
Breaking Buds—Stop and Go? Acid Invertase Activities in Apple Leaf Buds during Dormancy Release until Bud Break

{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Determination of the Adjusted Dry Weight [aDW]
2.3. Extraction Procedure for Intracellular and Cell Wall-Bound Proteins
2.4. Acid Invertase Activity Assay
2.5. Air Temperature
3. Results
3.1. Phenological Growth Stages of Apple Leaf Buds
3.2. Air Temperature
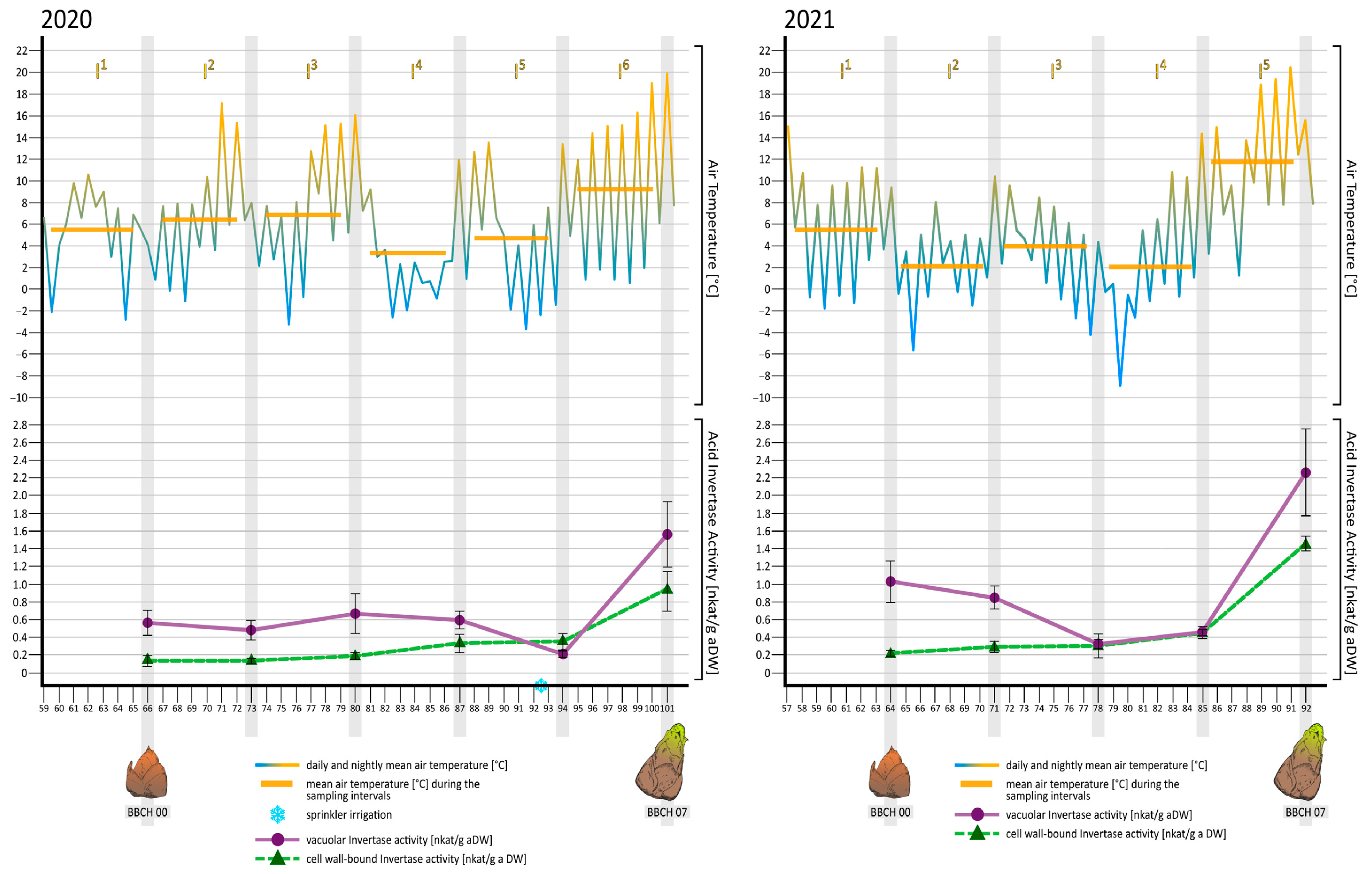
3.2.1. Air Temperature during the Observation Period in 2020
3.2.2. Air Temperature during the Observation Period in 2021
3.3. Acid Invertase Activities in Apple Leaf Buds
3.3.1. Acid Invertase Activities during the Observation Period in 2020
3.3.2. Acid Invertase Activities during the Observation Period in 2021
4. Discussion
5. Conclusions and Outlook
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lang, G.A. Dormancy: A New Universal Terminology. HortScience 1987, 22, 817–820. [Google Scholar] [CrossRef]
- Lang, G.A.; Early, J.D.; Martin, G.C.; Darnell, R.L. Endo-, Para-, and Ecodormancy: Physiological Terminology and Classification for Dormancy Research. HortScience 1987, 22, 371–377. [Google Scholar] [CrossRef]
- de Rosa, V.; Falchi, R.; Moret, E.; Vizzotto, G. Insight into Carbohydrate Metabolism and Signaling in Grapevine Buds during Dormancy Progression. Plants 2022, 11, 1027. [Google Scholar] [CrossRef] [PubMed]
- Leida, C.; Conejero, A.; Arbona, V.; Gómez-Cadenas, A.; Llácer, G.; Badenes, M.L.; Ríos, G. Chilling-dependent release of seed and bud dormancy in peach associates to common changes in gene expression. PLoS ONE 2012, 7, e35777. [Google Scholar] [CrossRef] [PubMed]
- Lempe, J.; Peil, A.; Flachowsky, H. Time-Resolved Analysis of Candidate Gene Expression and Ambient Temperature during Bud Dormancy in Apple. Front. Plant Sci. 2021, 12, 803341. [Google Scholar] [CrossRef]
- Naor, A.; Flaishman, M.; Stern, R.; Moshe, A.; Erez, A. Temperature Effects on Dormancy Completion of Vegetative Buds in Apple. J. Am. Soc. Hortic. Sci. 2003, 128, 636–641. [Google Scholar] [CrossRef]
- Heide, O.M.; Prestrud, A.K. Low temperature, but not photoperiod, controls growth cessation and dormancy induction and release in apple and pear. Tree Physiol. 2005, 25, 109–114. [Google Scholar] [CrossRef]
- Vitra, A.; Lenz, A.; Vitasse, Y. Frost hardening and dehardening potential in temperate trees from winter to budburst. New Phytol. 2017, 216, 113–123. [Google Scholar] [CrossRef]
- Bilavcik, A.; Zamecnik, J.; Faltus, M. Cryotolerance of apple tree bud is independent of endodormancy. Front. Plant Sci. 2015, 6, 695. [Google Scholar] [CrossRef]
- Faust, M.; Liu, D.; Millard, M.M.; Stutte, G.W. Bound versus Free Water in Dormant Apple Buds—A Theory for Endodormancy. HortScience 1991, 26, 887–890. [Google Scholar] [CrossRef]
- Erez, A.; Fishman, S.; Linsley-Noakes, G.C.; Allan, P. The Dynamic Model for Rest Completion in Peach Buds. Acta Hortic. 1990, 276, 165–174. [Google Scholar] [CrossRef]
- Farokhzad, A.; Nobakht, S.; Alahveran, A.; Sarkhosh, A.; Mohseniazar, M. Biochemical changes in terminal buds of three different walnut (Juglans regia L.) genotypes during dormancy break. Biochem. Syst. Ecol. 2018, 76, 52–57. [Google Scholar] [CrossRef]
