
Overcoming Challenges for Shoot Tip Cryopreservation of Root and Tuber Crops


 ,
,  ,
,  ,
, 
Abstract
1. Introduction
1.1. Importance of Root and Tuber Crops to Humans
1.2. Conserving RTCs as the Filed, Greenhouse, and In Vitro Collections
1.3. Cryopreservation of RTCs and Its Challenges
2. Overview of Protocols to Cryopreserve Shoot Tip of RTCs
2.1. From Two-Step (Programmed) Cooling to Ultra-Rapid Freezing
2.1.1. Two-Step (Programmed) Cooling
2.1.2. Ultra-Rapid Freezing
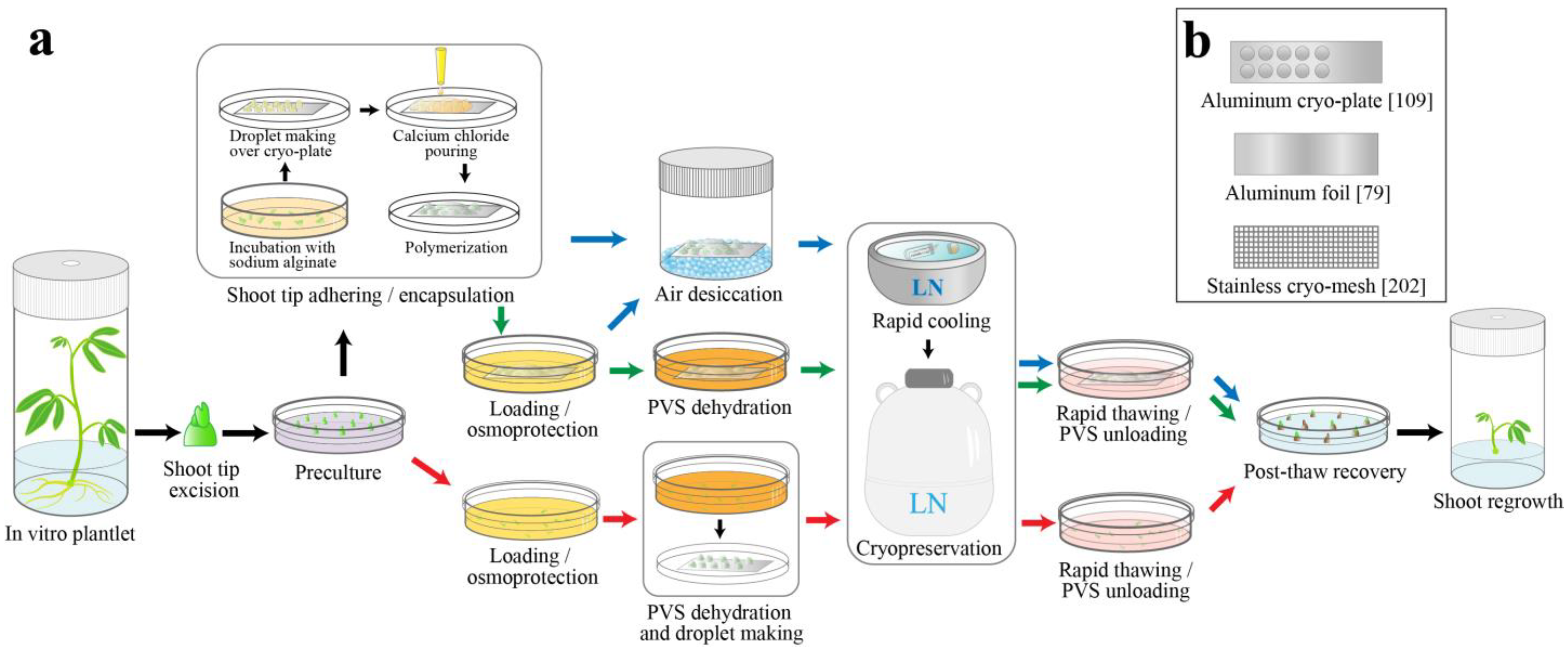
2.2. Development of Air-Drying Based Methods
2.2.1. Encapsulation-Dehydration Method (En-De)
2.2.2. Dehydration Cryo-Plate Method (D Cryo-Plate)
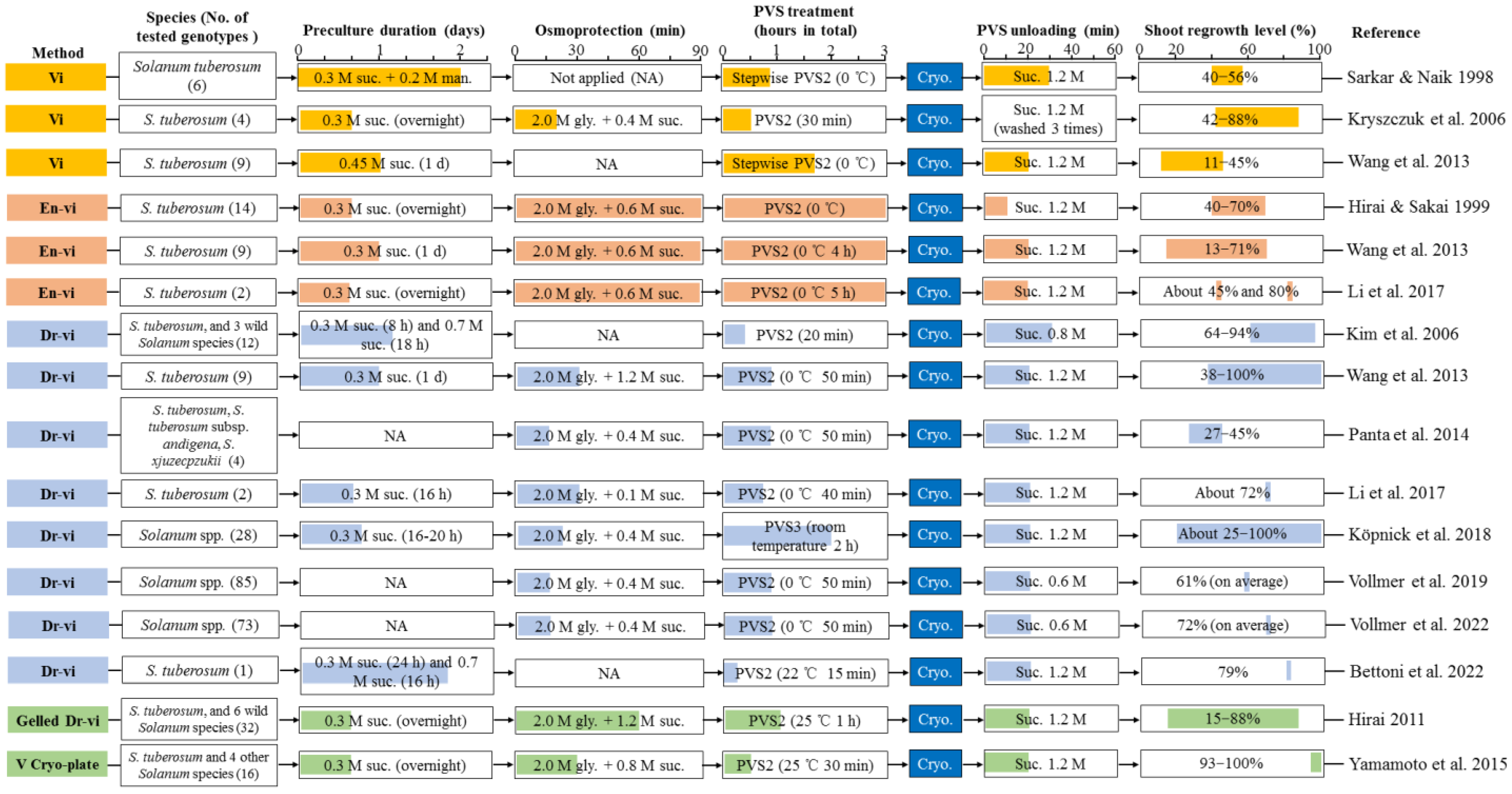
2.3. Development of Vitrification Solution-Based Methods
2.3.1. Vitrification and Vitrification Solution Method (Vi)
2.3.2. Encapsulation-Vitrification Method (En-vi)
2.3.3. Droplet-Vitrification Method (Dr-vi)
2.3.4. Vitrification Cryo-Plate Method (V Cryo-Plate)

{kind=link}
{kind=link}
{kind=link}
| Plant Species (No. of Tested Genotypes) | Pretreatment of Donor Plants and Preculture of Explants | Osmoprotection | PVS Dehydration and Unloading | Post-Thaw Culture (PTC) | Shoot Regrowth (%) | Ref. |
|---|---|---|---|---|---|---|
| Cassava [Manihot esculenta (48)] | Meristems (0.7 mm) excised from in vitro seedlings that are 3 to 4 weeks old, without preculture | 2 M gly with 0.4 M suc for 20 min at 24 °C | PVS2 for 30 min on ice; 2 M gly with 0.4 M suc for 15 min for unloading | With 0.3 M suc, 1.07 μM NAA, 0.23 μM GA3 and 0.66 μM BA for the first day and the same medium with 0.1 M suc for the rest of the recovery. The first 8 days of PTC was in darkness | 6–86 | [146] |
| M. esculenta (9) | Meristems (0.5 mm) excised from cultures that are 3 weeks old, without preculture | 2 M gly with 0.4 M suc for 20–60 min | PVS2 for 30 min on ice; 1.2 M gly with 0.4 M suc for unloading | With 0.3 M suc, 1 mg L−1 ascorbic acid and then transfer to standard medium for final recovery. The first 7 days of PTC was in darkness | About 60–98 | [70] |
| M. esculenta (100) which showed recalcitrance to En-de (potst-thaw regrowth rate <30%) | Shoot tips (2 mm) excised from cultures that are 3 months old, without preculture | 2 M gly with 0.4 M suc for 2 h | PVS2 for 30 min on ice; 1.2 M suc for unloading | With 0.3 M suc and 0.2% active charcoal for 2 days in darkness, and then transfer to basal medium with 2.32 μM Kinetin, 0.72 μM GA3 and vitamins | Higher than 30% for 70–75% tested accessions | [72] |
| Sweet potato [Ipomoea batatas (1)] | Apical shoot tips (0.5–1.0 mm) excised from cultures that are 4 to 8 weeks old, cultured on liquid basal medium with 2% suc for 1 day, followed by preculture with 0.3 M suc for another day | 2 M gly with 0.4 M suc for 2 h | PVS2 for 16 min at 22 °C; 1.2 M suc for unloading | With 1 μM NAA, 0.5 μM BA, 0.1 μM Kinetin for 2 days in the dark, under dim light for 3 days, before being transferred to normal conditions | 62 | [141] |
| I. betatas (1) | Shoot tips (length of 2.5 and a width of 1.5 mm) excised from cultures that are 5 to 7 weeks old and stepwise precultured with 0.3 and 0.5 M suc for 31 and 17 h, respectively | 2 M gly with 0.5 M suc for 50 min | PVS3 for 1 h at room temperature; 0.9 M suc for unloading | With 1 g L−1 casein hydrolysate, 1 mg L−1 GA3, 0.5 mg L−1 BA for 1 week in the dark, and then cultured with 1 g L−1 casein hydrolysate, 0.5 mg L−1 GA3 for further recovery | 19 | [128] |
| I. betatas (30) | Apical shoot tips 1.0–1.2 mm excised from 4–8-week-old cultures and precultured 1 d with 0.35 M suc | 2 M gly with 0.4 M suc for 20 min | PVS2 for 30 min on ice; 1.2 M suc for unloading | First 9 days of PTC in darkness with 10 mg L−1 GA3, 10 mL coconut water, and 0.3, 0.1, 0.09 M stepwise with decreased suc level, each level for 3 days, and then transferred to fresh medium with 0.09 M suc under diffuse light for 4 days before being moved to the normal conditions | 2–66 | [142] |
| I. betatas (10) | Axillary meristems (1 mm) excised from cultures that are 3 to 9 weeks old, without preculture | 2 M gly with 0.4 M suc for 20 min | PVS2 for 30 min on ice; 1.2 M suc for unloading | With 0.3 M suc overnight in the dark, and then moved to regeneration medium with 2.22 μM BA for 7 days in the dark, before being transferred to the normal conditions | 10–84 | [73] |
| Yam [Dioscorea bulbifera, D. alata, D. cayenensis, D. polystachya (4)] | Apical shoot tips (2–4 mm) excised from shoots after 3 weeks of cold-hardening and precultured with 0.3 or 0.44 M sucrose for 3 days | 2 M gly with 0.4 M suc for 20 min | PVS2 for 20 min at 23 °C; thawing was performed with 0.09 M suc for 3 min followed by 5 min unloading with 1.17 M suc | Recovered on MS with 2 mg L−1 BA and 0.1 mg L−1 NAA | 0 for D. alata and 27–47 for the other species | [147] |
| D. alata, D. rotundata (21) | Meristems (0.5 mm) excised from cultures that are 3 weeks old, without preculture | 2 M gly with 0.4 M suc for 20–60 min | PVS2 for 30 min on ice; 1.2 M gly with 0.4 M suc for unloading | With 0.3 M suc and 1 mg L−1 ascorbic acid, and then transfer to standard meristem medium for final recovery. The first 7 days of PTC was in darkness | 0–60 | [70] |
| D. deltoidei (15) | Shoot tips (2 mm) excised from cultures that are 4 weeks old and precultured with 0.3 M sucrose for 16 h | 2 M gly with 0.4 M suc for 20 min | PVS2 for 90 min at 0 °C; 1.2 M suc for unloading | With 1.5 mg L−1 BA, 0.2 mg L−1 NAA, 0.2 mg L−1 GA3 for 10 days, and then transferred to medium with 0.5 mg L−1 zeatin. The first 5 days of PTC was in darkness | 21–51 | [74] |
| Oca [Oxalis tuberosa (4)] | Shoot tips (2 mm) excised from cultures that are 3 weeks old, without preculture | 2 M gly with 0.4 M suc for 20 min | PVS2 for 1 h on ice; 1.2 M suc for unloading | With 0.04 mg L−1 kinetin, 0.1 mg L−1 GA3 and 0.3 M suc for the first 2 days in the dark, and then with 0.1 M suc for another 2 days in the dark, and finally plated with 0.07 M suc and 2 mg L−1 calcium pantothenate for final recovery | 7–15 | [75] |
