
Transmissible Effects of a Biochar Amendment to an Upper Soil Zone Upon an Associated Lower Zone, Including Attenuation of Soybean Cyst Nematode (Heterodera glycines)
 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Biochar Preparation
2.2. Soil Preparation
2.3. Soil Columns
2.4. Mung Bean Cultivation
2.5. Leaching
2.6. Soil Sampling and Analysis
2.6.1. Chemical Properties
2.6.2. Microbial Activity and Enzyme Activities
2.6.3. Nematode Extraction
2.6.4. DNA Assay for Quantifying the Density of Soybean Cyst Nematode
2.7. Statistical Analysis
3. Results
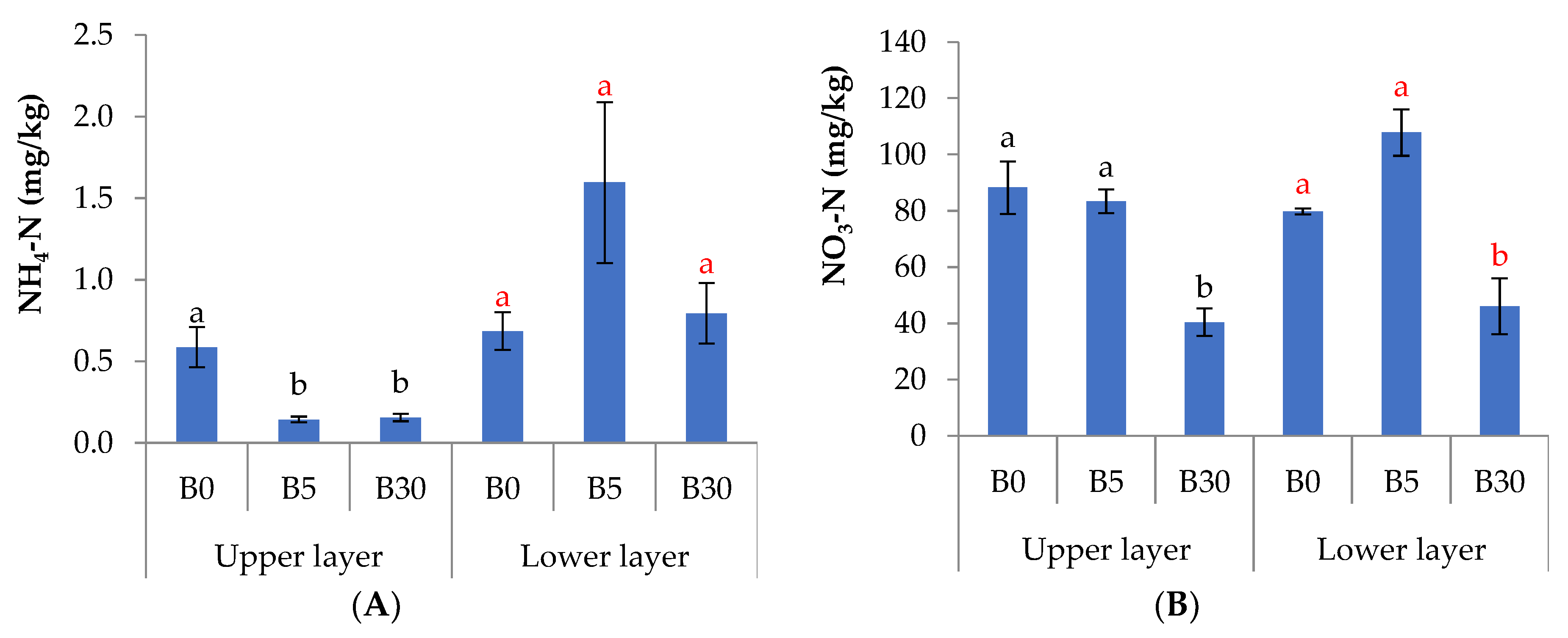
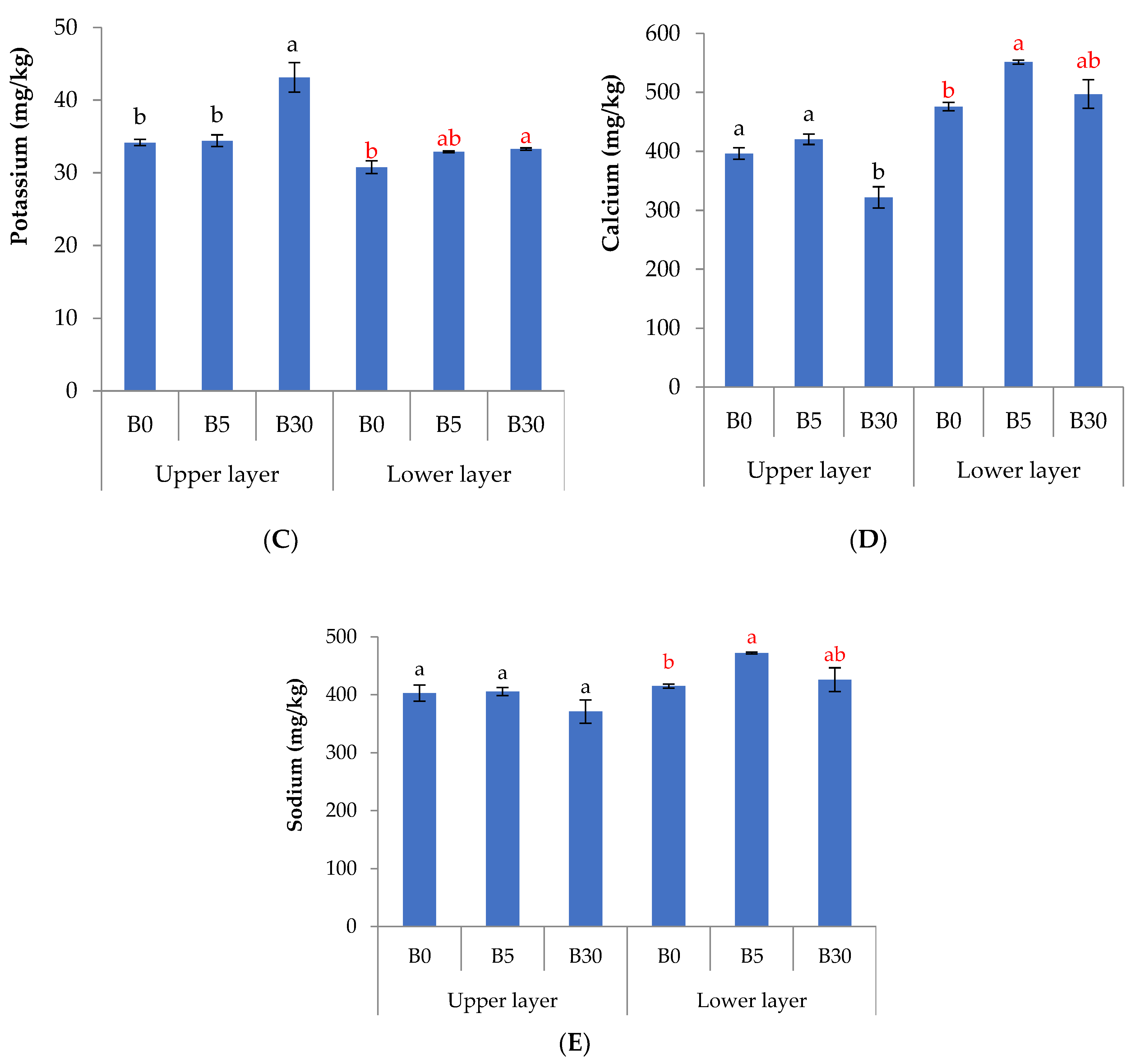
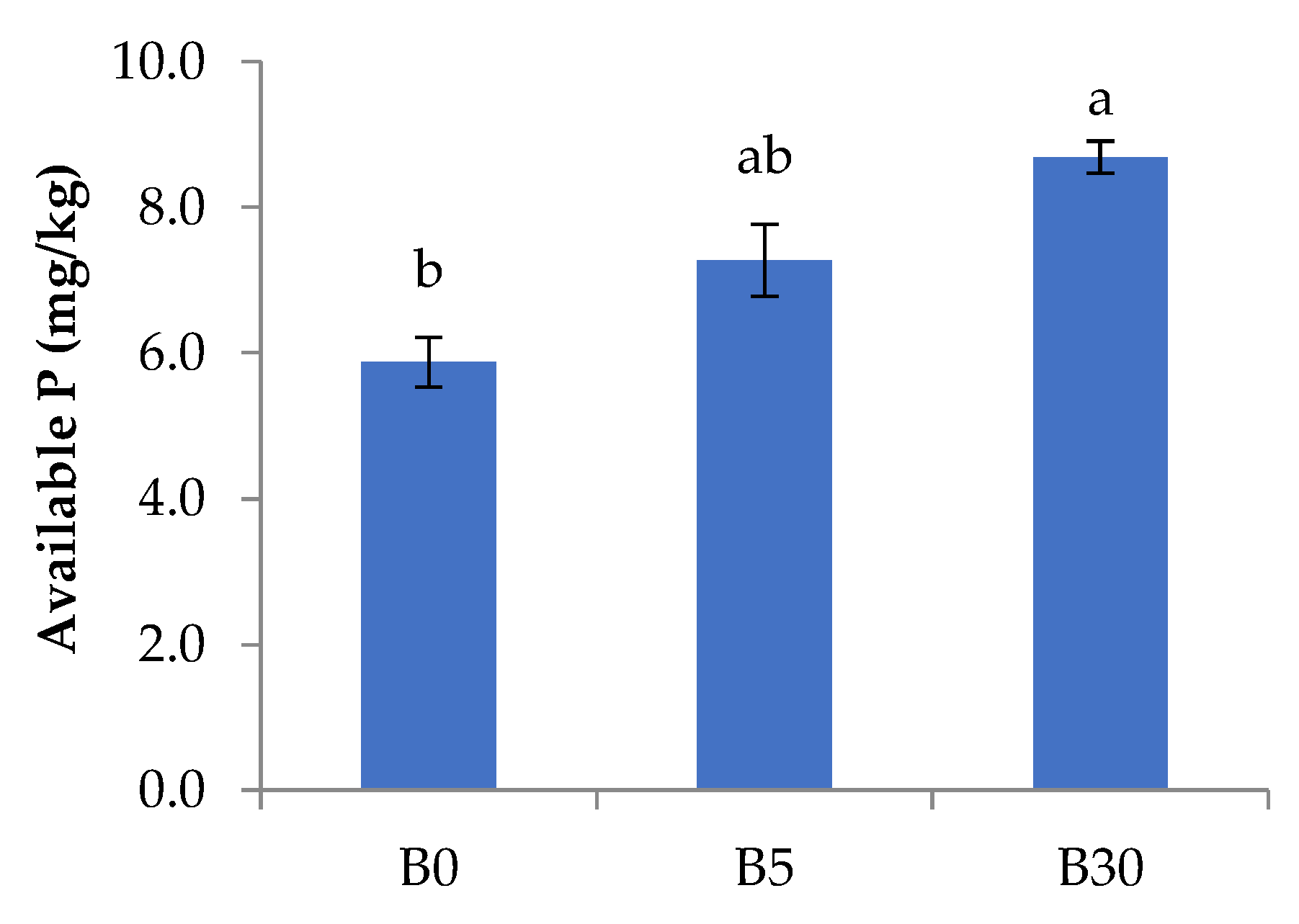
3.1. Effects of Biochar Amendment to Upper Layer on the Chemical Properties in Upper and Lower Layer Soils
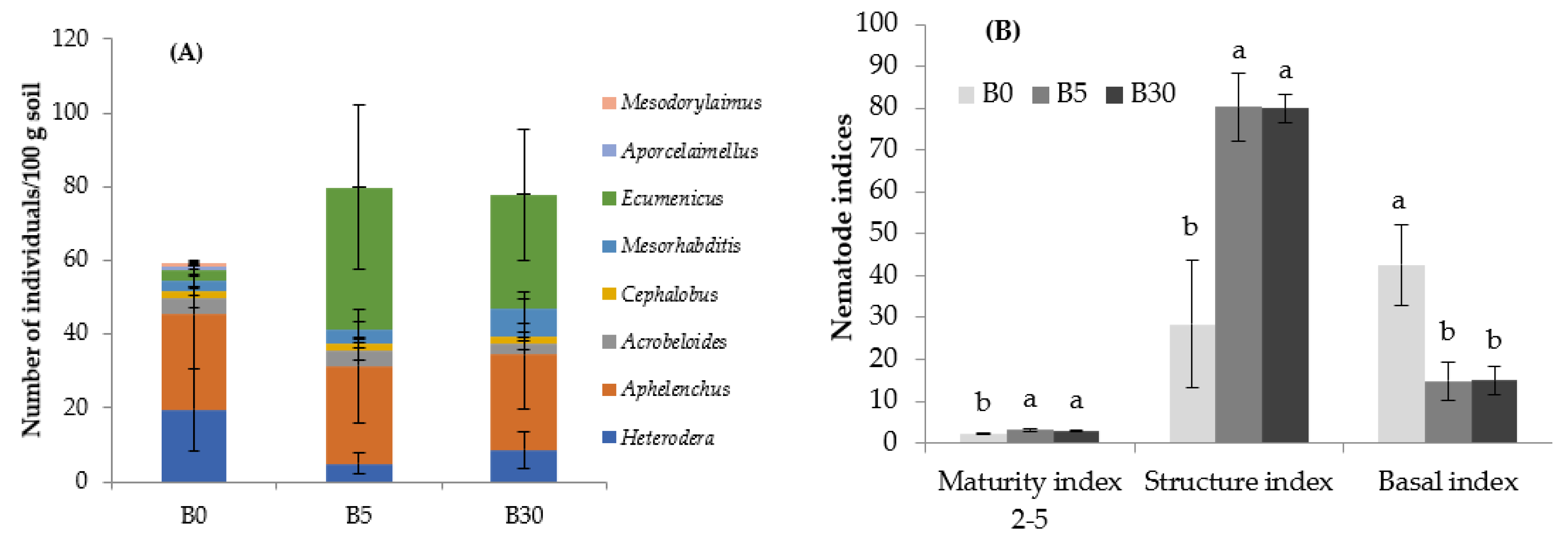
3.2. Effects of Biochar Amendment to Upper Layer on the Nematode Abundance and Composition in Upper Layer Soil
3.3. Effects of Biochar Amendment to Upper Layer on the Nematode Abundance and Composition in Lower Layer Soil
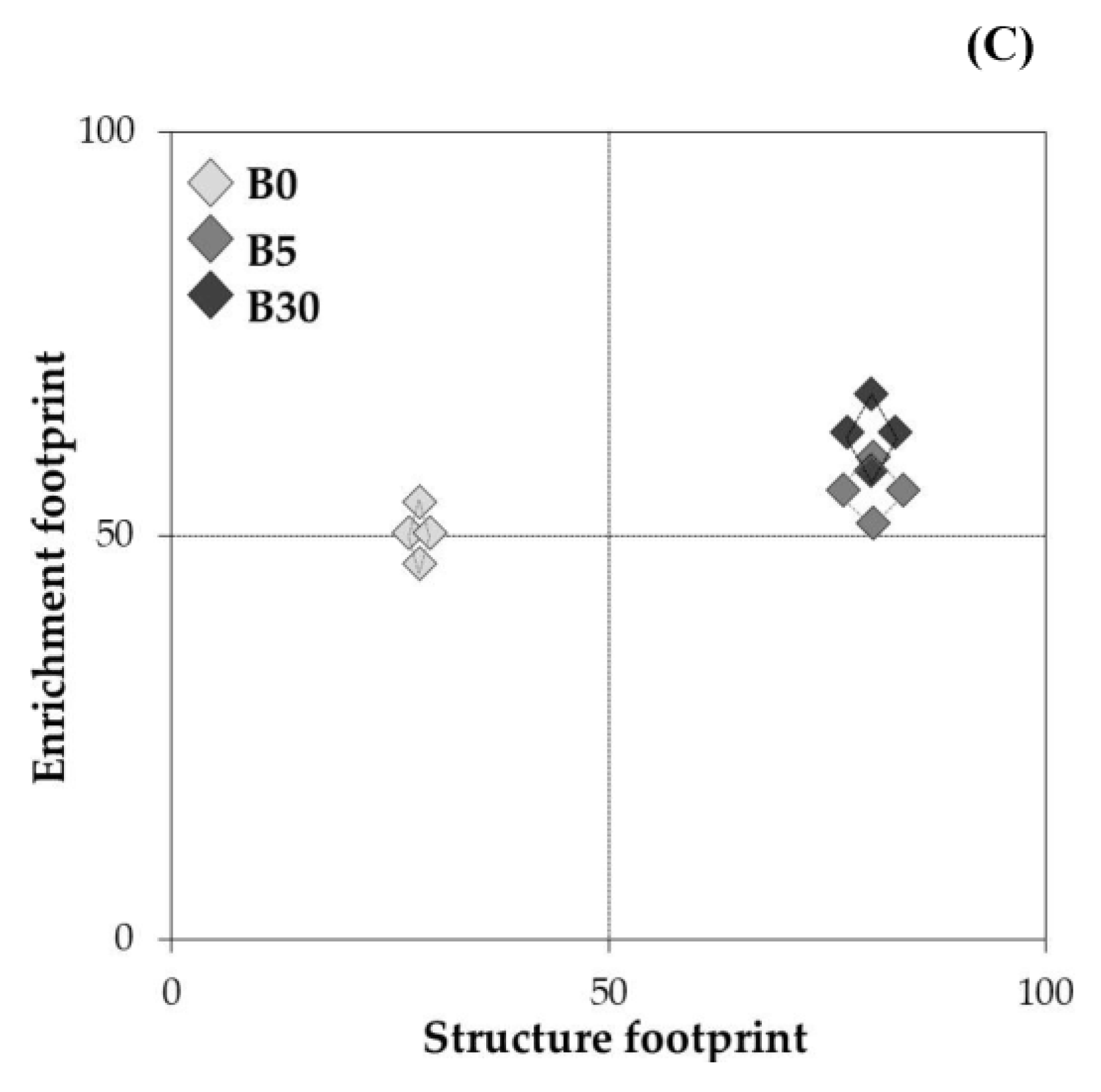
3.3.1. Nematode Community, Nematode Indices, and Metabolic Footprint
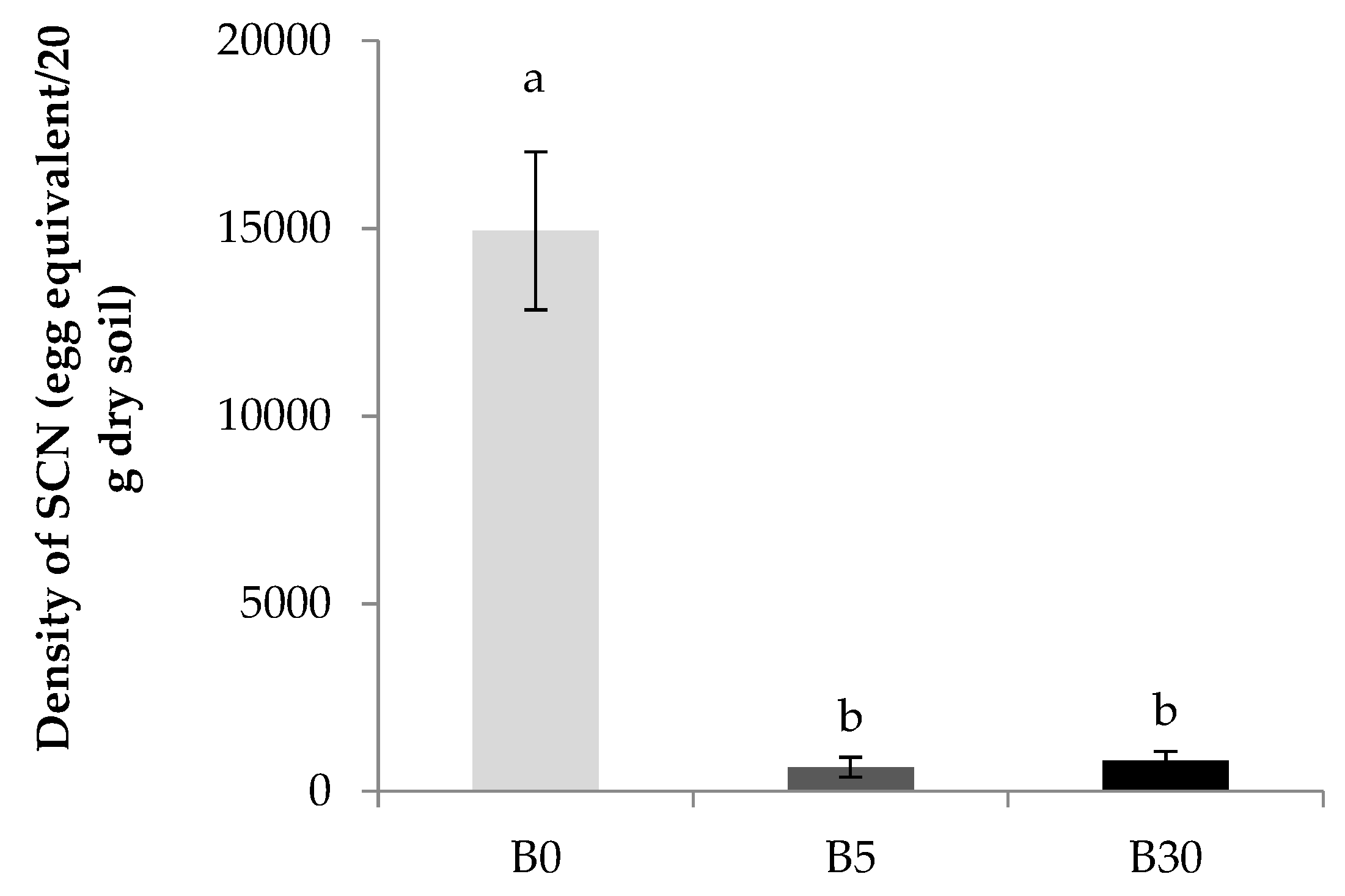
3.3.2. Density of Soybean Cyst Nematode
3.3.3. Microbial Activity and Enzyme Activity
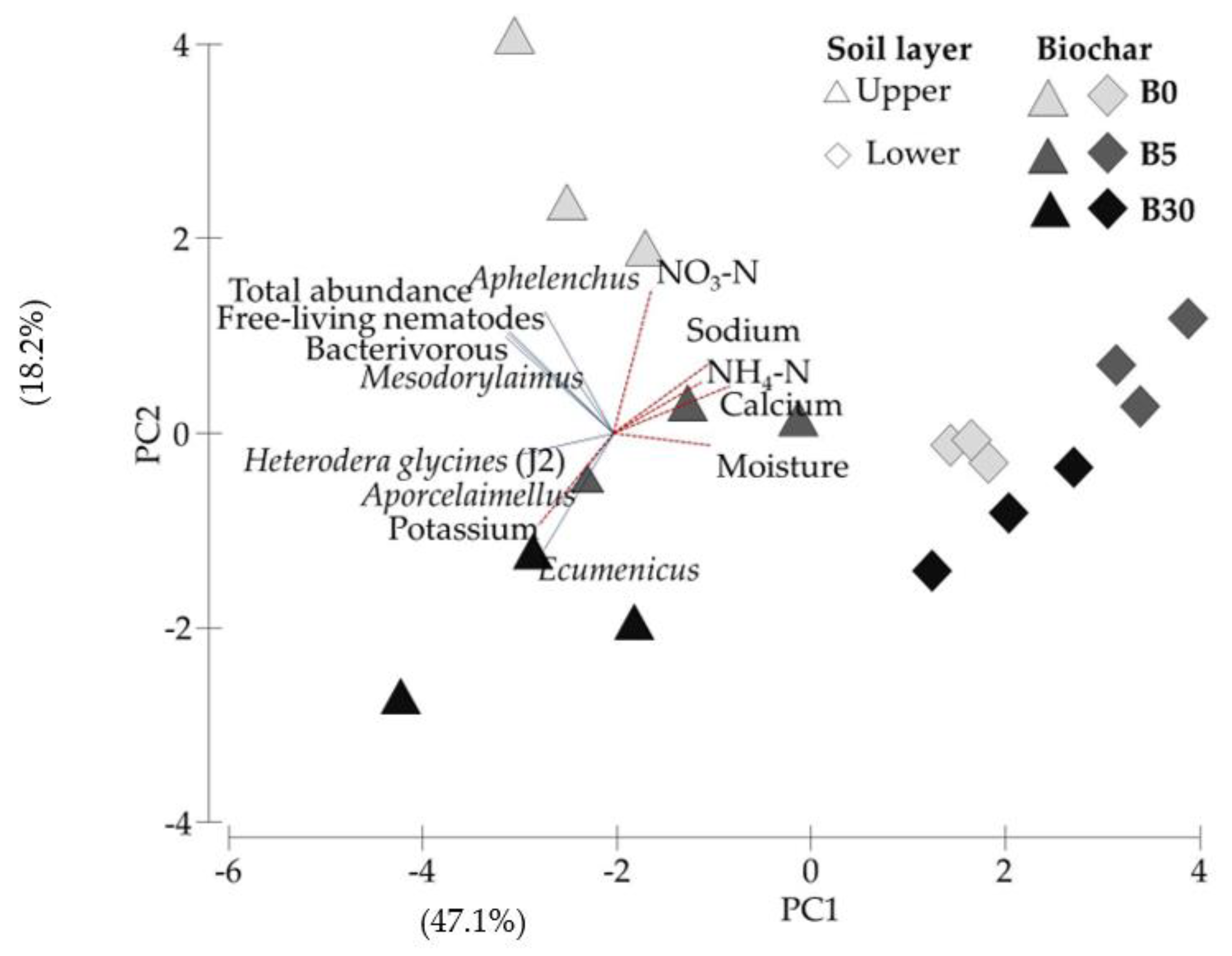
3.4. Principal Component Analysis
4. Discussion
4.1. Effects of Biochar Amendment to Upper Layer on the Chemical Properties in Upper and Lower Layer Soils
4.2. Biochar Reduced Bacterivorous in the Upper Layer but Stimulated Greater Trophic Guilds in the Lower Layer
4.3. Effect of Biochar Amendment to Upper Layer on the Density of Soybean Cyst Nematode (SCN) in the Lower Layer Soil
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lee, X.; Yang, F.; Xing, Y.; Huang, Y.; Xu, L.; Liu, Z.; Holtzman, R.; Kan, I.; Li, Y.; Zhang, L.; et al. Use of biochar to manage soil salts and water: Effects and mechanisms. Catena 2022, 211, 106018. [Google Scholar] [CrossRef]
- Joseph, S.; Cowie, A.L.; Van Zwieten, L.; Bolan, N.; Budai, A.; Buss, W.; Cayuela, M.L.; Graber, E.R.; Ippolito, J.A.; Kuzyakov, Y.; et al. How biochar works, and when it doesn’t: A review of mechanisms controlling soil and plant responses to biochar. GCB Bioenergy 2021, 13, 1731–1764. [Google Scholar] [CrossRef]
- Lu, T.; Wang, X.; Du, Z.; Wu, L. Impacts of continuous biochar application on major carbon fractions in soil profile of North China plain’s cropland: In comparison with straw incorporation. Agric. Ecosyst. Environ. 2021, 315, 107445. [Google Scholar] [CrossRef]
- Adebajo, S.O.; Oluwatobi, F.; Akintokun, P.O.; Ojo, A.E.; Akintokun, A.K.; Gbodope, I.S. Impacts of rice-husk biochar on soil microbial biomass and agronomic performances of tomato (Solanum lycopersicum L.). Sci. Rep. 2022, 12, 1787. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Fang, Z.; Van Zwieten, L.; Bolan, N.; Dong, D.; Quin, B.F.; Meng, J.; Li, F.; Wu, F.; Wang, H.; et al. A critical review of biochar-based nitrogen fertilizers and their effects on crop production and the environment. Biochar 2022, 4, 36. [Google Scholar] [CrossRef]