- Ashworth, E.N. Properties of peach flower buds which facilitate supercooling. Plant Physiol. 1982, 70, 1475–1479. [Google Scholar] [CrossRef]
- Proebsting, E.L. The role of air temperatures and bud development in determining hardiness of dormant Elberta peach fruit buds. Proc. Amer. Soc. Hort. Sc. 1963, 83, 259–269. [Google Scholar]
- Chmielewski, F.-M.; Götz, K.-P. Metabolites in Cherry Buds to Detect Winter Dormancy. Metabolites 2022, 12, 247. [Google Scholar] [CrossRef]
- Chmielewski, F.M.; Götz, K.-P.; Homann, T.; Huschek, G.; Rawel, H.M. Identification of Endodormancy Release for Cherries (Prunus avium L.) by Abscisic Acid and Sugars. J. Hortic. 2017, 4, 1000210. [Google Scholar] [CrossRef]
- Vitasse, Y.; Lenz, A.; Körner, C. The interaction between freezing tolerance and phenology in temperate deciduous trees. Front. Plant Sci. 2014, 5, 541. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Reighard, G.L.; Abbott, A.G.; Bielenberg, D.G. Dormancy-associated MADS genes from the EVG locus of peach [Prunus persica (L.) Batsch] have distinct seasonal and photoperiodic expression patterns. J. Exp. Bot. 2009, 60, 3521–3530. [Google Scholar] [CrossRef]
- IPCC. 2022: Climate Change 2022: Impacts, Adaptation and Vulnerability. In Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner, H.-O., Roberts, D.C., Tignor, M., Poloczanska, E.S., Mintenbeck, K., Alegría, A., Craig, M., Langsdorf, S., Löschke, S., Möller, V., et al., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; p. 3056. [Google Scholar] [CrossRef]
- Sapkota, S.; Liu, J.; Islam, M.T.; Ravindran, P.; Kumar, P.P.; Sherif, S.M. Contrasting bloom dates in two apple cultivars linked to differential levels of phytohormones and heat requirements during ecodormancy. Sci. Hortic. 2021, 288, 110413. [Google Scholar] [CrossRef]
- Unterberger, C.; Brunner, L.; Nabernegg, S.; Steininger, K.W.; Steiner, A.K.; Stabentheiner, E.; Monschein, S.; Truhetz, H. Spring frost risk for regional apple production under a warmer climate. PLoS ONE 2018, 13, e0200201. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAOSTA: Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data (accessed on 24 September 2023).
- Olhnuud, A.; Liu, Y.; Makowski, D.; Tscharntke, T.; Westphal, C.; Wu, P.; Wang, M.; van der Werf, W. Pollination deficits and contributions of pollinators in apple production: A global meta-analysis. J. Appl. Ecol. 2022, 59, 2911–2921. [Google Scholar] [CrossRef]
- Fadón, E.; Fernandez, E.; Behn, H.; Luedeling, E. A Conceptual Framework for Winter Dormancy in Deciduous Trees. Agronomy 2020, 10, 241. [Google Scholar] [CrossRef]
- Beauvieux, R.; Wenden, B.; Dirlewanger, E. Bud Dormancy in Perennial Fruit Tree Species: A Pivotal Role for Oxidative Cues. Front. Plant Sci. 2018, 9, 657. [Google Scholar] [CrossRef]
- Tixier, A.; Gambetta, G.A.; Godfrey, J.; Orozco, J.; Zwieniecki, M.A. Non-structural Carbohydrates in Dormant Woody Perennials; The Tale of Winter Survival and Spring Arrival. Front. For. Glob. Chang. 2019, 2, 18. [Google Scholar] [CrossRef]
- Götz, K.-P.; Chmielewski, F.-M. Response of sweet cherry buds and twigs to temperature changes—Evaluated by the determination of the degradation and synthesis of sucrose. Hortic. Sci. 2021, 48, 149–157. [Google Scholar] [CrossRef]
