, 1360–1385. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Shekoofa, A.; Emam, Y. Plant growth regulator (ethepon) alters maize (Zea mays L.) growth: Water use and grain yield under water stress. J. Agron. 2008, 7, 41–48. [Google Scholar] [CrossRef]
- Cox, W.J.; Andrade, H.F. Growth, yield, and yield components of maize as influenced by ethephon. Crop Sci. 1988, 28, 536–542. [Google Scholar] [CrossRef]
- Gaska, J.M.; Oplinger, E.S. Yield, lodging and growth characteristics in sweet corn as influenced by ethephon timing and rate. Agron. J. 1988, 80, 722–726. [Google Scholar] [CrossRef]
- Norberg, O.S.; Mason, S.C.; Lowry, S.R. Ethephon influence on harvestable yield, grain quality, and lodging of corn. Agron. J. 1988, 80, 768–772. [Google Scholar] [CrossRef]
- Sanvicente, P.; Lazarevitch, S.; Blouet, A.; Guckert, A. Morphological and anatomical modifications in winter barley culm after late plant growth regulator treatment. Eur. J. Agron. 1990, 11, 45–51. [Google Scholar] [CrossRef]
- Tang, Q.; Ren, J.; Du, X.; Niu, S.; Liu, S.; Wei, D.; Zhang, Y.; Bian, D.; Cui, Y.; Gao, Z. Reduced stem nonstructural carbohydrates caused by plant growth retardant had adverse effects on maize yield under low density. Front. Plant Sci. 2022, 13, 1035254. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.S.; Qu, S.J.; Huang, G.M.; Guo, Y.L.; Zhang, M.C.; Li, Z.H. Improving maize grain yield by formulating plant growth regulator strategies in north China. J. Integr. Agric. 2021, 20, 622–632. [Google Scholar] [CrossRef]
- Huang, G.M.; Liu, Y.R.; Guo, Y.L.; Peng, C.X.; Tan, W.M.; Zhang, M.C. A novel plant growth regulator improves the grain yield of high-density maize crops by reducing stalk lodging and promoting a compact plant type. Field Crop. Res. 2021, 260, 107982. [Google Scholar] [CrossRef]
- Shanahan, J.F.; Nielsen, D.C. Influence of growth retardants (Anti-Gibberllins) on corn vegetative growth, water use, and grain yield under different levels of water stress. Agron. J. 1987, 79, 103–109. [Google Scholar] [CrossRef]
- Kasele, I.N.; Nyirenda, F.; Shanahan, J.F.; Nielsen, D.C.; d’Andria, R. Ethephon alters corn growth, water use, and grain yield under drought stress. Agron. J. 1994, 86, 283–288. [Google Scholar] [CrossRef]
- Peltonen-Sainio, P.; Peltonen, J. Breaking uniculm growth habit of spring cereals at high latitudes by crop management. I. Leaf area index and biomass accumulation. J. Agron. Crop Sci. 1997, 178, 79–86. [Google Scholar] [CrossRef]
- Kochaki, A. Optimal Agronomic and Optimal Breeding in Dry Farming; Mashhad Jahad-University Publishers: Mashhad, Iran, 1997; p. 302. [Google Scholar]
- Rosenberg, N.J.; Blad, B.L.; Verma, S.B. Microclimate: The Biological Environment, 2nd ed.; Wiley-Interscience: New York, NY, USA, 1983; pp. 209–287. [Google Scholar]
- D’Andria, R.; Chiaranda, F.Q.; Lavini, A.; Mori, M. Grain yield and water consumption of ethephon-treated corn under different irrigation regimes. Agron. J. 1997, 89, 104–112. [Google Scholar] [CrossRef]
- Iqbal, N.; Umar, S.; Per, T.S.; Khan, N.A. Ethephon increases photosynthetic-nitrogen use efficiency, proline and antioxidant metabolism to alleviate decrease in photosynthesis under salinity stress in mustard. Plant Signal. Behav. 2017, 12, e1297000. [Google Scholar] [CrossRef]
- Achard, P.; Cheng, H.; De Grauwe, L.; Decat, J.; Schoutteten, H.; Moritz, T.; Van Der Straeten, D.; Peng, J.; Harberd, N.P. Integration of plant responses to environmentally activated phytohormonal signals. Science 2006, 311, 91–94. [Google Scholar] [CrossRef]
- Hendgen, M.; Gunther, S.; Schubert, S.; Lohnertz, O. Ethephon activates the transcription of senescence-associated genes and nitrogen mobilization in grapevine leaves (Vitis vinifera cv. Riesling). Plants 2021, 10, 333. [Google Scholar] [CrossRef]
- Lee, B.R.; Zaman, R.; La, V.H.; Bae, D.W.; Kim, T.H. Ethephon-induced ethylene enhances starch degradation and sucrose transport with an interactive abscisic acid-mediated manner in mature leaves of oilseed rape (Brassica napus L.). Plants 2021, 10, 1670. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Nazar, R.; Syeed, S.; Masood, A.; Khan, N.A. Exogenously-sourced ethylene increases stomatal conductance, photosynthesis and growth under optimal and deficient nitrogen fertilization in mustard. J. Exp. Bot. 2011, 62, 4955–4963. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, B.R.; Islam, M.T.; La, V.H.; Park, S.H.; Bae, D.W.; Kim, T.H. Cultivar variation in hormone- and sugar-response reveals abscisic acid responsive sucrose phloem loading at the early regenerative stage is a significant determinant of seed yield in Brassica napus. Environ. Exp. Bot. 2020, 169, 103917. [Google Scholar] [CrossRef]
- Kasai, M. Regulation of leaf photosynthetic rate correlating with leaf carbohydrate status and activation state of Rubisco under a variety of photosynthetic source/sink balances. Physiol. Plant. 2008, 134, 216–226. [Google Scholar] [CrossRef]
- Durand, M.; Mainson, D.; Porcheron, B.; Maurousset, L.; Lemoine, R.; Pourtau, N. Carbon source-sink relationship in Arabidopsis thaliana: The role of sucrose transporters. Planta 2018, 247, 587–611. [Google Scholar] [CrossRef]
- Coruzzi, G.M.; Zhou, L. Carbon and nitrogen sensing and signaling in plants: Emerging ‘matrix effects’. Curr. Opin. Plant Biol. 2001, 4, 247–253. [Google Scholar] [CrossRef]
- Aharoni, A.; Dixit, S.; Jetter, R.; Thoenes, E.; van Arkel, G.; Pereira, A. The SHINE clade of AP2 domain transcription factors activates wax biosynthesis, alters cuticle properties, and confers drought tolerance when overexpressed in Arabidopsis. Plant Cell 2004, 16, 2463–2480. [Google Scholar] [CrossRef]
- Wan, L.; Zhang, J.; Zhang, H.; Zhang, Z.; Quan, R.; Zhou, S.; Huang, R. The transcriptional regulation of OsDERF1 in OsERF3 and OsAP 2–39 suppresses ethylene synthesis and decreases drought tolerance in rice. PLoS ONE 2011, 6, e25216. [Google Scholar] [CrossRef]
- Wang, Y.; Wan, L.; Zhang, L.; Zhang, Z.; Zhang, H.; Quan, R.; Zhou, S.; Huang, R. An ethylene response factor OsWR1 responsible to drought stress transcriptionally activates wax synthesis related genes and increases wax production in rice. Plant Mol. Biol. 2012, 8, 275–288. [Google Scholar] [CrossRef]
- Mao, Z. Environmental impact of water-saving irrigation for rice. In Irrigation Scheduling: From Theory to Practice, Proceedings of the ICID/FAO Workshop on Irrigation Scheduling, Rome, Italy, 12–13 September 1995; International Commission on Irrigation and Drainage: New Delhi, India, 1996. [Google Scholar]