| D. deltoidei (15) | Shoot tips (2 mm) excised from cultures that are 4 weeks old and precultured with 0.3 M sucrose for 16 h | 2 M gly with 0.4 M suc for 20 min | PVS2 for 90 min at 0 °C; 1.2 M suc for unloading | With 1.5 mg L−1 BA, 0.2 mg L−1 NAA, 0.2 mg L−1 GA3 for 10 days, and then transferred to medium with 0.5 mg L−1 zeatin. The first 5 days of PTC was in darkness | 21–51 | [74] |
| Oca [Oxalis tuberosa (4)] | Shoot tips (2 mm) excised from cultures that are 3 weeks old, without preculture | 2 M gly with 0.4 M suc for 20 min | PVS2 for 1 h on ice; 1.2 M suc for unloading | With 0.04 mg L−1 kinetin, 0.1 mg L−1 GA3 and 0.3 M suc for the first 2 days in the dark, and then with 0.1 M suc for another 2 days in the dark, and finally plated with 0.07 M suc and 2 mg L−1 calcium pantothenate for final recovery | 7–15 | [75] |
| Taro [Colocasia esculenta (18)] | Apical hoot tips (0.8–1 mm) excised from in vitro shoots cultured on MS with 0.26 M suc for 4–8 weeks | 2 M gly with 0.4 M suc for 20 min | PVS2 for 20–40 min on ice; 1.2 M suc for unloading | With 0.3 M suc overnight in the dark, and then with 0.1 M suc for another 5 days in the dark and 10 days under dim light | 73–100 | [148] |
| C. esculenta (1) | Apical shoot tips (0.8–1.0 mm) excised from cultures that are 3 months old and precultured with 0.3 M sucrose overnight | 2 M gly with 0.4 M suc and 5% DMSO for 20 min | PVS3 for 10 min; 1.2 M suc for unloading | With 0.3 M suc for 2 days in the dark, and then transferred to basal medium with 0.1 M for final recovery for the next 13 days | 78 (survival) | [154] |
| Ulluco [Ullucus tuberosus (1)] | Apical shoot tips excised from the lateral shoots of in vitro nodal sections cultured on MS with 2 M suc for 5 days and precultured with 0.7 M suc for 12 h | Not specified | PVS3 for 1.5 h; thawing was performed with cryovials by immersing into a 40 °C water bath for 30 min. Unloading of PVS3 was exempted | PTC was performed with 0.5 mg L−1 kinetin and IAA, and 0.2 mg L−1 GA3 | 52.5 | [155] |
| U. tuberosus (4) | Shoot tips (2 mm) excised from cultures that are 3 weeks old, without preculture | 2 M gly with 0.4 M suc for 20 min | PVS2 for 1 h on ice; 1.2 M suc for unloading | Similar to the PTC protocol of Oca [Sánchez et al. 2011] | 11–35 | [75] |
| Yacon [Smallanthus sonchifolius (5)] | Apical shoot tips (2–3 mm) excised from cultures that are 2–3 weeks old and precultured with 0.3 M sucrose in darkness overnight | 2 M gly with 0.4 M suc for 20 min | PVS2 (0 °C) or PVS3 (22 °C) for 60 min; 1.2 M and 0.3 M suc were used for unloading PVS2 and PVS3, respectively | For the shoot tips treated with PVS2, PTC begins with 0.3 M suc for 1 day in darkness followed by basal MS medium for another 6 days in the dark. For the shoot tips treated with PVS3, PTC was performed with basal MS medium for the first 7 days in the dark | 66–75 | [149] |
| Callerya speciosa (Champ.) Schot | Axillary shoot tips (1.5 mm) excised from cultures that are 40 days old | 2 M gly with 0.4 M suc for 4 h on a shaker at 150 rpm | PVS2 (0 °C) for 60 min on a shaker at 150 rpm; 1.2 M suc for unloading | PTC begins with 0.3 M suc for 2 days, and were then transferred to MS with 2.2 μM BA, 0.6 μM IAA and 0.1 Μm GA3 for final recovery, first 6 days of PTC were in the dark | 60 | [156] |
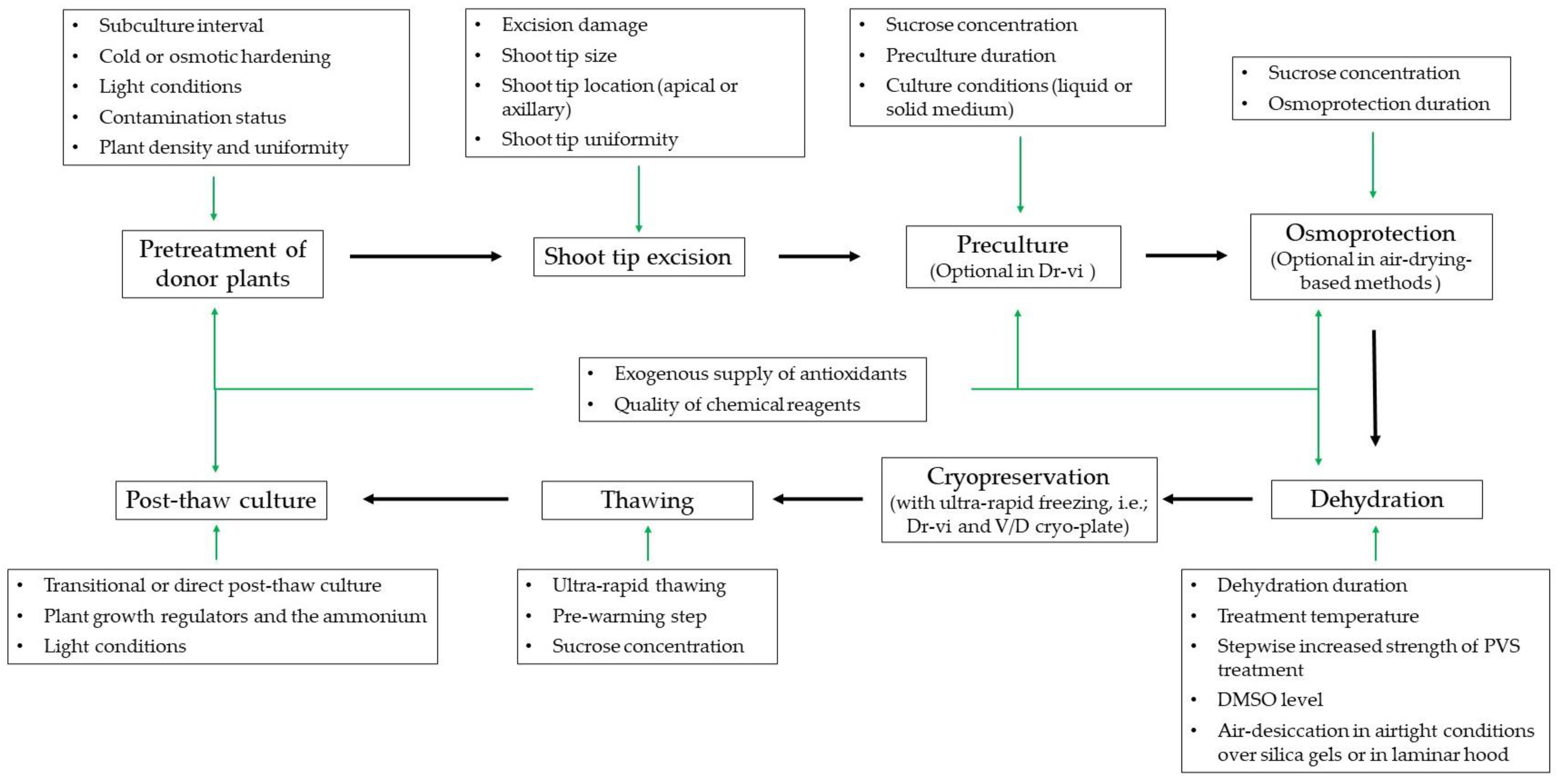
3. Key Strategies for Improving the Shoot Regrowth of RTCS after Cryopreservation
3.1. Pretreatment and Type of Plant Materials
3.2. Pathogen-Free Status
3.3. Preculture and Osmoprotection Conditions
3.4. Dehydration Methods
3.5. Alleviation of Oxidative Stress and Freezing Injury
3.6. Improved Thawing and Post-Thaw Culture
3.6.1. The Level of Sucrose in the Thawing Process
3.6.2. The Level of Sucrose and Ammonium in Post-Thaw Culture
3.6.3. The Influence of Plant Growth Regulators on Post-Thaw Recovery
3.7. Light Conditions
4. Exploring the Scientific Basis behind Shoot Tip Recovery after Cryopreservation
4.1. Metabolic Responses to the Cold and Osmotic Treatments
4.2. Anti-Oxidative/Freezing Responses behind Shoot Tip Cryopreservation
4.3. Understanding the Responses of Cell and Tissue to Cryogenic Treatment under Microscope
5. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zierer, W.; Rüscher, D.; Sonnewald, U.; Sonnewald, S. Tuber and tuberous root development. Annu. Rev. Plant Biol. 2021, 72, 551–580. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekara, A.; Kumar, T.J. Roots and tuber crops as functional foods: A review on phytochemical constituents and their potential health benefits. Int. J. Food Sci. 2016, 2016, 3631647. [Google Scholar] [CrossRef]
- Bradshaw, J.E.; Bonierbale, M. Potatoes. In Root and Tuber Crops; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 1–52. [Google Scholar] [CrossRef]
- Lebot, V. Sweet Potato. In Root and Tuber Crops; Bradshw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 97–125. [Google Scholar]
- Epping, J.; Laibach, N. An underutilized orphan tuber crop—Chinese yam: A review. Planta 2020, 252, 58. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.K. A comprehensive review of sweet potato (Ipomoea batatas [L.] Lam): Revisiting the associated health benefits. Trends Food Sci. Technol. 2021, 115, 512–529. [Google Scholar] [CrossRef]
- Leidi, E.O.; Altamirano, A.M.; Mercado, G.; Rodriguez, J.P.; Ramos, A.; Alandia, G.; Sørensen, M.; Jacobsen, S.-E. Andean roots and tubers crops as sources of functional foods. J. Funct. Foods 2018, 51, 86–93. [Google Scholar] [CrossRef]
- FAOSTAT. 2020. Available online: https://www.fao.org/faostat/en/#data/QC (accessed on 23 June 2022).