- Poveda, J.; Martínez-Gómez, Á.; Fenoll, C.; Escobar, C. The use of biochar for plant pathogen control. Phytopathology 2021, 111, 1490–1499. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Xiong, X.; Zhu, H.; Xu, H.; Leng, P.; Li, J.; Tang, C.; Xu, J. Association of biochar properties with changes in soil bacterial, fungal and fauna communities and nutrient cycling processes. Biochar 2021, 3, 239–254. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, Y.; Liu, B.; Amonette, J.E.; Lin, Z.; Liu, G.; Ambus, P.; Xie, Z. How does biochar influence soil N cycle? A meta-analysis. Plant Soil 2018, 426, 211–225. [Google Scholar] [CrossRef]
- Dawar, K.; Fahad, S.; Jahangir, M.M.R.; Munir, I.; Alam, S.S.; Khan, S.A.; Mian, I.A.; Datta, R.; Saud, S.; Banout, J.; et al. Biochar and urease inhibitor mitigate NH3 and N2O emissions and improve wheat yield in a urea fertilized alkaline soil. Sci. Rep. 2021, 11, 17413. [Google Scholar] [CrossRef]
- Borchard, N.; Schirrmann, M.; Cayuela, M.L.; Kammann, C.; Wrage-Mönnig, N.; Estavillo, J.M.; Fuertes-Mendizábal, T.; Sigua, G.; Spokas, K.; Ippolito, J.A.; et al. Biochar, soil and land-use interactions that reduce nitrate leaching and N2O emissions: A meta-analysis. Sci. Total Environ. 2019, 651, 2354–2364. [Google Scholar] [CrossRef]
- Tran Sy, N.; Huynh Van, T.; Nguyen Huu, C.; Nguyen Van, C.; Mitsunori, T. Rice husk and melaleuca biochar additions reduce soil CH4 and N2O emissions and increase soil physicochemical properties. F1000Research 2022, 10, 1128. [Google Scholar] [CrossRef]
- Karhu, K.; Kalu, S.; Seppanen, A.; Kitzler, B.; Virtanen, E. Potential of biochar soil amendments to reduce N leaching in boreal field conditions estimated using the resin bag method. Agric. Ecosyst. Environ. 2021, 316, 107452. [Google Scholar] [CrossRef]
- Gelardi, D.L.; Ainuddin, I.H.; Rippner, D.A.; Patiño, J.E.; Abou Najm, M.; Parikh, S.J. Biochar alters hydraulic conductivity and impacts nutrient leaching in two agricultural soils. SOIL 2021, 7, 811–825. [Google Scholar] [CrossRef]
- An, X.; Wu, Z.; Shi, W.; Qi, H.; Zhang, L.; Xu, X.; Yu, B. Biochar for simultaneously enhancing the slow-release performance of fertilizers and minimizing the pollution of pesticides. J. Hazard. Mater. 2021, 407, 124865. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Savla, N.; Pandit, C.; Pandit, S.; Gupta, P.K.; Pant, M.; Khilari, S.; Kumar, Y.; Agarwal, D.; Nair, R.R.; et al. Use of biomass-derived biochar in wastewater treatment and power production: A promising solution for a sustainable environment. Sci. Total Environ. 2022, 825, 153892. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elgawad, M.M.M. Plant-parasitic nematode threats to global food security. J. Nematol. 2014, 46, 130. [Google Scholar]