- Hadi, M.H.S.; Darzi, M.; Ashoorabadi, E.S. Study the effects of conventional and low input production system on energy efficiency of Silybum marianum L. World Acad. Sci. Eng. Technol. 2009, 54, 364–366. [Google Scholar]
- Laskari, M.; Menexes, G.; Kalfas, I.; Gatzolis, I.; Dordas, C. Water Stress Effects on the Morphological, Physiological Characteristics of Maize (Zea mays L.), and on Environmental Cost. Agronomy 2022, 12, 2386. [Google Scholar] [CrossRef]
- Luo, X.; Keenan, T.F. Tropical extreme droughts drive long-term increase in atmospheric CO2 growth rate variability. Nat. Commun. 2022, 13, 1193. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Bowman, K.W.; Schimel, D.S.; Parazoo, N.C.; Jiang, Z.; Lee, M.; Bloom, A.A.; Wunch, D.; Frankenberg, C.; Sun, Y.; et al. Contrasting carbon cycle responses of the tropical continents to the 2015–2016 El Niño. Science 2017, 358, eaam5690. [Google Scholar] [CrossRef] [PubMed]
- Rodenbeck, C.; Zaehle, S.; Keeling, R.; Heimann, M. History of El Niño impacts on the global carbon cycle 1957–2017: A quantification from atmospheric CO2 data. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2018, 373, 20170303. [Google Scholar] [CrossRef]
- Wigneron, J.P.; Fan, L.; Ciais, P.; Bastos, A.; Brandt, M.; Chave, J.; Saatchi, S.; Baccini, A.; Fensholt, R. Tropical forests did not recover from the strong 2015–2016 El Niño event. Sci. Adv. 2020, 6, eaay4603. [Google Scholar] [CrossRef]
- Yang, H.; Ciais, P.; Wigneron, J.P.; Chave, J.; Cartus, O.; Chen, X.; Fan, L.; Green, J.K.; Huang, Y.; Joetzjer, E.; et al. Climatic and biotic factors influencing regional declines and recovery of tropical forest biomass from the 2015/16 El Niño. PNAS 2022, 119, e2101388119. [Google Scholar] [CrossRef]
- Zhao, X.; Pu, C.; Ma, S.T.; Liu, S.L.; Xue, J.F.; Wang, X.; Wang, Y.Q.; Li, S.S.; Lal, R.; Chen, F.; et al. Management-induced greenhouse gases emission mitigation in global rice production. Sci. Total Environ. 2019, 649, 1299–1306. [Google Scholar] [CrossRef]
- Shakoor, A.; Ashraf, F.; Shakoor, S.; Mustafa, A.; Rehman, A.; Altaf, M.M. Biogeochemical transformation of greenhouse gas emissions from terrestrial to atmospheric environment and potential feedback to climate forcing. Environ. Sci. Pollut. Res. 2020, 27, 38513–38536. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). Software for National Gas Inventories, Intergovernmental Panel on Climate Change. Available online: https://www.ipcc-nggip.iges.or.jp/public/2006gl/ (accessed on 25 September 2023).
- Amnuaylojaroen, T.; Chanvichit, P.; Janta, R.; Surapipith, V. Projection of Rice and Maize Productions in Northern Thailand under Climate Change Scenario RCP8.5. Agriculture 2021, 11, 23. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Growth and physiological responses to supra-optimal nitrogen and pre-anthesis drought stress in maize. Agric. Nat. Resour. 2014, 48, 676–688. [Google Scholar]
- Molla, M.S.H.; Nakasathien, S.; Sarobol, E.; Vichukit, V. Effects of nitrogen and water on maize varieties under short and prolonged drought. J. Dev. Sustain. Agric. 2014, 9, 97–110. [Google Scholar] [CrossRef]
- Farahani, A.; Lebaschi, H.; Hussein, M.; Hussein, S.A.; Reza, V.A.; Jahanfar, D. Effects of arbuscular mycorrhizal fungi, different levels of phosphorus and drought stress on water use efficiency, relative water content and proline accumulation rate of Coriander (Coriandrum sativum L.). J. Med. Plant Res. 2008, 2, 125–131. [Google Scholar]
- Diaz-Lopez, E.; Aguilar-Luna, J.M.; Loeza-Corte, J.M. Net assimilation rate and agronomic efficiency of nitrogen in tartago (Ricinus communis L.) (Euphorbiaceae) in dry climate. Scientifica 2020, 2020, 7064745. [Google Scholar] [CrossRef]
- Zhao, J.; Marek, T.H.; Liu, K.; Harrison, M.T.; Xue, Q. Drought tolerant maize hybrids have higher yields and lower water use under drought conditions at a regional scale. Agric. Water Manag. 2022, 274, 107978. [Google Scholar] [CrossRef]
- Singh, J.; Hadda, M.S. Phenology and thermal indices of maize (Zea mays L.) influenced by subsoil compaction and nitrogen fertilization under semi-arid irrigated conditions. J. Appl. Nat. Sci. 2014, 6, 349–355. [Google Scholar] [CrossRef]
- Molla, M.S.H.; Nakasathien, S.; Ali, M.A.; Khan, A.S.M.M.R.; Alam, M.R.; Hossain, A.; Farooq, M.; El Sabagh, A. Influence of nitrogen application on dry biomass allocation and translocation in two maize varieties under short pre-anthesis and prolonged bracketing flowering periods of drought. Arch. Agron. Soil Sci. 2019, 65, 928–944. [Google Scholar] [CrossRef]
- Dionisio-Sese, M.L.; Tobita, S. Antioxidant responses of rice seedlings to salinity stress. Plant Sci. 1998, 135, 1–9. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Emerich, D.W.; Sanchez-Diaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Balbaa, M.G.; Osman, H.T.; Kandil, E.E.; Javed, T.; Lamlom, S.F.; Ali, H.M.; Kalaji, H.M.; Wróbel, J.; Telesiñski, A.; Brysiewicz, A.; et al. Determination of morpho-physiological and yield traits of maize inbred lines (Zea mays L.) under optimal and drought stress conditions. Front. Plant Sci. 2022, 13, 959203. [Google Scholar] [CrossRef]
- Hou, L.; Yang, Y.; Zhang, X.; Jiang, C. Carbon footprint for wheat and maize production modulated by farm size: A study in the North China plain. Int. J. Clim. Change Strateg. Manag. 2021, 13, 302–319. [Google Scholar] [CrossRef]
- Moser, S.B.; Feil, B.; Jampatong, S.; Stamp, P. Effects of pre-anthesis drought, nitrogen fertilizer rate, and variety on grain yield, yield components, and harvest index of tropical maize. Agric. Water Manag. 2006, 81, 41–58. [Google Scholar] [CrossRef]
- Nabizadeh, E.; Banifazel, M.; Taherifard, E. The effects of plant growth promoting on some of traits in maize (cv. S.C.704) under drought stress condition. Eur. J. Exp. Biol. 2012, 2, 875–881. [Google Scholar]
- Anjum, S.A.; Xie, X.; Wang, L.C.; Saleem, M.F.; Man, C.; Lei, W. Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 2011, 6, 2026–2032. [Google Scholar] [CrossRef]
- Li, L.L.; Gu, W.R.; Li, C.F.; Li, W.H.; Chen, X.C.; Zhang, L.G.; Wei, S. Dual application of ethephon and DCPTA increases maize yield and stalk strength. Agron. J. 2019, 111, 612–627. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Y.; Liu, C.; Ye, D.; Ren, D.; Li, Z.; Zhang, M. Ethephon Reduces Maize Nitrogen Uptake but Improves Nitrogen Utilization in Zea mays L. Front. Plant Sci. 2022, 12, 762736. [Google Scholar] [CrossRef]
- Karimi, A.; Amirnia, R.; Tajbakhsh, M.; Eivaz, A.R.; Karimi, K. Effect of plant growth inducers on morpho-physiological traits of corn (Zea mays L). Life Sci. 2012, 9, 1683–1688. [Google Scholar]
