1. Introduction
The adoption of high-density planting systems in mango (Mangifera indica L.) production is becoming increasingly popular in Australia due to its potential for earlier economic returns, sustained high yields of high-quality fruit, and reduced labor costs. However, the substantial establishment costs pose challenges to growers, requiring careful financial planning and risk assessment based on an understanding of how these new production systems differ from conventional systems. In the long run, those stated benefits of high-density planting systems are expected to outweigh the increased investment, making them a promising adaptation for the future of mango production.
The amount of light captured by interception, and how it is distributed within a canopy drives photosynthetic energy for tree growth, flowering, fruit set, quality and colour development [
1,
2,
3]. Interception and distribution are influenced by genetics, planting density, tree architecture and agronomic practices [
4,
5,
6,
7]. By planting trees at higher density, earlier cropping and higher yields have been achieved in some orchard crops because those additional trees/ha intercept more light/ha in the establishment years, before canopy closure is achieved, and can lead to a faster return on investment [
8]. The amount of light intercepted/ha increases curvilinearly with increasing tree densities through increased canopy volume, a superior leaf area index (LAI) and a more even light distribution within the canopy [
5,
9,
10,
11]. A linear relationship exists between total light interception and biomass production and is a key driver of yield [
12,
13,
14], with studies in some crops, including apple (
Malus × domestica), showing that manipulation of tree architecture [
15,
16], with tree training and the removal of branches, increased yield compared with control trees. The relationship tends to break down at relatively high light interception values: in tree crops, maximum yield is often found at light interception between 60–80% of sunlight [
17,
18].
Mango is an evergreen tree crop grown commercially in over 100 countries in tropical and subtropical climatic regions [
19]. It is the fifth most popular fruit worldwide [
17]. Mango crops are terminal bearers and have been cultivated for at least 1500 years [
18]. In Australia and in many other countries, mango is a relatively underdeveloped fruit tree crop and the orchard systems have undergone only slight changes over the past 50 years, integrating different varieties and mechanical pruning [
19]. Most commercial orchards are planted conventionally at low densities (100–200 trees/ha) in anticipation of trees growing vigorously, producing large specimens at maturity. Kensington Pride (KP) is the most popular mango variety grown in Australia, grown throughout Australia’s subtropical and tropical regions. Because of KP’s vigour, trees are traditionally planted at relatively wide spacing 10 × 10 m or 12 × 12 m (giving 100 and 70 trees/ha, respectively). Over time, these trees fill their allocated space and reach heights of 6 m or more. In recent years, mango plant spacings have reduced to an average plant spacing of 6 × 9 m (185 trees/ha). Previous attempts to increase planting density in KP failed due to overcrowding and tree vigour requiring excessive pruning that caused yield reduction [
20]. Growers using these closer spacings have committed to annual pruning to maintain a smaller canopy size. Generally, canopies are grown to touch in the row to maximise light interception and heights are kept to ~5 m for ease of spraying and picking. However, there are few reports available on the relationship between productivity and light interception in mango orchards.
The estimation of productivity and light interception in mango orchards over time is difficult as some cultivars have biennial and irregular bearing patterns and there can be strong rootstock effects [
21]. However, KP grown in northern Australia had nearly linear increases in fruit production from the first year of production through to 10 years [
22]. Singh et al. [
23] reported that productivity of conventionally planted mango orchards started to decline when trees reached 23–26 years old and had poor total orchard light interception.
Tree training methods, such as espalier systems, can be employed to manipulate leaf distribution and light penetration within a canopy [
24]. These techniques can be tailored to manage the canopy’s architecture and light interception, driving more efficient use of the available light resources. This manipulation of tree shoot architecture changes light interception and distribution within canopy change in an attempt to optimize radiation use efficiency [
24].
The mango industry’s transition from conventional, large-tree orchards to smaller tree orchards has potential to improve profits for mango growers. However, there is little understanding of how the total light interception of mango orchards and light distribution are related to mango training systems and productivity. The objectives of this research were to document the relationships between light interception, canopy size, and yield in young mango trees using higher density planting systems in combination with three different varieties. Additionally, we conducted a study in conventional orchards to compare light relations with alternative planting systems. Our study aims to provide valuable information for farmers, agronomists, and scientists, which will be useful for optimizing canopy management and light interception to maximize fruit productivity in the industry.
2. Materials and Methods
2.1. Light Interception in Low-Density, Conventionally Trained Trees in Commercial Mango Orchards
We conducted a survey to establish baseline light relationships in commercial KP mango orchards using conventional planting system in the Mareeba/Dimbula district of Queensland with soils varying from granitic sands to volcanic light clays. All orchards in this study were irrigated using channel water from lake Tinaroo. We measured total light interception, canopy volume and tree yield using nine orchard blocks of different ages, (2, 4, 8, 9, 10, 13, 26, 28 and 31 years old) producing data in developing orchards with varying canopy size in the low-density plantings. Each block was planted at conventional low-density, with 185, 185, 185, 156, 185, 173, 128, 139 and 196 trees/ha respectively, and conventionally trained. Within each orchard block, five trees were selected to represent an individual experimental unit, with the central three used as datum trees. In July 2015, the transmission of total photosynthetic active radiation (PAR) under the canopy of each tree was measured using an AccuPAR linear ceptometer (model LP-80, Decagon Devices Inc., Pullman, WA, USA) for calculation of the proportion of light intercepted by the canopy. Canopy dimensions (tree height, width, depth, and skirt) were measured to calculate tree volume and yield/m3 canopy volume. Tree yield was estimated for all datum trees in each orchard prior to harvest by counting the number of fruit/tree and multiplying by the average fruit weight of a sub-sample of 10 fruit/tree in 2015. Yield (kg/ha) was calculated using the average of the three tree yields multiplied by trees/ha.