- Fábia, S.O.L.; Valdir, R.C.; Sônia, R.N.; Patrícia, R.R.S. Nematodes affecting soybean and sustainable practices for their management. In Soybean—The Basis of Yield, Biomass and Productivity; Kasai, M., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- Gendron St-Marseille, A.-F.; Bourgeois, G.; Brodeur, J.; Mimee, B. Simulating the impacts of climate change on soybean cyst nematode and the distribution of soybean. Agric. For. Meteorol. 2019, 264, 178–187. [Google Scholar] [CrossRef]
- Tefft, P.M.; Rende, J.F.; Bone, L.W. Factors influencing egg hatching of the soybean cyst nematode Heterodera glycines race-3. Proc. Helminthol. Soc. Wash. 1982, 49, 258–265. [Google Scholar]
- Ochola, J.; Coyne, D.; Cortada, L.; Haukeland, S.; Ng’ang’a, M.; Hassanali, A.; Opperman, C.; Torto, B. Cyst nematode bio-communication with plants: Implications for novel management approaches. Pest Manage. Sci. 2021, 77, 1150–1159. [Google Scholar] [CrossRef]
- Acharya, K.; Yan, G.; Plaisance, A. Effects of cover crops on population reduction of soybean cyst nematode (Heterodera glycines). Plant Dis. 2021, 105, 764–769. [Google Scholar] [CrossRef]
- Rocha, L.F.; Pimentel, M.F.; Bailey, J.; Wyciskalla, T.; Davidson, D.; Fakhoury, A.M.; Bond, J.P. Impact of wheat on soybean cyst nematode population density in double-cropping soybean production. Front. Plant Sci. 2021, 12, 640714. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.S.; Chikamatsu, S.; Kato, R.; Chau, M.K.; Nguyen, T.K.P.; Ritz, K.; Toyota, K. A biochar improves the efficacy of green manure-based strategies to suppress soybean cyst nematode (Heterodera glycines) and promotes free-living nematode populations. J. Soil Sci. Plant Nutr. 2022, 22, 3414–3427. [Google Scholar] [CrossRef]
- Chikamatsu, S.; Takeda, A.; Ohta, K.; Imura, T.; Perry, R.N.; Toyota, K. Suppression of the soybean cyst nematode, Heterodera glycines, by short-term field cultivation and soil incorporation of mung bean. Nematology 2021, 23, 305–315. [Google Scholar] [CrossRef]
- Linh, T.B.; Van, K.L.; Van Elsacker, S.; Cornelis, W.M. Effect of cropping system on physical properties of clay soil under intensive rice cultivation. Land. Degrad. Dev. 2016, 27, 973–982. [Google Scholar] [CrossRef]
- Nawaz, M.F.; Bourrie, G.; Trolard, F. Soil compaction impact and modelling. A review. Agron. Sustain. Dev. 2013, 33, 291–309. [Google Scholar] [CrossRef]
- Knox, O.; Polain, K.; Fortescue, E.; Griffiths, B. Distribution and restricted vertical movement of nematodes in a heavy clay soil. Agronomy 2020, 10, 221. [Google Scholar] [CrossRef]
- Bray, R.H.; Kurtz, L.T. Determination of total, organic, and available forms of phosphorus in soils. Soil Sci. 1945, 59, 39–46. [Google Scholar] [CrossRef]