- Begum, N.; Ahanger, M.A.; Su, Y.; Lei, Y.; Mustafa, N.S.A.; Ahmad, P.; Zhang, L. Improved Drought Tolerance by AMF Inoculation in Maize (Zea mays) Involves Physiological and Biochemical Implications. Plants 2019, 8, 579. [Google Scholar] [CrossRef]
- Kępczyńska, E.; Zielińska, S. The role of endogenous ethylene in carbohydrate metabolism of Medicago sativa L. somatic embryos in relation to their regenerative ability. J. Plant Growth Regul. 2013, 32, 191–199. [Google Scholar] [CrossRef]
- Golbashy, M.; Ebrahimi, M.; Khavari, K.S.; Choucan, R. Evaluation of drought tolerance of some corn (Zea mays L.) hybrids in Iran. Afr. J. Agric. Res. 2010, 5, 2714–2719. [Google Scholar]
- Araus Ortega, J.L.; Serret Molins, M.D.; Edmeades, G.O. Phenotyping maize for adaptation to drought. Front. Physiol. 2012, 3, 305. [Google Scholar] [CrossRef]
- Coque, M.; Gallais, A. Genetic variation for nitrogen remobilization and post-silking nitrogen uptake in maize recombinant inbred lines: Heritabilities and correlations among traits. Crop Sci. 2007, 47, 1787–1796. [Google Scholar] [CrossRef]
- Chen, Y.L.; Xiao, C.X.; Wu, D.L.; Xia, T.T.; Chen, Q.W.; Chen, F.J. Effects of nitrogen application rate on grain yield and grain nitrogen concentration in two maize hybrids with contrasting nitrogen remobilization efficiency. Eur. J. Agron. 2015, 62, 79–89. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef] [PubMed]
- Maillard, A.; Diquélou, S.; Billard, V.; Laîné, P.; Garnica, M.; Prudent, M.; Garcia-Mina, J.-M.; Yvin, J.-C.; Ourry, A. Leaf mineral nutrient remobilization during leaf senescence and modulation by nutrient deficiency. Front. Plant Sci. 2015, 6, 317. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Aziz, T.; Wahid, A.; Lee, D.J.; Siddique, K.H. Chilling tolerance in maize: Agronomic and physiological approaches. Crop Pasture Sci. 2009, 60, 501–516. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2021, 27, 699–716. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Janz, B.; Engedal, T.; Neergaard, A. Effect of irrigation regimes and nitrogen rates on water use efficiency and nitrogen uptake in maize. Agric. Water Manag. 2016, 179, 271–276. [Google Scholar] [CrossRef]
- Malo, M.; Ghosh, A. Studies on different agrometeorological indices and thermal use efficiencies of rice in New Alluvial Zone of West Bengal. Bull. Env. Pharmacol. Life Sci. 2018, 7, 72–78. [Google Scholar]
- Horvath, E.; Gombos, B.; Szeles, A. Evaluation phenology, yield and quality of maize genotypes in drought stress and non-stress environments. Agron. Res. 2021, 19, 408–422. [Google Scholar] [CrossRef]
- Ye, D.; Wang, Y.; Zhou, L.; Li, J.; Duan, L.; Zhang, M.; Li, Z. Effect of ethephon and nitrogen fertilizer on nitrogen uptake, nitrogen use efficiency and yield of summer maize. Acta Agron. Sin. 2015, 41, 1701–1710. [Google Scholar] [CrossRef]
- Yu, H.; Zhang, Y.; Xie, Y.; Wang, Y.; Duan, L.; Zhang, M.; Li, Z. Ethephon improved drought tolerance in maize seedlings by modulating cuticular wax biosynthesis and membrane stability. J. Plant Physiol. 2017, 214, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kirkham, M.B. Water status of drought-resistant and drought-sensitive sorghum treated with ethephon. J. Plant Growth Regulat. 1990, 9, 189–194. [Google Scholar] [CrossRef]
- Shen, S.; Li, B.B.; Deng, T.; Xiao, Z.D.; Chen, X.M.; Hu, H.; Zhang, B.C.; Wu, G.; Li, F.; Zhao, X.; et al. The equilibrium between sugars and ethylene is involved in shading-and drought-induced kernel abortion in maize. Plant Growth Regul. 2020, 91, 101–111. [Google Scholar] [CrossRef]
- Bolaños, J.; Edmeades, G.O. The importance of the anthesis-silking interval in breeding for drought tolerance in tropical maize. Field Crop. Res. 1996, 48, 65–80. [Google Scholar] [CrossRef]
- El-Badawy, M.; El, M.; Mehasen, S.A.S. Multivariate Analysis For Yield and Its Components In Maize Under Zinc and Nitrogen Fertilization Levels. Aust. J. Basic Appl. Sci. 2011, 5, 3008–3015. [Google Scholar]
- Chen, K.; Camberato, J.J.; Vyn, T.J. Maize Grain Yield and Kernel Component Relationships to Morpho-physiological Traits in Commercial Hybrids Separated by Four Decades. Crop Sci. 2017, 57, 1641–1657. [Google Scholar] [CrossRef]
- Dewey, D.R.; Lu, K.H. A correlation and path coefficient analysis of components of crescent wheat grass seed production. Agron. J. 1959, 51, 515–518. [Google Scholar] [CrossRef]
- Aslam, M.; Khan, N.A.; Mirza, M.S.; Khan, A.R. Correlation and path coefficient analysis for yield components in soybean. Pak. J. Agric. Res. 1992, 13, 20–25. [Google Scholar]
- Lenka, D.; Mishra, B. Path coefficient analysis of yield in rice varieties. Ind. J. Agric. Sci. 1973, 43, 376–379. [Google Scholar]
- Ahmadizadeh, M.; Valizadeh, M.; Zaefizadeh, M.; Shahbazi, H. Antioxidative protection and electrolyte leakage in durum wheat under drought stress condition. J. Appl. Sci. Res. 2011, 7, 236–246. [Google Scholar]
- Sarvajeet, S.G.; Narendra, T. Reactive oxygen species and antioxidant machinery in a biotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 3, 909–930. [Google Scholar] [CrossRef]
- Poudel, S.; Bhattarai, S.; Sherpa, T.; Karki, A.; Hyun, D.; Kafle, S. The energy input-output analysis of maize production in Sundarharaincha Municipality, Morang district, Nepal. IOP Conf. Ser. Earth Environ. Sci. 2019, 301, 012027. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Chai, M.; Yang, D.; van der Werf, W.; Evers, J. Use of EDAH improves maize morphological and mechanical traits related to lodging. Agron. J. 2019, 111, 581–591. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, C.; Zhang, K.; Zhou, Y.; Tan, W.; Zhang, L. Plant growth regulator and its interactions with environment and genotype affect maize optimal plant density and yield. Eur. J. Agron. 2017, 91, 34–43. [Google Scholar] [CrossRef]
- Huang, X.; Chen, C.; Qian, H.; Chen, M.; Deng, A.; Zhang, J.; Zhang, W. Quantification for carbon footprint of agricultural inputs of grains cultivation in China since 1978. J. Clean. Prod. 2017, 142, 1629–1637. [Google Scholar] [CrossRef]
- Cheng, K.; Pan, G.; Smith, P.; Luo, T.; Li, L.; Zheng, J.; Zhang, X.; Han, X.; Yan, M. Carbon footprint of China’s crop production—An estimation using agro-statistics data over 1993–2007. Agric. Ecosyst. Environ. 2011, 142, 231–237. [Google Scholar] [CrossRef]
- Yan, M.; Cheng, K.; Luo, T.; Yan, Y.; Pan, G.; Rees, R.M. Carbon footprint of grain crop production in China—Based on farm survey data. J. Clean. Prod. 2015, 104, 130–138. [Google Scholar] [CrossRef]
- Climate Change Knowledge Portal. Thailand 1991–2020. Available online: https://climateknowledgeportal.worldbank.org/country/thailand/climate-data-historical (accessed on 25 September 2023).