- OECD/FAO. OECD-FAO Agricultural Outlook 2021–2030; OECD Publishing: Paris, France, 2021. [Google Scholar] [CrossRef]
- Devaux, A.; Goffart, J.P.; Kromann, P.; Andrade-Piedra, J.; Polar, V.; Hareau, G. The potato of the future: Opportunities and challenges in sustainable Agri-food systems. Potato Res. 2021, 64, 681–720. [Google Scholar] [CrossRef]
- Machida-Hirano, R. Diversity of potato genetic resources. Breed Sci. 2015, 65, 26–40. [Google Scholar] [CrossRef]
- Devaux, A.; Goffart, J.-P.; Petsakos, A.; Kromann, P.; Gatto, M.; Okello, J.; Suarez, V.; Hareau, G. Global Food Security, Contributions from Sustainable Potato Agri-Food Systems. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 3–36. [Google Scholar] [CrossRef]
- Zaheer, K.; Akhtar, M.H. Potato Production, Usage, and Nutrition—A Review. Cri. Rev. Food Sci. Nutr. 2016, 56, 711–721. [Google Scholar] [CrossRef]
- Liu, B.; Gu, W.; Yang, Y.; Lu, B.; Wang, F.; Zhang, B.; Bi, J. Promoting potato as staple food can reduce the carbon-land-water impacts of crops in China. Nat. Food 2021, 2, 570–577. [Google Scholar] [CrossRef]
- Ceballos, H.; Okogbenin, E.; Pérez, J.C.; Augusto, L.; López-Valle, B.; Debouck, D. Cassava. In Root and Tuber Crops; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 53–96. [Google Scholar] [CrossRef]
- Muiruri, S.K.; Ntui, V.O.; Tripathi, L.; Tripathi, J.N. Mechanisms and approaches towards enhanced drought tolerance in cassava (Manihot esculenta). Curr. Plant Biol. 2021, 28, 100227. [Google Scholar] [CrossRef]
- Amelework, A.B.; Bairu, M.W.; Maema, O.; Vender, S.L.; Laing, M. Adoption and Promotion of Resilient Crops for Climate Risk Mitigation and Import Substitution: A Case Analysis of Cassava for South African Agriculture. Front. Sustain. Food Syst. 2021, 5, 617783. [Google Scholar] [CrossRef]
- Tang, C.; Lu, Y.; Jiang, B.; Chen, J.; Mo, X.; Yang, Y.; Wang, Z. Energy, Economic, and Environmental Assessment of Sweet Potato Production on Plantations of Various Sizes in South China. Agronomy 2022, 12, 1290. [Google Scholar] [CrossRef]
- Motsa, N.M.; Modi, A.T.; Mabhaudhi, T. Sweet potato (Ipomoea batatas L.) as a drought tolerant and food security crop. S. Afr. J. Sci. 2015, 111, 1–8. [Google Scholar] [CrossRef]
- Nimoh, F.; Asare, G.O.; Twumasi, I.; Anaman, R. Consumers’ Willingness to Consume Cassava Leaves as a Leafy Vegetable in the Kumasi Metropolis, Ghana. Int. J. Food Stud. 2018, 7, 38–50. [Google Scholar] [CrossRef]
- Li, M.; Zi, X.; Tang, J.; Xu, T.; Gu, L.; Zhou, H. Effects of cassava foliage on feed digestion, meat quality, and antioxidative status of geese. Poult. Sci. 2020, 99, 423–429. [Google Scholar] [CrossRef]
- Fathima, A.A.; Sanitha, M.; Tripathi, L.; Muiruri, S. Cassava (Manihot esculenta) dual use for food and bioenergy: A review. Food Energy Secur. 2022, 00, e380. [Google Scholar] [CrossRef]
- Tay, D. Tropical and Subtropical Root and Tuber Crops. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer: New York, NY, USA, 2013; pp. 249–298. [Google Scholar] [CrossRef]
- Lebot, V. Aroids, Cassava. In Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yams and Aroids, 2nd ed.; Stubbs, R., Ed.; CABI: Oxfordshire, UK, 2020; pp. 40–42. [Google Scholar]
- Arnau, G.; Abraham, K.; Sheela, M.N.; Chair, H.; Sartie, A.; Asiedu, R. Yams. In Root and Tuber Crops; Bradshaw, J.E., Ed.; Springer New York: London, UK, 2010; pp. 127–148. [Google Scholar] [CrossRef]
- Quero-Garcia, J.; Ivancic, A.; Lebot, V. Taro and Cocoyam. In Root and Tuber Crops; Bradshaw, J.E., Ed.; Springer: New York, NY, USA, 2010; pp. 149–172. [Google Scholar] [CrossRef]
- Li Pun, H.H.; Mares, V.; Quiroz, R.; León Velarde, C.U.; Valdivia, R.; Reinoso, J. Pursuing the Millennium Development Goals in the Andean Altiplano. Mt. Res. Dev. 2006, 26, 15–19. [Google Scholar] [CrossRef]
- Olusegun, O.V.; Obi-Egbedi, O.; Adeniran, L.O. Root and tuber expansion programme and poverty reduction among farmers in Southwest Nigeria. J. Dev. Agric. Econ. 2015, 7, 332–343. [Google Scholar] [CrossRef]
- Aboajah, F.N.; Ejechi, M.E.; Viashima, S.S.; Adeyongu, S.K.; Muogbo, P.C. Sweet Potato Production for Poverty Alleviation in Nasarawa State, Nigeria. Int. J. Environ. Agric. Biotech. 2018, 3, 380–387. [Google Scholar] [CrossRef]
- Agbarevo, M.N.B.; Okringbo, J.I. Effect of Technologies of National Root Crops Research Institute, Umudike on Poverty Reduction among Farmers in Umuahia Agricultural Zone, Abia State. J. Community Commun. Res. 2020, 5, 83–90. [Google Scholar]
- van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A meta-analysis of projected global food demand and population at risk of hunger for the period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef]
- van Ittersuma, M.K.; van Bussela, L.G.J.; Wolfa, J.; Grassinib, P.; van Wartb, J.; Guilpartb, N.; Claessensc, L.; de Grootd, H.; Wiebee, K.; Mason-D’Croz, D.; et al. Can sub-Saharan Africa feed itself? Proc. Natl. Acad. Sci. USA 2016, 13, 14964–14969. [Google Scholar] [CrossRef] [PubMed]
- Galluzzi, G.; López Noriega, I. Conservation and Use of Genetic Resources of Underutilized Crops in the Americas—A Continental Analysis. Sustainability 2014, 6, 980–1017. [Google Scholar] [CrossRef]
- Ellis, D.; Salas, A.; Chavez, O.; Gomez, R.; Anglin, N. Ex Situ Conservation of Potato [Solanum Section Petota (Solanaceae)] Genetic Resources in Genebanks. In The Potato Crop; Campos, H., Ortiz, O., Eds.; Springer: Cham, Switzerland, 2020; pp. 109–138. [Google Scholar]
- Izquierdo, J.; Roca, W. Under-ultilized Andean Food Crops: Status and Prospects of Plant Biotechnology for The conservation and Sustainable Agricultural Use of Genetic Resources. Acta Hortic. 1998, 457, 157–172. [Google Scholar] [CrossRef]
- Flores, H.E.; Walker, T.S.; Guimarães, R.L.; Bais, H.P.; Vivanco, J.M. Andean Root and Tuber Crops: Underground Rainbows. HortScience 2003, 38, 161–167. [Google Scholar] [CrossRef]
- CIPotato. 2022. Available online: https://cipotato.org/potato/wild-potato-species/ (accessed on 16 July 2022).
- CIP Genebanks, CGIAR Genebank Platform. 2022. Available online: https://www.genebanks.org/genebanks/international-potato-centre/ (accessed on 1 October 2022).
- Jarvis, A.; Lane, A.; Hijmans, R.J. The effect of climate change on crop wild relatives. Agric. Ecosyst. Environ. 2008, 126, 13–23. [Google Scholar] [CrossRef]
- Brummitt, N.A.; Bachman, S.T.; Griffiths-Lee, J.; Lutz, M.; Moat, J.F.; Farjon, A.; Donaldson, J.S.; Hilton-Taylor, C.; Meagher, T.R.; Albuquerque, S.; et al. Green Plants in the Red: A Baseline Global Assessment for the IUCN Sampled Red List Index for Plants. PLoS ONE 2015, 10, e0135152. [Google Scholar] [CrossRef]
- FAO. Genebank Standards for Plant Genetic Resources for Food and Agriculture. 2014. Available online: https://www.fao.org/3/i3704e/i3704e.pdf (accessed on 2 January 2023).
