

Synergistic Interaction of Rhizobium tropici, Rhizophagus irregularis and Serendipita indica in Promoting Snap Bean Growth
 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant, Microbial Materials and Experimental Site
2.2. Plant Growth
- (1)
- Non-inoculated plants (control),
- (2)
- Plants inoculated with R. tropici CIAT 899 (C),
- (3)
- Plants inoculated with R. tropici CIAT 899 and Rhizophagus irregularis (C+Ri),
- (4)
- Plants inoculated with R. tropici CIAT 899 and Serendipita indica (C+Si),
- (5)
- Plants inoculated with R. tropici CIAT 899, Serendipita indica, and Rhizophagus irregularis (C+Ri+Si).
2.2.1. Inoculation with Bacterial Strains
2.2.2. Enumeration of Autochthonous Rhizobia
2.3. Studied Parameters
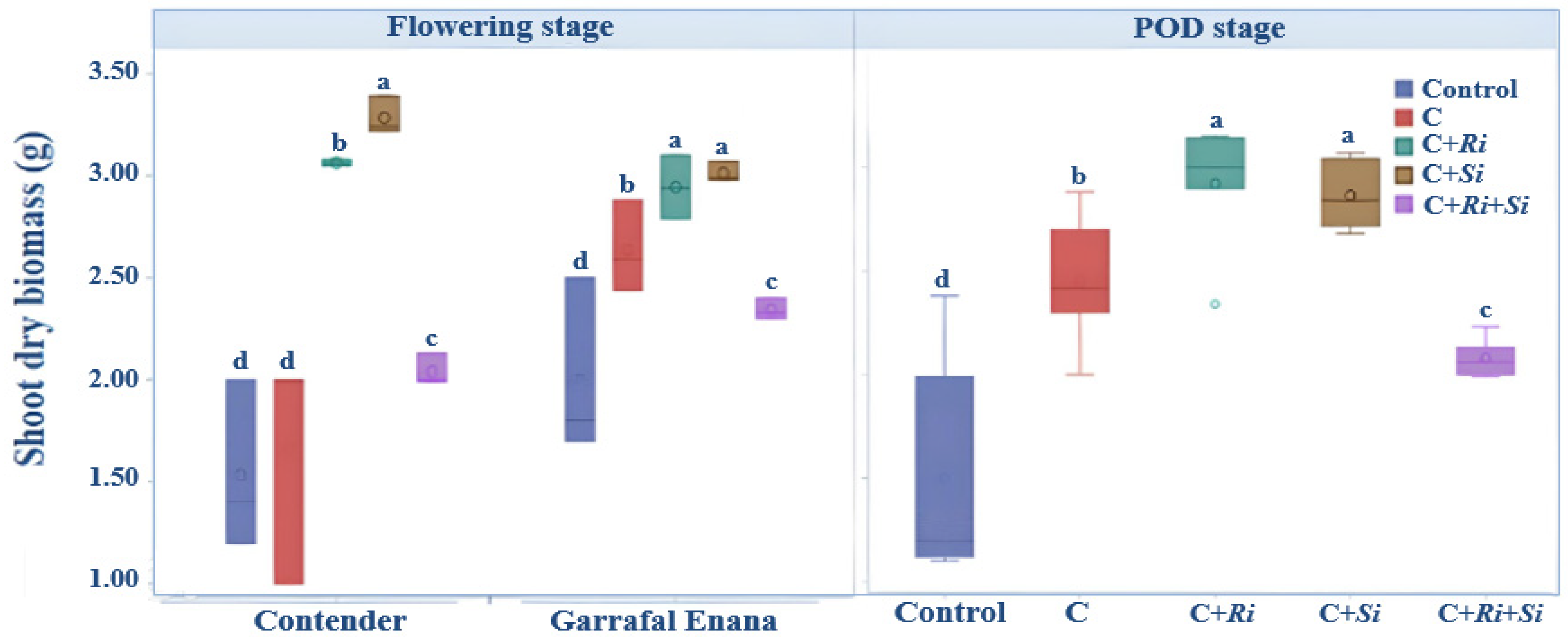
2.3.1. Biomass Yield
2.3.2. Nodulation
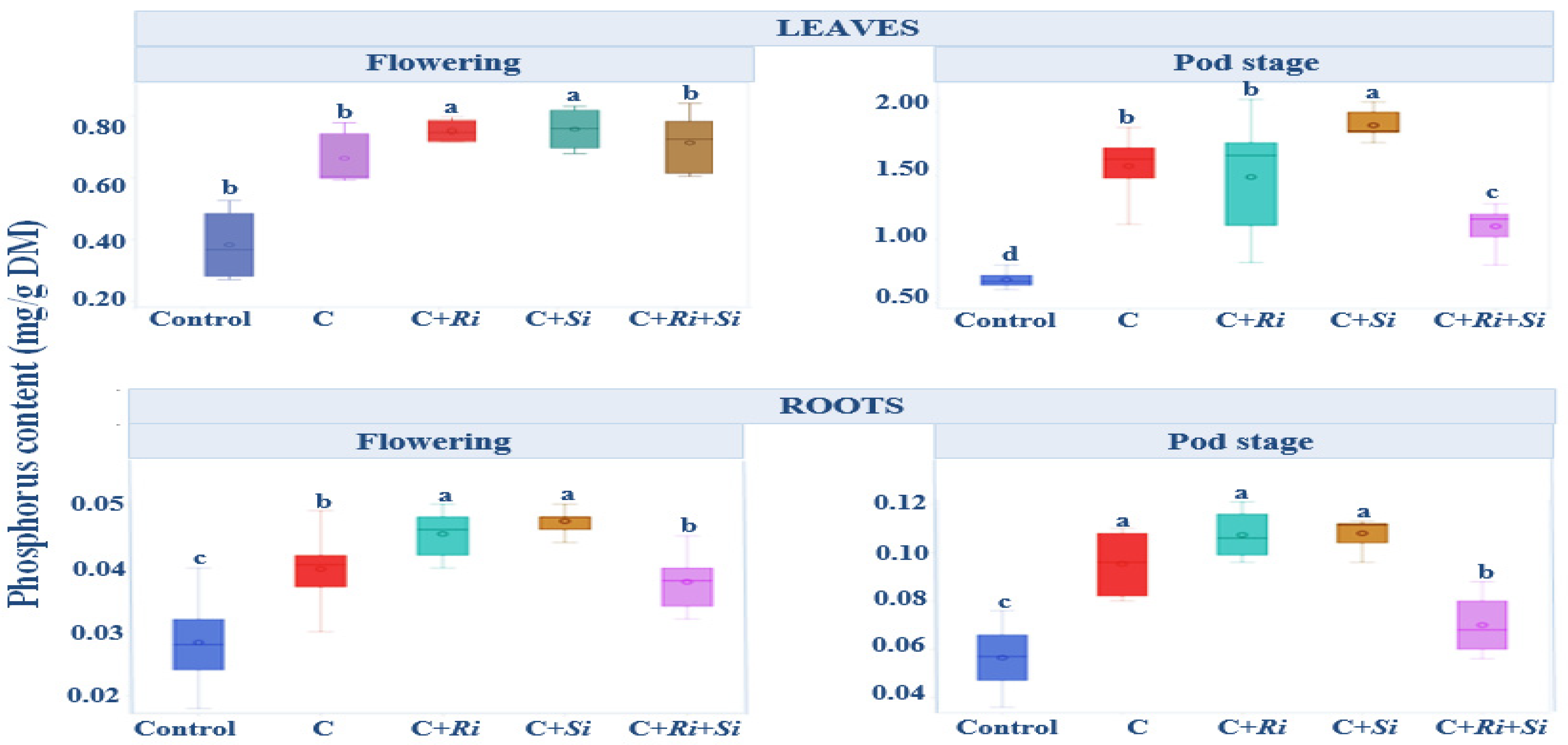
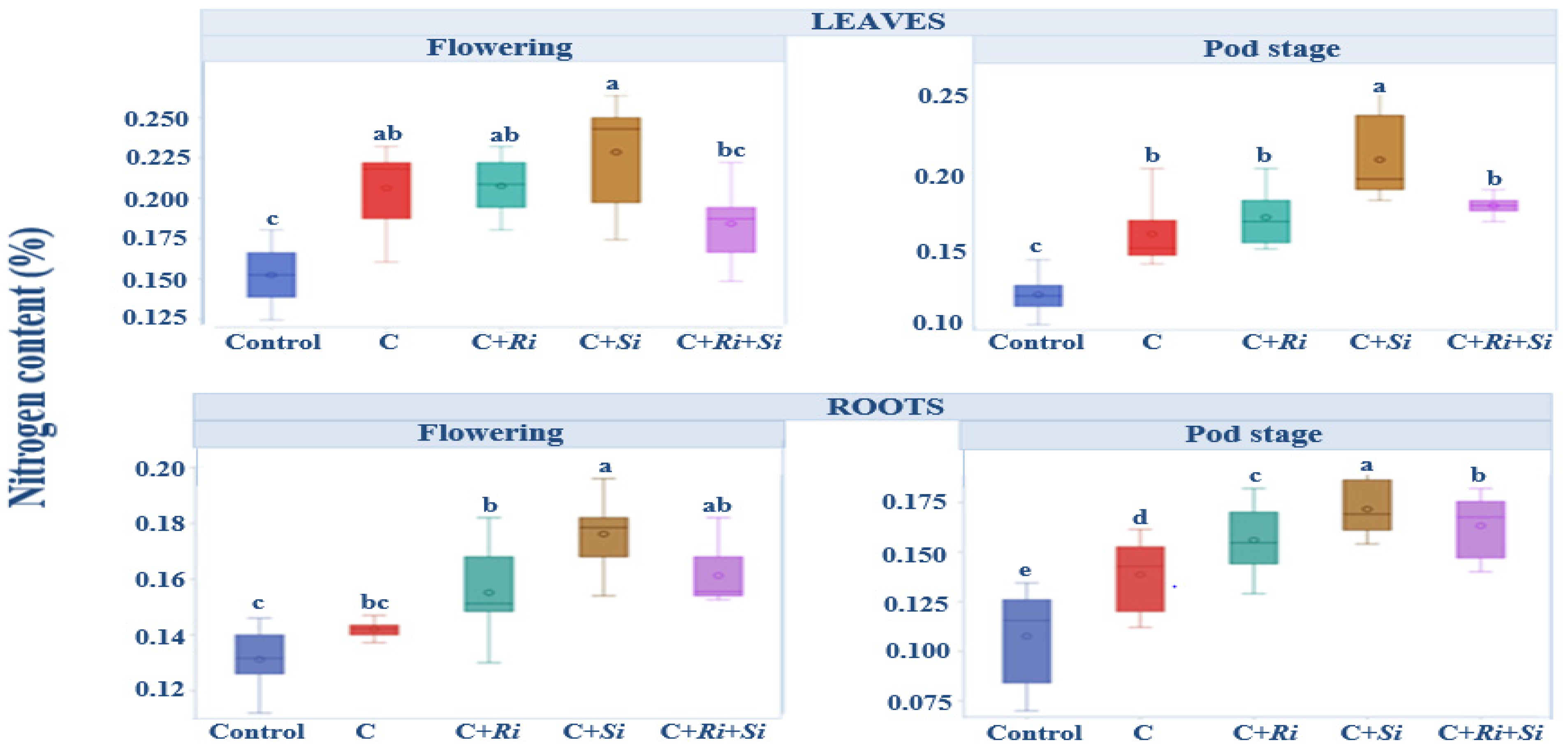
2.3.3. Nitrogen and Phosphorus Contents
2.3.4. Mycorrhization Rate
2.4. Statistical Analysis
3. Results
3.1. Plant Growth Parameters