- Lorzadeh, S.H.; Mahdavidamghani, A.; Enayatgholizadeh, M.R.; Yousefi, M. Agrochemical input application and energy use efficiency of maize production systems in Dezful, Iran. Middle East J. Sci. Res. 2011, 9, 153–156. [Google Scholar]
- Vural, H.; Efecan, I. An analysis of energy use and input costs for maize production in Turkey. J. Food Agric. Environ. 2012, 10, 613–616. [Google Scholar]
- Erdal, G.; Esengun, K.; Erdal, H.; Gunduz, O. Energy use and economical analysis of sugar beet production in Tokat province of Turkey. Energy 2007, 32, 35–41. [Google Scholar] [CrossRef]
- Zhelyazkova, T. Nutritive value and energy efficiency of winter pea production, treated with plant growth regulators. Trakia J. Sci. 2010, 8, 44–51. [Google Scholar]
- Shahin, R.; Mousavi-Avval, S.; Mohammadi, A. Modeling and sensitivity analysis of energy inputs for apple production in Iran. Energy 2010, 35, 3301–3306. [Google Scholar] [CrossRef]
- Yousefi, M.; Mohammadi, A. Economical analysis and energy use efficiency in alfalfa production systems in Iran. Sci. Res. Essays 2011, 6, 2332–2336. [Google Scholar] [CrossRef]
- Zhang, W.; Dou, Z.; Hea, P.; Jua, X.; Powlson, D.; Chadwick, D.; Norse, D.; Lu, Y.; Zhang, Y.; Wu, L.; et al. New technologies reduce greenhouse gas emissions from nitrogenous fertilizer in China. Proc. Natl. Acad. Sci. USA 2013, 110, 8375–8380. [Google Scholar] [CrossRef] [PubMed]
- West, T.; Marland, G. A synthesis of carbon sequestration, carbon emissions, and net carbon flux in agriculture: Comparing tillage practices in the United States. Agric. Ecosyst. Environ. 2002, 91, 217–232. [Google Scholar] [CrossRef]
- Zhu, Y.; Waqas, M.; Li, Y.; Zou, X.; Jiang, D.; Wilkes, A.; Qin, X.; Gao, Q.; Wan, Y.; Hasbagan, G. Large-scale farming operations are win-win for grain production, soil carbon storage and mitigation of greenhouse gases. J. Clean. Prod. 2018, 172, 2143–2152. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Sand (%) | Silt (%) | Clay (%) | pH (Acetate) | EC (dS m−1) | OM (%) | Total N (%) | Available P (mg kg−1) | Exchangeable K (mg kg−1) |
|---|---|---|---|---|---|---|---|---|---|
| Value | 10.00 | 37.33 | 52.67 | 7.77 | 0.22 | 2.01 | 0.28 | 166.00 | 296.67 |
| Status | Silty clay soil | Slightly alkaline | Normal | Medium | Medium | Very high | Very high | ||
| Source of Variance | df | Mean Sum Square | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PH-R3 (cm) | SD-R3 (cm) | FEGLA-V6 (cm2 plant−1) | FEGLA-V10 (cm2 plant−1) | FEGLA-R3 (cm2 plant−1) | LAI-R3 | SW-V6 (g plant−1) | SW-V10 (g plant−1) | SW-R3 (g plant−1) | GNP (no.) | 100-GW (g) | GY (t ha−1) | ||
| Replication | 3 | 616.3 | 0.07 | 18,455 | 115,975 | 271,988 | 0.09 | 2.64 | 18.81 | 571 | 1697.3 | 8.00 | 0.31 |
| Water levels (W) | 2 | 11,065.8 ** | 0.26 ** | 41,037 ** | 248.35 ns | 2615.07 ** | 8.52 ** | 7.16 ** | 1.33 ns | 208,132 ** | 3452.9 ** | 18.58 ** | 29.40 ** |
| Ethephon (E) | 6 | 3400.4 ** | 0.20 ** | 14,951 ns | 271.07 ** | 6674.07 ** | 21.83 ** | 1.94 ns | 2334.40 ** | 98,659 ** | 580.5 ns | 0.49 * | 30.99 ** |
| W × E | 12 | 13,384.1 ** | 0.19 * | 37,202 ns | 38,673.0 ns | 5,910,228 ** | 1.93 ** | 4.31 ns | 11.45 ns | 71,474 ** | 0.2 ns | 0.00 ns | 16.60 ** |
| Error | 60 | 3466.6 | 0.43 | 113,412 | 816,281 | 1,670,232 | 0.54 | 16.43 | 133.03 | 3760 | 10,457.9 | 49.30 | 2.06 |
| Source of Variance | df | Mean Sum Square | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| RGR-V6V10 (mg plant−1 day−1) | RGR-V10R3 (mg plant−1 day−1) | RGR-V6R3 (mg plant−1 day−1) | NAR-V6V10 (mg cm−2 day−1) | NAR-V10R3 (mg cm−2 day−1) | NAR-V6R3 (mg cm−2 day−1) | WP (kg m−3) | AGDD (°C days) | HUE (kg ha−1 °C days−1) | ||
| Replication | 3 | 16.19 | 5.09 | 7.17 | 0.002 | 0.01 | 0.01 | 0.01 | 39,964 | 0.10 |
| Water levels (W) | 2 | 38.96 ** | 661.73 ** | 380.63 ** | 0.005 ** | 1.11 ** | 1.20 ** | 0.03 ** | 93,870 ** | 12.86 ** |
| Ethephon (E) | 6 | 1875.72 ** | 225.72 ** | 197.52 ** | 0.322 ** | 0.35 ** | 0.60 ** | 1.25 ** | 47,406 ns | 9.35 ** |
| W × E | 12 | 32.33 ns | 210.33 ** | 115.34 ** | 0.008 * | 0.33 ** | 0.46 ** | 0.59 ** | 10,750 ns | 5.82 ** |
| Error | 60 | 116.01 | 31.07 | 46.15 | 0.018 | 0.03 | 0.04 | 0.08 | 251,875 | 0.68 |
| Source of Variance | df | Mean Sum Square | |||||
|---|---|---|---|---|---|---|---|
| SPAD-JAWSP | RSR-R3 (%) | RWC-JAWSP (%) | EL-JAWSP (%) | PrL-JAWSP (µmol g−1 FW) | TSSL-JAWSP (mg g−1 FW) | ||
