



Cover Crops Increase N and P Cycling and Rice Productivity in Temperate Cropping Systems

 ,
, 
 ,
,  , ,
, ,
Abstract
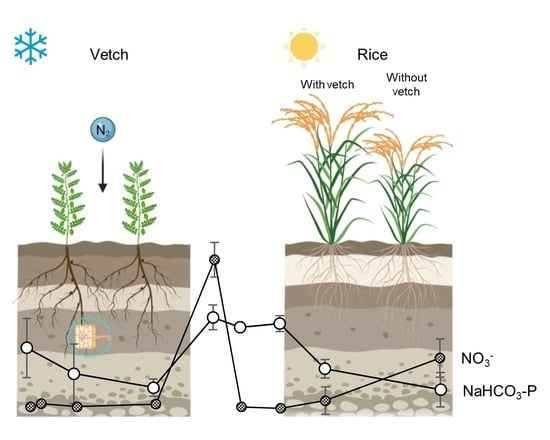
:
1. Introduction
2. Materials and Methods
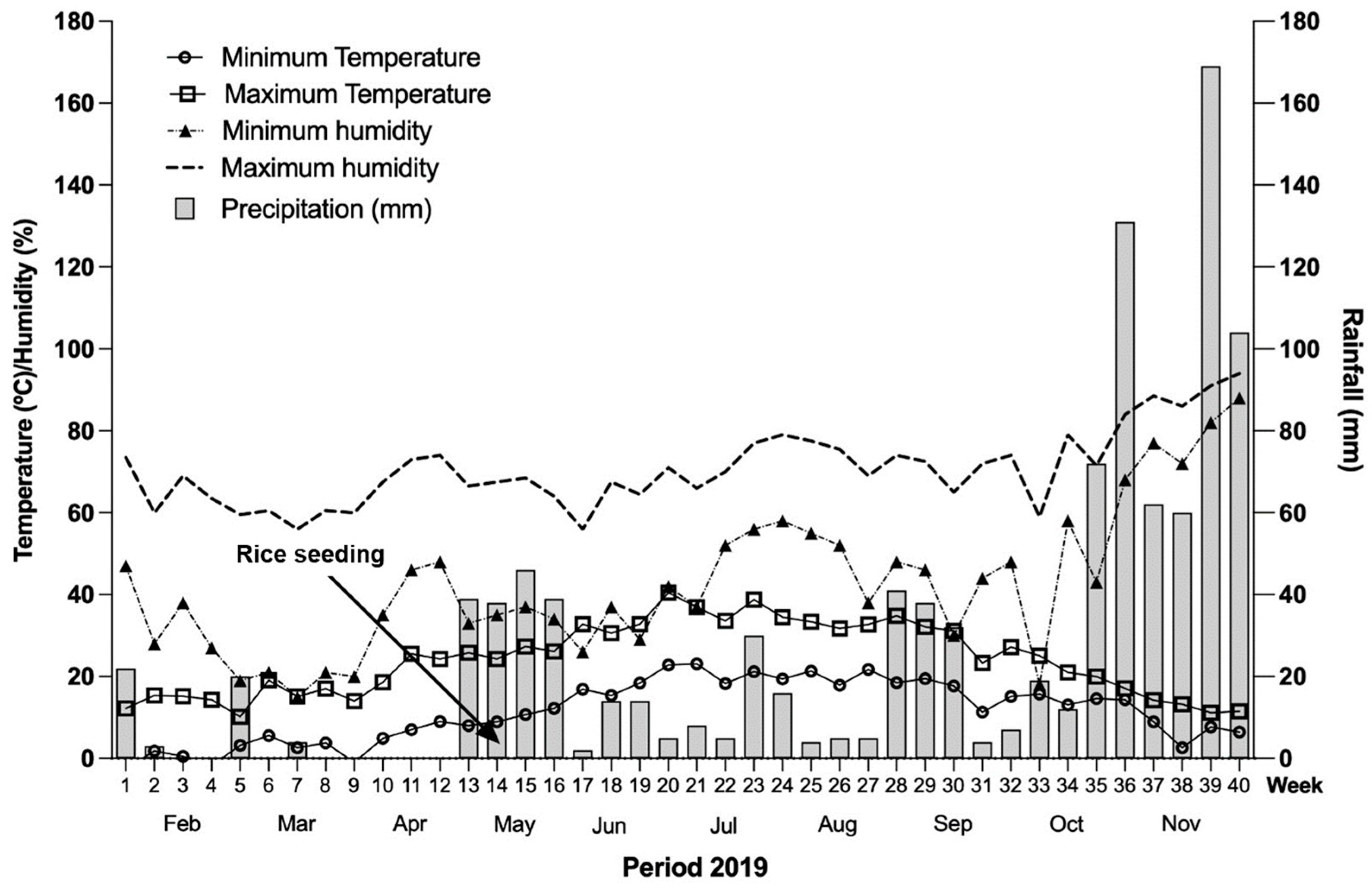