- Lebot, V. Aroids, Origin and History. In Tropical Root and Tuber Crops: Cassava, Sweet Potato, Yams and Aroids, 2nd ed.; Stubbs, R., Ed.; CABI: Oxfordshire, UK, 2020; pp. 329–334. [Google Scholar]
- Panis, B.; Nagel, M.; Van den houwe, I. Challenges and Prospects for the Conservation of Crop Genetic Resources in Field Genebanks, in In Vitro Collections and/or in Liquid Nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Lüttringhaus, S.; Pradel, W.; Suarez, V.; Manrique-Carpintero, N.C.; Anglin, N.L.; Ellis, D.; Hareau, G.; Jamora, N.; Smale, M.; Gómez, R. Dynamic guardianship of potato landraces by Andean communities and the genebank of the International Potato Center. CABI Agric. Biosci. 2021, 2, 1–16. [Google Scholar] [CrossRef]
- Reed, B.M.; Gupta, S.; Uchendu, E.E. In Vitro Genebanks for Preserving Tropical Biodiversity. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer: New York, NY, USA, 2013; pp. 77–106. [Google Scholar] [CrossRef]
- Bamberg, J.B.; Martin, M.W.; Abad, J.; Jenderek, M.M.; Tanner, J.; Donnelly, D.J.; Nassar, A.M.K.; Veilleux, R.E.; Novy, R.G. In vitro technology at the US Potato Genebank. In Vitro Cell. Dev. Biol.-Plant 2016, 52, 213–225. [Google Scholar] [CrossRef]
- Engelmann, F. In vitro conservation methods. In Biotechnology and Plant Genetic Resources: Conservation and Use; Ford-Lloyd, B.V., Newbury, J.H., Callow, J.A., Eds.; CABI International: Wallingford, UK, 1997; pp. 119–162. [Google Scholar]
- Ruta, C.; Lambardi, M.; Ozudogru, E.A. Biobanking of vegetable genetic resources by in vitro conservation and cryopreservation. Biodivers. Conserv. 2020, 29, 3495–3532. [Google Scholar] [CrossRef]
- Bairu, M.W.; Aremu, A.O.; van Staden, J. Somaclonal variation in plants: Causes and detection methods. Plant Growth Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Van den houwe, I.; Chase, R.; Sardos, J.; Ruas, M.; Kempenaers, E.; Guignon, V.; Massart, S.; Carpentier, S.; Panis, B.; Rouard, M.; et al. Safeguarding and using global banana diversity: A holistic approach. CABI Agric. Biosci. 2020, 1, 15. [Google Scholar] [CrossRef]
- Reed, B.M.; Engelmann, F.; Dulloo, M.E.; Engels, J.M.M. Technical guidelines for the management of field and in vitro germplasm collections. In IPGRI Handbooks for Genebanks No. 7; International Plant Genetic Resources Institute: Rome, Italy, 2004; p. 106. [Google Scholar]
- Sakai, A.; Engelmann, F. Vitrification, Encapsulation-vitrification and Droplet-Vitrification: A review. Cryo-Lett. 2007, 28, 151–172. [Google Scholar]
- Benson, E.E.; Harding, K. Cryopreservation of Shoot Tips and Meristems: An Overview of Contemporary Methodologies. In Plant Cell Culture Protocols, Methods in Molecular Biology; Loyola-Vargas, V.M., Ochoa-Alejo, N., Eds.; Humana: Totowa, NJ, USA, 2012; Volume 877, pp. 191–226. [Google Scholar] [CrossRef]
- Sadia, B.; Anthony, P.; Lowe, C.K.; Power, J.B.; Davey, M.R. Culture treatments for enhancing post-thaw recovery of cryopreserved suspension cells of potato cv. Desiree. Cell. Mol. Biol. Lett. 2003, 8, 979–989. [Google Scholar] [PubMed]
- Kaczmarczyk, A.; Rokka, V.-M.; Keller, E.R.J. Potato Shoot Tip Cryopreservation. A Review. Potato Res. 2011, 54, 45–79. [Google Scholar] [CrossRef]
- Yamamoto, S.; Wunna; Rafique, T.; Valle Arizaga, M.; Fukui, K.; Cruz Cutierrez, E.J.; Castillo Martinez, C.R.; Watanabe, K.; Niino, T. The Alumium Cryo-plate Increases Efficiency of Cryopreservation Protocols for Potato Shoot tips. Am. J. Potato Res. 2015, 92, 250–257. [Google Scholar] [CrossRef]
- Smyda-Dajmund, P. Cryopreservation of shoot tips and pollen of potato. Plant Breed. Seed Sci. 2017, 76, 75–80. [Google Scholar] [CrossRef]
- Vollmer, R.; Espirilla, J.; Villagaray, R.; Cárdenas, J.; Castro, M.; Sánchez, J.C.; Manrique-Carpintero, N.; Ellis, D.; Anglin, N.L. Cryopreservation of Potato Shoot Tips for Long-Term Storage. Methods Mol. Biol. 2021, 2354, 21–54. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Bonnart, R.; Volk, G.M. Challenges in implementing plant shoot tip cryopreservation technologies. Plant Cell Tissue Organ Cult. 2021, 144, 21–34. [Google Scholar] [CrossRef]
- Wang, M.-R.; Lambardi, M.; Engelmann, F.; Pathirana, R.; Panis, B.; Volk, G.M.; Wang, Q.-C. Advances in cryopreservation of in vitro-derived propagules: Technologies and explant sources. Plant Cell Tissue Organ Cult. 2021, 144, 7–20. [Google Scholar] [CrossRef]
- Zámečník, J.; Faltus, M.; Bilavcik, A. Vitrification Solutions for Plant Cryopreservation: Modification and Properties. Plants 2021, 10, 2623. [Google Scholar] [CrossRef] [PubMed]
- Westcott, R.J.; Henshaw, G.G.; Roca, W.M. Tissue culture storage of potato germplasm: Culture initiation and plant regeneration. Plant Sci. Lett. 1977, 9, 309–315. [Google Scholar] [CrossRef]
- Grout, B.W.W.; Henshaw, G.G. Freeze Preservation of Potato Shoot-tip Cultures. Ann. Bot. 1978, 42, 1227–1229. [Google Scholar] [CrossRef]
- Engelmann, F.; Arnao, M.T.G.; Wu, Y.; Escobar, R. Development of Encapsulation Dehydration. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 59–75. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Kretzschmar, A.A.; Bonnart, R.; Shepherd, A.; Volk, G.M. Cryopreservation of 12 Vitis species using apical shoot tips derived from plants grown in vitro. HortScience 2019, 54, 976–981. [Google Scholar] [CrossRef]
- Panis, B. Sixty years of plant cryopreservation: From freezing hardy mulberry twigs to establishing reference crop collections for future generations. Acta Hortic. 2019, 1234, 1–7. [Google Scholar] [CrossRef]
- Jenderek, M.M.; Reed, B.M. Cryopreserved storage of clonal germplasm in the USDA National Plant Germplasm System. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 299–308. [Google Scholar] [CrossRef]
- Vollmer, R.; Villagaray, R.; Castro, M.; Cárdenas, J.; Pineda, S.; Espirilla, J.; Anglin, N.; Ellis, D.; Azevedo, V. The world’s largest potato cryobank at the International Potato Center (CIP)—Status quo, protocol improvement through large-scale experiments and long-term viability monitoring. Front. Plant Sci. 2022, 13, 1059817. [Google Scholar] [CrossRef] [PubMed]
- Niino, T.; Arizaga, M.V. Cryopreservation for preservation of potato genetic resources. Breed Sci. 2015, 65, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Dumet, D.; Diebiru, E.; Adeyemi, A.; Akinyemi, O.; Gueye, B.; Franco, J. Cryopreservation for the ‘In Perpetuity’ Conservation of Yam and Cassava Genetic Resources. Cryo-Lett. 2013, 34, 107–118. [Google Scholar]
- Acker, J.P.; Adkins, S.; Alves, A.; Horna, D.; Toll, J. Feasibility Study for a Safety Back-Up Cropreservation Facility. Independent Expert Report; Bioversity International: Rome, Italy, 2017. [Google Scholar]
- Escobar, R.H.; Muñoz, L.; Rios, A.; Núñez, A.; Tohme, J. Using a Droplet-Vitrification Method to Partially Overcome the Recalcitrance of Cassava to Cryostorage. Acta Hortic. 2014, 1039, 227–232. [Google Scholar] [CrossRef]
- Wilms, H.; Sleziak, N.F.; Van der Auweraer, M.; Brands, M.; Verleije, M.; Hardeman, D.; Andre, E.; Panis, B. Development of a fast and user-friendly cryopreservation protocol for sweet potato genetic resources. Sci. Rep. 2020, 10, 14674. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Malhotra, E.V.; Chandra, R.; Gowthami, R.; Sultan, S.M.; Bansal, S.; Shankar, M.; Agrawal, A. Cryopreservation and genetic stability assessment of regenerants of the critically endangered medicinal plant Dioscorea deltoidea Wall. ex Griseb. for cryobanking of germplasm. In Vitro Cell. Dev. Biol.-Plant 2022, 58, 521–529. [Google Scholar] [CrossRef]
- Sánchez, D.F.; Panta, A.; Tay, D.; Roca, W. Cryopreservation of Ulluco (Ullucus tuberosus Cal.) and Oca (Oxalis tuberosa Mol.) Shoot Tips Using the PVS2 Droplet-Vitrification Method. Acta Hortic. 2011, 908, 339–346. [Google Scholar] [CrossRef]
- Bajaj, Y.P.S. Initiation of shoots and callus from potato-tuber sprouts and axillary buds frozen at −196 °C. Crop Improv. 1977, 4, 48–53. [Google Scholar]
- Kartha, K.K.; Leung, N.L.; Mroginski, L.A. In vitro Growth Responses and Plant Regeneration from Cryopreserved Meristems of Cassava (Manihot esculenta Crantz). Z. Pflanzenphysiol. 1982, 107, 133–140. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Panis, B.; Piette, B.; Swennen, R. Droplet vitrification of apical meristems: A cryopreservation protocol applicable to all Musaceae. Plant Sci. 2005, 168, 45–55. [Google Scholar] [CrossRef]
- Elliott, G.D.; Wang, S.; Fuller, B.J. Cryoprotectants: A review of the actions and applications of cryoprotective solutes that modulate cell recovery from ultra-low temperatures. Cryobiology 2017, 76, 74–91. [Google Scholar] [CrossRef]
- Schäfer-Menuhr, A.; Schumacher, H.M.; Mix-Wagner, G. Langzeitlagerung alter Kartoffelsorten durch Kryokonservierung der Meristeme. Landbauforsch. Völkenrode 1994, 44, 301–313. [Google Scholar]
- Schäfer-Menuhr, A.; Müller, E.; Mix-Wagner, G. Cryopreservation: An alternative for the long-term storage of old potato cultivars. Potato Res. 1996, 39, 507–513. [Google Scholar] [CrossRef]
- Schäfer-Menuhr, A.; Schumacher, H.M.; Mix-Wagner, G. Cryopreservation of potato cultivars-design of a method for routine application in genebanks. Acta Hortic. 1997, 447, 447–482. [Google Scholar] [CrossRef]
- Mix-Wagner, G.; Schumacher, H.M.; Cross, R.J. Recovery of Potato Apices After Several Years of Storage in Liquid Nitrogen. Cryo-Lett. 2002, 24, 33–41. [Google Scholar]