3.1.1. Shoot Dry Biomass
3.1.2. Root Dry Mass
3.1.3. Number of Nodules
3.1.4. POD Number
3.1.5. Percent Mycorrhization
3.1.6. Nitrogen Content
3.1.7. Phosphorus Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mutari, B.; Sibiya, J.; Matova, P.M.; Gasura, E.; Simango, K. Drought stress impact on agronomic, shoot, physiological, canning and nutritional quality traits of navy beans (Phaseolus vulgaris L.) under field conditions in Zimbabwe. Field Crops Res. 2023, 292, 108826. [Google Scholar] [CrossRef]
- de Almeida Costa, G.E.; da Silva Queiroz-Monici, K.; Reis, S.M.P.M.; de Oliveira, A.C. Chemical composition, dietary fibre and resistant starch contents of raw and cooked pea, common bean, chickpea and lentil legumes. Food Chem. 2006, 94, 327–330. [Google Scholar] [CrossRef]
- El Sheikha, A.F.; Allam, A.Y.; Taha, M.; Varzakas, T. How Does the Addition of Biostimulants Affect the Growth, Yield, and Quality Parameters of the Snap Bean (Phaseolus vulgaris L.)? How Is This Reflected in Its Nutritional Value? Appl. Sci. 2022, 12, 776. [Google Scholar] [CrossRef]
- Mecha, E.; Figueira, M.E.; Vaz Patto, M.C.; do Rosário Bronze, M. Two sides of the same coin: The impact of grain legumes on human health: Common bean (Phaseolus vulgaris L.) as a case study. In Legume Seed Nutraceutical Research; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef]
- Ratna, B.; Sajia, S.; Sandip, M.; Kaynath, A.; Limon, D.; Kamruzzaman, M.; Akhtaruzzaman Khan, M. Production risk and 509 technical inefficiency of bean (Phaseolus vulgaris) cultivation in Bangladesh: Do socio-economic factors matter? Soc. Sci. Humanit. Open 2023, 7, 1. [Google Scholar] [CrossRef]
- Raza, T.; Qureshi, K.N.; Imran, S.; Eash, N.S.; Bortone, I. Associated health risks from heavy metal-laden effluent into point drainage channels in Faisalabad. Pak. J. Agric. Sci. 2021, 34, 487–494. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT Database. Pesticides Use. 2022. Available online: http://www.fao.org/faostat/en/#data/RP (accessed on 20 August 2023).