| Replication | 3 | 25.88 | 0.06 | 93.87 | 0.42 | 7.74 | 52,462.9 |
| Water levels (W) | 2 | 1026.69 ** | 4.83 ** | 2108.57 ** | 177.23 ** | 5266.61 ** | 5544.07 ** |
| Ethephon (E) | 6 | 190.05 ** | 1.18 ** | 695.38 ** | 22.05 ** | 866.21 ** | 1925.07 ** |
| W × E | 12 | 44.65 ns | 0.65 ** | 196.65 ns | 10.80 ** | 225.28 ** | 4,130,239 ** |
| Error | 60 | 159.38 | 0.40 | 588.42 | 3.17 | 55.83 | 470,809 |
| Source of Variation | PH-R3 (cm) | SD-R3 (cm) | FEGLA-V6 (cm2 plant−1) | FEGLA-V10 (cm2 plant−1) | FEGLA-R3 (cm2 plant−1) | LAI-R3 | SW-V6 (g plant−1) | SW-V10 (g plant−1) | SW-R3 (g plant−1) | GNP (no.) | 100-GW (g) | GY (t ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W1 | 219.25a | 2.39a | 1172.26b | 3134.62 | 5367.53a | 3.07a | 14.02b | 39.93 | 275.02a | 363.27a | 24.91a | 5.71a |
| W2 | 210.63b | 2.30b | 1207.18a | 3131.14 | 4455.24b | 2.55b | 14.61a | 40.19 | 186.00b | 358.40a | 24.64a | 4.82b |
| W3 | 195.09c | 2.25b | 1153.89b | 3134.94 | 4030.16c | 2.30c | 13.97b | 40.19 | 158.35c | 347.90b | 23.81b | 4.27c |
| F test (W) | ** | ** | ** | ns | ** | ** | ** | ns | ** | ** | ** | ** |
| LSD0.05 | 4.06 | 0.05 | 23.24 | 62.36 | 89.20 | 0.06 | 0.28 | 0.80 | 4.23 | 7.06 | 0.48 | 0.10 |
| Source of Variation | PH-R3 (cm) | SD-R3 (cm) | FEGLA-V6 (cm2 plant−1) | FEGLA-V10 (cm2 plant−1) | FEGLA-R3 (cm2 plant−1) | LAI-R3 | SW-V6 (g plant−1) | SW-V10 (g plant−1) | SW-R3 (g plant−1) | GNP (no.) | 100-GW (g) | GY (t ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E1 | 217.62ab | 2.35ab | 1159.65b | 3063.74b | 5142.09c | 2.94c | 14.09ab | 38.69b | 219.37c | 356.12 | 24.52ab | 5.11c |
| E2 | 204.78c | 2.31bc | 1193.87ab | 3079.82b | 3781.06f | 2.16f | 14.01b | 38.85b | 174.67f | 354.34 | 24.43ab | 4.49d |
| E3 | 217.92ab | 2.39a | 1173.00ab | 3527.48a | 5427.25b | 3.10b | 14.15ab | 45.30a | 260.67a | 357.44 | 24.90a | 5.96a |
| E4 | 211.98b | 2.26cd | 1176.52ab | 2592.77c | 4122.33e | 2.36e | 14.33ab | 33.18c | 185.17e | 355.58 | 24.44ab | 4.53d |
| E5 | 204.73c | 2.24d | 1175.46ab | 2601.95c | 3234.95g | 1.85g | 14.06ab | 33.04c | 158.83g | 353.35 | 24.17b | 4.18e |
| E6 | 203.58c | 2.30bcd | 1199.92a | 3523.98a | 4667.30d | 2.67d | 14.44a | 45.42a | 243.50b | 356.67 | 24.44ab | 5.63b |
| E7 | 218.64a | 2.33ab | 1166.01ab | 3545.22a | 5948.52a | 3.40a | 14.32ab | 46.25a | 202.98d | 352.14 | 24.38ab | 4.63d |
| F test (W) | ** | ** | ns | ** | ** | ** | ns | ** | ** | ns | * | ** |
| LSD0.05 | 6.21 | 0.07 | 35.50 | 95.25 | 135.25 | 0.12 | 0.43 | 1.22 | 6.46 | 10.78 | 0.64 | 0.15 |
| Source of Variation | PH-R3 (cm) | SD-R3 (cm) | FEGLA-V6 (cm2 plant−1) | FEGLA-V10 (cm2 plant−1) | FEGLA-R3 (cm2 plant−1) | LAI-R3 | SW-V6 (g plant−1) | SW-V10 (g plant−1) | SW-R3 (g plant−1) | GNP (no.) | 100-GW (g) | GY (t ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| W1E1 | 223.11c | 2.39bc | 1177.03 | 3058.12 | 5944.34c | 3.40b | 13.88 | 39.11 | 278.12d | 362.85 | 24.98 | 5.32ef |
| W1E2 | 204.66ef | 2.39bc | 1217.79 | 3106.05 | 4398.85h | 2.51ef | 14.11 | 39.22 | 221.50f | 361.04 | 24.89 | 5.03gh |
| W1E3 | 221.21cd | 2.43b | 1157.08 | 3536.11 | 6242.67b | 3.57b | 13.88 | 45.38 | 317.00b | 364.21 | 25.07 | 6.74ab |
| W1E4 | 211.48de | 2.30c-g | 1124.87 | 2561.13 | 4429.98gh | 2.53e | 13.78 | 32.81 | 240.50e | 362.31 | 24.90 | 4.89hi |
| W1E5 | 200.38fg | 2.29c-g | 1160.24 | 2629.20 | 3769.79ij | 2.15hi | 13.83 | 32.34 | 203.50h | 360.03 | 24.82 | 4.67ij |
| W1E6 | 225.64bc | 2.34b-e | 1211.43 | 3521.31 | 5379.77de | 3.07c | 14.35 | 45.12 | 295.00c | 363.42 | 24.90 | 6.50b |
| W1E7 | 248.25a | 2.57a | 1157.38 | 3530.42 | 7407.31a | 4.23a | 14.30 | 45.52 | 369.50a | 369.00 | 24.84 | 6.79a |
| W2E1 | 231.21bc | 2.38bc | 1168.31 | 3046.10 | 5159.51ef | 2.95c | 14.28 | 38.34 | 221.00fg | 357.99 | 24.71 | 5.62cd |
| W2E2 | 225.87bc | 2.31b-f | 1188.56 | 3032.05 | 3593.63j | 2.05hi | 14.00 | 38.51 | 167.50i | 356.20 | 24.62 | 4.55jk |
| W2E3 | 234.65b | 2.39bc | 1222.58 | 3551.12 | 5388.99de | 3.08c | 14.71 | 45.00 | 240.50e | 359.32 | 24.79 | 5.75c |
| W2E4 | 230.70bc | 2.25d-g | 1248.60 | 2605.04 | 3948.50i | 2.26gh | 14.80 | 33.39 | 172.50i | 357.46 | 24.63 | 4.65ij |
| W2E5 | 229.36bc | 2.23efg | 1198.70 | 2598.34 | 3049.42l | 1.74jk | 14.43 | 33.86 | 148.00kl | 355.21 | 24.55 | 4.16lm |
| W2E6 | 182.76i | 2.28c-g | 1237.85 | 3540.21 | 4641.48g | 2.65de | 15.09 | 45.40 | 225.50f | 358.54 | 24.63 | 5.29efg |