- Towill, L.E. Improved Survival after Cryogenic Exposure of Shoot Tips Derived from In Vitro Plantlet Cultures of Potato. Cryobiology 1983, 20, 567–573. [Google Scholar] [CrossRef]
- Towill, L.E. Survival at ultra-low temperatures of shoot tips from Solanum tuberosum groups Andigena, Phureja, Stenotomum, Tuberosum, and other tuber-bearing Solanum species. Cryo-Lett. 1984, 5, 319–326. [Google Scholar]
- Benson, E.E.; Harding, K.; Smith, H. Variation in recovery of cryopreserved shoot-tips of Solanum tuberosum exposed to different pre- and post-freeze light regimes. Cryo-Lett. 1989, 10, 323–344. [Google Scholar]
- Escobar, R.H.; Mafla, G.; Roca, W.M. A methodology for recovering cassava plants from shoot tips maintained in liquid nitrogen. Plant Cell Rep. 1997, 16, 474–478. [Google Scholar] [CrossRef]
- Bhatti, M.H.; Percival, T.; Davey, C.D.M.; Henshaw, G.G.; Blakesley, D. Cryopreservation of embryogenic tissue of a range of genotypes of sweet potato (Ipomoea Batatas [L.] Lam.) using an encapsulation protocol. Plant Cell Rep. 1997, 16, 802–806. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.-A.; Dhital, S.P.; Fang, Y.-L.; Khu, D.-M.; Song, Y.-S.; Park, E.-J.; Kang, C.-W.; Lim, H.-T. Application of Slow-Freezing Cryopreservation Method for the Conservation of Diverse Potato (Solanum tuberosum L.) Genotypes. J. Plant Biotechnol. 2005, 7, 183–186. [Google Scholar]
- Vollmer, R.; Villagaray, R.; Cárdenas, J.; Castro, M.; Chávez, O.; Anglin, N.L.; Ellis, D. A large-scale viability assessment of the potato cryobank at the International Potato Center (CIP). In Vitro Cell. Dev. Biol.-Plant 2017, 53, 309–317. [Google Scholar] [CrossRef]
- Fabre, J.; Dereuddre, J. Encapsulation dehydration a new approach to cryopreservation of Solanum shoot-tips. Cryo-Lett. 1990, 11, 413–426. [Google Scholar]
- Bouafia, S.; Jelti, N.; Lairy, G.; Blanc, A.; Bonnel, E.; Dereuddre, J. Cryopreservation of potato shoot tips by encapsulation-dehydration. Potato Res. 1996, 39, 69–78. [Google Scholar] [CrossRef]
- Grospietsch, M.; Stodulková, E.; Zámecnik, J. Effect of osmotic stress on the dehydration tolerance of Solanum tuberosum shoot tips. Cryo-Lett. 1999, 20, 339–346. [Google Scholar]
- Matsumoto, T.; Sakai, A. An approach to enhance dehydration tolerance of alginate-coated dried meristems cooled to −196 °C. Cryo-Lett. 1995, 16, 299–306. [Google Scholar]
- Takagi, H. Recent developments in cryopreservation of shoot tips of tropical species. In Cryopreservation of Tropical Plant Germplasm; Engelmann, F., Takagi, H., Eds.; International Plant Genetics Research Institute: Rome, Italy, 2000; pp. 178–193. [Google Scholar]
- Engelmann, F.; Benson, E.E.; Chabrillange, N.; Gonzalez Arnao, M.T.; Mari, S.; Michaux-Ferriere, N.; Paulet, F.; Glaszmann, J.C.; Charrier, A. Cryopreservation of Several Tropical Plant Species using Encapsulation/Dehydration of Apices. In Current Issues in Plant Molecular and Cellular Biology. Current Plant Science and Biotechnology in Agriculture; Terzi, M., Cella, R., Falavigna, A., Eds.; Springer: Dordrecht, The Netherlands, 1995; Volume 22, pp. 315–320. [Google Scholar] [CrossRef]
- Benson, E.E.; Harding, K.; Ryan, M.; Petrenko, A.; Petrenko, Y.; Fuller, B. Alginate Encapsulation to Enhance Biopreservation Scope and Success: A Multidisciplinary Review of Current Ideas and Applications in Cryopreservation and Non-freezing Storage. Cryo-Lett. 2018, 39, 14–38. [Google Scholar]
- Pennycooke, J.C.; Towill, L.E. Medium alternations improve regrowth of sweet potato (Ipomoea batatas [L.] Lam.) shoot tips cryopreserved by vitrification and encapsulation-dehydration. Cryo-Lett. 2001, 22, 381–389. [Google Scholar]
- Mandal, B.B.; Chandel, K.P.S.; Dwivedi, S. Cryopreservation of Yam (Dioscorea spp.) Shoot Apices by Encapsulation-Dehydration. Cryo-Lett. 1996, 17, 165–174. [Google Scholar]
- Malaurie, B.; Trouslot, M.F.; Engelmann, F.; Chabrillange, N. Effect of Pretreatment Conditions on the Cryopreservation of in vitro-cultured Yam (Dioscorea alata ‘Brazo Fuerte’ and D. bulbifera ‘Nouméa Imboro’) Shoot Apices by Encapsulation-Dehydration. Cryo-Lett. 1998, 19, 15–26. [Google Scholar]
- Hong, S.-R.; Yin, M.-H. A simple cryopreservation protocol for in vitro-grown shoot tips of Chinese genuine red bud taro (Colocasia esculenta L. Schott Var. Cormosus CV. Hongyayu) by encapsulation-dehydration. Sci. Hortic. 2013, 162, 226–233. [Google Scholar] [CrossRef]
- Valle Arizaga, M.; Yamamoto, S.I.; Tanaka, D.; Fukui, K.; Nohara, N.; Nishikawa, T.; Kazuo, W.; Niino, T. Cryopreservation of in vitro shoot tips of ulluco (Ullucus tuberosus Cal.) using D cryo-plate method. Cryo-Lett. 2017, 38, 419–427. [Google Scholar]
- Niino, T.; Yamamoto, S.; Fukui, K.; Martínez, C.R.C.; Arizaga, M.V.; Matsumoto, T.; Engelmann, F. Dehydration Improves Cryopreservation of Mat Rush (Juncus decipiens Nakai) Basal Stem Buds on Cryo-plates. Cryo-Lett. 2013, 34, 549–560. [Google Scholar]
- Scottez, C.; Chevreau, E.; Godard, N.; Arnaud, Y.; Duron, M.; Dereuddre, J. Cryopreservation of Cold-Acclimated Shoot Tips of Pear In Vitro Cultures after Encapsulation-Dehydration. Cryobiology 1992, 29, 691–700. [Google Scholar] [CrossRef]
- Niino, T.; Sakai, A. Cryopreservation of alginate coated in vitro-grown shoot tips of apple, pear and mulberry. Plant Sci. 1992, 87, 199–206. [Google Scholar] [CrossRef]
- Hirai, D.; Sakai, A. Cryopreservation of in vitro-grown meristems of potato (Solanum tuberosum L.) by encapsulation-vitrification. Potato Res. 1999, 42, 153–160. [Google Scholar] [CrossRef]
- Kim, H.-H.; Yoon, J.-W.; Park, Y.-E.; Cho, E.-G.; Sohn, J.-K.; Kim, T.-S.; Engelmann, F. Cryopreservation of Potato Cultivated Varieties and Wild Species: Critical Factors in Droplet Vitrification. Cryo-Lett. 2006, 27, 223–234. [Google Scholar]
- Yamamoto, S.-i.; Rafique, T.; Priyantha, W.S.; Fukui, K.; Matsumoto, T.; Niino, T. Development of a cryopreservation procedure using aluminum cryo-plates. Cryo-Lett. 2011, 32, 256–265. [Google Scholar]
- Niino, T.; Yamamoto, S.; Matsumoto, T.; Engelmann, F.; Arizaga, M.V.; Tanaka, D. Development of V and D cryo-plate methods as effective protocols for cryobanking. Acta Hortic. 2019, 1234, 249–262. [Google Scholar] [CrossRef]
- Valle Arizaga, M.; Navarro, O.F.V.; Martinez, C.R.C.; Gutiérrez, E.J.C.; Delgado, H.A.L.; Yamamoto, S.; Watanabe, K.; Niino, T. Improvement to the D Cryo-plate Protocol Applied to Practical Cryopreservation of In Vitro Grown Potato Shoot Tips. Hort. J. 2017, 86, 222–228. [Google Scholar] [CrossRef]
- Tanaka, D.; Sakuma, Y.; Yamamoto, S.; Valle Arizaga, M.; Niino, T.; Masumoto, T. Development of −80 °C storage for Allium shoot tips using D cryo-plate method. Plant Cell Tissue Organ Cult. 2021, 144, 115–122. [Google Scholar] [CrossRef]
- Faltus, M.; Bilavcik, A.; Zamecnik, J. Vitrification Ability of Combined and Single Cryoprotective Agents. Plants 2021, 10, 2392. [Google Scholar] [CrossRef]
- Matsumoto, T.; Sakai, A.; Yamada, K. Cryopreservation of in vitro-grown apical meristems of wasabi (Wasabia japonica) by vitrification and subsequent high plant regeneration. Plant Cell Rep. 1994, 13, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Sakai, A. Cryopreservation of axillary shoot tips of in vitro-grown grape (Vitis) by a two-step vitrification protocol. Euphytica 2003, 131, 299–304. [Google Scholar] [CrossRef]
- Zhu, G.-Y.; Geuns, J.M.C.; Dussert, S.; Swennen, R.; Panis, B. Change in sugar, sterol and fatty acid composition in banana meristems caused by sucrose-induced acclimation and its effects on cryopreservation. Physiol. Plant. 2006, 128, 80–94. [Google Scholar] [CrossRef]
- Leunufna, S.; Keller, E.R.J. Investigating a new cryopreservation protocol for yams (Dioscorea spp.). Plant Cell Rep. 2003, 21, 1159–1166. [Google Scholar] [CrossRef]
- Kryszczuk, A.; Keller, J.; Grübe, M.; Zimnoch-Guzowska, E. Cryopreservation of potato (Solanum tuberosum L.) shoot tips using vitrification and droplet method. J. Food. Agric. Environ. 2006, 4, 196–200. [Google Scholar]
- Sakai, A.; Kobayashi, S.; Oiyama, I. Cryopreservation of nucellar cells of navel orange (Citrus sinensis Osb. var. brasiliensis Tanaka) by vitrification. Plant Cell Rep. 1990, 9, 30–33. [Google Scholar] [CrossRef]
- Matsumoto, T.; Niino, T. Manual of Cryopreservation Methods Using Cryo-Plate: V and D Cryo-Plate Procedure as an Effective Protocol for Cryobanks; Niino, T., Matsumoto, T., Yamamoto, S.-I., Maki, S., Tanata, D., Engelmann, F., Eds.; Plant Tissue Culture and Cryopreservation Group (PTCCcryoG): Tsukuba, Japan, 2017; pp. 8–15. [Google Scholar]
- Matsumoto, T.; Niino, T. The development of plant vitrification solution 2 and recent PVS2-based vitrification protocols. Acta Hortic. 2014, 1039, 21–28. [Google Scholar] [CrossRef]
- Towill, L.E.; Jarrett, R.L. Cryopreservation of sweet potato (Ipomoea batatas [L.] Lam.) shoot tips by vitrification. Plant Cell Rep. 1992, 11, 175–178. [Google Scholar] [CrossRef]
- Sarkar, D.; Naik, P.S. Cryopreservation of shoot tips of tetraploid potato (Solanum tuberosum L.) clones by vitrification. Ann. Bot. 1998, 82, 455–461. [Google Scholar] [CrossRef]
- Wang, B.; Wang, R.; Li, J.; Ma, Y.; Sheng, W.; Li, M.; Wang, Q. Development of three vitrification-based cryopreservation procedures for shoot tips of China’s potato. Cryo-Lett. 2013, 34, 369–380. [Google Scholar]
- Charoensub, R.; Phansiri, S.; Sakai, A.; Yongmanitchai, W. Cryopreservation of cassava in vitro-grown shoot tips collet to −196 °C by vitrification. Cryo-Lett. 1999, 20, 89–94. [Google Scholar]