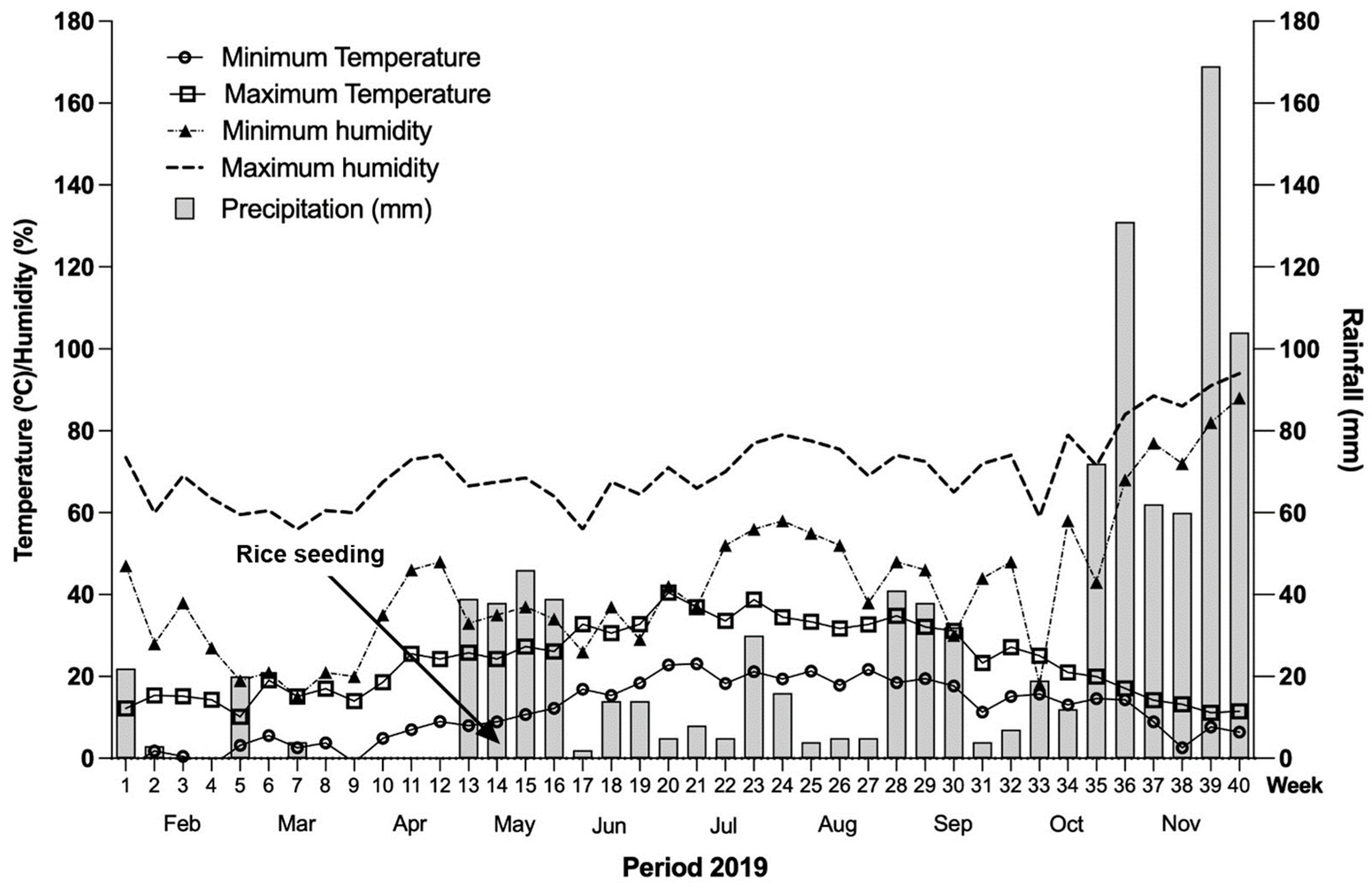
2.1. Study Site
2.2. Experimental Design
2.3. Crop Yields, Yield Components and Nutrient Contents
2.4. Soil and Vetch Sampling and Analyses
2.5. Statical Analysis
3. Results
3.1. Rice Yields and the Yield Components