2.2. Light Relationships in High-Density, Intensive Orchards with Alternative Tree Training Systems
We studied light relationships in intensive mango planting and training orchard systems to investigate how light relations were affected by planting density and canopy training. The planting systems experiment was designed as a split-split plot with density as the main plot, training system as sub-plot and variety as lower sub-sub-plot. The experimental site was located on the Queensland Department of Agriculture and Fisheries Research Facility in the Walkamin district of Queensland, Australia (Latitude: 17.13 °S Longitude: 145.43 °E, Elevation: 594 m). The site has a dry topical climate with a mean annual maximum 24.7 °C and minimum 17.1 °C temperature, 1026 mm mean annual rainfall and a mean of 7.8 h of sunshine per day [
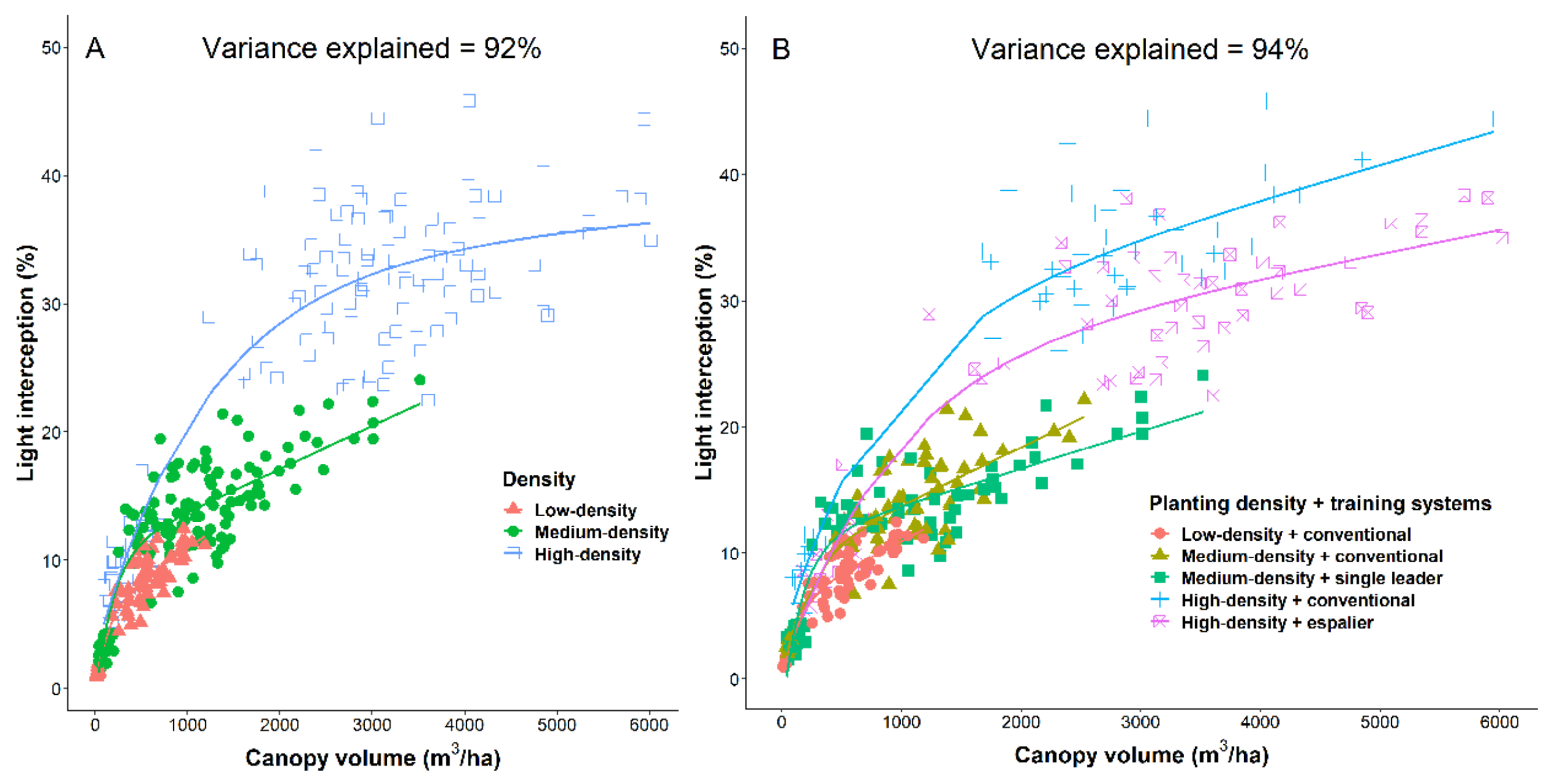
25]. Trees were planted in December 2014 with KP rootstock. Three varieties, National Mango Breeding Program NMBP1243, Keitt and Calypso were planted at 3 densities. Low-density 8 m × 6 m = 208 trees/ha, medium-density 6 m × 4 m = 417 trees/ha, high-density 4 m × 2 m = 1250 trees/ha. Within the medium-density treatment, there were two training systems: conventional and single leader. Within the high-density treatment, there were two training systems: espalier and conventional. Each planting system was replicated six times. Each sub-sub-plot comprised 3 datum trees, except the high-density espalier sub-sub-plots comprised 4 datum trees. Rows within the experiment run north to south. In the low-density treatment, trees were grown naturally with no training. In the medium-density treatment, trees were trained as either conventional or as single leader. In the high-density treatments, trees were trained as either conventional or as espalier on trellis. The main scaffold limbs in the single leader and espalier training systems were trained (bent) horizontally every 60 cm. Trees in each planting system were pruned annually just after harvest.