- Mukherjee, P.; Mandal, B.B.; Bhat, K.V.; Biswas, A.K. Cryopreservation of Asian Dioscorea bulbifera L. and D. alta L. by Vitrification: Importance of Plant Growth Regulators. Cryo-Lett. 2009, 20, 100–111. [Google Scholar]
- Takagi, H.; Thinh, N.T.; Islam, O.M.; Senboku, T.; Sakai, A. Cryopreservation of in vitro-grown shoot tips of taro (Colocasia esculenta (L.) Schott) by vitrification. 1. Investigation of basic conditions of the vitrification procedure. Plant Cell Rep. 1997, 16, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-U.; Kim, H.-H. Cryopreservation of Sweet Potato Shoot Tips Using a Droplet-Vitrification Procedure. Cryo-Lett. 2015, 36, 344–352. [Google Scholar]
- Li, J.-W.; Chen, H.-Y.; Li, X.-Y.; Zhang, Z.; Blystad, D.-R.; Wang, Q.-C. Cryopreservation and evaluations of vegetative growth, microtuber production and genetic stability in regenerants of purple-fleshed potato. Plant Cell Tissue Organ Cult. 2017, 128, 641–653. [Google Scholar] [CrossRef]
- Panta, A.; Panis, B.; Ynouye, C.; Swennen, R.; Roca, W. Development of a PVS2 Droplet Vitrification Method for Potato Cryopreservation. Cryo-Lett. 2014, 35, 255–266. [Google Scholar]
- Köpnick, C.; Grübe, M.; Stock, J.; Senula, A.; Mock, H.-P.; Nagel, M. Changes of soluble sugars and ATP content during DMSO droplet freezing and PVS3 droplet vitrification of potato shoot tips. Cryobiology 2018, 85, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, R.; Villagaray, R.; Castro, M.; Anglin, N.L.; Ellis, D. Cryopreserved potato shoot tips showed genotype-specific response to sucrose concentration in rewarming solution (RS). Plant Cell Tissue Organ Cult. 2019, 136, 353–363. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Mathew, L.; Pathirana, R.; Wiedow, C.; Hunter, D.A.; McLachlan, A.; Khan, S.; Tang, J.; Nadarajan, J. Eradication of Potato Virus S, Potato Virus A, and Potato Virus M from Infected in vitro Grown Potato Shoots Using in vitro Therapies. Front. Plant Sci. 2022, 13, 878733. [Google Scholar] [CrossRef]
- Hirai, D. Gelled Droplet Vitrification Improves Recovery of Cryopreserved Potato Germplasm. Cryo-Lett. 2011, 32, 287–296. [Google Scholar]
- Tannoury, M.; Ralambosoa, J.; Kaminski, M.; Dereuddre, J. Cryopreservation by vitrification of coated shoot tips of carnation (Diathus caryophyllus L.) cultured in vitro. C. R. Acad. Sci. Paris Ser. III 1991, 313, 633–638. [Google Scholar]
- Charoensub, R.; Hirai, D.; Sakai, A. Cryopreservation of in vitro-grown shoot tips of cassava by encapsulation-vitrification method. Cryo-Lett. 2004, 25, 51–58. [Google Scholar]
- Hirai, D.; Sakai, A. Simplified cryopreservation of sweet potato [Ipomoea batatas (L.) Lam.] by optimizing conditions for osmoprotection. Plant Cell Rep. 2003, 21, 961–966. [Google Scholar] [CrossRef] [PubMed]
- Yi, J.Y.; Lee, G.A.; Lee, Y.Y.; Gwag, J.G.; Son, E.H.; Park, H.J. Cryopreservation of in Vitro Grown Shoot Tips of Sweet Potato (Ipomoea batatas L.) by the Encapsulation-Vitrification Method. Korean J. Plant Res. 2016, 29, 635–641. [Google Scholar] [CrossRef]
- Panis, B.; Piette, B.; André, E.; Van den Houwe, I.; Swennen, R. Droplet vitrification: The first generic cryopreservation protocol for organized plant tissues? Acta Hortic. 2011, 908, 157–163. [Google Scholar] [CrossRef]
- Matsumoto, T. Cryopreservation of plant genetic resources: Conventional and new methods. Rev. Agric. Sci. 2017, 5, 13–20. [Google Scholar] [CrossRef]
- Pennycooke, J.C.; Towill, L.E. Cryopreservation of shoot tips from in vitro plants of sweet potato [Ipomoea batatas (L.) Lam.] by vitrification. Plant Cell Rep. 2000, 19, 733–737. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, R.; Panta, A.; Tay, D.; Roca, W.; Ellis, D. Effect of Sucrose Preculture and PVS2 Exposure on the Cryopreservation of Sweet Potato Shoot Tips [Ipomea batatas (L.) Lam.] Using the PVS2 Droplet Vitrification. Acta Hortic. 2014, 1039, 265–271. [Google Scholar] [CrossRef]
- Halmagyi, A.; Deliu, C.; Coste, A. Plant regrowth from potato shoot tips cryopreserved by a combined vitrification-droplet method. Cryo-Lett. 2005, 26, 313–322. [Google Scholar]
- Yoon, J.-W.; Kim, H.-H.; Ko, H.-C.; Hwang, H.-S.; Hong, E.-S.; Cho, E.-G.; Engelmann, F. Cryopreservation of Cultivated and Wild Potato Varieties by Droplet Vitrification: Effect of Subculture of Mother-Plants and of Preculture of Shoot Tips. Cryo-Lett. 2006, 27, 211–222. [Google Scholar]
- Panta, A.; Panis, B.; Ynouye, C.; Swennen, R.; Roca, W.; Tay, D.; Ellis, D. Improved cryopreservation method for the long-term conservation of the world potato germplasm collection. Plant Cell Tissue Organ Cult. 2015, 120, 117–125. [Google Scholar] [CrossRef]
- Dumet, D.; Korie, S.; Adeyemi, A. Cryobanking Cassava Germplasm at IITA. Acta Hortic. 2009, 908, 439–446. [Google Scholar] [CrossRef]
- Leunufna, S.; Keller, E.R.J. Cryopreservation of Yam Using Vitrification Modified by Including Droplet Method: Effects of Cold Acclimation and Sucrose. Cryo-Lett. 2005, 26, 93–102. [Google Scholar]
- Sant, R.; Panis, B.; Taylor, M.; Tyagi, A. Cryopreservation of shoot tips by droplet vitrification applicable to taro (Colocasia esculenta var. esculenta) accessions. Plant Cell Tissue Organ Cult. 2008, 92, 107–111. [Google Scholar] [CrossRef]
- Hammond, S.D.H.; Viehmannova, I.; Zámečník, J.; Panis, B.; Faltus, M. Droplet-vitrification methods for apical bud cryopreservation of yacon [Smallanthus sonchifolius (Poepp. and Endl.) H. Rob.]. Plant Cell Tissue Organ Cult. 2021, 147, 197–208. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Marković, Z.; Bi, W.; Volk, G.M.; Matsumoto, T.; Wang, Q.-C. Grapevine Shoot Tip Cryopreservation and Cryotherapy: Secure Storage of Disease-Free Plants. Plants 2021, 10, 2190. [Google Scholar] [CrossRef]
- CIP. Cryopreservation. Available online: https://cipotato.org/genebankcip/process/cryopreservation/ (accessed on 31 August 2022).
- Bettoni, J.C.; Bonnart, R.; Shepherd, A.; Kretzschmar, A.A.; Volk, G.M. Modifications to a Vitis shoot tip cryopreservation procedure: Effect of shoot tip size and use of cryoplates. Cryo-Lett. 2019, 40, 103–112. [Google Scholar]
- Agrawal, A.; Sharma, N.; Gupta, S.; Bansal, S.; Srivastava, V.; Malhotra, E.V.; Chander, S.; Gowthami, R.; Singh, K. Biotechnological applications for plant germplasm conservation at ICAR-National Bureau of Plant Genetic Resources, India—Recent achievements. Acta Hortic. 2022, 1339, 29–41. [Google Scholar] [CrossRef]
- Noor Camellia, N.A.; Abdul Muhaimin, A.K.; Umikalsum, M.B.; Rozlaily, Z. Cryopreservation of In vitro Grown Taro (Colocasia esculenta) Apical Meristem using a Droplet-vitrification PVS3 Technique. Tran. Malays. Soc. Plant Physiol. 2021, 28, 240–245. [Google Scholar]
- Zamecnikova, J.; Fernandez, E.; Viehmannova, I.; Zamecnik, J.; Faltus, M. Preparation of Shoot Tips by Sucrose and Dehydration Pre-Treatment of Ullucus tuberosus Cal. for Cryopreservation. Acta Hortic. 2011, 908, 331–338. [Google Scholar] [CrossRef]
- Li, Z.; Li, T.; Xu, L.; Panis, B. Cryopreservation of Callerya speciosa (Champ.) Schot Through Droplet-Vitrification. Propag. Ornam. Plants 2013, 13, 189–195. [Google Scholar]
- Agrawal, A.; Gowthami, R.; Chander, S.; Srivastava, V. Sustainability of In Vitro Genebanks and Cryogenebanks. Indian J. Genet. Plant Breed. 2022, 35, 180–184. [Google Scholar] [CrossRef]
- Ruiz-Sáenz, D.R.; Ayala-Hernández, D.D.; Niino, T.; Cruz-Gutiérrez, E.J.; Aquino-Martínez, J.G.; López-Delgado, H.A. Salicylic Acid-Cryotherapy Treatment for Elimination of Potato Virus S from Solanum Tuberosum. Am. J. Potato Res. 2019, 96, 225–234. [Google Scholar] [CrossRef]
- Folgado, R.; Panis, B.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F. Unravelling the effect of sucrose and cold pretreatment on cryopreservation of potato through sugar analysis and proteomics. Cryobiology 2015, 71, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.-H.; Tsai, S.-F.; Liaw, S.-L. Cryopreservation of Sweet Potato Using Vitrification Technique: Effects of the Vitrification Procedure and Sucrose Pretreatments. Propag. Ornam. Plants 2021, 21, 123–130. [Google Scholar]
- Edesi, J.; Pirttilä, A.M.; Häggman, H. Modified light spectral conditions prior to cryopreservation alter growth characteristics and cryopreservation success of potato (Solanum tuberosum L.) shoot tips in vitro. Plant Cell Tissue Organ Cult. 2017, 128, 409–421. [Google Scholar] [CrossRef]
- Charoensub, R.; Phansiri, S.; Yongmanitchai, W.; Sakai, A. Routine cryopreservation of in vitro-grown axillary apices of cassava (Manihot esculenta Crantz) by vitrification: Importance of a simple mononodal culture. Sci. Hortic. 2003, 98, 485–492. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Shukla, M.; Saxena, P.K.; Keller, J.E.R. Cryopreservation of potato microtubers: The critical roles of sucrose and desiccation. Plant Cell Tissue Organ Cult. 2016, 124, 649–656. [Google Scholar] [CrossRef]
- Volk, G.M.; Shepherd, A.N.; Bonnart, R. Successful cryopreservation of Vitis shoot tips: Novel pre-treatment combinations applied to nine species. Cryo-Lett. 2018, 39, 322–330. [Google Scholar]
- Wang, B.; Ma, Y.-L.; Zhang, Z.-B.; Wu, Z.-M.; Wu, Y.-F.; Wang, Q.-C.; Li, M.-F. Potato viruses in China. Crop Prot. 2011, 30, 1117–1123. [Google Scholar] [CrossRef]