| W2E7 | 202.84efg | 2.23efg | 1185.66 | 3545.12 | 5405.16d | 3.09c | 14.97 | 46.84 | 127.00n | 364.05 | 24.57 | 3.74n |
| W3E1 | 198.54fg | 2.29c-g | 1133.62 | 3087.01 | 4322.42h | 2.47efg | 14.11 | 38.61 | 159.00jk | 347.51 | 23.87 | 4.38kl |
| W3E2 | 183.81hi | 2.23efg | 1175.27 | 3101.35 | 3350.71k | 1.91ij | 13.92 | 38.83 | 135.00mn | 345.77 | 23.79 | 3.90mn |
| W3E3 | 197.90fg | 2.36bcd | 1139.33 | 3495.21 | 4650.09g | 2.66de | 13.86 | 45.52 | 224.50f | 348.80 | 23.96 | 5.38de |
| W3E4 | 193.76gh | 2.23efg | 1156.08 | 2612.14 | 3988.52i | 2.28fgh | 14.42 | 33.33 | 142.50lm | 346.99 | 23.80 | 4.05m |
| W3E5 | 184.47hi | 2.19g | 1167.44 | 2578.30 | 2885.64l | 1.65k | 13.90 | 32.92 | 125.00n | 344.80 | 23.72 | 3.70n |
| W3E6 | 202.33efg | 2.27c-g | 1150.48 | 3510.41 | 3980.65i | 2.27fgh | 13.89 | 45.75 | 210.00gh | 348.04 | 23.80 | 5.10fgh |
| W3E7 | 204.82ef | 2.20fg | 1155.00 | 3560.13 | 5033.11f | 2.88cd | 13.69 | 46.40 | 112.45o | 353.39 | 23.74 | 3.37o |
| F test (W) | ** | * | ns | ns | ** | ** | ns | ns | ** | ns | ns | ** |
| LSD0.05 | 10.75 | 0.12 | - | - | 235.99 | 0.25 | - | - | 11.20 | - | - | 0.26 |
| Source of Variation | RGR-V6V10 (mg plant−1 day−1) | RGR-V10R3 (mg plant−1 day−1) | RGR-V6R3 (mg plant−1 day−1) | NAR-V6V10 (mg cm−2 day−1) | NAR-V10R3 (mg cm−2 day−1) | NAR-V6R3 (mg cm−2 day−1) | WP (kg m−3) | AGDD (°C days) | HUE (kg ha−1 °C days−1) |
|---|---|---|---|---|---|---|---|---|---|
| W1. | 37.55a | 22.54a | 26.21a | 0.47a | 0.66a | 0.84a | 0.97b | 1705.72b | 3.34a |
| W2 | 36.33b | 17.80b | 22.34b | 0.46b | 0.46b | 0.61b | 0.95c | 1774.20a | 2.72b |
| W3 | 37.92a | 15.85c | 21.26c | 0.47a | 0.39c | 0.56c | 1.00a | 1778.84a | 2.40c |
| F test (W) | ** | ** | ** | ** | ** | ** | ** | ** | ** |
| LSD0.05 | 0.74 | 0.38 | 0.47 | 0.001 | 0.01 | 0.01 | 0.02 | 34.64 | 0.06 |
| Source of Variation | RGR-V6V10 (mg plant−1 day−1) | RGR-V10R3 (mg plant−1 day−1) | RGR-V6R3 (mg plant−1 day−1) | NAR-V6V10 (mg cm−2 day−1) | NAR-V10R3 (mg cm−2 day−1) | NAR-V6R3 (mg cm−2 day−1) | WP (kg m−3) | AGDD (°C days) | HUE (kg ha−1 °C days−1) |
|---|---|---|---|---|---|---|---|---|---|
| E1 | 36.56b | 20.07ab | 24.11c | 0.45c | 0.52c | 0.67b | 1.01c | 1757.47ab | 2.91c |
| E2 | 36.92b | 17.40d | 22.18d | 0.45c | 0.46e | 0.63c | 0.89d | 1734.69b | 2.60d |
| E3 | 42.12a | 20.41a | 25.73a | 0.53ab | 0.57a | 0.79a | 1.18a | 1757.47ab | 3.39a |
| E4 | 30.39c | 19.90ab | 22.47d | 0.38d | 0.54b | 0.64c | 0.90d | 1734.69b | 2.62d |
| E5 | 30.93c | 18.19c | 21.31e | 0.38d | 0.50d | 0.62c | 0.83e | 1723.13b | 2.43e |
| E6 | 41.49a | 19.58b | 24.95b | 0.52b | 0.57a | 0.79a | 1.11b | 1762.46ab | 3.20b |
| E7 | 42.45a | 15.56e | 22.15d | 0.54a | 0.37f | 0.54d | 0.89d | 1800.53a | 2.60d |
| F test (W) | ** | ** | ** | ** | ** | ** | ** | ns | ** |
| LSD0.05 | 1.14 | 0.59 | 0.72 | 0.01 | 0.02 | 0.02 | 0.03 | 52.91 | 0.09 |
| Source of Variation | RGR-V6V10 (mg plant−1 day−1) | RGR-V10R3 (mg plant−1 day−1) | RGR-V6R3 (mg plant−1 day−1) | NAR-V6V10 (mg cm−2 day−1) | NAR-V10R3 (mg cm−2 day−1) | NAR-V6R3 (mg cm−2 day−1) | WP (kg m−3) | AGDD (°C days) | HUE (kg ha−1 °C days−1) |
|---|---|---|---|---|---|---|---|---|---|
| W1E1 | 37.49 | 23.03bc | 26.57bc | 0.46d | 0.65bc | 0.80d | 0.91efg | 1723.12 | 3.09bcd |
| W1E2 | 37.01 | 20.32e | 24.41de | 0.45d | 0.58d | 0.74ef | 0.86gh | 1688.35 | 2.98de |
| W1E3 | 42.86 | 22.82bc | 27.73ab | 0.54a | 0.67b | 0.89b | 1.15bc | 1723.12 | 3.91a |
| W1E4 | 31.39 | 23.38b | 25.34cd | 0.39e | 0.71a | 0.83c | 0.83hi | 1688.35 | 2.90ef |
| W1E5 | 30.74 | 21.59d | 23.83e | 0.37f | 0.63c | 0.76de | 0.79i | 1688.35 | 2.77fg |
| W1E6 | 41.46 | 22.04cd | 26.80b | 0.51bc | 0.67b | 0.89b | 1.11c | 1705.64 | 3.81a |
| W1E7 | 41.91 | 24.58a | 28.82a | 0.53ab | 0.73a | 0.94a | 1.16bc | 1723.12 | 3.94a |
| W2E1 | 35.75 | 20.56e | 24.28de | 0.44d | 0.53e | 0.68g | 1.11c | 1774.65 | 3.17bc |
| W2E2 | 36.63 | 17.25hi | 22.00f | 0.45d | 0.46f | 0.63h | 0.90fg | 1757.87 | 2.59hi |
| W2E3 | 40.47 | 19.67ef | 24.77de | 0.50c | 0.52e | 0.71fg | 1.13c | 1774.65 | 3.24b |
| W2E4 | 29.45 | 19.27f | 21.77fg | 0.36f | 0.51e | 0.60h | 0.92ef | 1757.87 | 2.65gh |
| W2E5 | 30.86 | 17.31hi | 20.63ghi | 0.39ef | 0.48f | 0.60h | 0.82hi | 1740.53 | 2.39jk |
| W2E6 | 39.87 | 18.81fg | 23.97e | 0.50c | 0.52e | 0.72ef | 1.04d | 1774.65 | 2.98de |
| W2E7 | 41.28 | 11.71l | 18.95jk | 0.54ab | 0.21i | 0.36k | 0.74j | 1839.23 | 2.03n |