- Houba, V.J.G.; Vanderlee, J.J.; Novozamsky, I. Soil and plant analysis: A series of syllabi. In Part 5b Soil Analysis Procedures Other Procedures, 6th ed.; Wageningen Agricultural University, Department of Soil Science and Plant Nutrition: Wageningen, The Netherlands, 1995. [Google Scholar]
- Kaneko, S.; Inagaki, M.; Morishita, T. A simple method for the determination of nitrate in potassium chloride extracts from forest soils. In Proceedings of the 19th World Congress of Soil Science, Soil Solutions for a Changing World, Brisbane, Australia, 1–6 August 2010; Published on DVD: Brisbane, Australia. [Google Scholar]
- Tabatabai, M. Soil enzymes. In Methods of Soil Analysis: Part 2 Microbiological and Biochemical Properties; Soil Science Society of America: Madison, WI, USA, 1994; Volume 5, pp. 775–833. [Google Scholar]
- Van Bezooijen, J. Methods and Techniques for Nematology; Wageningen University: Wageningen, The Netherlands, 2006. [Google Scholar]
- De Grisse, A. Redescription ou modifications de quelques techniques utililisées dans l’etude des nematodes phytoparasitaires. Meded. Rijksfac. Landbouwwet. Gent 1969, 34, 351–369. [Google Scholar]
- Eyualem-Abebe; Andrássy, I.; Traunspurger, W. Freshwater Nematodes: Ecology and Taxonomy; CAB International Publishing: Surrey, UK, 2006. [Google Scholar]
- Andrassy, I. A Taxonomic Review of the Suborder Rhabditina (Nematode: Secernnentia); Zoosystematica and Ecologica Institute, Eôtvôs Lorand University: Budapest, Hungary, 1983. [Google Scholar]
- Zulini, A. Identification Manual for Freshwater Nematode Genera; Lecture Book for Msc in Nematology Ghent University: Ghent, Belgium, 2010; p. 112. [Google Scholar]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G.M. Ninja: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Ferris, H. Form and function: Metabolic footprints of nematodes in the soil food web. Eur. J. Soil Biol. 2010, 46, 97–104. [Google Scholar] [CrossRef]
- Sato, E.; Min, Y.Y.; Toyota, K.; Takada, A. Relationships between the damage to radish caused by the root-lesion nematode Pratylenchus penetrans, its density prior to cultivation and the soil nematode community structure evaluated by polymerase chain reaction-denaturing gradient gel electrophoresis. Soil Sci. Plant Nutr. 2009, 55, 478–484. [Google Scholar] [CrossRef]
- Shirai, S.; Toyota, K. Optimisation of a species-specific primer set to quantify the soybean cyst nematode, Heterodera glycines, in soil using real-time PCR. Nematology 2019, 21, 1037–1042. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial (Plymouth Routines in Multivariate Ecological Research); PRIMER-E Ltd.: Plymouth, UK, 2006. [Google Scholar]
- Ding, Y.; Liu, Y.; Liu, S.; Li, Z.; Tan, X.; Huang, X.; Zeng, G.; Zhou, L.; Zheng, B. Biochar to improve soil fertility. A review. Agron. Sustain. Dev. 2016, 36, 36. [Google Scholar] [CrossRef]