3.2. Rice Nutrient Uptake and N Apparent Recovery
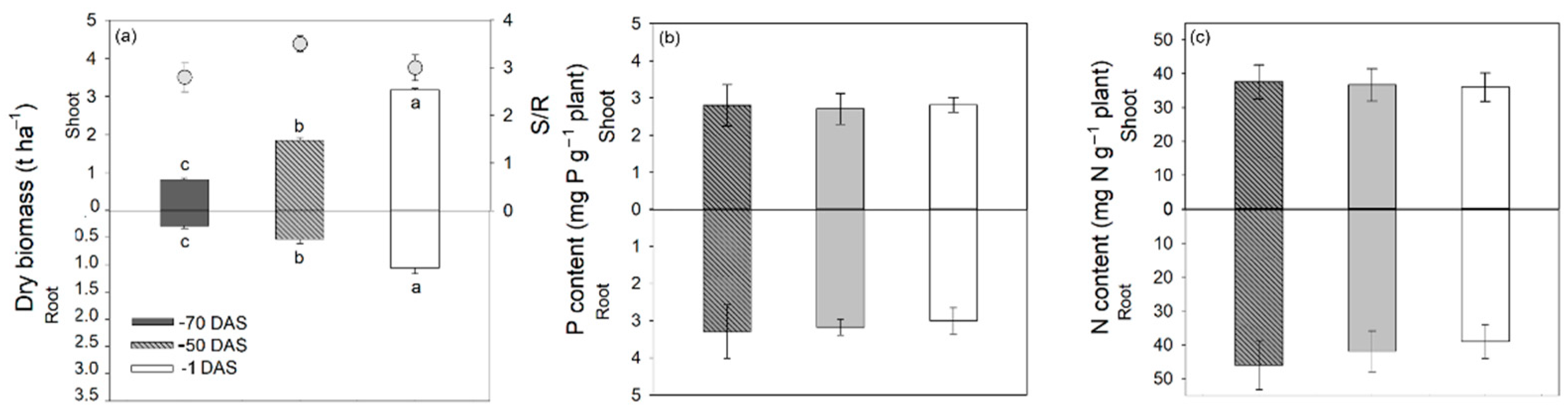
3.3. Vetch Biomass and Nutrient Uptake
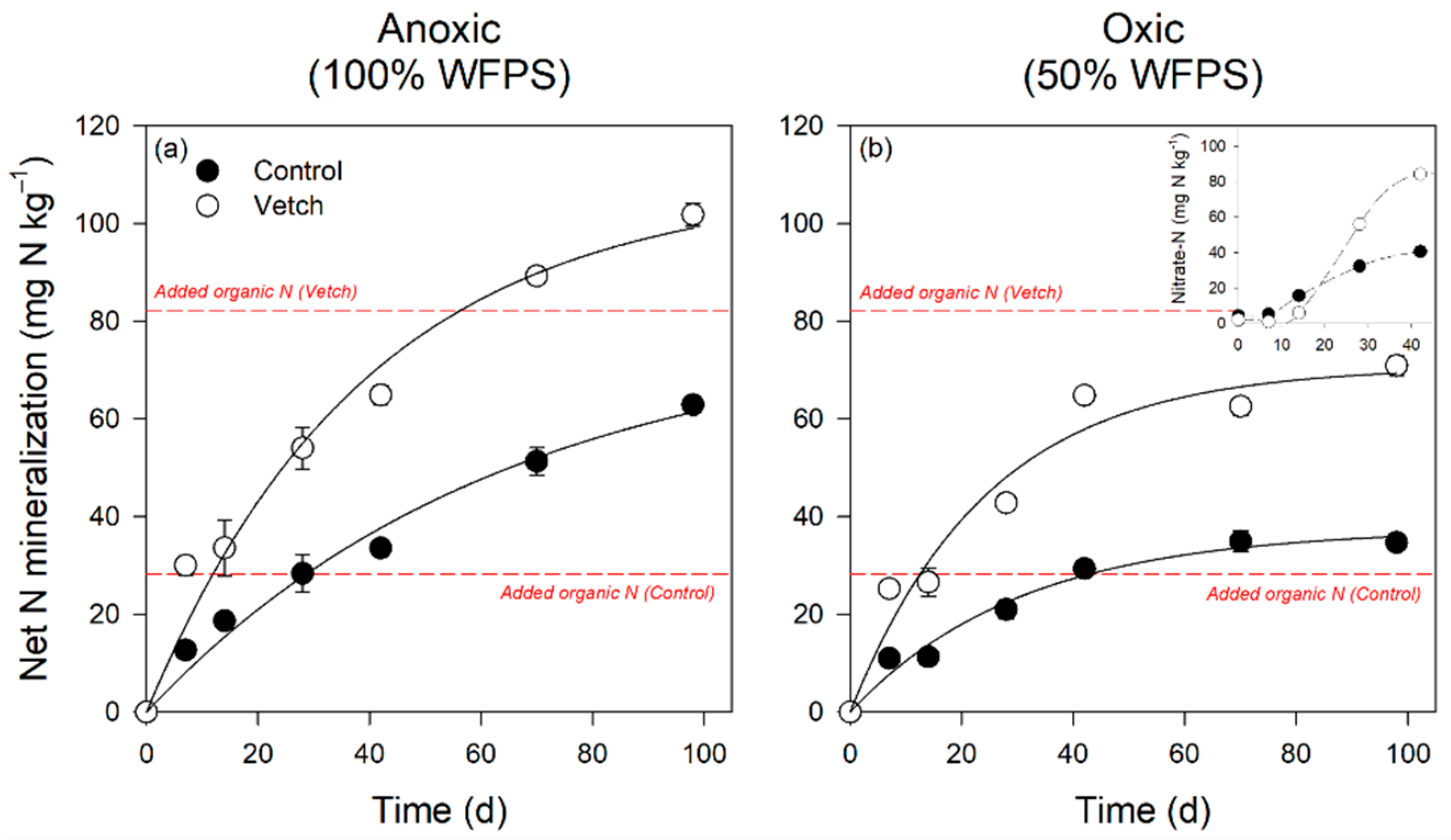
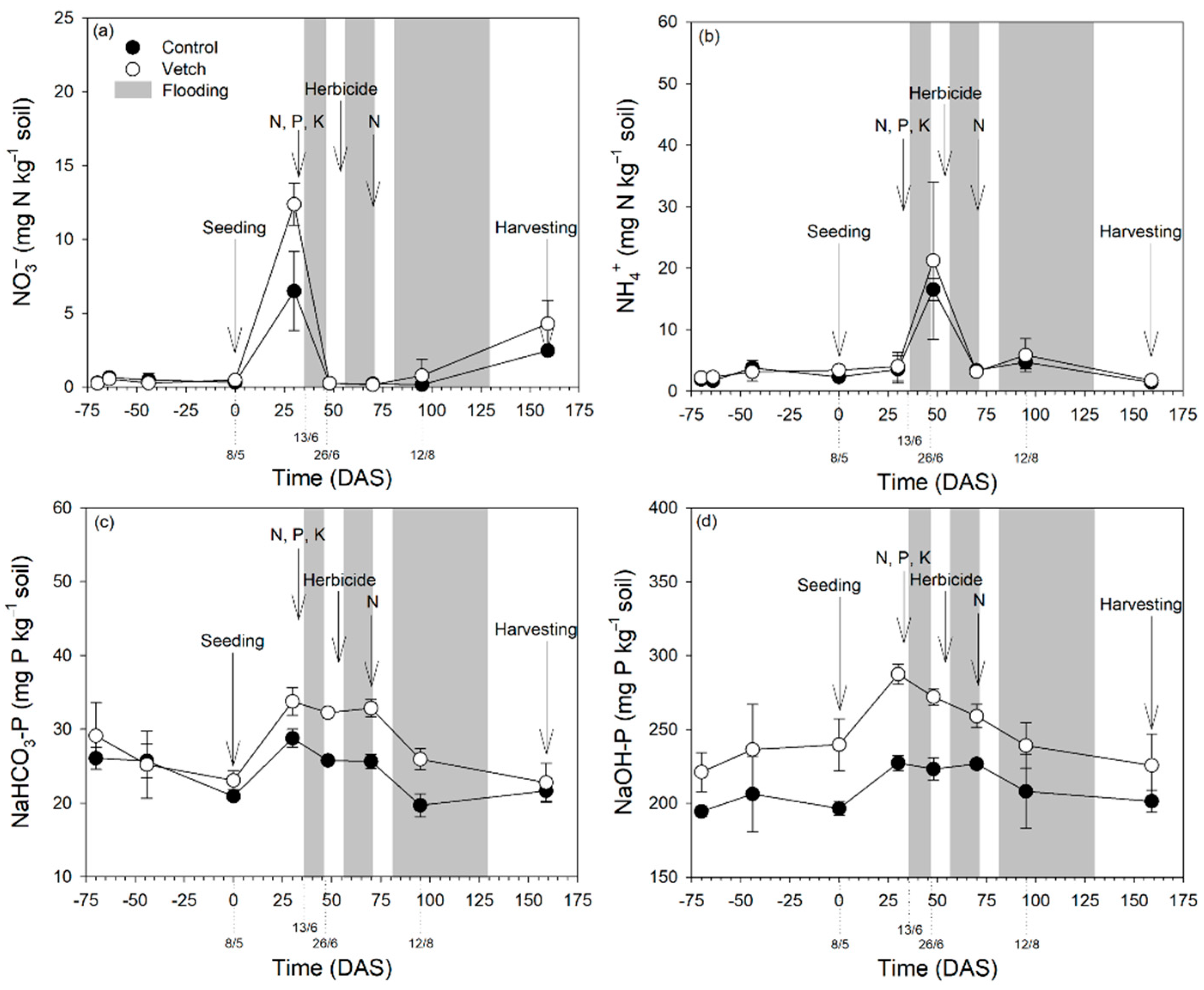