- FAO. Pesticides Use, Pesticides Trade and Pesticides Indicators—Global, Regional and Country Trends, 1990–2020; FAOSTAT Analytical Briefs no. 46; FAO: Rome, Italy, 2022. [Google Scholar] [CrossRef]
- Raza, T.; Abbas, M.; Amna; Imran, S.; Khan, M.Y.; Rebi, A.; Rafie-Rad, Z.; Eash, N.S. Impact of silicon on plant nutrition and significance of silicon mobilizing bacteria in agronomic practices. Silicon 2023, 15, 3797–3817. [Google Scholar] [CrossRef]
- Gavrilescu, M. Water, Soil, and Plants Interactions in a Threatened Environment. Water 2021, 13, 2746. [Google Scholar] [CrossRef]
- Wahab, A.; Muhammad, M.; Munir, A.; Abdi, G.; Zaman, W.; Ayaz, A.; Khizar, C.; Reddy, S.P.P. Role of Arbuscular Mycorrhizal Fungi in Regulating Growth, Enhancing Productivity, and Potentially Influencing Ecosystems under Abiotic and Biotic Stresses. Plants 2023, 12, 3102. [Google Scholar] [CrossRef]
- Álvaro Santana, M.; Ruth, R.R.; Antonio, V.; Herrera, H.; José Elías, C.G.; Bárbara, S.R. Food safety assessment of wines commercialized in the Canary Islands by monitoring of pesticide residues from 2017 to 2019. Food Control 2023, 153, 109957. [Google Scholar] [CrossRef]
- Bhanse, P.; Kumar, M.; Singh, L.; Awasthi, M.K.; Qureshi, A. Role of plant growth-promoting rhizobacteria in boosting the phytoremediation of stressed soils: Opportunities, challenges, and prospects. Chemosphere 2022, 303, 134954. [Google Scholar] [CrossRef]
- Naamala, J.; Smith, D.L. Relevance of Plant Growth Promoting Microorganisms and Their Derived Compounds, in the Face of Climate Change. Agronomy 2020, 10, 1179. [Google Scholar] [CrossRef]
- Gerke, J. The Central Role of Soil Organic Matter in Soil Fertility and Carbon Storage. Soil Syst. 2022, 6, 33. [Google Scholar] [CrossRef]
- Murindangabo, Y.T.; Kopecký, M.; Perná, K.; Nguyen, T.G.; Ghorbani, M.; Konvalina, P.; Bohatá, A.; Kavková, M.; Hoang, T.N.; Kabelka, D.; et al. Enhancing Soil Organic Matter Transformation through Sustainable Farming Practices: Evaluating Labile Soil Organic Matter Fraction Dynamics and Identifying Potential Early Indicators. Agriculture 2023, 13, 1314. [Google Scholar] [CrossRef]
- Raza, T.; Khan, M.Y.; Nadeem, S.M.; Imran, S.; Qureshi, K.N.; Mushtaq, M.N.; Sohaib, M.; Schmalenberger, A.; Eash, N.S. Biological management of selected weeds of wheat through co-application of allelopathic rhizobacteria and sorghum extract. Biol. Control 2021, 164, 104775. [Google Scholar] [CrossRef]
- Syed, A.; Raza, T.; Bhatti, T.T.; Eash, N.S. Climate Impacts on the agricultural sector of Pakistan: Risks and solutions. Environ. Chall. 2022, 6, 100433. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Lisek, A.; Sumorok, B.; Derkowska, E.; Szymańska, M.; Sas-Paszt, L. Arbuscular Mycorrhizal Fungi as an Important Factor Enabling the Adaptation of Anthyllis vulneraria L. to Zn-Pb-Polluted Tailings. Plants 2023, 12, 2092. [Google Scholar] [CrossRef]
- Zhou, J.; Chai, X.; Zhang, L.; George, T.S.; Wang, F.; Feng, G. Different Arbuscular Mycorrhizal Fungi Cocolonizing on a Single Plant Root System Recruit Distinct Microbiomes. mSystems 2020, 5, e00929-20. [Google Scholar] [CrossRef]
- Huey, C.J.; Gopinath, S.C.B.; Uda, M.N.A.; Zulhaimi, H.I.; Jaafar, M.N.; Kasim, F.H.; Yaakub, A.R.W. Mycorrhiza: A natural resource assists plant growth under varied soil conditions. 3 Biotech 2020, 10, 204. [Google Scholar] [CrossRef]
- de Andrade, L.A.; Santos, C.H.B.; Frezarin, E.T.; Sales, L.R.; Rigobelo, E.C. Plant Growth-Promoting Rhizobacteria for Sustainable Agricultural Production. Microorganisms 2023, 11, 1088. [Google Scholar] [CrossRef] [PubMed]
- Chamkhi, I.; El Omari, N.; Balahbib, A.; El Menyiy, N.; Benali, T.; Cherki Ghoulam, C. Is the rhizosphere a source of applicable multi-beneficial microorganisms for plant enhancement? Saudi J. Biol. Sci. 2022, 29, 1246–1259. [Google Scholar] [CrossRef] [PubMed]
- Chiaranunt, P.; White, J.F. Plant Beneficial Bacteria and Their Potential Applications in Vertical Farming Systems. Plants 2023, 12, 400. [Google Scholar] [CrossRef]