- Liu, S.N.; Meng, J.; Jiang, L.L.; Yang, X.; Lan, Y.; Cheng, X.Y.; Chen, W.F. Rice husk biochar impacts soil phosphorous availability, phosphatase activities and bacterial community characteristics in three different soil types. Appl. Soil Ecol. 2017, 116, 12–22. [Google Scholar] [CrossRef]
- Munda, S.; Bhaduri, D.; Mohanty, S.; Chatterjee, D.; Tripathi, R.; Shahid, M.; Kumar, U.; Bhattacharyya, P.; Kumar, A.; Adak, T.; et al. Dynamics of soil organic carbon mineralization and C fractions in paddy soil on application of rice husk biochar. Biomass Bioenergy 2018, 115, 1–9. [Google Scholar] [CrossRef]
- Phuong, N.T.K.; Khoi, C.M.; Ritz, K.; Linh, T.B.; Minh, D.D.; Duc, T.A.; Sinh, N.V.; Linh, T.T.; Toyota, K. Influence of rice husk biochar and compost amendments on salt contents and hydraulic properties of soil and rice yield in salt-affected fields. Agronomy 2020, 10, 1101. [Google Scholar] [CrossRef]
- Gluba, Ł.; Rafalska-Przysucha, A.; Szewczak, K.; Łukowski, M.; Szlązak, R.; Vitková, J.; Kobyłecki, R.; Bis, Z.; Wichliński, M.; Zarzycki, R.; et al. Effect of fine size-fractionated sunflower husk biochar on water retention properties of arable sandy soil. Materials 2021, 14, 1335. [Google Scholar] [CrossRef]
- Ahmed, R.; Li, Y.; Mao, L.; Xu, C.; Lin, W.; Ahmed, S.; Ahmed, W. Biochar effects on mineral nitrogen leaching, moisture content, and evapotranspiration after 15N urea fertilization for vegetable crop. Agronomy 2019, 9, 331. [Google Scholar] [CrossRef]
- Pratiwi, E.P.A.; Shinogi, Y. Rice husk biochar application to paddy soil and its effects on soil physical properties, plant growth, and methane emission. Paddy. Water Environ. 2016, 14, 521–532. [Google Scholar] [CrossRef]
- Huang, H.; Reddy, N.G.; Huang, X.; Chen, P.; Wang, P.; Zhang, Y.; Huang, Y.; Lin, P.; Garg, A. Effects of pyrolysis temperature, feedstock type and compaction on water retention of biochar amended soil. Sci. Rep. 2021, 11, 7419. [Google Scholar] [CrossRef]
- Phuong, N.T.K.; Khoi, C.M.; Ritz, K.; Sinh, N.V.; Tarao, M.; Toyota, K. Potential use of rice husk biochar and compost to improve P availability and reduce GHG emissions in acid sulfate soil. Agronomy 2020, 10, 685. [Google Scholar] [CrossRef]
- Sánchez-García, M.; Roig, A.; Sánchez-Monedero, M.A.; Cayuela, M.L. Biochar increases soil N2O emissions produced by nitrification-mediated pathways. Front. Environ. Sci. 2014, 2, 25. [Google Scholar] [CrossRef]
- Wang, X.-F.; Li, J.; Li, G.; Zhang, G.-L.; Wang, Z.-W.; Zhi, Y.-C.; Wu, M.-L.; Lai, X.; Yang, D.-L.; Ren, T.-Z. Biochar application affects Nitrobacter rather than Nitrospira in plastic greenhouse vegetable soil. Appl. Soil Ecol. 2022, 175, 104449. [Google Scholar] [CrossRef]