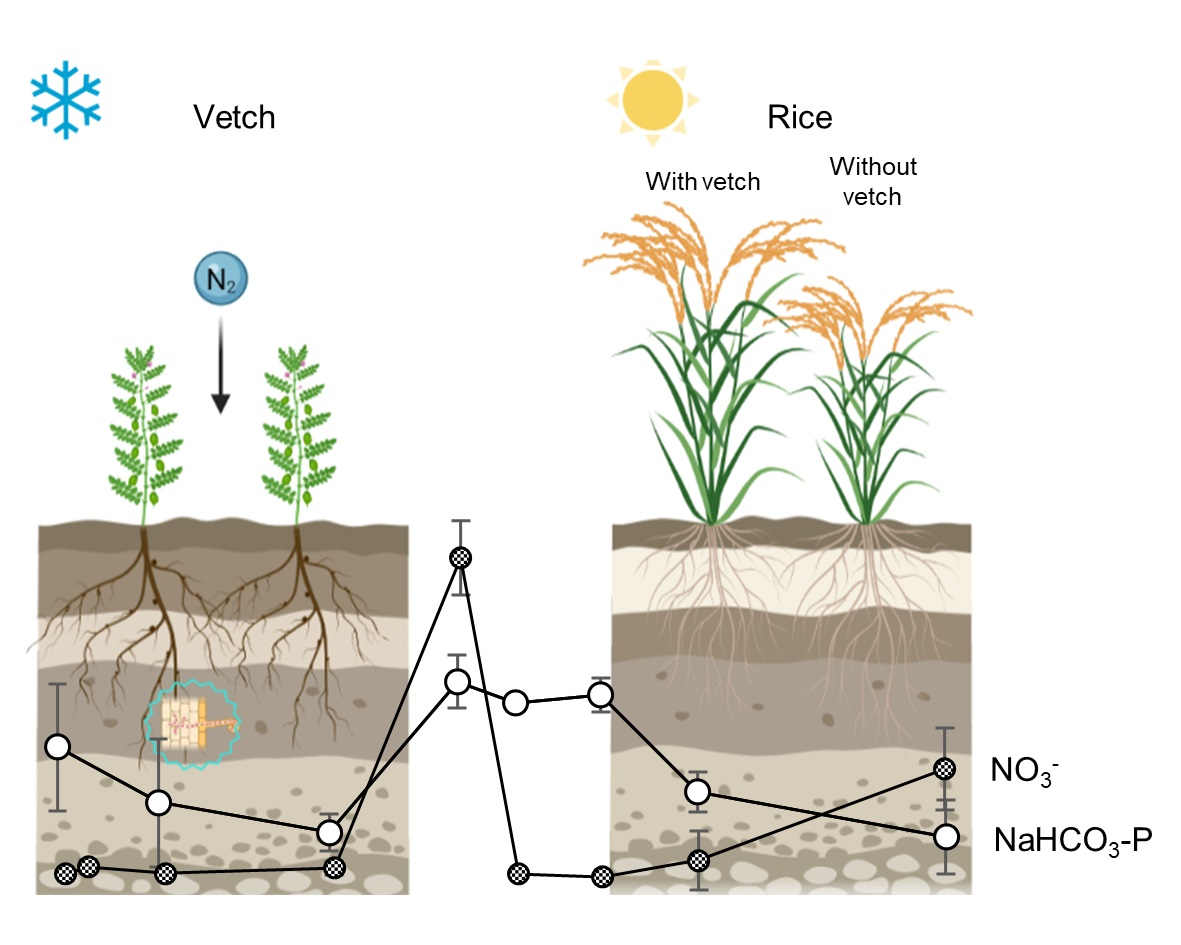
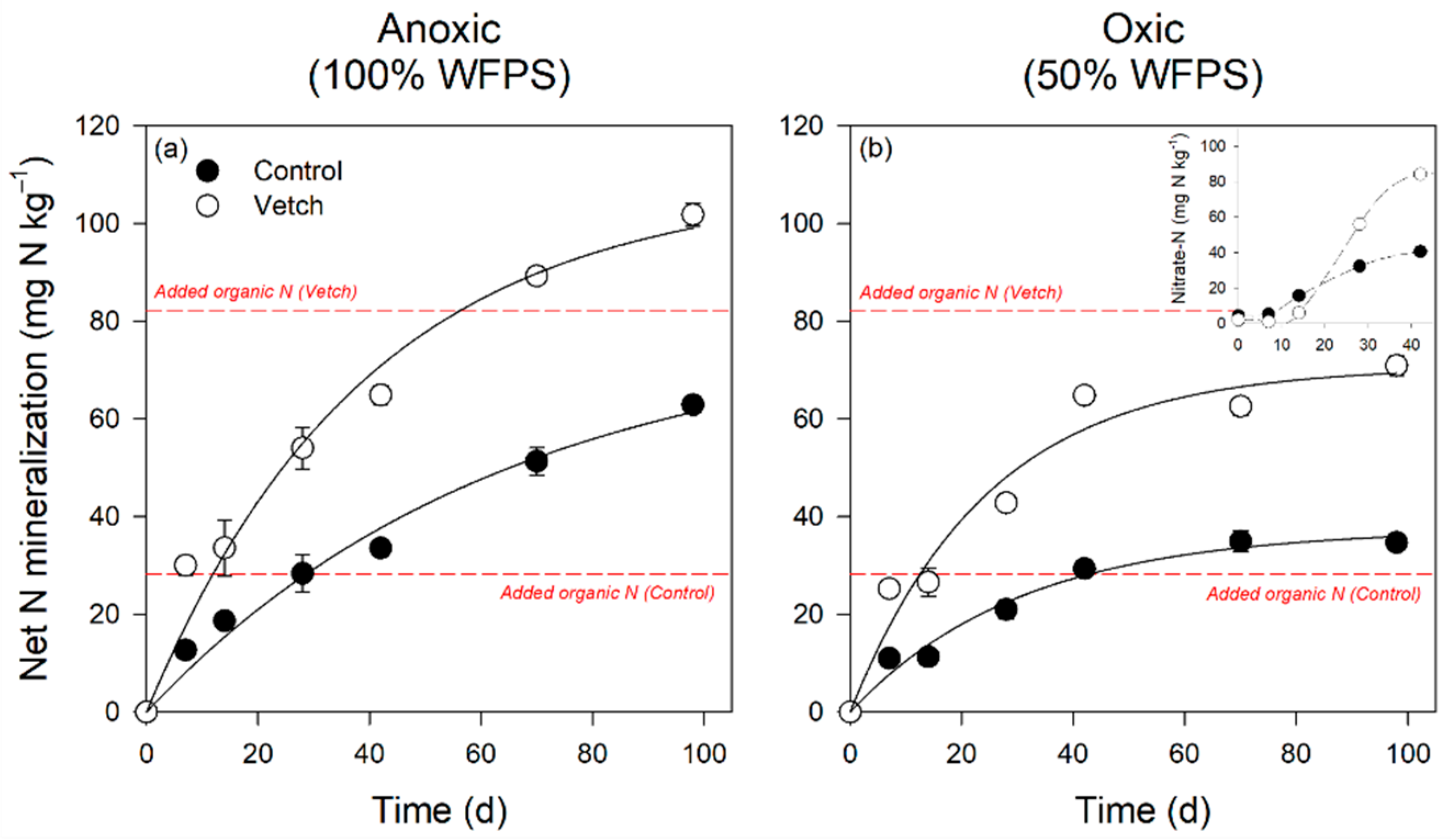
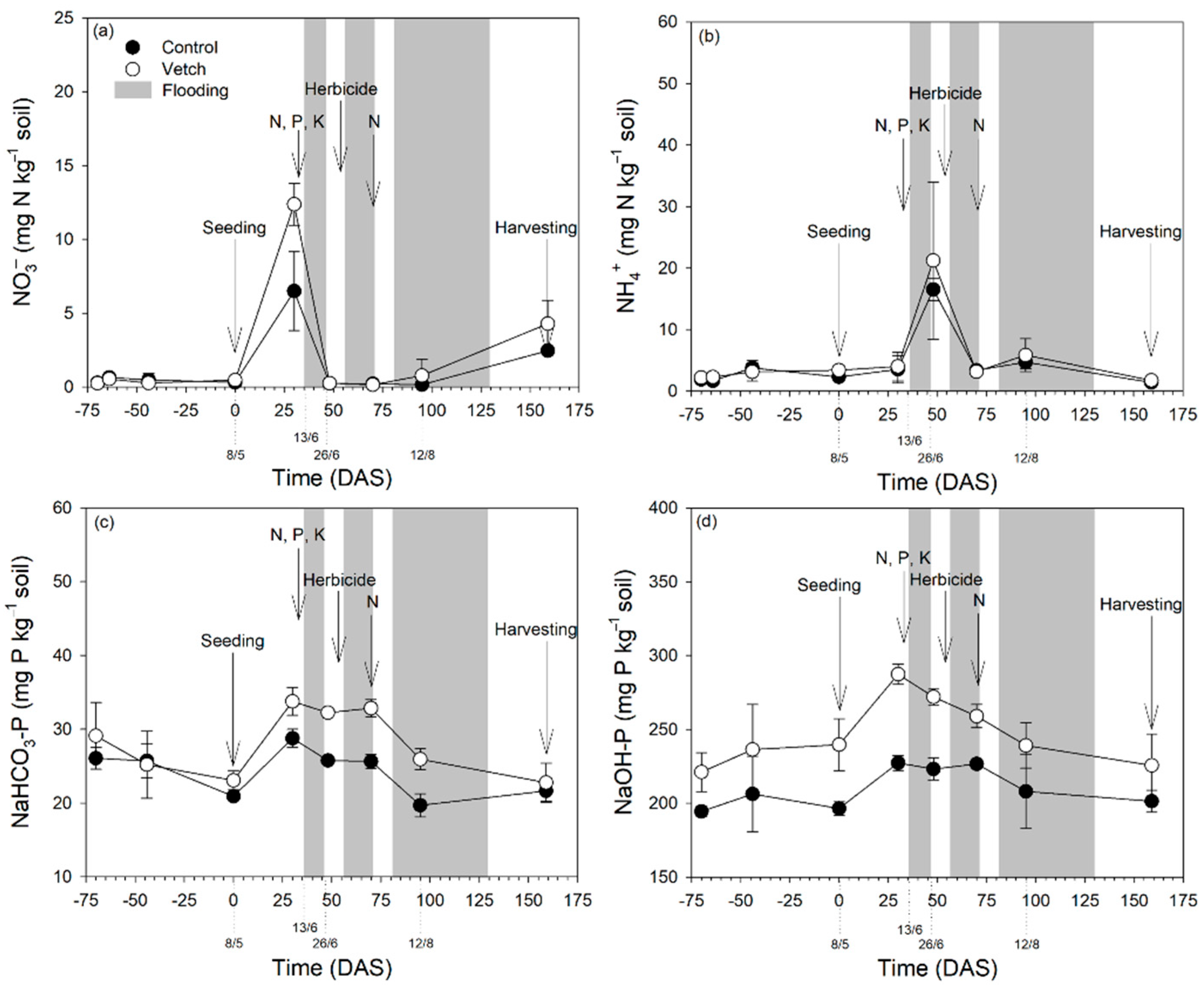
3.4. Soil Nitrogen and Phosphorus Forms and Temporal Dynamics
4. Discussion
4.1. Hairy Vetch Increases the Rice Yield Performance as a Function of N Fertilizer Levels
4.2. Hairy Vetch Increases Soil N and P Availability
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations. FAOSTAT. FAO: Rome, Italy, 2019. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 30 January 2020).