- Liang, D.; Huang, X.; Shen, Y.; Shen, T.; Zhang, H.; Lin, L.; Wang, J.; Deng, Q.; Lyu, X.; Xia, H. Hydrogen cyanamide induces grape bud endodormancy release through carbohydrate metabolism and plant hormone signaling. BMC Genom. 2019, 20, 1034. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, W.; Gao, Z.; Wen, L.; Huo, X.; Cai, B.; Zhang, Z. Metabolic changes upon flower bud break in Japanese apricot are enhanced by exogenous GA4. Hortic. Res. 2015, 2, 15046. [Google Scholar] [CrossRef]
- Marafon, A.C.; Citadin, I.; Amarante, L.d.; Herter, F.G.; Hawerroth, F.J. Chilling privation during dormancy period and carbohydrate mobilization in Japanese pear trees. Sci. Agric. 2011, 68, 462–468. [Google Scholar] [CrossRef]
- Girault, T.; Abidi, F.; Sigogne, M.; Pelleschi-Travier, S.; Boumaza, R.; Sakr, S.; Leduc, N. Sugars are under light control during bud burst in Rosa sp. Plant Cell Environ. 2010, 33, 1339–1350. [Google Scholar] [CrossRef]
- Bonhomme, M.; Rageau, R.; Lacointe, A.; Gendraud, M. Influences of cold deprivation during dormancy on carbohydrate contents of vegetative and floral primordia and nearby structures of peach buds (Prunus persica L. Batch). Sci. Hortic. 2005, 105, 223–240. [Google Scholar] [CrossRef]
- Marquat, C.; Vandamme, M.; Gendraud, M.; Pétel, G. Dormancy in vegetative buds of peach: Relation between carbohydrate absorption potentials and carbohydrate concentration in the bud during dormancy and its release. Sci. Hortic. 1999, 79, 151–162. [Google Scholar] [CrossRef]
- Hussain, S.; Liu, G.; Liu, D.; Ahmed, M.; Hussain, N.; Teng, Y. Study on the expression of dehydrin genes and activities of antioxidative enzymes in floral buds of two sand pear (Pyrus pyrifolia Nakai) cultivars requiring different chilling hours for bud break. Turk. J. Agric. For. 2015, 39, 930–939. [Google Scholar] [CrossRef]
- Ito, A.; Hayama, H.; Kashimura, Y. Sugar metabolism in buds during flower bud formation: A comparison of two Japanese pear [Pyrus pyrifolia (Burm.) Nak.] cultivars possessing different flowering habits. Sci. Hortic. 2002, 96, 163–175. [Google Scholar] [CrossRef]
- Ben Mohamed, H.; Vadel, A.M.; Geuns, J.M.; Khemira, H. Carbohydrate changes during dormancy release in Superior Seedless grapevine cuttings following hydrogen cyanamide treatment. Sci. Hortic. 2012, 140, 19–25. [Google Scholar] [CrossRef]
- Peng, Y.; Zhu, L.; Tian, R.; Wang, L.; Su, J.; Yuan, Y.; Ma, F.; Li, M.; Ma, B. Genome-wide identification, characterization and evolutionary dynamic of invertase gene family in apple, and revealing its roles in cold tolerance. Int. J. Biol. Macromol. 2023, 229, 766–777. [Google Scholar] [CrossRef] [PubMed]
- Roitsch, T.; González, M.-C. Function and regulation of plant invertases: Sweet sensations. Trends Plant Sci. 2004, 9, 606–613. [Google Scholar] [CrossRef]
- Klotz, K.L.; Finger, F.L.; Shelver, W.L. Characterization of two sucrose synthase isoforms in sugarbeet root. Plant Physiol. Biochem. 2003, 41, 107–115. [Google Scholar] [CrossRef]