- Zimmerman, A.R.; Gao, B.; Ahn, M.-Y. Positive and negative carbon mineralization priming effects among a variety of biochar-amended soils. Soil Biol. Biochem. 2011, 43, 1169–1179. [Google Scholar] [CrossRef]
- Abell, J.; Laverman, A.M.; Van Cappellen, P. Bioavailability of organic matter in a freshwater estuarine sediment: Long-term degradation experiments with and without nitrate supply. Biogeochemistry 2009, 94, 13–28. [Google Scholar] [CrossRef]
- Zavalloni, C.; Alberti, G.; Biasiol, S.; Vedove, G.D.; Fornasier, F.; Liu, J.; Peressotti, A. Microbial mineralization of biochar and wheat straw mixture in soil: A short-term study. Appl. Soil Ecol. 2011, 50, 45–51. [Google Scholar] [CrossRef]
- Hussain, M.; Farooq, M.; Nawaz, A.; Al-Sadi, A.M.; Solaiman, Z.M.; Alghamdi, S.S.; Ammara, U.; Ok, Y.S.; Siddique, K.H.M. Biochar for crop production: Potential benefits and risks. J. Soils Sed. 2017, 17, 685–716. [Google Scholar] [CrossRef]
- Brtnicky, M.; Datta, R.; Holatko, J.; Bielska, L.; Gusiatin, Z.M.; Kucerik, J.; Hammerschmiedt, T.; Danish, S.; Radziemska, M.; Mravcova, L.; et al. A critical review of the possible adverse effects of biochar in the soil environment. Sci. Total Environ. 2021, 796, 148756. [Google Scholar] [CrossRef]
- Liu, T.; Yang, L.H.; Hu, Z.K.; Xue, J.R.; Lu, Y.Y.; Chen, X.Y.; Griffiths, B.S.; Whalen, J.K.; Liu, M.Q. Biochar exerts negative effects on soil fauna across multiple trophic levels in a cultivated acidic soil. Biol. Fertil. Soils 2020, 56, 597–606. [Google Scholar] [CrossRef]
- Nguyen, V.S.; Kato, R.; Doan, T.T.L.; Nguyen, T.K.P.; Toyota, K. Influence of rice husk biochar on soil nematode community under upland and flooded conditions: A microcosm experiment. Agronomy 2022, 12, 378. [Google Scholar]
- Wang, C.Y.; Wang, Y.D.; Herath, H. Polycyclic aromatic hydrocarbons (PAHs) in biochar—Their formation, occurrence and analysis: A review. Org. Geochem. 2017, 114, 1–11. [Google Scholar] [CrossRef]
- Li, J.; Chen, Y.X.; Zhang, G.L.; Ruan, W.B.; Shan, S.J.; Lai, X.; Yang, D.L.; Yu, Z.G. Integration of behavioural tests and transcriptome sequencing of C. elegans reveals how the nematode responds to peanut shell biochar amendment. Sci. Total Environ. 2020, 707, 136024. [Google Scholar] [CrossRef] [PubMed]
- Domene, X.; Mattana, S.; Sanchez-Moreno, S. Biochar addition rate determines contrasting shifts in soil nematode trophic groups in outdoor mesocosms: An appraisal of underlying mechanisms. Appl. Soil Ecol. 2021, 158, 103788. [Google Scholar] [CrossRef]
- Domene, X.; Hanley, K.; Enders, A.; Lehmann, J. Short-term mesofauna responses to soil additions of corn stover biochar and the role of microbial biomass. Appl. Soil Ecol. 2015, 89, 10–17. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Z.; Yang, K.; Wang, P.; Wang, H.; Guo, L.; Zhu, S.; Zhu, Y.; He, X. Biochar application alleviated negative plant-soil feedback by modifying soil microbiome. Front. Microbiol. 2020, 11, 799. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Eyre, M.; Venette, R.C.; Lau, S.S. Population energetics of bacterial-feeding nematodes: Stage-specific development and fecundity rates. Soil Biol. Biochem. 1996, 28, 271–280. [Google Scholar] [CrossRef]