| W3E1 | 36.44 | 16.61ij | 21.47fgh | 0.45d | 0.39g | 0.54i | 1.02d | 1774.65 | 2.47ij |
| W3E2 | 37.12 | 14.63k | 20.14ij | 0.45d | 0.35h | 0.52ij | 0.91ef | 1757.87 | 2.22lm |
| W3E3 | 43.04 | 18.73fg | 24.68de | 0.55a | 0.52e | 0.76e | 1.26a | 1774.65 | 3.03cde |
| W3E4 | 30.33 | 17.05hi | 20.30hi | 0.38ef | 0.39g | 0.50j | 0.95e | 1757.87 | 2.30kl |
| W3E5 | 31.21 | 15.66j | 19.47ijk | 0.39ef | 0.40g | 0.52ij | 0.87fgh | 1740.53 | 2.13mn |
| W3E6 | 43.15 | 17.89gh | 24.07e | 0.55a | 0.52e | 0.76de | 1.19b | 1807.09 | 2.82f |
| W3E7 | 44.17 | 10.39m | 18.66k | 0.55a | 0.18j | 0.33k | 0.79i | 1839.23 | 1.83o |
| F test (W) | ns | ** | ** | * | ** | ** | ** | ns | ** |
| LSD0.05 | - | 1.02 | 1.24 | 0.02 | 0.03 | 0.04 | 0.05 | - | 0.15 |
| Source of Variation | SPAD-JAWSP | RSR-R3 (%) | RWC-JAWSP (%) | EL-JAWSP (%) | PrL-JAWSP (µmol g−1 FW) | TSSL-JAWSP (mg g−1 FW) |
|---|---|---|---|---|---|---|
| W1 | 48.39a | 2.41a | 89.84a | 4.26c | 16.02c | 1054.79c |
| W2 | 42.09b | 1.89b | 80.36b | 6.41b | 35.42a | 2852.31a |
| W3 | 40.22c | 1.92b | 78.35c | 7.79a | 25.91b | 2692.75b |
| F test (W) | ** | ** | ** | ** | ** | ** |
| LSD0.05 | 0.87 | 0.04 | 1.67 | 0.12 | 0.52 | 47.36 |
| Source of Variation | SPAD-JAWSP | RSR-R3 (%) | RWC-JAWSP (%) | EL-JAWSP (%) | PrL-JAWSP (µmol g−1 FW) | TSSL-JAWSP (mg g−1 FW) |
|---|---|---|---|---|---|---|
| E1 | 44.92ab | 1.95d | 82.47c | 6.01cd | 27.04b | 2086.07e |
| E2 | 42.25d | 2.15b | 81.76c | 5.90d | 27.06b | 2458.83b |
| E3 | 46.10a | 2.01c | 88.19a | 5.69e | 28.78a | 2254.49d |
| E4 | 42.58cd | 2.17b | 82.19c | 6.21b | 26.38bc | 2349.31c |
| E5 | 41.40d | 2.26a | 81.43c | 6.11bc | 26.03c | 2587.05a |
| E6 | 43.74bc | 2.06c | 85.42b | 5.82de | 27.04b | 2566.32a |
| E7 | 43.99b | 1.91d | 78.49d | 7.35a | 18.17d | 1097.58f |
| F test (W) | ** | ** | ** | ** | ** | ** |
| LSD0.05 | 1.33 | 0.07 | 2.56 | 0.19 | 0.79 | 72.34 |
| Source of Variation | SPAD-JAWSP | RSR-R3 (%) | RWC-JAWSP (%) | EL-JAWSP (%) | PrL-JAWSP (µmol g−1 FW) | TSSL-JAWSP (mg g−1 FW) |
|---|---|---|---|---|---|---|
| W1E1 | 49.38 | 2.11cd | 89.03 | 4.28j | 16.57ij | 610.18h |
| W1E2 | 47.51 | 2.49b | 88.37 | 4.23j | 17.60hi | 987.72g |
| W1E3 | 49.42 | 2.19c | 97.10 | 4.22j | 16.68i | 1112.19fg |
| W1E4 | 47.01 | 2.53b | 88.08 | 4.31j | 16.70i | 1173.17f |
| W1E5 | 46.14 | 2.68a | 88.02 | 4.24j | 15.22j | 1508.87e |
| W1E6 | 48.91 | 2.46b | 89.04 | 4.26j | 16.71i | 1456.89e |
| W1E7 | 50.37 | 2.42b | 89.24 | 4.31j | 12.70k | 534.50h |
| W2E1 | 44.25 | 1.80hi | 80.12 | 6.12gh | 35.86c | 2860.15c |
| W2E2 | 40.13 | 1.95fg | 79.89 | 5.92hi | 36.72bc | 3208.87a |
| W2E3 | 45.77 | 1.91gh | 84.13 | 5.76i | 40.68a | 2861.12c |
| W2E4 | 40.52 | 1.97efg | 81.03 | 6.35g | 37.12bc | 3054.08b |
| W2E5 | 40.05 | 2.03def | 79.14 | 6.31g | 36.74bc | 3140.25ab |
| W2E6 | 41.78 | 1.92fg | 84.10 | 5.97hi | 37.26b | 3111.50ab |
| W2E7 | 42.15 | 1.64j | 74.12 | 8.42b | 23.56g | 1730.23d |
| W3E1 | 41.12 | 1.93fg | 78.25 | 7.63d | 28.71d | 2787.88c |
| W3E2 | 39.11 | 2.00defg | 77.02 | 7.54de | 26.85e | 3179.90a |
| W3E3 | 43.10 | 1.94fg | 83.35 | 7.10f | 28.98d | 2790.16c |
| W3E4 | 40.21 | 2.02defg | 77.45 | 7.97c | 25.33f | 2820.68c |
| W3E5 | 38.01 | 2.08cde | 77.12 | 7.78cd | 26.13ef | 3112.04ab |
| W3E6 | 40.52 | 1.79i | 83.13 | 7.24ef | 27.15e | 3130.56ab |
| W3E7 | 39.45 | 1.67j | 72.12 | 9.31a | 18.24h | 1028.01g |
| F test (W) | ns | ** | ns | ** | ** | ** |
| LSD0.05 | - | 0.12 | 0.33 | 1.36 | 125.29 |
| Trait | Indirect Effect via Following Morphological Traits | Total Correlation with GY | Trait | Indirect Effect via Following Physio-Biochemical Traits | Total Correlation with GY | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PH-R3 | SD-R3 | LAI-R3 | SW-R3 | GNP | 100-GW | NAR-V6R3 | SPAD-JAWSP | RWC-JAWSP | EL-JAWSP | PrL-JAWSP | TSSL-JAWSP | ||||
| PH-R3 | 0.00 | 0.39 | 0.37 | 0.07 | −0.04 | 0.67 | NAR-V6R3 | 0.14 | 0.17 | −0.19 | 0.07 | 0.08 | 0.95 | ||
| SD-R3 | 0.04 | 0.19 | 0.93 | 0.00 | −0.16 | 1.00 | SPAD-JAWSP | 0.42 | 0.11 | −0.09 | 0.04 | 0.00 | 0.71 | ||
| LAI-R3 | 0.11 | 0.00 | 0.26 | 0.07 | −0.07 | 0.83 | RWC-JAWSP | 0.68 | 0.14 | −0.19 | 0.07 | 0.08 | 0.95 | ||
| SW-R3 | 0.05 | 0.01 | 0.11 | 0.00 | −0.12 | 1.00 | EL-JAWSP | −0.60 | −0.10 | −0.15 | −0.09 | −0.11 | −0.84 | ||
| GNP | 0.11 | 0.00 | 0.40 | −0.03 | −0.01 | 0.76 | PrL-JAWSP | 0.52 | 0.10 | 0.14 | −0.20 | 0.10 | 0.75 | ||