2.3. Light Interception, Tree Volume and Yield Data Collection in Intensive Planting Systems
Photosynthetic photon flux (PPF) was measured using 15 Apogee SQ -100 PAR point sensors (Apogee Instruments, Logan, UT, USA) mounted atop a 427 cm long aluminium arm with spacing of 30 cm. The aluminium arm was mounted to a modified trolley and the sensors were connected to a CR1000X Series (Campbell Scientific, Garbutt, Australia) data logger. A PC200W (Data Logger Starter Software, Version 4.5, Campbell Scientific, Inc., Logan, UT, USA) data logger was used for data collection. Light measurements were recorded for all 288 datum trees for intensive mango planting systems treatments. Light interception was measured on cloud-free days, five times throughout the day: early morning, mid-morning, solar noon, mid-afternoon and late afternoon. The value was generated by averaging these readings. The time for each of these sampling events were calculated as 1.5 h and 3 h before and after solar noon on the day the measures were taken. With rows running due North/South, data collected at the corresponding time from the morning or afternoon was substituted if data was not able to be recorded at the equal but opposite time of the day. Above canopy light data was collected in each density at the end of every two rows under the condition of full sun. We calculated light interception for each sub-sub-plot as the proportion of incident above canopy PPF reaching the orchard floor for each measurement, using the average of all the measurements within the sub-sub-plot. The light interception and canopy volume/ha were measured in the planting systems experiment in 2015, 2016, 2017 and 2018 when the trees were 2, 3, 4 and 5 years old.
We measured tree canopy volume for each datum tree and the mean canopy volume/ha was calculated to give one corresponding value per sub-sub-plot. Canopy volume for conventional trees was calculated using length × width × canopy height. Canopy volume in the medium-density single leader trees was calculated as the volume of a cylinder (πr2h) (where r is the radius and h is the canopy height) multiplied by the percentage of canopy on each 50 cm interval. Canopy volume in the high-density espalier trees was calculated as the volume of a trapezoidal prism LH(A+B)/2 where: L is the depth of the canopy; H is the height of the total canopy minus the gaps in between the scaffolds of canopy; A+B is the length of the canopy along the row at the top of the canopy plus the length of canopy along the row of the bottom of the canopy.
We harvested the datum trees and weighed fruit between late December and early February in 2016, 2017 and 2018. No yield data was recorded in 2015 (2 year old trees). Every individual fruit in each datum tree was weighed in 2016 and 2018 whereas a random selection of 25 fruit/tree were weighed in 2017 to estimate average fruit weight/tree.
2.4. Statistical Methods
Analysis of variance (ANOVA) was used to analyse the mean light interception in both experiments for each individual measurement event. All significance testing was performed at the 0.05 level and where a significant effect was found, the pairwise 95% least significant difference (lsd) was used to make comparisons. Linear and non-linear models were used to investigate relationships between variables measured within the light interception experiments. A log10 transformation was applied when needed to satisfy the assumption of homogeneity of variance and normality underlying the ANOVA performed on data from the planting systems experiment.
4. Discussion
Mango is a high value fruit and growers want to maximise their returns by increasing the amount of light intercepted, and the rate at which that energy is converted to salable fruit, so are investigating planting higher-density planting systems. The authors have observed growers adopting annual pruning methods. Case studies wherein orchards planted under high-density or ultra-high-density but without adopting the mandatory techniques, resulting in poor yield and more pest and disease incidences are discussed [
26]. Therefore, consistent tree training or pruning is necessary. In addition, increased light interception results in increased dry matter accumulation/ha [
11], but not necessarily fruit if this energy is used for vegetative growth. These relationships highlight important trends with canopy volume, management and yield where there is competition between vegetative growth and fruiting. Studies on apple showed that the initial establishment costs of trellis and branch training and pruning of espalier and non-trellis single leader systems are high, with the risk of the extra pruning stimulating unwanted vegetative growth in the place of reproductive growth [
27]. This study provides some physiologically-based measurement of canopy response to training method and planting density, in the hope that sensible guidelines can be developed to assist growers to manage blocks planted at high-density profitably.