- Sturm, A.; Tang, G.Q. The sucrose-cleaving enzymes of plants are crucial for development, growth and carbon partitioning. Trends Plant Sci. 1999, 4, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Wu, L.; Yang, Y.; Zhou, G.; Ruan, Y.-L. Evolution of Sucrose Metabolism: The Dichotomy of Invertases and Beyond. Trends Plant Sci. 2018, 23, 163–177. [Google Scholar] [CrossRef]
- Braun, D.M.; Wang, L.; Ruan, Y.-L. Understanding and manipulating sucrose phloem loading, unloading, metabolism, and signalling to enhance crop yield and food security. J. Exp. Bot. 2014, 65, 1713–1735. [Google Scholar] [CrossRef]
- Ruan, Y.-L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L.; Jin, Y.; Yang, Y.-J.; Li, G.-J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Sturm, A. Invertases. Primary structures, functions, and roles in plant development and sucrose partitioning. Plant Physiol. 1999, 121, 1–8. [Google Scholar] [CrossRef] [PubMed]
- González, M.-C.; Roitsch, T.; Cejudo, F.J. Circadian and developmental regulation of vacuolar invertase expression in petioles of sugar beet plants. Planta 2005, 222, 386–395. [Google Scholar] [CrossRef]
- Proels, R.K.; Roitsch, T. Extracellular invertase LIN6 of tomato: A pivotal enzyme for integration of metabolic, hormonal, and stress signals is regulated by a diurnal rhythm. J. Exp. Bot. 2009, 60, 1555–1567. [Google Scholar] [CrossRef] [PubMed]
- Ahiakpa, J.K.; Karikari, B.; Magdy, M.; Munir, S.; Mumtaz, M.A.; Li, F.; Wang, Y.; Shang, L.; Zhang, Y. Regulation of invertase and sucrose for improving tomato fruit flavor: A review. Veg. Res. 2021, 1, 10. [Google Scholar] [CrossRef]
- Ruan, Y.-L.; Jin, Y.; Huang, J. Capping invertase activity by its inhibitor: Roles and implications in sugar signaling, carbon allocation, senescence and evolution. Plant Signal. Behav. 2009, 4, 983–985. [Google Scholar] [CrossRef]
- Coluccio Leskow, C.; Conte, M.; Del Pozo, T.; Bermúdez, L.; Lira, B.S.; Gramegna, G.; Baroli, I.; Burgos, E.; Zavallo, D.; Kamenetzky, L.; et al. The cytosolic invertase NI6 affects vegetative growth, flowering, fruit set, and yield in tomato. J. Exp. Bot. 2021, 72, 2525–2543. [Google Scholar] [CrossRef]
- Qian, W.; Xiao, B.; Wang, L.; Hao, X.; Yue, C.; Cao, H.; Wang, Y.; Li, N.; Yu, Y.; Zeng, J.; et al. CsINV5, a tea vacuolar invertase gene enhances cold tolerance in transgenic Arabidopsis. BMC Plant Biol. 2018, 18, 228. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L. CWIN-sugar transporter nexus is a key component for reproductive success. J. Plant Physiol. 2022, 268, 153572. [Google Scholar] [CrossRef]
- Eschrich, W. Free Space Invertase, its Possible Role in Phloem Unloading1. Ber. Dtsch. Bot. Ges. 1980, 93, 363–378. [Google Scholar] [CrossRef]
- Koch, K. Sucrose metabolism: Regulatory mechanisms and pivotal roles in sugar sensing and plant development. Curr. Opin. Plant Biol. 2004, 7, 235–246. [Google Scholar] [CrossRef]
- Winter, H.; Huber, S.C. Regulation of sucrose metabolism in higher plants: Localization and regulation of activity of key enzymes. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 253–289. [Google Scholar] [CrossRef] [PubMed]
- Tymowska-Lalanne, Z.; Kreis, M. The Plant Invertases: Physiology, Biochemistry and Molecular Biology. Adv. Bot. Res. 1998, 28, 71–117. [Google Scholar] [CrossRef]