- Venette, R.C.; Ferris, H. Influence of bacterial type and density on population growth of bacterial-feeding nematodes. Soil Biol. Biochem. 1998, 30, 949–960. [Google Scholar] [CrossRef]
- Bouwman, L.A.; Arts, W.B.M. Effects of soil compaction on the relationships between nematodes, grass production and soil physical properties. Appl. Soil Ecol. 2000, 14, 213–222. [Google Scholar] [CrossRef]
- Singh, H.; Northup, B.K.; Rice, C.W.; Prasad, P.V.V. Biochar applications influence soil physical and chemical properties, microbial diversity, and crop productivity: A meta-analysis. Biochar 2022, 4, 8. [Google Scholar] [CrossRef]
- Charlson, D.V.; Tylka, G.L. Heterodera glycines cyst components and surface disinfestants affect H. glycines hatching. J. Nematol. 2003, 35, 458–464. [Google Scholar]
- Chikamatsu, S.; Xiaoman, W.; Ito, D.; Yamada, E.; Toyota, K. Effect of short-term growth of mung bean and its soil incorporation on the density of the soybean cyst nematode, Heterodera glycines, in pot experiments. Nematology 2017, 19, 1147–1155. [Google Scholar] [CrossRef]
- Fukuzawa, A.; Furusaki, A.; Ikura, M.; Masamune, T. Glycinoeclepin A, a natural hatching stimulus for the soybean cyst nematode. J. Chem. Soc. Chem. Commun. 1985, 4, 222–224. [Google Scholar] [CrossRef]
- Tefft, P.M.; Bone, L.W. Zinc-mediated hatching of eggs of soybean cyst nematode, Heterodera glycines. J. Chem. Ecol. 1984, 10, 361–372. [Google Scholar] [CrossRef]
- Clarke, A.J.; Perry, R.N. The role of egg-shell calcium in the hatching of Heterodera schachtii. Nematologica 1985, 31, 151–158. [Google Scholar] [CrossRef]
- Atkinson, H.J.; Ballantyne, A.J. Evidence for the involvement of calcium in the hatching of Globodera rostochiensis. Ann. Appl. Biol. 1979, 93, 191–198. [Google Scholar] [CrossRef]
- Clarke, A.J.; Perry, R.N. Egg-shell calcium and the hatching of Globodera rostochiensis. Int. J. Parasitol. 1985, 15, 511–516. [Google Scholar] [CrossRef]
- Gao, X.; Zhang, S.; Zhao, X.; Wu, Q. Potassium-induced plant resistance against soybean cyst nematode via root exudation of phenolic acids and plant pathogen-related genes. PLoS ONE 2018, 13, e0200903. [Google Scholar] [CrossRef] [PubMed]
- Li, T.J.; Wang, D.Y.; Li, B.X.; Wu, H.Y. The hatching of cereal cyst nematode (Heterodera avenae) in response to different inorganic ions. Sci. Agric. 2022, 79, e20200072. [Google Scholar] [CrossRef]
- Hodda, M. Phylum nematoda: Feeding habits for all valid genera using a new, universal scheme encompassing the entire phylum, with descriptions of morphological characteristics of the stoma, a key, and discussion of the evidence for trophic relationships. Zootaxa 2022, 5114, 318–451. [Google Scholar] [CrossRef]