- Alrajhei, K.; Saleh, I.; Abu-Dieyeh, M.H. Biodiversity of arbuscular mycorrhizal fungi in plant roots and rhizosphere soil from different arid land environment of Qatar. Plant Direct 2022, 6, e369. [Google Scholar] [CrossRef]
- Fall, A.F.; Nakabonge, G.; Ssekandi, J.; Founoune-Mboup, H.; Apori, S.O.; Ndiaye, A.; Badji, A.; Ngom, K. Roles of Arbuscular Mycorrhizal Fungi on Soil Fertility: Contribution in the Improvement of Physical, Chemical, and Biological Properties of the Soil. Front. Fungal Biol. 2022, 3, 723892. [Google Scholar] [CrossRef]
- Soussani, F.E.; Boutasknit, A.; Ben-Laouane, R.; Benkirane, R.; Baslam, M.; Meddich, A. Arbuscular Mycorrhizal Fungi and Compost-Based Biostimulants Enhance Fitness, Physiological Responses, Yield, and Quality Traits of Drought-Stressed Tomato Plants. Plants 2023, 12, 1856. [Google Scholar] [CrossRef] [PubMed]
- Ladha, J.K.; Peoples, M.B.; Reddy, P.M.; Biswas, J.C.; Bennett, A.; Jat, M.L.; Krupnik, T.J. Biological nitrogen fixation and prospects for ecological intensification in cereal-based cropping systems. Field Crops Res. 2022, 283, 108541. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, K.; Abdul Rahman, N.S.N. The Microbial Connection to Sustainable Agriculture. Plants 2023, 12, 2307. [Google Scholar] [CrossRef]
- Jawahar, S.; Mariel, C.I.A.; Oswaldo, V.L. Harnessing the Potential of Symbiotic Associations of Plants in Phosphate-Deficient Soil for Sustainable Agriculture. Plant Cell Physiol. 2023, 64, 850–857. [Google Scholar] [CrossRef]
- El-Saadony, M.T.; Saad, A.M.; Soliman, S.M.; Salem, H.M.; Ahmed, A.; Mahmood, M.; El-Tahan, A.M.; Ebrahim, A.A.M.; Abd El-Mageed, T.A.; Negm, S.H.; et al. Plant growth-promoting microorganisms as biocontrol agents of plant diseases: Mechanisms, challenges and future perspectives. Front. Plant Sci. 2022, 13, 923880. [Google Scholar] [CrossRef]
- Pu, Z.; Zhang, R.; Wan, H.; Li, Q.; Zhang, J.; Wang, X.X. Root morphological and physiological traits and arbuscular mycorrhizal fungi shape phosphorus-acquisition strategies of 12 vegetable species. Front. Plant Sci. 2023, 14, 1150832. [Google Scholar] [CrossRef]
- Hao, Z.; Xie, W.; Jiang, X.; Wu, Z.; Zhang, X.; Chen, B. Arbuscular Mycorrhizal Fungus Improves Rhizobium–Glycyrrhiza Seedling Symbiosis under Drought Stress. Agronomy 2019, 9, 572. [Google Scholar] [CrossRef]
- Mazoyon, C.; Firmin, S.; Bensaddek, L.; Pecourt, A.; Chabot, A.; Faucon, M.P.; Sarazin, V.; Dubois, F.; Duclercq, J. Optimizing Crop Production with Bacterial Inputs: Insights into Chemical Dialogue between Sphingomonas sediminicola and Pisum sativum. Microorganisms 2023, 11, 1847. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.M. Amanual for the Practical Study of Root-Nodule Bacteria. J. Basic Microbiol. 1970, 12, 440. [Google Scholar] [CrossRef]
- Kaefer, E. Meiotic and mitotic recombination in Aspergillus and its chromosomal aberrations. Adv. Genet. 1977, 19, 33–131. [Google Scholar] [CrossRef]
- Jackson, N.E.; Franklin, R.E.; Miller, R.H. Effects of vesicular-arbuscular mycorrhizae on growth and phosphorus content of three agronomic crops. Soil Sci. Soc. Am. J. 1972, 36, 64–67. [Google Scholar] [CrossRef]
- Brockwell, J. Accuracy of a plant-infection technique for counting populations of Rhizobium trifolii. Appl. Microbiol. 1963, 11, 377–383. [Google Scholar] [CrossRef]
- Sáez-Plaza, P.; Navas, M.J.; Wybraniec, S.; Michałowski, T.; Asuero, A.G. An Overview of the Kjeldahl Method of Nitrogen Determination. Part II. Sample Preparation, Working Scale, Instrumental Finish, and Quality Control. Crit. Rev. Anal. Chem. 2013, 43, 224–272. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Cruz-Silva, A.; Figueiredo, A.; Sebastiana, M. First Insights into the Effect of Mycorrhizae on the Expression of Pathogen Effectors during the Infection of Grapevine with Plasmopara viticola. Sustainability 2021, 13, 1226. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS® 9.4 Language Reference: Concepts, 6th ed.; SAS Institute Inc.: Cary, NC, USA, 2016. [Google Scholar]
- Guo, X.; Wang, P.; Wang, X.; Li, Y.; Ji, B. Specific Plant Mycorrhizal Responses Are Linked to Mycorrhizal Fungal Species Interactions. Front. Plant Sci. 2022, 13, 930069. [Google Scholar] [CrossRef]