- Legg, J.P.; Kumar, P.L.; Makeshkumar, T.; Tripathi, L.; Ferguson, M.; Kanju, E.; Ntawuruhunga, P.; Cuellar, W. Chapter Four—Cassava Virus Diseases: Biology, Epidemiology, and Management. Adv. Virus Res. 2015, 91, 85–142. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.P. Sweet Potato: Ipomoea batatas. In Plant Protection in Tropical Root and Tuber Crops; Springer: New Delhi, India, 2015; pp. 119–134. [Google Scholar]
- Luo, G.F.; Podolyan, A.; Kidanemariam, D.B.; Pilotti, C.; Houliston, G.; Sukal, A.C. A Review of Viruses Infecting Yam (Dioscorea spp.). Viruses 2022, 14, 662. [Google Scholar] [CrossRef] [PubMed]
- Priegnitz, U.; Lommen, W.J.M.; van der Vlugt, R.A.A.; Struik, P.C. Potato yield and yield components as affected by positive selection during several generations of seed multiplication in southwestern Uganda. Potato Res. 2000, 63, 507–543. [Google Scholar] [CrossRef]
- Thomas-Sharma, S.; Abdurahman, A.; Ali, S.; Andrade-Piedra, J.L.; Bao, S.; Charkowski, A.O.; Crook, D.; Kadian, M.; Kromann, P.; Struik, P.C.; et al. Seed degeneration in potato: The need for an integrated seed health strategy to mitigate the problem in developing countries. Plant Pathol. 2015, 65, 3–16. [Google Scholar] [CrossRef]
- Hadidi, A.; Barba, M. Economic impact of pome and stone fruit viruses and viroids. In Virus and Virus-Like Diseases of Pome and Stone Fruits; Hadidi, A., Barba, M., Candresse, T.H., Jelkmann, W., Eds.; American Phytopathological Society: St. Paul, MN, USA, 2011; pp. 1–7. [Google Scholar] [CrossRef]
- Anžlovar, S.; Kovac, M.; Ravnikar, M. Photosynthetic pigments in healthy and virus-infected potato plantlets (Solanum tuberosum L.) grown in vitro. Phyton 1996, 36, 221–230. [Google Scholar]
- Li, J.-W.; Wang, B.; Song, X.-M.; Wang, R.-R.; Chen, L.; Zhang, H.; Zhang, Z.-B.; Wang, Q.-C. Potato leafroll virus (PLRV) and Potato virus Y (PVY) influence vegetative growth, physiological metabolism, and microtuber production of in vitro-grown shoots of potato (Solanum tuberosum L.). Plant Cell Tissue Organ Cult. 2013, 114, 313–324. [Google Scholar] [CrossRef]
- Chen, J.; Tang, H.-H.; Li, L.; Qin, S.-J.; Wang, G.-P.; Hong, N. Effects of virus infection on plant growth, root development and phytohormone levels in in vitro-cultured pear plants. Plant Cell Tissue Organ Cult. 2017, 131, 359–368. [Google Scholar] [CrossRef]
- Li, J.-W.; Chen, H.-Y.; Li, J.; Zhang, Z.; Blystad, D.-R.; Wang, Q.-C. Growth, microtuber production and physiological metabolism in virus-free and virus-infected potato in vitro plantlets grown under NaCl-induced salt stress. Eur. J. Plant Pathol. 2018, 152, 417–432. [Google Scholar] [CrossRef]
- Wang, M.R.; Hao, X.Y.; Zhao, L.; Cui, Z.H.; Volk, G.M.; Wang, Q.C. Virus infection reduces shoot proliferation of in vitro stock cultures and ability of cryopreserved shoot tips to regenerate into normal shoots in ‘Gala’ apple (Malus × domestica). Cryobiology 2018, 82, 52–58. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, M.R.; Li, J.W.; Cui, Z.H.; Volk, G.M.; Wang, Q.C. Cryobiotechnology: A double-edged sword for obligate plant pathogens. Plant Dis. 2019, 103, 1058–1067. [Google Scholar] [CrossRef]
- Wang, M.R.; Bi, W.-L.; Bettoni, J.C.; Zhang, D.; Volk, G.M.; Wang, Q.-C. Shoot tip cryotherapy for plant pathogen eradication. Plant Pathol. 2022, 71, 1241–1254. [Google Scholar] [CrossRef]
- Panis, B. Cryopreservation of Musa germplasm. In Technical Guidelines of Bioversity International; Benson, E., Engelmann, F., Eds.; Bioversity International: Rome, Italy, 2009; pp. 8–9. [Google Scholar]
- Benson, E.E. Cryopreservation of phytodiversity: A critical appraisal of theory & practice. Crit. Rev. Plant. Sci. 2008, 27, 141–219. [Google Scholar] [CrossRef]
- Engelmann, F. Plant cryopreservation: Progress and prospects. In Vitro Cell. Dev. Biol.-Plant 2004, 40, 427–433. [Google Scholar] [CrossRef]
- Folgado, R.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F.; Panis, B. Changes in sugar content and proteome of potato in response to cold and dehydration stress and their implications for cryopreservation. J. Proteom. 2014, 98, 99–111. [Google Scholar] [CrossRef]
- Kyesmu, P.M.; Takagi, H.; Yashima, S. Cryopreservation of white yam (Dioscorea rotundata) shoot apices by vitrification. In Proceedings of the Annual Meeting of Japan Molecular Biology, Kumamoto, Japan, 20–12 July 1997; p. 162. [Google Scholar]
- Sherlock, G.; Block, W.; Benson, E.E. Thermal Analysis of the Plant Encapsulation-Dehydration Cryopreservation Protocol Using Silica Gel as the Desiccant. Cryo-Lett. 2005, 26, 45–54. [Google Scholar]
- Teixeira, A.S.; Faltus, M.; Zámečcník, J.; González-Benito, M.E.; Molina-García, A.D. Glass transition and heat capacity behaviors of plant vitrification solutions. Thermochim. Acta 2014, 593, 43–49. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Costa, M.D.; Gardin, J.P.P.; Kretzschmar, A.A.; Pathirana, R. Cryotherapy: A new technique to obtain grapevine plants free of viruses. Rev. Bras. Frutic. 2016, 38, e-833. [Google Scholar] [CrossRef]
- Wang, Q.; Tanne, E.; Arav, A.; Gafny, R. Cryopreservation of in vitro-grow shoot tips of grapevine by encapsulation-dehydration. Plant Cell Tissue Organ. Cult. 2000, 63, 41–46. [Google Scholar] [CrossRef]
- Carra, A.; Carimi, F.; Bettoni, J.C.; Pathirana, R. Progress and Challenges in the Application of Synthetic Seed Technology for Ex Situ Germplasm Conservation in Grapevine (Vitis spp.). In Synthetic Seeds; Faisal, A., Alatar, A.A., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 439–467. [Google Scholar] [CrossRef]
- Nishizawa, S.; Sakai, A.; Amano, Y.; Matsuzawa, T. Cryopreservation of asparagus (Asparagus officinalis L.) embryogenic suspension cells and subsequent plant regeneration by vitrification. Plant Sci. 1993, 91, 67–73. [Google Scholar] [CrossRef]
- Yamada, T.; Sakai, A.; Matsumura, T.; Higuchi, S. Cryopreservation of apical meristems of white clover (Trifolium repens L.) by vitrification. Plant Sci. 1991, 78, 81–87. [Google Scholar] [CrossRef]
- Manamela, M.T.; Mycock, D.J. Responses of the Buds of Three South African Sweet Potato (Ipomoea Batatas) Accessions to Different Cryoprotectants. Cryo-Lett. 2019, 40, 357–366. [Google Scholar]
- Sakai, A. Development of cryopreservation techniques. In Cryopresevation of Tropical Plant Germplasm, Current Progress and Application; Engelmann, F., Takagi, H., Eds.; IPGRI: Rome, Italy, 2000; pp. 1–7. [Google Scholar]
- Uchendu, E.; Muminova, M.; Gupta, S.; Reed, B.M. Antioxidant and anti-stress compounds improve regrowth of cryopreserved Rubus shoot tips. In Vitro Cell. Dev. Biol.-Plant 2010, 46, 386–393. [Google Scholar] [CrossRef]
- Ren, L.; Wang, M.R.; Wang, Q.-C. ROS-induced oxidative stress in plant cryopreservation: Occurrence and alleviation. Planta 2021, 254, 124. [Google Scholar] [CrossRef]
- Ren, L.; Zhang, D.; Jiang, X.; Gai, Y.; Wang, W.; Reed, B.M.; Shen, X. Peroxidation due to cryoprotectant step is a vital factor for cell survival in Arabidopsis cryopreservation. Plant Sci. 2013, 212, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhang, D.; Jiang, X.-N.; Gai, Y.; Wang, W.-M.; Reed, B.M.; Shen, X.-H. Transcriptomic profiling revealed the regulatory mechanism of Arabidopsis seedlings response to oxidative stress from cryopreservation. Plant Cell Rep. 2015, 34, 2161–2178. [Google Scholar] [CrossRef]
- Prudente, D.O.; Paiva, R.; Nery, F.C.; Paiva, P.D.O.; Alves, J.D.; Máximo, W.P.F.; Silva, L.C. Compatible solutes improve regrowth, ameliorate enzymatic antioxidant systems, and reduce lipid peroxidation of cryopreserved Hancornia speciosa Gomes lateral buds. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 352–362. [Google Scholar] [CrossRef]
- Vianna, M.G.; Garcia, R.O.; Mansur, E.; Engelmann, F.; Pacheco, G. Oxidative stress during the cryopreservation of Passiflora suberosa L. shoot tips using the V-Cryo-plate technique: Determination of the critical stages of the protocol. Plant Cell Tissue Organ Cult. 2019, 139, 369–379. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. ROS are good. Trend Plant Sci. 2017, 22, 11–19. [Google Scholar] [CrossRef]
- Wang, M.-R.; Bi, W.; Shukla, M.R.; Ren, L.; Hamborg, Z.; Blystad, D.-R.; Saxena, P.K.; Wang, Q.-C. Epigenetic and Genetic Integrity, Metabolic Stability, and Field Performance of Cryopreserved Plants. Plants 2021, 10, 1889. [Google Scholar] [CrossRef]
- Funnekotter, B.; Bunn, E.; Mancera, R.L. Cryo-mesh: A Simple Alternative Cryopreservation Protocol. Cryo-Lett. 2017, 38, 155–159. [Google Scholar]
- Mathew, L.; Burritt, D.J.; McLachlan, A.; Pathirana, R. Combined pre-treatments enhance antioxidant metabolism and improve survival of cryopreserved kiwifruit shoot tips. Plant Cell Tissue Organ Cult. 2019, 138, 193–205. [Google Scholar] [CrossRef]
- Bi, W.L.; Hao, X.Y.; Cui, Z.H.; Volk, G.M.; Wang, Q. Droplet-vitrification cryopreservation of in vitro-grown shoot tips of grapevine (Vitis spp.). In Vitro Cell. Dev. Biol.-Plant 2018, 54, 590–599. [Google Scholar] [CrossRef]
- Uchendu, E.E.; Keller, E.R.J. Melatonin-loaded alginate beads improve cryopreservation of yam (Dioscorea alata and D. cayenensis). Cryo-Lett. 2016, 37, 77–87. [Google Scholar]
- Wang, Z.C.; Deng, X.X. Cryopreservation of shoot-tips of citrus using vitrification: Effect of reduced form of glutathione. Cryo-Lett. 2004, 25, 43–50. [Google Scholar]
- Wang, Q.C.; Valkonen, J.P.T. Improved recovery of cryotherapy- treated shoot tips following thermotherapy of in vitro grown stock shoots of raspberry (Rubus idaeus L.). Cryo-Lett. 2009, 30, 171–182. [Google Scholar]