- Moore, J.C.; McCann, K.; Setälä, H.; De Ruiter, P.C. Top-down is bottom-up: Does predation in the rhizosphere regulate aboveground dynamics? Ecology 2003, 84, 846–857. [Google Scholar] [CrossRef]
- Khan, Z.; Kim, Y.H. A review on the role of predatory soil nematodes in the biological control of plant parasitic nematodes. Appl. Soil Ecol. 2007, 35, 370–379. [Google Scholar] [CrossRef]
- Zhang, X.K.; Li, Q.; Liang, W.J.; Zhang, M.; Bao, X.L.; Xie, Z.B. Soil nematode response to biochar addition in a Chinese wheat field. Pedosphere 2013, 23, 98–103. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depths | Biochar | Total Abundance | Bacterivorous | Ecumenicus | Heterodera (J2) | Aphelenchus |
|---|---|---|---|---|---|---|
| Upper layer | B0 | 4307 ± 340 a | 3915 ± 150 a | 12 ± 8 a | 90 ± 26 a | 286 ± 178 a |
| B5 | 2042 ± 401 b | 1648 ± 291 b | 85 ± 57 a | 140 ± 50 a | 167 ± 31 a | |
| B30 | 2263 ± 163 b | 1876 ± 207 b | 168 ± 31 a | 126 ± 78 a | 91 ± 19 a | |
| Lower layer | B0 | 59 ± 8 a | 9 ± 2 a | 3 ± 2 b | 20 ± 5 a | 26 ± 7 a |
| B5 | 80 ± 4 a | 10 ± 4 a | 39 ± 9 a | 5 ± 4 a | 27 ± 8 a | |
| B30 | 78 ± 5 a | 13 ± 4 a | 31 ± 2 a | 9 ± 3 a | 26 ± 5 a |
| Treatments | Microbial Activity | Β-Glucosidase | Acid Phosphatase | Alkaline Phosphatase |
|---|---|---|---|---|
| B0 | 0.70 ± 0.06a | 17.0 ± 0.4 a | 227 ± 6 a | 196 ± 4 a |
| B5 | 1.09 ± 0.08a | 18.0 ± 0.5 a | 219 ± 1 a | 190 ± 3 a |
| B30 | 1.07 ± 0.22a | 17.1 ± 0.4 a | 234 ± 7 a | 190 ± 2 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sinh, N.V.; Linh, D.T.T.; Phuong, N.T.K.; Ritz, K.; Toyota, K. Transmissible Effects of a Biochar Amendment to an Upper Soil Zone Upon an Associated Lower Zone, Including Attenuation of Soybean Cyst Nematode (Heterodera glycines). Agronomy 2023, 13, 53. https://doi.org/10.3390/agronomy13010053
Sinh NV, Linh DTT, Phuong NTK, Ritz K, Toyota K. Transmissible Effects of a Biochar Amendment to an Upper Soil Zone Upon an Associated Lower Zone, Including Attenuation of Soybean Cyst Nematode (Heterodera glycines). Agronomy. 2023; 13(1):53. https://doi.org/10.3390/agronomy13010053
Chicago/Turabian StyleSinh, Nguyen Van, Doan Thi Truc Linh, Nguyen Thi Kim Phuong, Karl Ritz, and Koki Toyota. 2023. "Transmissible Effects of a Biochar Amendment to an Upper Soil Zone Upon an Associated Lower Zone, Including Attenuation of Soybean Cyst Nematode (Heterodera glycines)" Agronomy 13, no. 1: 53. https://doi.org/10.3390/agronomy13010053
APA StyleSinh, N. V., Linh, D. T. T., Phuong, N. T. K., Ritz, K., & Toyota, K. (2023). Transmissible Effects of a Biochar Amendment to an Upper Soil Zone Upon an Associated Lower Zone, Including Attenuation of Soybean Cyst Nematode (Heterodera glycines). Agronomy, 13(1), 53. https://doi.org/10.3390/agronomy13010053