- Merrild, M.P.; Ambus, P.; Rosendahl, S.; Jakobsen, I. Common arbuscular mycorrhizal networks amplify competition for phos-545 phorus between seedlings and established plants. New Phytol. 2013, 200, 229–240. [Google Scholar] [CrossRef]
- Xu, Z.; Pehlivan, N.; Ghorbani, A.; Wu, C. Effects of Azorhizobium caulinodans and Piriformospora indica Co-Inoculation on Growth and Fruit Quality of Tomato (Solanum lycopersicum L.) under Salt Stress. Horticulturae 2022, 8, 302. [Google Scholar] [CrossRef]
- Heydari, S.; Pirzad, A. Mycorrhizal fungi and Thiobacillus co-inoculation improve the physiological indices of Lallemantia iberica under salinity stress. Curr. Microbiol. 2020, 77, 2523–2534. [Google Scholar] [CrossRef] [PubMed]
- Sheteiwy, M.S.; Abd Elgawad, H.; Xiong, Y.C.; Macovei, A.; Brestic, M.; Skalicky, M.; Shaghaleh, H.; Alhaj Hamoud, Y.; El-Sawah, A.M. Inoculation with Bacillus amyloliquefaciens and mycorrhiza confers tolerance to drought stress and improve seed yield and quality of soybean plant. Physiol. Plant. 2021, 172, 2153–2169. [Google Scholar] [CrossRef] [PubMed]
- Kocira, A.; Lamorska, J.; Kornas, R.; Nowosad, N.; Tomaszewska, M.; Leszczyńska, D.; Kozłowicz, K.; Tabor, S. Changes in Biochemistry and Yield in Response to Biostimulants Applied in Bean (Phaseolus vulgaris L.). Agronomy 2020, 10, 189. [Google Scholar] [CrossRef]
- Sadak, M.S.H.; Abdelhamid, M.T.; Schmidhalter, U. Effect of foliar application of aminoacids on plant yield and some physiological parameters in bean plants irrigated with seawater. Acta Biol. Colomb. 2015, 20, 141–152. [Google Scholar]
- Shafeek, M.R.; Hafez, M.M.; Mahmoud, A.R.; Ali, A.R. Comparative effect on N-fixing bacterial with foliar application of amino acid mixed on growth and yield of pea plants (Pisum sativum L.). Middle East J. Appl. Sci. 2014, 4, 755–761. [Google Scholar]
- Zewail, R.M.Y. Effect of seaweed extract and amino acids on growth and productivity and some biocostituents of common bean (Phaseolus vulgaris L.) plants. J. Plant. Prod. Mansoura Univ. 2014, 5, 1441–1453. [Google Scholar] [CrossRef]
- Sangiorgio, D.; Spinelli, F.; Vandelle, E. The unseen effect of pesticides: The impact on phytobiota structure and functions. Front. Agron. 2022, 4, 936032. [Google Scholar] [CrossRef]
- Stępkowski, T.; Banasiewicz, J.; Granada, C.E.; Andrews, M.; Passaglia, L.M.P. Phylogeny and Phylogeography of Rhizobial Symbionts Nodulating Legumes of the Tribe Genisteae. Genes 2018, 9, 163. [Google Scholar] [CrossRef]
- Shrestha, S.; Gautam, T.P.; Raut, J.K.; Goto, B.T.; Chaudhary, S.; Mandal, T.N. Edaphic factors and elevation gradient influence arbuscular mycorrhizal colonization and spore density in the rhizosphere of Shorea robusta Gaertn. Acta Ecol. Sin. 2023, in press. [Google Scholar] [CrossRef]
- Di Martino, C.; Torino, V.; Minotti, P.; Pietrantonio, L.; Del Grosso, C.; Palmieri, D.; Palumbo, G.; Crawford, T.W.J.; Carfagna, S. Mycorrhized Wheat Plants and Nitrogen Assimilation in Coexistence and Antagonism with Spontaneous Colonization of Pathogenic and Saprophytic Fungi in a Soil of Low Fertility. Plants 2022, 11, 924. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, E.; Gamper, H.A.; Ciccolini, V.; Ercoli, L. Forage Rotations Conserve Diversity of Arbuscular Mycorrhizal Fungi and Soil Fertility. Front. Microbiol. 2020, 10, 2969. [Google Scholar] [CrossRef] [PubMed]
- Rosendahl, S.; McGee, P.; Morton, J.B. Lack of global population genetic differentiation in the arbuscular mycorrhizal fungus Glomus mosseae suggests a recent range expansion which may have coincided with the spread of agriculture. Mol. Ecol. 2009, 18, 4316–4329. [Google Scholar] [CrossRef] [PubMed]
- Chafai, W.; El Gabardi, S.; Douira, A.; Khalid, A. Diversity and mycorrhizal potential of arbuscular mycorrhizal fungi in two natural soils in the eastern region of Morocco. Asian J. Agric. Biol. 2022, 2, 202102101. [Google Scholar] [CrossRef]
- Wu, M.; Wei, Q.; Xu, L.; Li, H.; Oelmüller, R.; Zhang, W. Piriformospora indica enhances phosphorus absorption by stimulating acid phosphatase activities and organic acid accumulation in Brassica napus. Plant Soil 2018, 432, 333–344. [Google Scholar] [CrossRef]