- Cassman, K.G.; Peng, S.; Olk, D.C.; Ladha, J.K.; Reichardt, W.; Dobermann, A.; Singh, U. Opportunities for increased nitrogen-use efficiency from improved resource management in irrigated rice systems. Field Crops Res. 1998, 56, 7–39. [Google Scholar] [CrossRef]
- Eagle, A.J.; Bird, J.A.; Horwáth, W.R.; Linquist, B.A.; Brouder, S.M.; Hill, J.E.; van Kessel, C. Rice yield and nitrogen utilization efficiency under alternative straw management practices. Agron. J. 2000, 92, 1096–1103. [Google Scholar] [CrossRef]
- Montgomery, D.R. Soil erosion and agricultural sustainability. Proc. Natl. Acad. Sci. USA 2007, 104, 13268–13272. [Google Scholar] [CrossRef]
- Weerasekara, C.S.; Udawatta, R.P.; Gantzer, C.J.; Kremer, R.J.; Jose, S.; Veum, K.S. Effects of Cover Crops on Soil Quality: Selected Chemical and Biological Parameters. Commun. Soil Sci. Plant Anal. 2017, 48, 2074–2082. [Google Scholar] [CrossRef]
- Amundson, R.; Berhe, A.A.; Hopmans, J.W.; Olson, C.; Sztein, A.E.; Sparks, D.L. Soil and human security in the 21st century. Science 2015, 348, 1261071. [Google Scholar] [CrossRef]
- Delgado, J.A.; Gantzer, C.J. The 4Rs for cover crops and other advances in cover crop management for environmental quality. J. Soil Water Conserv. 2015, 70, 142A–145A. [Google Scholar] [CrossRef]
- Prell, J.; Poole, P. Metabolic changes of rhizobia in legume nodules. Trends Microbiol. 2006, 14, 161–168. [Google Scholar] [CrossRef]
- Parr, M.; Grossman, J.M.; Reberg-Horton, S.C.; Brinton, C.; Crozier, C. Nitrogen delivery from legume cover crops in no-till organic corn production. Agron. J. 2011, 103, 1578–1590. [Google Scholar] [CrossRef]
- Cassman, K.G.; Dobermann, A.; Walters, D.T. Agroecosystems, nitrogen-use efficiency, and nitrogen management. Ambio 2002, 31, 132–140. [Google Scholar] [CrossRef]
- Miniotti, E.F.; Romani, M.; Said-Pullicino, D.; Facchi, A.; Bertora, C.; Peyron, M.; Sacco, D.; Bischetti, G.B.; Lerda, C.; Tenni, D.; et al. Agro-environmental sustainability of different water management practices in temperate rice agro-ecosystems. Agric. Ecosys. Environ. 2016, 222, 235–248. [Google Scholar] [CrossRef]
- Wyland, L.J.; Jackson, L.E.; Chaney, W.E.; Klonsky, K.; Koike, S.T.; Kimple, B. Winter cover crops in a vegetable cropping system: Impacts on nitrate leaching, soil water, crop yield, pests and management costs. Agric. Ecosys. Environ. 1996, 59, 1–17. [Google Scholar] [CrossRef]
- Brandsæter, L.O.; Heggen, H.; Riley, H.; Stubhaug, E.; Henriksen, T.M. Winter survival, biomass accumulation and N mineralization of winter annual and biennial legumes sown at various times of year in Northern Temperate Regions. Eur. J. Agron. 2008, 28, 437–448. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, X.; Liao, Y.; Lu, Y.; Nie, J.; Cao, W. Co-incorporation of rice straw and green manure benefits rice yield and nutrient uptake. Crop Sci. 2019, 59, 749–759. [Google Scholar] [CrossRef]
- Devêvre, O.C.; Horwáth, W.R. Stabilization of fertilizer nitrogen-15 into humic substances in aerobic vs. waterlogged soil following straw incorporation. Soil Sci. Soc. Am. J. 2001, 65, 499–510. [Google Scholar] [CrossRef]
- Cucu, M.A.; Said-Pullicino, D.; Maurino, V.; Bonifacio, E.; Romani, M.; Celi, L. Influence of redox conditions and rice straw incorporation on nitrogen availability in fertilized paddy soils. Biol. Fertil. Soils 2014, 50, 755–764. [Google Scholar] [CrossRef]
- Said-Pullicino, D.; Cucu, M.A.; Sodano, M.; Birk, J.J.; Glaser, B.; Celi, L. Nitrogen immobilization in paddy soils as affected by redox conditions and rice straw incorporation. Geoderma 2014, 228–229, 44–53. [Google Scholar] [CrossRef]
- Alamgir, M.D.; McNeill, A.; Tang, C.; Marschner, P. Changes in soil P pools during legume residue decomposition. Soil Biol. Biochem. 2012, 49, 70–77. [Google Scholar] [CrossRef]
- Damon, P.M.; Bowden, B.; Rose, T.; Rengel, Z. Crop residue contributions to phosphorus pools in agricultural soils: A review. Soil Biol. Biochem. 2014, 74, 127–137. [Google Scholar] [CrossRef]
- Oehl, F.; Oberson, A.; Probst, M.; Fliessbach, A.; Roth, H.R.; Frossard, E. Kinetics of microbial phosphorus uptake in cultivated soils. Biol. Fertil. Soils 2001, 34, 31–41. [Google Scholar] [CrossRef]