- Zhu, J.; Qi, J.; Fang, Y.; Xiao, X.; Li, J.; Lan, J.; Tang, C. Characterization of Sugar Contents and Sucrose Metabolizing Enzymes in Developing Leaves of Hevea brasiliensis. Front. Plant Sci. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Pantin, F.; Simonneau, T.; Muller, B. Coming of leaf age: Control of growth by hydraulics and metabolics during leaf ontogeny. New Phytol. 2012, 196, 349–366. [Google Scholar] [CrossRef]
- Le Hir, R.; Leduc, N.; Jeannette, E.; Viemont, J.-D.; Pelleschi-Travier, S. Variations in sucrose and ABA concentrations are concomitant with heteroblastic leaf shape changes in a rhythmically growing species (Quercus robur). Tree Physiol. 2006, 26, 229–238. [Google Scholar] [CrossRef]
- He, X.; Wei, Y.; Kou, J.; Xu, F.; Chen, Z.; Shao, X. PpVIN2, an acid invertase gene family member, is sensitive to chilling temperature and affects sucrose metabolism in postharvest peach fruit. Plant Growth Regul. 2018, 86, 169–180. [Google Scholar] [CrossRef]
- Xu, X.-X.; Hu, Q.; Yang, W.-N.; Jin, Y. The roles of call wall invertase inhibitor in regulating chilling tolerance in tomato. BMC Plant Biol. 2017, 17, 195. [Google Scholar] [CrossRef]
- Wang, L.; Zheng, Y.; Ding, S.; Zhang, Q.; Chen, Y.; Zhang, J. Molecular cloning, structure, phylogeny and expression analysis of the invertase gene family in sugarcane. BMC Plant Biol. 2017, 17, 109. [Google Scholar] [CrossRef]
- Wang, X.; Chen, Y.; Jiang, S.; Xu, F.; Wang, H.; Wei, Y.; Shao, X. PpINH1, an invertase inhibitor, interacts with vacuolar invertase PpVIN2 in regulating the chilling tolerance of peach fruit. Hortic. Res. 2020, 7, 168. [Google Scholar] [CrossRef]
- McKenzie, M.J.; Chen, R.K.Y.; Harris, J.C.; Ashworth, M.J.; Brummell, D.A. Post-translational regulation of acid invertase activity by vacuolar invertase inhibitor affects resistance to cold-induced sweetening of potato tubers. Plant Cell Environ. 2013, 36, 176–185. [Google Scholar] [CrossRef]
- Hubmann, A.M.; Jammer, A.; Monschein, S. Activities of H2O2-Converting Enzymes in Apple Leaf Buds during Dormancy Release in Consideration of the Ratio Change between Bud Scales and Physiologically Active Tissues. Horticulturae 2022, 8, 982. [Google Scholar] [CrossRef]
- Jammer, A.; Gasperl, A.; Luschin-Ebengreuth, N.; Heyneke, E.; Chu, H.; Cantero-Navarro, E.; Großkinsky, D.K.; Albacete, A.A.; Stabentheiner, E.; Franzaring, J.; et al. Simple and robust determination of the activity signature of key carbohydrate metabolism enzymes for physiological phenotyping in model and crop plants. J. Exp. Bot. 2015, 66, 5531–5542. [Google Scholar] [CrossRef]
- Fimognari, L.; Dölker, R.; Kaselyte, G.; Jensen, C.N.G.; Akhtar, S.S.; Großkinsky, D.K.; Roitsch, T. Simple semi-high throughput determination of activity signatures of key antioxidant enzymes for physiological phenotyping. Plant Methods 2020, 16, 42. [Google Scholar] [CrossRef] [PubMed]
- Federal Institute for Geology, Geophysics, Climatology and Meteorology. Auf- und Untergangszeiten von Sonne und Mond, Ephemeriden in Graz für Das Jahr 2021. Available online: https://www.zamg.ac.at/cms/de/klima/klimauebersichten/ephemeriden/graz?jahr=2021 (accessed on 24 September 2023).