- Reed, B.M. Antioxidants and cryopreservation, the new normal? Acta Hortic. 2014, 1039, 41–48. [Google Scholar] [CrossRef]
- Chen, G.Q.; Ren, L.; Zhang, J.; Reed, B.M.; Zhang, D.; Shen, X.H. Cryopreservation affects ROS-induced oxidative stress and antioxidant response in Arabidopsis seedlings. Cryobiology 2015, 70, 38–47. [Google Scholar] [CrossRef]
- Chen, G.Q.; Ren, L.; Zhang, D.; Shen, X.H. Glutathione improves survival of cryopreserved embryogenic calli of Agapanthus praecox subsp. orientalis. Acta Physiol. Plant 2016, 38, 250. [Google Scholar] [CrossRef]
- Seo, J.H.; Naing, A.H.; Jeon, S.M.; Kim, C.K. Anti-freezing-protein type III strongly influences the expression of relevant genes in cryopreserved potato shoot tips. Plant Mol. Biol. 2018, 97, 347–355. [Google Scholar] [CrossRef]
- Reed, B.M.; Uchendu, E. Controlled Rate Cooling. In Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; pp. 77–92. [Google Scholar]
- Britto, D.T.; Kronzucker, H.J. NH4+ toxicity in higher plants: A critical review. J. Plant Physiol. 2002, 159, 567–584. [Google Scholar] [CrossRef]
- Liu, Y.; von Wirén, N. Ammonium as a signal for physiological and morphological responses in plants. J. Exp. Bot. 2017, 68, 2581–2592. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.; Cakmak, T.; Cooper, A.; Lager, I.; Rasmusson, A.G.; Escobar, M.A. Distinct signaling pathways and transcriptome response signatures differentiate ammonium- and nitrate-supplied plants. Plant Cell Environ. 2010, 33, 1486–1501. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.J. (Eds.) The components of plant tissue culture media I: Macro-and micro-nutrients. In Plant Propagation by Tissue Culture; Springer: Dordrecht, The Netherlands, 2008; pp. 65–113. [Google Scholar] [CrossRef]
- Feng, C.; Yin, Z.; Ma, Y.; Zhang, Z.; Chen, L.; Wang, B.; Li, B.; Huang, Y.; Wang, Q. Cryopreservation of sweetpotato (Ipomoea batatas) and its pathogen eradication by cryotherapy. Biotechnol. Adv. 2011, 29, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, F. Use of biotechnologies for the conservation of plant biodiversity. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Escobar, R.H.; Debouck, D.; Roca, W.M. Development of cassava cryopreservation. In Cryopreservation of Tropical Plant Germplasm; Engelmann, F., Takagi, H., Eds.; JIRCAS: Tsukuba, Japan; IPGRI: Rome, Italy, 2000; pp. 222–226. [Google Scholar]
- Macro-Medina, A.; Casas, J.L.; Swennen, R.; Panis, B. Cryopreservation of Thymus moroderi by Droplet Vitrification. Cryo-Lett. 2010, 31, 14–23. [Google Scholar]
- Vollmer, R.; Villagaray, R.; Egúsquiza, V.; Espirilla, J.; García, M.; Torres, A.; Rojas, E.; Panta, A.; Barkley, N.A.; Ellis, D. The Potato Cryobank at the International Potato Center (CIP): A Model for Long Term Conservation of Clonal Plant Genetic Resources Collections of the Future. Cryo-Lett. 2016, 37, 318–329. [Google Scholar]
- Edesi, J.; Kotkas, K.; Pirttilä, A.M.; Häggman, H. Does light spectral quality affect survival and regeneration of potato (Solanum tuberosum L.) shoot tips after cryopreservation? Plant Cell Tissue Organ Cult. 2014, 119, 599–607. [Google Scholar] [CrossRef]
- Kaczmarczyk, A.; Shvachko, N.; Lupysheva, Y.; Hajirezaei, M.-R.; Keller, E.R.J. Influence of alternating temperature preculture on cryopreservation results for potato shoot tips. Plant Cell Rep. 2008, 27, 1551–1558. [Google Scholar] [CrossRef]
- Criel, B.; Hausman, F.; Oufir, M.; Swennen, R.; Panis, B.; Renaut, J. Proteome and sugar analysis of abiotic stress underlying cryopreservation in potato. Commun. Agric. Appl. Biol. Sci. 2006, 71, 3–6. [Google Scholar]
- Folgado, R.; Panis, B.; Sergeant, K.; Renaut, J.; Swennen, R.; Hausman, J.-F. Differential Protein Expression in Response to Abiotic Stress in Two Potato Species: Solanum commersonii Dun and Solanum tuberosum L. Int. J. Mol. Sci. 2013, 14, 4912–4933. [Google Scholar] [CrossRef]
- Wang, M.-R.; Zhang, Z.; Zámečník, J.; Bilavčík, A.; Blystad, D.-R.; Haugslien, S.; Wang, Q.-C. Droplet-vitrification for shoot tip cryopreservation of shallot (Allium cepa var. aggregatum): Effects of PVS3 and PVS2 on shoot regrowth. Plant Cell Tissue Organ Cult. 2020, 140, 185–195. [Google Scholar] [CrossRef]
- Senula, A.; Nagel, M. Cryopreservation of Plant Shoot Tips of Potato, Mint, Garlic, and Shallot Using Plant Vitrification Solution 3. Methods Mol. Biol. 2021, 2180, 647–661. [Google Scholar] [CrossRef] [PubMed]
- Whelehan, L.M.; Funnekotter, B.; Bunn, E.; Mancera, R.L. Review: The case for studying mitochondrial function during plant cryopreservation. Plant Sci. 2022, 315, 111134. [Google Scholar] [CrossRef]
- Ruiz-Sáenz, D.R.; López-Delgado, H.A.; Hernández, D.D.A.; Trejo, C.; Mora-Herrera, M.E.; Mortera, E.U. Induction of tolerance to cryogenic protocols in Solanum tuberosum by salicylic acid is mediated by enzymatic antioxidant activity and hydrogen peroxide. J. Hortic. Sci. Biotechnol. 2022, 97, 86–95. [Google Scholar] [CrossRef]
- Grout, B.W.W.; Henshaw, G.G. Structural Observations on the Growth of Potato Shoot-tip Cultures after Thawing from Liquid Nitrogen. Ann. Bot. 1980, 46, 243–248. [Google Scholar] [CrossRef]
- Golmirzaie, A.M.; Panta, A.; Delgado, C. Structural observations on potato shoot-tips after thawing from liquid nitrogen. In Cryopreservation of Tropical Plant Germplasm. Current Research Progress and Application; Engelmann, F., Takagi, H., Eds.; Japan International Research Center for Agricultural Sciences: Tsukuba, Japan, 2000; pp. 388–389. [Google Scholar]
- Kaczmarczyk, A.; Rutten, T.; Melzer, M.; Keller, E.R.J. Ultrastructural Changes Associated with Cryopreservation of Potato (Solanum tuberosum L.). Cryo-Lett. 2008, 29, 145–156. [Google Scholar]
- Wang, B.; Li, J.-W.; Zhang, Z.-B.; Wang, R.-R.; Ma, Y.-L.; Blystad, D.-R.; Keller, E.R.J.; Wang, Q.-C. Three vitrification-based cryopreservation procedures cause different cryo-injuries to potato shoot tips while all maintain genetic integrity in regenerants. J. Biotech. 2014, 184, 47–55. [Google Scholar] [CrossRef]



| Plant Species (No. of Tested Genotypes) | Age of Culture, Pretreatment of Donor Plants and Excised Explant | Encapsulation Method 1 | Preculture | Duration of Dehydration (Residual Water Content) 2 | Shoot Tip Regrowth in Percentage | Ref. |
|---|---|---|---|---|---|---|
| Cassava [Manihot esculenta (3)] | Apices (not specified) | 3% sodium alginate to form beads, the calcium chloride concentration was not specified | 0.75 M suc for 1 day or 0.5 M for 3 days | 6 h | <60 (survival) | [97] |
| Potato [Solanum phureja, S. Phureja × chacoense, S. tuberosum (5)] | Shoot tips (0.5 mm) excised from in vitro shoots cultured for 2 weeks | 3% sodium alginate to form beads of 3–4 mm in diameter | 0.5 M or 0.75 M suc for 2 days | 4.5 h (0.2–0.22 g per g of dry weight) | 50–78 | [93] |
| S. tuberosum, S. chacoence, S. phureja, S. bulbocastanum, S. pinnatisectum (16) | Shoot tips (1.5–2.0 mm) excised from in vitro shoots cultured for 2 weeks | 2% sodium alginate and 0.4 M suc to attach shoot tips to aluminum plates; osmoprotection with 2 M glycerol and 0.8 M suc for 30 min | 0.3 M suc overnight | 2 h | 80–100 | [56] |
| Sweet potato [Ipomoea batatas (1)] | Shoot tips (0.5–1.0 mm) excised from cultures that are 4 to 8 weeks old | 3% sodium alginate to form beads of 4–5 mm in diameter | Stepwise daily increased of suc levels 0.25, 0.5 and 0.75 M, | Till water content of 18.1% | 67 | [99] |
| Taro [Colocasia esculenta (1)] | Shoot tips (0.8–1.0 mm) excised from 8-week-old cultures preconditioned with 0.29 M suc | 2% sodium alginate and 0.4 M suc to form beads of 4 mm in diameter | 0.75 M suc for 2 days | 6 h in laminar airflow and 12 h over silica gel (18–19%) | 65 | [102] |
| Ulluco [Ullucus tuberosus (11)] | Shoot tips (1.0–1.5 mm) excised from cultures that are 3 to 4 weeks old were cold-hardened at 5 °C, followed by preculture with sucrose at 0.3 M for 16 h | With 2% sodium alginate and 0.4 M suc to attach shoot tips to aluminum plates | 2.0 M gly and 1.0 M suc for 90 min | 45 min | 73–97 | [103] |
| Yam [Dioscorea alata, D. wallichii, D. bulbifera, D. floribunda (4)] | Apical shoot tips (0.6–1.2 mm) excised from cultures that are 6 to 6 weeks old | 3% sodium alginate to form beads | 0.75 M suc for 3 days | 4 h | 21% for D. alata, 37% for D. wallichii and 0% for others | [100] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, A.-L.; Wang, M.-R.; Li, Z.; Panis, B.; Bettoni, J.C.; Vollmer, R.; Xu, L.; Wang, Q.-C. Overcoming Challenges for Shoot Tip Cryopreservation of Root and Tuber Crops. Agronomy 2023, 13, 219. https://doi.org/10.3390/agronomy13010219
Zhang A-L, Wang M-R, Li Z, Panis B, Bettoni JC, Vollmer R, Xu L, Wang Q-C. Overcoming Challenges for Shoot Tip Cryopreservation of Root and Tuber Crops. Agronomy. 2023; 13(1):219. https://doi.org/10.3390/agronomy13010219
Chicago/Turabian StyleZhang, A-Ling, Min-Rui Wang, Zhiying Li, Bart Panis, Jean Carlos Bettoni, Rainer Vollmer, Li Xu, and Qiao-Chun Wang. 2023. "Overcoming Challenges for Shoot Tip Cryopreservation of Root and Tuber Crops" Agronomy 13, no. 1: 219. https://doi.org/10.3390/agronomy13010219
APA StyleZhang, A.-L., Wang, M.-R., Li, Z., Panis, B., Bettoni, J. C., Vollmer, R., Xu, L., & Wang, Q.-C. (2023). Overcoming Challenges for Shoot Tip Cryopreservation of Root and Tuber Crops. Agronomy, 13(1), 219. https://doi.org/10.3390/agronomy13010219