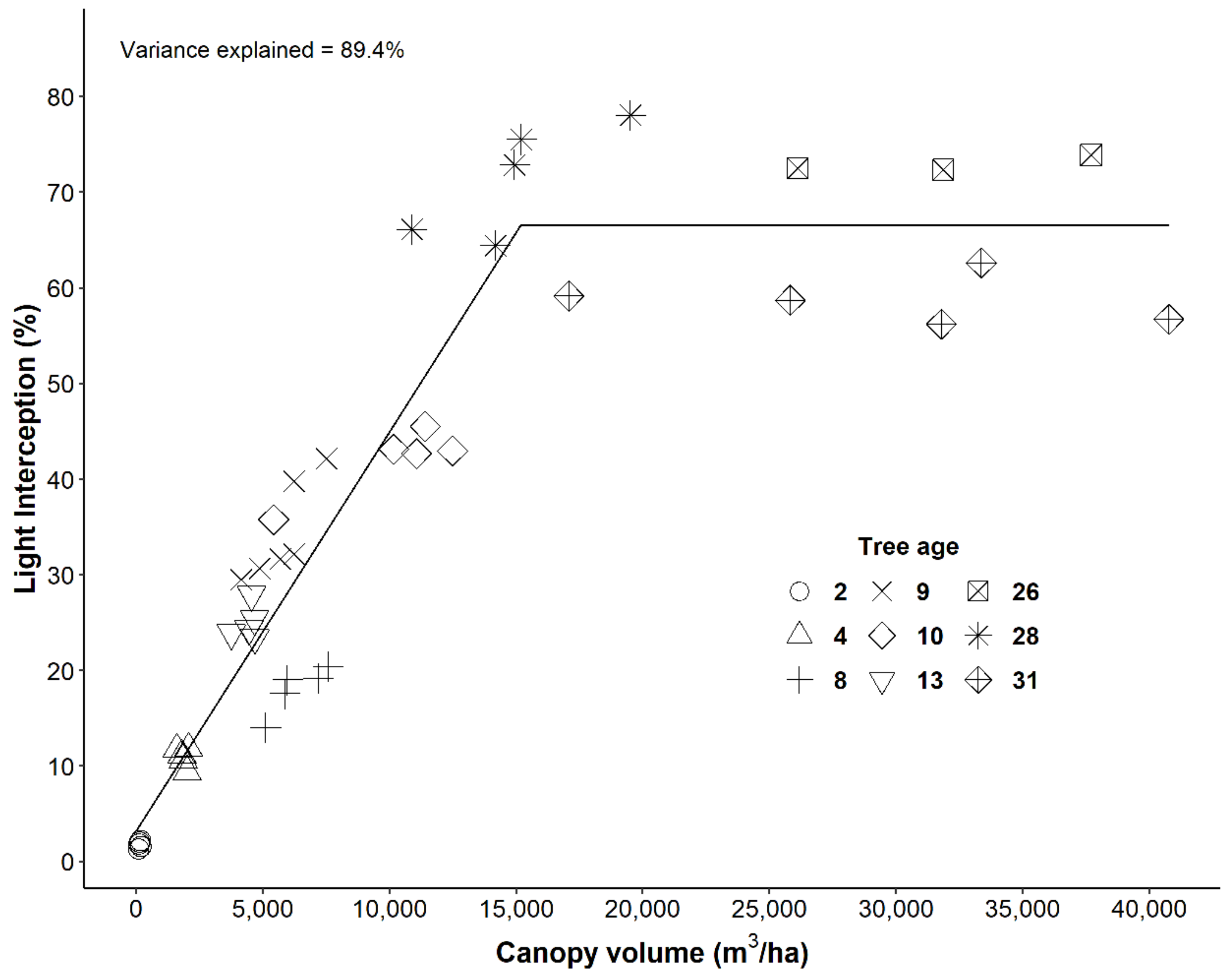
In our observations of traditional low-density mango orchards, the trees reached canopy volumes of up to 15,200 m
3/ha with light interception of 67% between their 13th and 26th year (
Figure 1). This maximum observed level of light interception is similar to those commonly measured in apple orchards (~60–70%) [
6]. Our yield observations in traditional mango orchards showed yields reached a mean maximum of 16,000 kg/ha when the canopy intercepted 55% of incident light. Yields declined at higher light interception (
Figure 2), similar to that seen in apple orchards, where yields declined after 60% light interception [
17,
27]. Beyond that 55% light threshold, those traditional low-density mango trees increased light interception up to 67%, but the energy demand by the fruit reduced and resources can be partitioned and shifted to vegetative growth [
28]. Mango trees may have very low photosynthetic rates per unit of leaf area, especially in the inner- and lower-canopy area because they may have very low light distribution within their canopies. It is worthwhile to do further study to understand this better.