- Federal Institute for Geology, Geophysics, Climatology and Meteorology. Auf- und Untergangszeiten von Sonne und Mond, Ephemeriden in Graz für das Jahr 2020. Available online: https://www.zamg.ac.at/cms/de/klima/klimauebersichten/ephemeriden/graz?jahr=2020 (accessed on 24 September 2023).
- Meier, U.; Graf, H.; Hack, H.; Heß, M.; Kennel, W.; Klose, R.; Mappes, D.; Seipp, D.; Strauß, R.; Streif, J.; et al. Phänologische Entwicklungsstadien des Kernobstes (Malus domestica Borkh. und Pyrus communis L.), des Steinobstes (Prunus-Arten), der Johannisbeere (Ribes-Arten) und der Erdbeere (Fragaria ananassa Duch.). Nachr. Dtsch Pflanzenschutzd. 1994, 46, 141–153. [Google Scholar]
- Baumgarten, F.; Zohner, C.M.; Gessler, A.; Vitasse, Y. Chilled to be forced: The best dose to wake up buds from winter dormancy. New Phytol. 2021, 230, 1366–1377. [Google Scholar] [CrossRef]
- Luedeling, E.; Kunz, A.; Blanke, M.M. Identification of chilling and heat requirements of cherry trees—A statistical approach. Int. J. Biometeorol. 2013, 57, 679–689. [Google Scholar] [CrossRef]
- Eccel, E.; Rea, R.; Caffarra, A.; Crisci, A. Risk of spring frost to apple production under future climate scenarios: The role of phenological acclimation. Int. J. Biometeorol. 2009, 53, 273–286. [Google Scholar] [CrossRef]
- Viti, R.; Bartolini, S.; Andreini, L. Apricot flower bud dormancy: Main morphological, anatomical and physiological features related to winter climate influence. Adv. Hortic. Sci. 2013, 27, 5–17. [Google Scholar]
- Cai, F.; Jin, X.; Tian, Y.; Huang, Z.; Wang, X.; Zhang, Y.; Sun, Y.; Shao, C. Molecular regulation of bud dormancy in perennial plants. Plant Growth Regul. 2023. [Google Scholar] [CrossRef]
- Yamane, H.; Singh, A.K.; Cooke, J.E.K. Plant dormancy research: From environmental control to molecular regulatory networks. Tree Physiol. 2021, 41, 523–528. [Google Scholar] [CrossRef]
- Yamane, H.; Andrés, F.; Bai, S.; Luedeling, E.; Or, E. Editorial: Environmental and molecular control of bud dormancy and bud break in woody perennials: An integrative approach. Front. Plant Sci. 2023, 14, 1104108. [Google Scholar] [CrossRef] [PubMed]
- Noguer, C.G.; Delgado, A.; Else, M.; Hadley, P. Apple (Malus × domestica Borkh.) dormancy—A review of regulatory mechanisms and agroclimatic requirements. Front. Hortic. 2023, 2, 1217689. [Google Scholar] [CrossRef]
- Maurya, J.P.; Bhalerao, R.P. Photoperiod- and temperature-mediated control of growth cessation and dormancy in trees: A molecular perspective. Ann. Bot. 2017, 120, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Cooke, J.E.K.; Eriksson, M.E.; Junttila, O. The dynamic nature of bud dormancy in trees: Environmental control and molecular mechanisms. Plant Cell Environ. 2012, 35, 1707–1728. [Google Scholar] [CrossRef]
- Heide, O.M. Interaction of photoperiod and temperature in the control of growth and dormancy of Prunus species. Sci. Hortic. 2008, 115, 309–314. [Google Scholar] [CrossRef]
- Zhu, L.H.; Borsboom, O.; Tromp, J. The effect of temperature on flower-bud formation in apple including some morphological aspects. Sci. Hortic. 1997, 70, 1–8. [Google Scholar] [CrossRef]
- Thakur, P.; Kumar, S.; Malik, J.A.; Berger, J.D.; Nayyar, H. Cold stress effects on reproductive development in grain crops: An overview. Environ. Exp. Bot. 2010, 67, 429–443. [Google Scholar] [CrossRef]