- Das, J.; Ramesh, K.V.; Maithri, U.; Mutangana, D.; Suresh, C.K. Response of aerobic rice to Piriformospora indica. Indian J. Exp. Biol. 2014, 52, 237–251. [Google Scholar] [PubMed]
- Yang, L.; Zou, Y.N.; Tian, Z.H.; Wu, Q.S.; Kuča, K. Effects of beneficial endophytic fungal inoculants on plant growth and nutrient absorption of trifoliate orange seedlings. Sci. Hortic. 2021, 277, 109815. [Google Scholar] [CrossRef]
- Ganugi, P.; Masoni, A.; Pietramellara, G.; Benedettelli, S. A Review of Studies from the Last Twenty Years on Plant–Arbuscular Mycorrhizal Fungi Associations and Their Uses for Wheat Crops. Agronomy 2019, 9, 840. [Google Scholar] [CrossRef]
- El-Sawah, A.M.; El-Keblawy, A.; Ali, D.F.I.; Ibrahim, H.M.; El-Sheikh, M.A.; Sharma, A.; Alhaj Hamoud, Y.; Shaghaleh, H.; Brestic, M.; Skalicky, M.; et al. Arbuscular Mycorrhizal Fungi and Plant Growth-Promoting Rhizobacteria Enhance Soil Key Enzymes, Plant Growth, Seed Yield, and Qualitative Attributes of Guar. Agriculture 2021, 11, 194. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.M.; Elkelish, A. Beneficial Features of Biochar and Arbuscular Mycorrhiza for Improving Spinach Plant Growth, Root Morphological Traits, Physiological Properties, and Soil Enzymatic Activities. J. Fungi 2021, 7, 571. [Google Scholar] [CrossRef]
- Beltayef, H.; Melki, M.; Saidi, W.; Hajri, R.; Cruz, C.; Muscolo, A.; ben Youness, M. Potential Piriformospora indica effect on growth and mineral nutrition of Phaseolus vulgaris crop under low phosphorus intake. J. Plant. Nutr. 2021, 44, 498–507. [Google Scholar] [CrossRef]
- Seshagiri, S.; Tallapragada, P. Study of Acid Phosphatase in Solubilization of Inorganic Phosphates by Piriformospora indica. Pol. J. Microbiol. 2017, 65, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Kalpana, N.; Lourdes, B.; Manoj, K.A.; Xochitl, A.A.; Federico, S.; Miguel, L. Nitrate regulates rhizobial and mycorrhizal symbiosis in common bean (Phaseolus vulgaris L.). J Integr Plant Biol. 2014, 56, 281–298. [Google Scholar] [CrossRef]
- Beltayef, H.; Melki, M.; Saidi, W.; Samaali, S.; Muscolo, A.; Cruz, C.; Garoui, T. Betterment of biological nitrogen fixation in snap bean under Mediterranean semi-arid conditions. Bulg. J. Agric. Sci. 2018, 24, 244–251. [Google Scholar]
- Sherameti, I.; Shahollari, B.; Venus, Y.; Altschmied, L.; Varma, A.; Oelmüller, R. The endophytic fungus Piriformospora indica stimulates the expression of nitrate reductase and the starch-degrading enzyme glucan-water dikinase in tobacco and Arabidopsis roots through a homeodomain transcription factor that binds to a conserved motif in their promoters. J. Biol. Chem. 2005, 280, 26241–26247. [Google Scholar] [CrossRef]
- Liu, C.-W.; Murray, J.D. The Role of Flavonoids in Nodulation Host-Range Specificity: An Update. Plants 2016, 5, 33. [Google Scholar] [CrossRef]
- Acosta-Jurado, S.; Fuentes-Romero, F.; Ruiz-Sainz, J.E.; Janczarek, M.; Vinardell, J.M. Rhizobial Exopolysaccharides: Genetic Regulation of Their Synthesis and Relevance in Symbiosis with Legumes. Int. J. Mol. Sci. 2021, 22, 6233. [Google Scholar] [CrossRef]
- Janczarek, M.; Rachwał, K.; Marzec, A.; Grzadziel, J.; Palusińska-Szysz, M. Signal molecules and cell-surface components involved in early stages of the legume-Rhizobium interactions. Appl. Soil Ecol. 2015, 85, 94–113. [Google Scholar] [CrossRef]
- López-Baena, F.J.; Ruiz-Sainz, J.E.; Rodríguez-Carvajal, M.A.; Vinardell, J.M. Bacterial molecular signals in the Sinorhizobium fredii-soybean symbiosis. Int. J. Mol. Sci. 2016, 17, 755. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Unit | Value |
|---|---|---|
| Sand | % | 29.5 |
| Clay | 47.0 | |
| Silt | 24.5 | |
| Texture | -- | Clay |
| pH | -- | 8.3 |
| Organic matter | % | 1.73 |
| Total N | 0.04 | |
| P | mg kg−1 | 180.41 |
| K | 241.80 |
| Parameter Variables | DF | F Value | (Pr > F) | |
|---|---|---|---|---|
| SDB (at flowering) | Treat | 4 | 28.46 | <0.0001 |
| Var | 1 | 32.13 | 0.02 | |
| Treat*Var | 4 | 4.91 | 0.008 | |
| SDB (at harvest) | Treat | 4 | 32.48 | <0.0001 |
| Var | 1 | 0.50 | ns | |
| Treat*Var | 4 | 2.84 | ns | |
| RDB (at flowering) | Treat | 4 | 27.14 | <0.0001 |