| 100-GW | 0.03 | 0.01 | 0.20 | 0.82 | 0.00 | 0.90 | TSSL-JAWSP | 0.44 | 0.01 | 0.11 | −0.19 | 0.08 | 0.57 | ||
| Water Level | Energy Efficiency | |||
|---|---|---|---|---|
| Ethephon Application | Well-Watered Conditions (W1) | Short Water Stress (W2) | Prolonged Water Stress (W3) | |
| E1 = 281 g a.i. ha−1 at V6 stage | 1.48cde | 1.59bc | 1.26fg | |
| E2 = 281 g a.i. ha−1 at V6 + 281 g a.i. ha−1 at V10 stage | 1.40def | 1.29fg | 1.12h | |
| E3 = 281 g a.i. ha−1 at V10 stage | 1.88a | 1.63b | 1.55bc | |
| E4 = 562 g a.i. ha−1 at V6 stage | 1.36ef | 1.31f | 1.16gh | |
| E5 = 562 g a.i. ha−1 at V6 + 562 g a.i. ha−1 at V10 stage | 1.30fg | 1.17gh | 1.06hi | |
| E6 = 562 g a.i. ha−1 at V10 stage | 1.81a | 1.50bcd | 1.47cde | |
| E7 = no ethephon | 1.89a | 1.06hi | 0.97i | |
| F test (W) | ** | |||
| LSD0.05 | 0.14 | |||
| Inputs | Input Amount | E1 | E2 | E3 | E4 | E5 | E6 | E7 | |
|---|---|---|---|---|---|---|---|---|---|
| Well watered (W1) | Nitrogen (N) | 194 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 |
| Phosphorus (P2O5) | 114.5 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | |
| Potassium (K2O) | 60.5 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | |
| Pesticides and PGR | E7 = 0.31; E1, E3 = 0.591; E2, E4, E6 = 0.872; E5 = 1.434 | 10.64 | 15.70 | 10.64 | 15.70 | 25.81 | 15.70 | 5.58 | |
| Diesel | 524.2 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | |
| Electricity | 720.1 | 576.08 | 576.08 | 576.08 | 576.08 | 576.08 | 576.08 | 576.08 | |
| Seed | 25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | |
| Total emission CO2-eq | 3768.28 | 3773.34 | 3768.28 | 3773.34 | 3783.45 | 3773.34 | 3763.22 | ||
| Short water stress (W2) | Nitrogen (N) | 194 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 |
| Phosphorus (P2O5) | 114.5 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | |
| Potassium (K2O) | 60.5 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | |
| Pesticides and PGR | E7 = 0.31; E1, E3 = 0.591; E2, E4, E6 = 0.872; E5 = 1.434 | 10.64 | 15.70 | 10.64 | 15.70 | 25.81 | 15.70 | 5.58 | |
| Diesel | 524.2 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | |
| Electricity | 617.23 | 493.78 | 493.78 | 493.78 | 493.78 | 493.78 | 493.78 | 493.78 | |
| Seed | 25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | |
| Total emission CO2-eq | 3685.98 | 3691.04 | 3685.98 | 3691.04 | 3701.16 | 3691.04 | 3680.93 | ||
| Prolong water stress (W3) | Nitrogen (N) | 194 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 | 1610.20 |
| Phosphorus (P2O5) | 114.5 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | 69.85 | |
| Potassium (K2O) | 60.5 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | 26.62 | |
| Pesticides and PGR | E7 = 0.31; E1, E3 = 0.591; E2, E4, E6 = 0.872; E5 = 1.434 | 10.64 | 15.70 | 10.64 | 15.70 | 25.81 | 15.70 | 5.58 | |
| Diesel | 524.2 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | 1378.65 | |
| Electricity | 514.36 | 411.49 | 411.49 | 411.49 | 411.49 | 411.49 | 411.49 | 411.49 | |
| Seed | 25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | 96.25 | |
| Total emission CO2-eq | 3603.69 | 3608.75 | 3603.69 | 3608.75 | 3618.86 | 3608.75 | 3598.63 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Molla, M.S.H.; Kumdee, O.; Worathongchai, N.; Khongchiu, P.; Ali, M.A.; Anwar, M.M.; Wongkaew, A.; Nakasathien, S. Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field. Agronomy 2023, 13, 2673. https://doi.org/10.3390/agronomy13112673
Molla MSH, Kumdee O, Worathongchai N, Khongchiu P, Ali MA, Anwar MM, Wongkaew A, Nakasathien S. Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field. Agronomy. 2023; 13(11):2673. https://doi.org/10.3390/agronomy13112673
Chicago/Turabian StyleMolla, Md. Samim Hossain, Orawan Kumdee, Nattaporn Worathongchai, Phanuphong Khongchiu, M. Akkas Ali, Md. Mazharul Anwar, Arunee Wongkaew, and Sutkhet Nakasathien. 2023. "Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field" Agronomy 13, no. 11: 2673. https://doi.org/10.3390/agronomy13112673
APA StyleMolla, M. S. H., Kumdee, O., Worathongchai, N., Khongchiu, P., Ali, M. A., Anwar, M. M., Wongkaew, A., & Nakasathien, S. (2023). Efforts to Stimulate Morpho-Physio-Biochemical Traits of Maize for Efficient Production under Drought Stress in Tropics Field. Agronomy, 13(11), 2673. https://doi.org/10.3390/agronomy13112673