- Jammer, A.; Akhtar, S.S.; Amby, D.B.; Pandey, C.; Mekureyaw, M.F.; Bak, F.; Roth, P.M.; Roitsch, T. Enzyme activity profiling for physiological phenotyping within functional phenomics: Plant growth and stress responses. J. Exp. Bot. 2022, 73, 5170–5198. [Google Scholar] [CrossRef]
- Jammer, A.; Albacete, A.; Schulz, B.; Koch, W.; Weltmeier, F.; van der Graaff, E.; Pfeifhofer, H.W.; Roitsch, T.G. Early-stage sugar beet taproot development is characterized by three distinct physiological phases. Plant Direct 2020, 4, e00221. [Google Scholar] [CrossRef]
- Biais, B.; Bénard, C.; Beauvoit, B.; Colombié, S.; Prodhomme, D.; Ménard, G.; Bernillon, S.; Gehl, B.; Gautier, H.; Ballias, P.; et al. Remarkable reproducibility of enzyme activity profiles in tomato fruits grown under contrasting environments provides a roadmap for studies of fruit metabolism. Plant Physiol. 2014, 164, 1204–1221. [Google Scholar] [CrossRef]
- Marchi, S.; Sebastiani, L.; Gucci, R.; Tognetti, R. Sink-source Transition in Peach Leaves during Shoot Development. J. Am. Soc. Hortic. Sci. 2005, 130, 928–935. [Google Scholar] [CrossRef]
- Turgeon, R. The Sink-Source Transition in Leaves. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1989, 40, 119–138. [Google Scholar] [CrossRef]
- Maurel, K.; Leite, G.B.; Bonhomme, M.; Guilliot, A.; Rageau, R.; Pétel, G.; Sakr, S. Trophic control of bud break in peach (Prunus persica) trees: A possible role of hexoses. Tree Physiol. 2004, 24, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Eguchi, Y.; Inada, Y.; Onishi, K.; Hishikawa, K. Successive Induction of Invertase Isoforms During Flower Development in Eustoma. Hort. J. 2021, 90, 334–340. [Google Scholar] [CrossRef]
- Zanol, G.C.; Bartolini, S. Changes in intracellular pH in apricot buds during the winter season. Adv. Hortic. Sci. 2003, 17, 97–101. [Google Scholar]
- Rady, M.M.; Seif El-Yazal, M.A. Response of “Anna” apple dormant buds and carbohydrate metabolism during floral bud break to onion extract. Sci. Hortic. 2013, 155, 78–84. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Wang, X.-L.; Wang, X.-F.; Xia, G.-H.; Pan, Q.-H.; Fan, R.-C.; Wu, F.-Q.; Yu, X.-C.; Zhang, D.-P. A shift of Phloem unloading from symplasmic to apoplasmic pathway is involved in developmental onset of ripening in grape berry. Plant Physiol. 2006, 142, 220–232. [Google Scholar] [CrossRef]
- Chourey, P.S.; Jain, M.; Li, Q.-B.; Carlson, S.J. Genetic control of cell wall invertases in developing endosperm of maize. Planta 2006, 223, 159–167. [Google Scholar] [CrossRef]
- Signorelli, S.; Shaw, J.; Hermawaty, D.; Wang, Z.; Verboven, P.; Considine, J.A.; Considine, M.J. The initiation of bud burst in grapevine features dynamic regulation of the apoplastic pore size. J. Exp. Bot. 2020, 71, 719–729. [Google Scholar] [CrossRef]
- Kutschera, U. Cell Expansion in Plant Development. Braz. J. Plant Physiol. 2000, 12, 65–95. [Google Scholar]
- Feng, Z.; Zheng, F.; Wu, S.; Li, R.; Li, Y.; Zhong, J.; Zhao, H. Functional Characterization of a Cucumber (Cucumis sativus L.) Vacuolar Invertase, CsVI1, Involved in Hexose Accumulation and Response to Low Temperature Stress. Int. J. Mol. Sci. 2021, 22, 9365. [Google Scholar] [CrossRef]