| Var | 1 | 6.20 | ns | |
| Treat*Var | 4 | 3.77 | 0.02 | |
| RDB (at harvest) | Treat | 4 | 102.12 | <0.0001 |
| Var | 1 | 1.01 | ns | |
| Treat*Var | 4 | 4.34 | 0.01 | |
| NOD numbers (at flowering) | Treat | 4 | 55.04 | <0.0001 |
| Var | 1 | 4.00 | ns | |
| Treat*Var | 4 | 3.16 | 0.04 | |
| NOD numbers (at harvest) | Treat | 4 | 7.68 | 0.001 |
| Var | 1 | 0.28 | ns | |
| Treat*Var | 4 | 3.30 | 0.03 | |
| POD numbers | Treat | 4 | 177.06 | <0.0001 |
| Var | 1 | 121.00 | 0.008 | |
| Treat*Var | 4 | 7.02 | 0.001 | |
| Treatments | Root Dry Biomass at Flowering (g) | Root Dry Biomass at Harvest (g) | Nodule Number/Plant at Flowering | Nodule Number/Plant at Harvest | POD Number/Plant | |
|---|---|---|---|---|---|---|
| Contender | Control | 1.37 ± 0.47 d | 1.11 ± 0.15 d | 97 ± 5.21 e | 86 ± 0.47 b | 5 ± 0.002 d |
| C | 2.39 ± 0.17 c | 2.80 ± 0.06 b | 251 ± 4.5 c | 92 ± 5.13 a | 9 ± 0.007 b | |
| C+Ri | 2.76 ± 0.09 b | 2.97 ± 0.08 a | 273 ± 2.84 b | 91 ± 6.6 a | 11 ± 0.009 a | |
| C+Si | 3.05 ± 0.03 a | 2.98 ± 0.18 a | 295 ± 4.63 a | 87 ± 1.05 b | 12 ± 0.01 a | |
| C+Ri+Si | 2.37 ± 0.13c | 2.11 ± 0.17 c | 185 ± 5.29 d | 83 ± 6.8 c | 7 ± 0.006 c | |
| Garrafal Enana | Control | 1.19 ± 0.06 d | 1.21 ± 0.02 d | 100 ± 1.52 d | 81 ± 2.12 c | 4 ± 0.002 d |
| C | 2.47 ± 0.19 b | 2.43 ± 0.06 b | 253 ± 7.37 b | 93 ± 3 a | 11 ± 0.008 b | |
| C+Ri | 2.98 ± 0.12 a | 2.84 ± 0.01 a | 303 ± 3.18 a | 92 ± 7.09 a | 13 ± 0.01 a | |
| C+Si | 2.99 ± 0.01 a | 2.85 ± 0.02 a | 313 ± 2.96 a | 91 ± 1.41 ab | 13 ± 0.008 a | |
| C+Ri+Si | 2.33 ± 0.05 c | 2.15 ± 0.1 c | 199 ± 3.5 c | 90 ± 6.55 ab | 8 ± 0.003 c | |
| Parameters | Variables | DF | F Value | (Pr > F) |
|---|---|---|---|---|
| Leaves phosphorus content (flowering stage) | Treat | 4 | 21.70 | <0.0001 |
| Var | 1 | 0.01 | ns | |
| Treat*Var | 4 | 0.47 | ns | |
| Leaves phosphorus content (at harvest) | Treat | 4 | 40.55 | <0.0001 |
| Var | 1 | 0.22 | ns | |
| Treat*Var | 4 | 2.95 | ns | |
| Root phosphorus content (flowering stage) | Treat | 4 | 30.35 | <0.0001 |
| Var | 1 | 4.95 | ns | |
| Treat*Var | 4 | 2.92 | ns | |
| Root phosphorus content (at harvest) | Treat | 4 | 39.13 | <0.0001 |
| Var | 1 | 2.67 | ns | |
| Treat*Var | 4 | 3.77 | ns | |
| Leaves nitrogen content (flowering stage) | Treat | 4 | 9.08 | 0.0005 |
| Var | 1 | 0.04 | ns | |
| Treat*Var | 4 | 0.34 | ns | |
| Leaves nitrogen content (at harvest) | Treat | 4 | 22.86 | <0.0001 |
| Var | 1 | 17.64 | 0.05 | |
| Treat*Var | 4 | 1.28 | ns | |
| Root nitrogen content (flowering stage) | Treat | 4 | 18.93 | <0.0001 |
| Var | 1 | 37.64 | 0.02 | |
| Treat*Var | 4 | 5.11 | 0.007 | |
| Root nitrogen content (at harvest) | Treat | 4 | 10.88 | 0.0002 |
| Var | 1 | 7.29 | ns | |
| Treat*Var | 4 | 0.63 | ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beltayef, H.; Mechri, M.; Saidi, W.; Raza, T.; Hajri, R.; Othmani, A.; Bouajila, K.; Cruz, C.; Hashem, A.; Abd_Allah, E.F.; et al. Synergistic Interaction of Rhizobium tropici, Rhizophagus irregularis and Serendipita indica in Promoting Snap Bean Growth. Agronomy 2023, 13, 2619. https://doi.org/10.3390/agronomy13102619
Beltayef H, Mechri M, Saidi W, Raza T, Hajri R, Othmani A, Bouajila K, Cruz C, Hashem A, Abd_Allah EF, et al. Synergistic Interaction of Rhizobium tropici, Rhizophagus irregularis and Serendipita indica in Promoting Snap Bean Growth. Agronomy. 2023; 13(10):2619. https://doi.org/10.3390/agronomy13102619
Chicago/Turabian StyleBeltayef, Hayet, Mouna Mechri, Wafa Saidi, Taqi Raza, Rim Hajri, Afef Othmani, Khedija Bouajila, Cristina Cruz, Abeer Hashem, Elsayed Fathi Abd_Allah, and et al. 2023. "Synergistic Interaction of Rhizobium tropici, Rhizophagus irregularis and Serendipita indica in Promoting Snap Bean Growth" Agronomy 13, no. 10: 2619. https://doi.org/10.3390/agronomy13102619
APA StyleBeltayef, H., Mechri, M., Saidi, W., Raza, T., Hajri, R., Othmani, A., Bouajila, K., Cruz, C., Hashem, A., Abd_Allah, E. F., & Melki, M. (2023). Synergistic Interaction of Rhizobium tropici, Rhizophagus irregularis and Serendipita indica in Promoting Snap Bean Growth. Agronomy, 13(10), 2619. https://doi.org/10.3390/agronomy13102619