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Li, S.; Lei, X.; Qin, L.; Sun, X.; Wang, L.; Zhao, S.; Wang, M.; Chen, S. Fe(III) reduction due to low pe + pH contributes to reducing Cd transfer within a soil-rice system. J. Hazard. Mater. 2021, 415, 125668. [Google Scholar] [CrossRef] [PubMed]
- Oberson, A.; Frossard, E. Phosphorus Management for Organic Agriculture. In Phosphorus: Agriculture and the Environment; Sims, T., Sharpley, A.N., Daniel, T.C., Eds.; American Society of Agronomy, Inc.; Crop Science Society of America, Inc.; Soil Science Society of America, Inc.: Madison, WI, USA, 2005; Volume 46. [Google Scholar]
- Celi, L.; Prati, M.; Magnacca, G.; Santoro, V.; Martin, M. Role of crystalline iron oxides on stabilization of inositol phosphates in soil. Geoderma 2020, 374, 114442. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, X.; Xu, J.; Cao, K.; Wang, J.; Xu, C.; Cao, W. Green manure incorporation with reductions in chemical fertilizer inputs improves rice yield and soil organic matter accumulation. J. Soils Sediments 2020, 20, 2784–2793. [Google Scholar] [CrossRef]
- Gao, X.; He, Y.; Zhang, T.; An, Y.; Sun, C.; Xu, H.; Wang, X. Alfalfa green manure amendment improved P use efficiency and reduced P losses from paddy fields. Nutr. Cycl. Agroecosyst. 2022, 123, 35–47. [Google Scholar] [CrossRef]
- Lizcano Toledo, R. Strategies to Improve Phosphorus Availability and Reduce Environmental Impact in Rice Agrosystems. Ph.D. Thesis, Universidad de Granada, Granada, Spain, 2022. [Google Scholar]
- Ohno, T.; Zibilske, L.M. Determination of low concentration of phosphorus in soil extracts using malachite green. Soil Sci. Soc. Am. J. 1991, 55, 892–895. [Google Scholar] [CrossRef]
- Olsen, S.R.; Cole, C.V.; Watanabe, F.S.; Dean, L.A. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; USDA Circular No. 939; US Department of Agriculture: Washington, DC, USA, 1954. [Google Scholar]
- Compton, J.E.; Cole, D.W. Phosphorus cycling and soil P fractions in Douglas-fir and red alder stands. For. Ecol. Manag. 1998, 110, 101–112. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Sahrawat, K.L.; Ponnamperuma, F.N. Measurement of Exchangeable NH4+ in Tropical Rice Soils. Soil Sci. Soc. Am. J. 1978, 42, 282–283. [Google Scholar] [CrossRef]
- Waring, S.; Bremner, J. Ammonium production in soil under waterlogged conditions as an index of nitrogen availability. Nature 1964, 201, 951–952. [Google Scholar] [CrossRef]
- Haque, M.M.; Kim, S.Y.; Pramanik, P.; Kim, G.Y.; Kim, P.J. Optimum application level of winter cover crop biomass as green manure under considering methane emission and rice productivity in paddy soil. Biol. Fertil. Soils 2013, 49, 487–493. [Google Scholar] [CrossRef]
- Zhu, B.; Yi, L.X.; Hu, Y.G.; Zeng, Z.H.; Lin, C.W.; Tang, H.M.; Yang, G.L.; Xiao, X.P. Nitrogen release from incorporated 15N-labelled chinese milk vetch (Astragalus Sinicus L.) residue and its dynamics in a double rice cropping system. Plant Soil 2014, 374, 331–344. [Google Scholar] [CrossRef]
- Chen, J.R.; Qin, W.J.; Chen, X.F.; Cao, W.D.; Qian, G.M.; Jia, L.I.U.; Xu, C.X. Application of Chinese milk vetch affects rice yield and soil productivity in a subtropical double-rice cropping system. J. Integr. Agric. 2020, 19, 2116–2126. [Google Scholar] [CrossRef]
- Zhou, G.; Cao, W.; Bai, J.; Xu, C.; Zeng, N.; Gao, S.; Rees, R.M.; Dou, F. Co-incorporation of rice straw and leguminous green manure can increase soil available nitrogen (N) and reduce carbon and N losses: An incubation study. Pedosphere 2020, 30, 661–670. [Google Scholar] [CrossRef]
- Kaewpradit, W.; Toomsan, B.; Cadisch, G.; Vityakon, P.; Limpinuntana, V.; Saenjan, P.; Jogloy, S.; Patanothai, A. Mixing groundnut residues and rice straw to improve rice yield and N use efficiency. Field Crops Res. 2009, 110, 130–138. [Google Scholar] [CrossRef]
- Nie, J.; Yi, L.; Xu, H.; Liu, Z.; Zeng, Z.; Dijkstra, P.; Koch, G.W.; Hungate, B.A.; Zhu, B. Leguminous Cover Crop Astragalus sinicus Enhances Grain Yields and Nitrogen Use Efficiency through Increased Tillering in an Intensive Double-Cropping Rice System in Southern China. Agronomy 2019, 9, 554. [Google Scholar] [CrossRef]