- Wang, L.; Cook, A.; Patrick, J.W.; Chen, X.-Y.; Ruan, Y.-L. Silencing the vacuolar invertase gene GhVIN1 blocks cotton fiber initiation from the ovule epidermis, probably by suppressing a cohort of regulatory genes via sugar signaling. Plant J. 2014, 78, 686–696. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, X.-R.; Lian, H.; Ni, D.-A.; He, Y.; Chen, X.-Y.; Ruan, Y.-L. Evidence that high activity of vacuolar invertase is required for cotton fiber and Arabidopsis root elongation through osmotic dependent and independent pathways, respectively. Plant Physiol. 2010, 154, 744–756. [Google Scholar] [CrossRef]
- Kutschera, U.; Niklas, K.J. Cell division and turgor-driven stem elongation in juvenile plants: A synthesis. Plant Sci. 2013, 207, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Gao, K.; Su, X.; Rao, P.; An, X. Genome-Wide Identification of the Invertase Gene Family in Populus. PLoS ONE 2015, 10, e0138540. [Google Scholar] [CrossRef]
- Bhaskar, P.B.; Wu, L.; Busse, J.S.; Whitty, B.R.; Hamernik, A.J.; Jansky, S.H.; Buell, C.R.; Bethke, P.C.; Jiang, J. Suppression of the vacuolar invertase gene prevents cold-induced sweetening in potato. Plant Physiol. 2010, 154, 939–948. [Google Scholar] [CrossRef]
- Usadel, B.; Bläsing, O.E.; Gibon, Y.; Poree, F.; Höhne, M.; Günter, M.; Trethewey, R.; Kamlage, B.; Poorter, H.; Stitt, M. Multilevel genomic analysis of the response of transcripts, enzyme activities and metabolites in Arabidopsis rosettes to a progressive decrease of temperature in the non-freezing range. Plant Cell Environ. 2008, 31, 518–547. [Google Scholar] [CrossRef] [PubMed]
- Dahro, B.; Wang, Y.; Alhag, A.; Li, C.; Guo, D.; Liu, J.-H. Genome-wide identification and expression profiling of invertase gene family for abiotic stresses tolerance in Poncirus trifoliata. BMC Plant Biol. 2021, 21, 559. [Google Scholar] [CrossRef]
- Ashworth, E.N.; Davis, G.A.; Wisniewski, M.E. The formation and distribution of ice within dormant and deacclimated peach flower buds. Plant Cell Environ. 1989, 12, 521–528. [Google Scholar] [CrossRef]
- Kalberer, S.R.; Wisniewski, M.; Arora, R. Deacclimation and reacclimation of cold-hardy plants: Current understanding and emerging concepts. Plant Sci. 2006, 171, 3–16. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hubmann, A.M.; Roth, A.; Monschein, S. Breaking Buds—Stop and Go? Acid Invertase Activities in Apple Leaf Buds during Dormancy Release until Bud Break. Agronomy 2023, 13, 2659. https://doi.org/10.3390/agronomy13102659
Hubmann AM, Roth A, Monschein S. Breaking Buds—Stop and Go? Acid Invertase Activities in Apple Leaf Buds during Dormancy Release until Bud Break. Agronomy. 2023; 13(10):2659. https://doi.org/10.3390/agronomy13102659
Chicago/Turabian StyleHubmann, Anna M., Alexandra Roth, and Stephan Monschein. 2023. "Breaking Buds—Stop and Go? Acid Invertase Activities in Apple Leaf Buds during Dormancy Release until Bud Break" Agronomy 13, no. 10: 2659. https://doi.org/10.3390/agronomy13102659
APA StyleHubmann, A. M., Roth, A., & Monschein, S. (2023). Breaking Buds—Stop and Go? Acid Invertase Activities in Apple Leaf Buds during Dormancy Release until Bud Break. Agronomy, 13(10), 2659. https://doi.org/10.3390/agronomy13102659