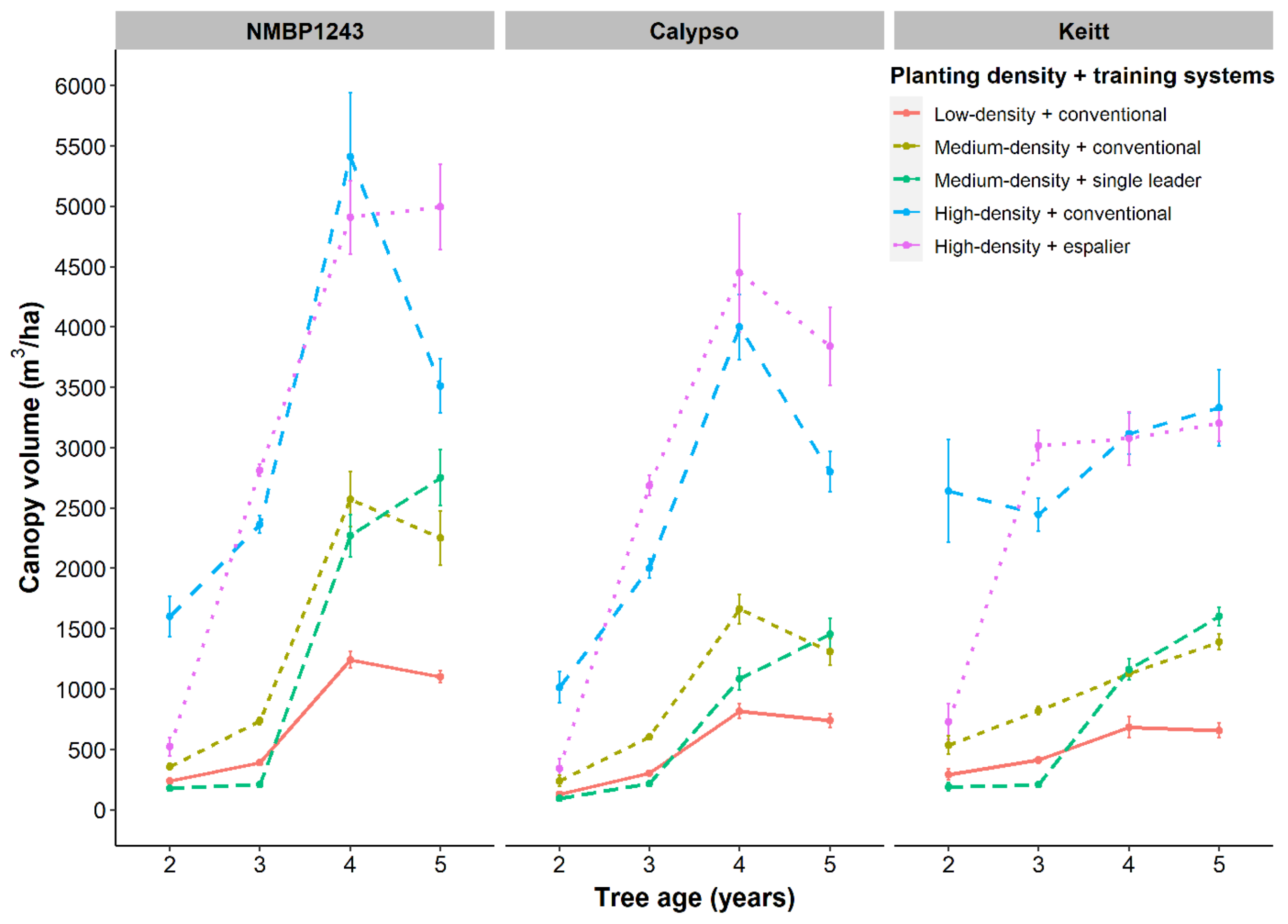
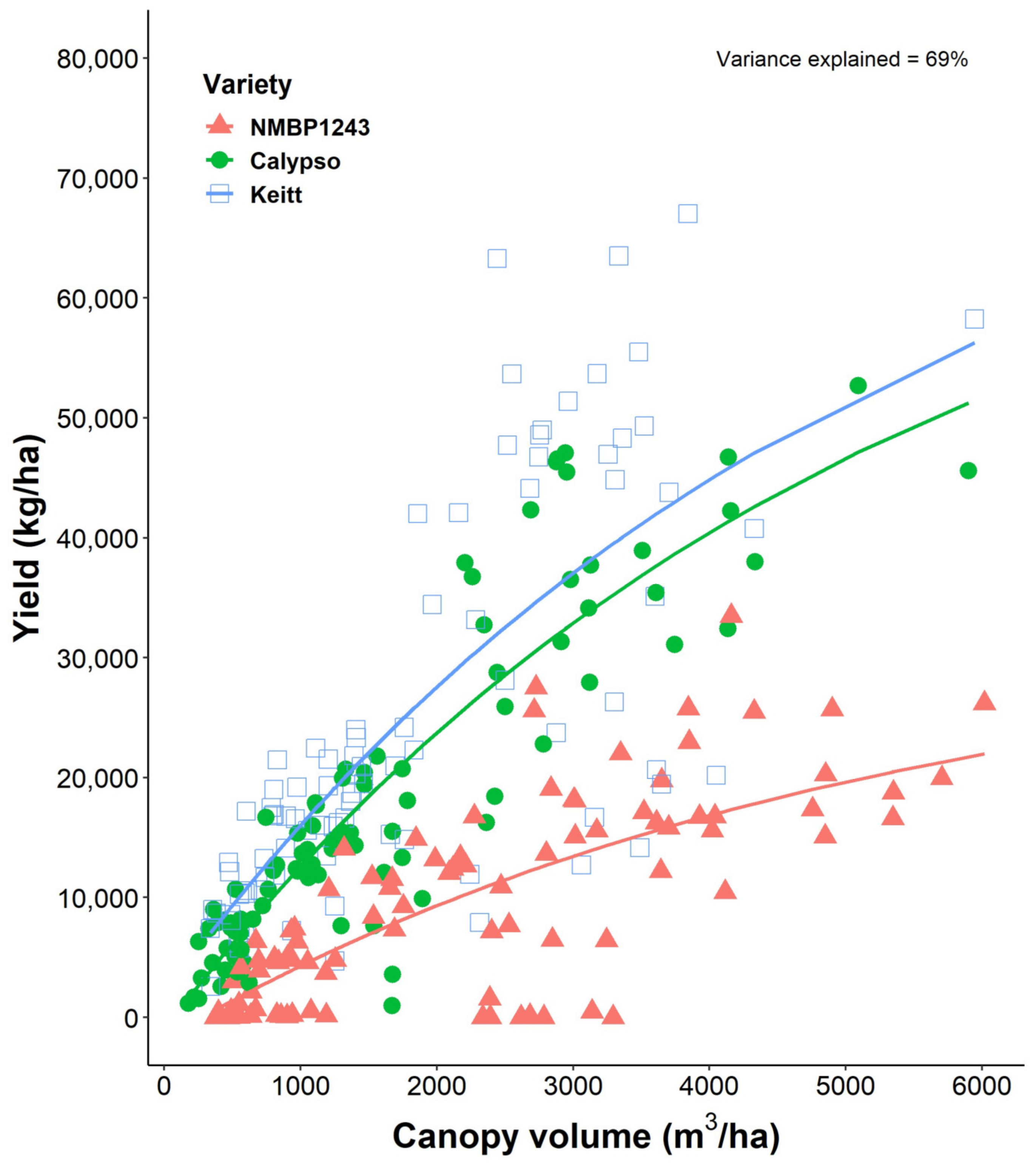
As 4-year-old trees, conventional planting Kensington Pride (180 trees/ha) had grown its canopy to ~1900 m
3/ha, or 10.55 m
3/tree (
Figure 2). At the same age, NMBP1243, Calypso and Keitt, planted at 204 trees/ha, had grown their canopies to 1243, 816 and 683 m
3/ha, respectively, or 6.1, 4.0 and 3.3 m
3/tree, respectively (
Figure 3). In the fifth and final year of our study, NMBP1243 mango trees, the fastest growing in our intensive planting systems experiment, planted at high-density had reached ~40% of the canopy volume achieved in mature, conventional KP orchards at canopy closure (6000 m
3/ha cf. 15,200 m
3/ha,
Figure 3 and
Figure 9). These high-density NMBP1243 trees, planted at 1250 trees/ha, correspond to a similar amount intercepted as that shown in 8-, 9- and 10-year-old KP orchards planted at low-density (180 trees/ha). On a per-tree basis, those same KP trees grew at a rate of 3.7 m
3/tree/year, compared with 0.96 m
3/tree/year for NMBP1243, demonstrating KP’s known status as a vigorous mango variety.
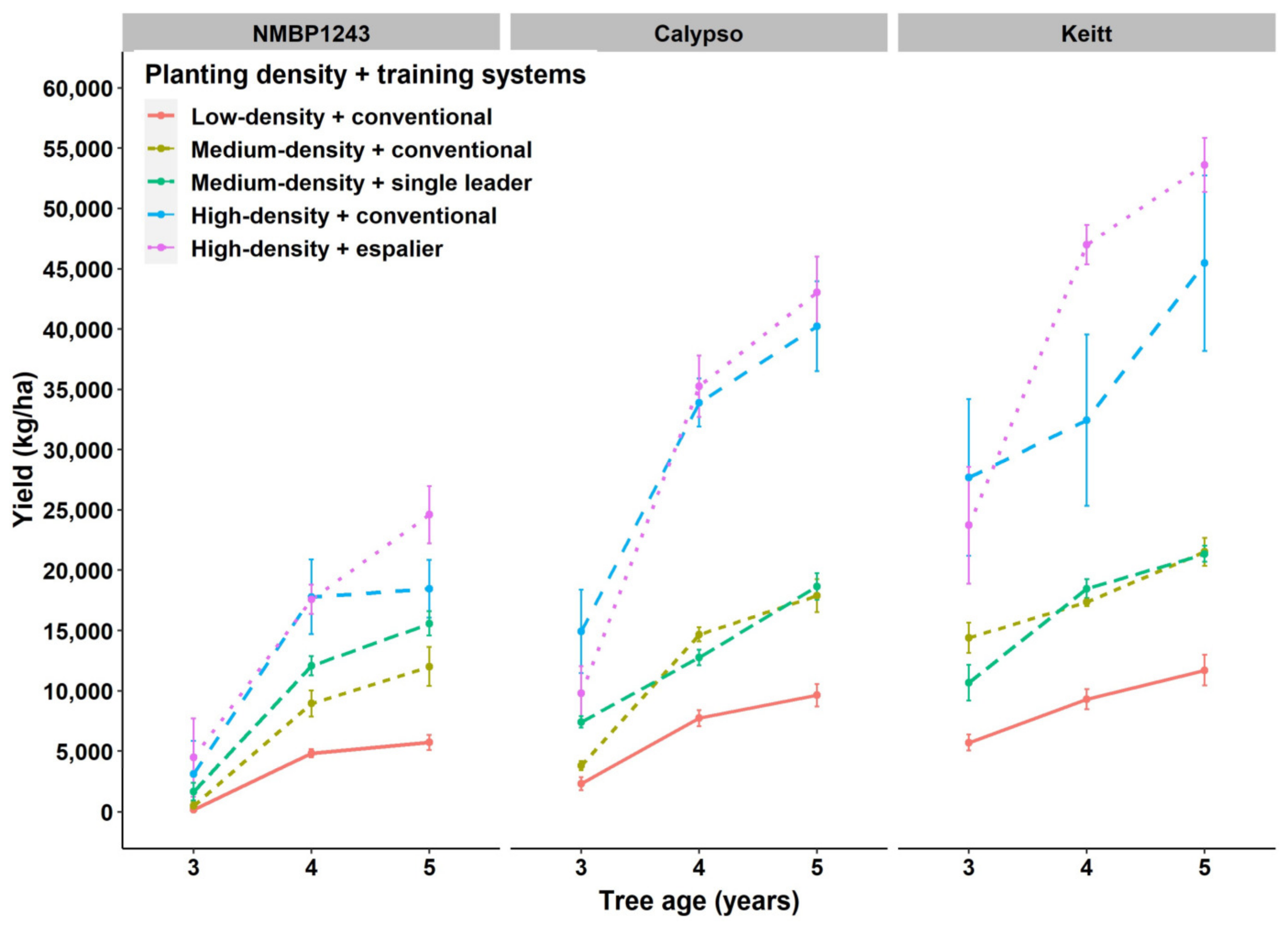
Our research has shown that higher density planting in mango increased the canopy volume/ha leading to an increase in light interception in the orchard compared to the conventional low- density conventional training systems in coming years, light interception will increase in the espalier and non-trellis single leader training systems once the tree structure is shaped and has reached their allotted space. The young trees in high density training systems reached ~55,000 kg/ha (
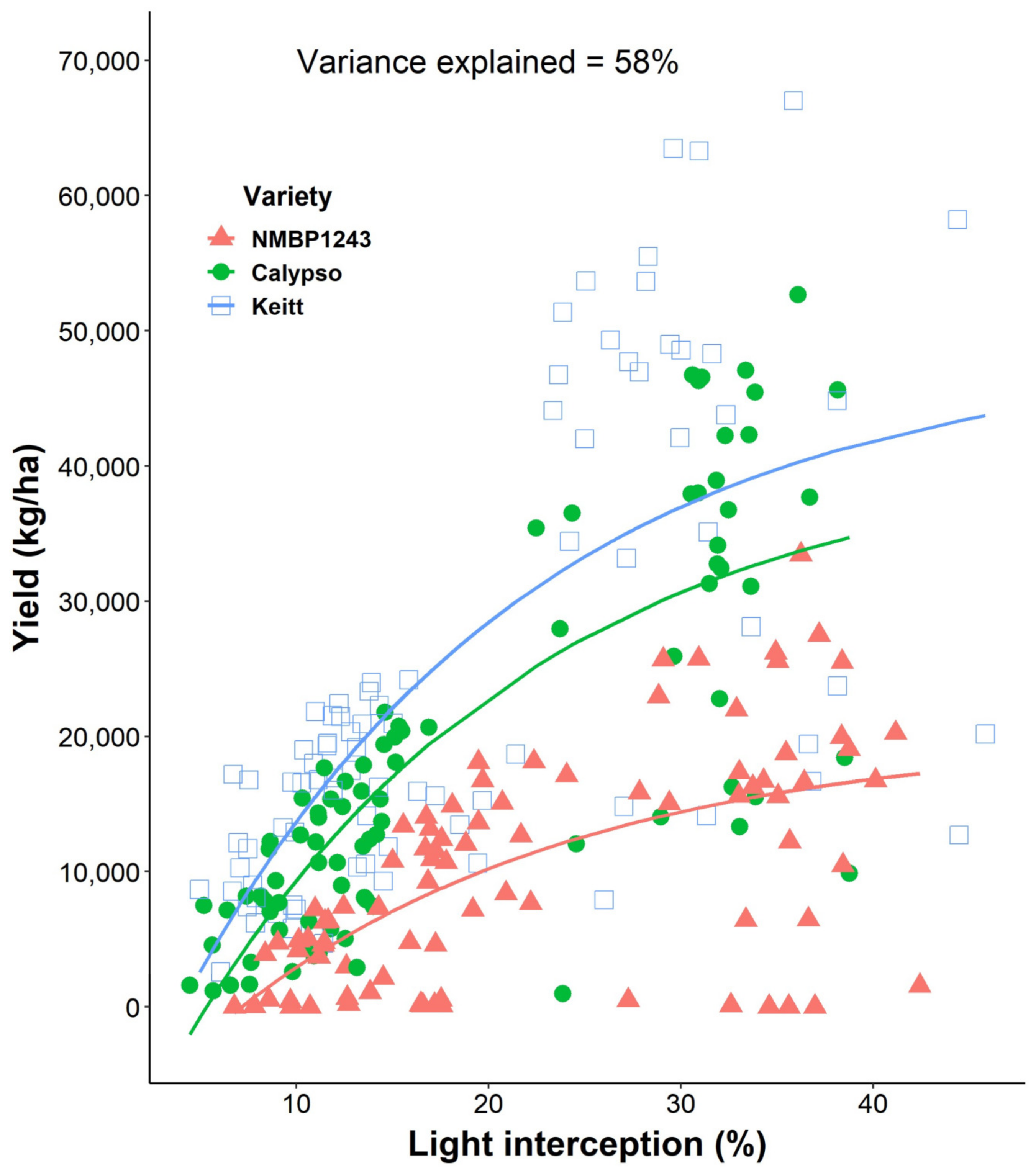
Figure 6). In this research, yield was the result of a combination of genotype and their biennial bearing characteristics. Keitt had the highest yield compared to Calypso and NMBP1243, despite a smaller canopy volume (
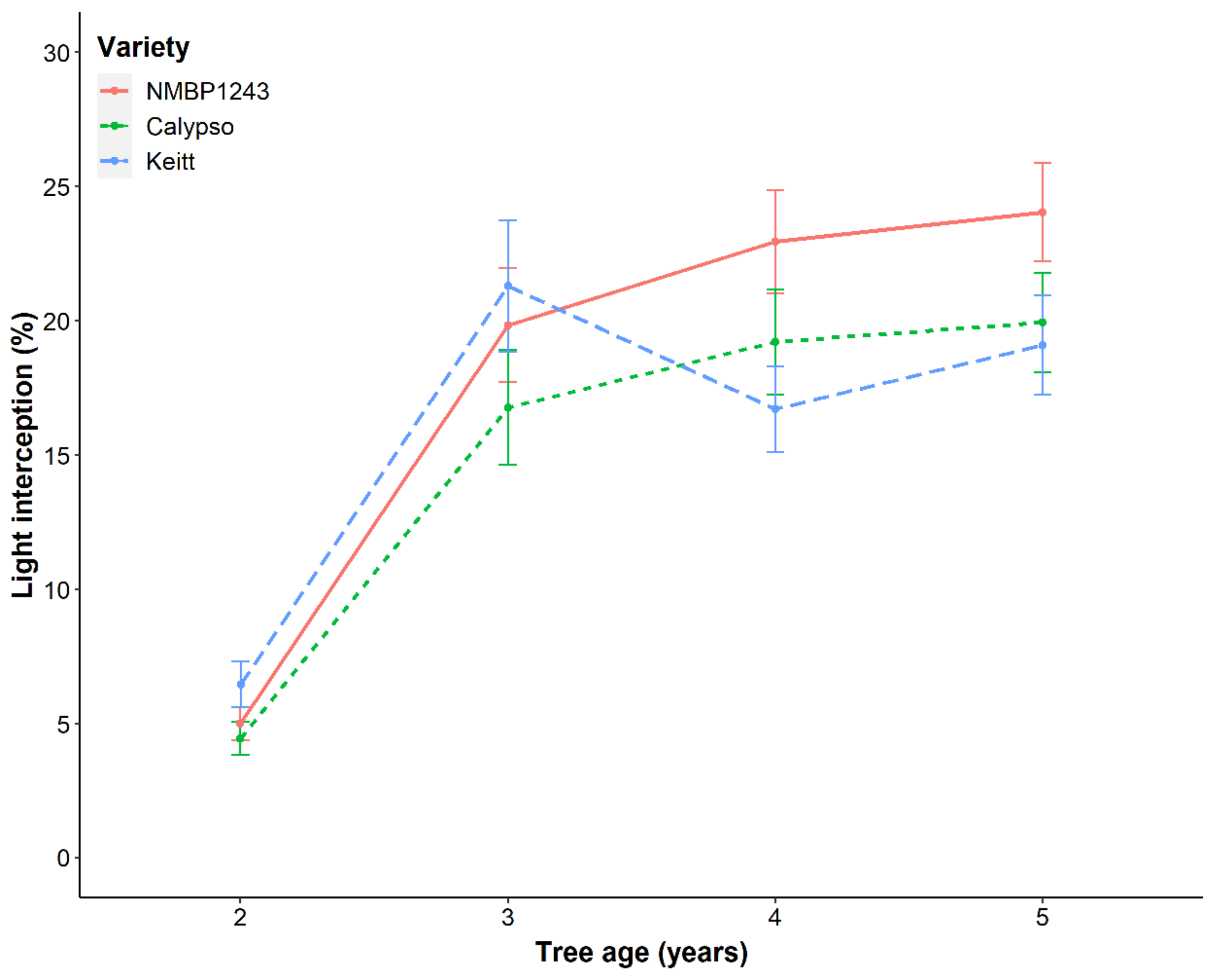
Figure 3) and lower light interception (
Table 1 and
Figure 5). The lower canopy volume in Keitt was not due to pruning or training. It is a result of the natural growth habit and tree architecture of Keitt. The decrease in canopy volume of Calypso and NMBP1243 in high density (
Figure 3) in year 5 was due to pruning and training of espalier and hedging of the conventional trees. In addition, no relationship between light interception and yield was found, showing the irregular nature of mature mango canopies to fruit [
29,
30]. The yield was greater in year 5 compared to year 4, which was opposite to the trend observed between canopy volume (
Figure 3) and light interception (
Figure 4). We observed a trend toward a further increase in light capture by the tree canopy using different canopy training systems. Training systems can change light distribution through the canopy and how it is intercepted across the orchard floor [
11,
24].
Therefore, we propose that high-density training systems lead to greater economic yield earlier in the life of the tree compared to conventional orchard systems [
31]. This is tempered by the significant difference between varieties. Jackson and Palmer [
5,
10] suggested that by implementing different training methods we can increase tree height, and with appropriate pruning and training we can increase light use efficiency, due to taller thinner canopies and lower leaf density which positively influences internal canopy irradiance. The optimisation of light interception and canopy irradiance are considered as prerequisites for effective production and high fruit quality [
32]. Our results show how light interception can be increased in young mango canopies by increasing planting density and by using alternative single leader training systems which leads to early yield gain per unit land area compared to low tree density. The espalier and single leader systems suggested that such training may increase trellis and training cost, but ease canopy management in the longer term [
31]. Mango growers can choose a suitable training system based on their personal preference.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}