- Zhou, G.; Gao, S.; Chang, D.; Rees, R.M.; Cao, W. Using milk vetch (Astragalus sinicus L.) to promote rice straw decomposition by regulating enzyme activity and bacterial community. Bioresour. Technol. 2021, 319, 124215. [Google Scholar] [CrossRef]
- Zhou, G.; Cao, W.; Bai, J.; Xu, C.; Zeng, N.; Gao, S.; Rees, R.M. Non-additive responses of soil C and N to rice straw and hairy vetch (Vicia villosa Roth L.) mixtures in a paddy soil. Plant Soil 2019, 436, 229–244. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.; Kong, X.; Hou, D.; Gu, J.; Liu, L.; Wang, Z.; Yang, J. Progressive integrative crop managements increase grain yield, nitrogen use efficiency and irrigation water productivity in rice. Field Crop Res. 2018, 215, 1–11. [Google Scholar] [CrossRef]
- Hirzel, J.; Rodríguez, F. Response of rice to increasing nitrogen rates in five soils with different mineralizable nitrogen levels. J. Plant Nutr. 2017, 40, 382–390. [Google Scholar] [CrossRef]
- Campiglia, E.; Caporali, F.; Radicetti, E.; Mancinelli, R. Hairy vetch (Vicia villosa Roth.) cover crop residue management for improving weed control and yield in no-tillage tomato (Lycopersicon esculentum Mill.) production. Eur. J. Agron. 2010, 33, 94–102. [Google Scholar] [CrossRef]
- Anugroho, F.; Kitou, M.; Nagumo, F.; Kinjo, K.; Jayasinghe, G.Y. Potential growth of hairy vetch as a winter legume cover crops in subtropical soil conditions. Soil Sci. Plant Nutr. 2010, 56, 254–262. [Google Scholar] [CrossRef]
- Liang, X.; Jin, Y.; Zhao, Y.; Wang, Z.; Yin, R.; Tian, G. Release and migration of colloidal phosphorus from a typical agricultural field under long-term phosphorus fertilization in southeastern China. J. Soils Sediments 2016, 16, 842–853. [Google Scholar] [CrossRef]
- Reddy, K.R.; DeLaune, R.D. Biogeochemistry of Wetlands: Science and Applications; CRC: Boca Raton, FL, USA, 2008. [Google Scholar]
- Akter, M.; Deroo, H.; De Grave, E.; Van Alboom, T.; Abdul Kader, M.; Pierreux, S.; Begum, M.A.; Boeckx, P.; Sleutel, S. Link between paddy soil mineral nitrogen release and iron and manganese reduction examined in a rice pot growth experiment. Geoderma 2018, 326, 9–21. [Google Scholar] [CrossRef]
- Choi, B.; Lim, J.E.; Sung, J.K.; Jeon, W.T.; Lee, S.S.; Oh, S.E.; Ok, Y.S. Effect of rapeseed green manure amendment on soil properties and rice productivity. Commun. Soil Sci. Plant Anal. 2014, 45, 751–764. [Google Scholar] [CrossRef]
- Lowry, C.J.; Robertson, G.P.; Brainard, D.C. Strip-tillage decreases soil nitrogen availability and increases the potential for N losses in a cover cropped organic system. Agric. Ecosys. Environ. 2021, 319, 107524. [Google Scholar] [CrossRef]
- Kuo, S.; Huang, B.; Bembenek, R. Effects of long-term phosphorus fertilization and winter cover cropping on soil phosphorus transformations in less weathered soil. Biol. Fertil. Soils 2005, 41, 116–123. [Google Scholar] [CrossRef]
- Wang, X.C.; Ma, X.T.; Han, M.; Cao, W.D.; Zhang, H.L.; Bai, J.S.; Zeng, N.H.; Gao, S.J.; Zhou, G.P.; Wang, Y.Q. Screening of rhizobia of common vetch (Vicia sativa) in Qinghai, and assessment of symbiont salt tolerance. Acta Pratacult. Sin. 2016, 25, 145–153. [Google Scholar]
- Tarui, A.; Matsumura, A.; Asakura, S.; Yamawaki, K.; Hattori, R.; Daimon, H. Evaluation of mixed cropping of oat and hairy vetch as green manure for succeeding corn production. Plant Prod. Sci. 2013, 16, 383–392. [Google Scholar] [CrossRef]
- Solangi, F.; Bai, J.; Gao, S.; Yang, L.; Zhou, G.; Cao, W. Improved Accumulation Capabilities of Phosphorus and Potassium in Green Manures and Its Relationship with Soil Properties and Enzymatic Activities. Agronomy 2019, 9, 708. [Google Scholar] [CrossRef]
- Xiao, Y.; Tang, J.L.; Wang, M.K.; Zhai, L.; Zhang, X. Impacts of soil properties on phosphorus adsorption and fractions in purple soils. J. Mt. Sci. 2017, 14, 2420–2431. [Google Scholar] [CrossRef]
- Kalbitz, K.; Kaiser, K.; Fiedler, S.; Kölbl, A.; Amelung, W.; Bräuer, T.; Cao, Z.; Don, A.; Grootes, P.; Jahn, R.; et al. The carbon count of 2000 years of rice cultivation. Glob. Change Biol. 2013, 19, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Said-Pullicino, D.; Miniotti, E.F.; Sodano, M.; Bertora, C.; Lerda, C.; Chiaradia, E.A.; Romani, M.; Cesari de Maria, S.; Sacco, D.; Celi, L. Linking dissolved organic carbon cycling to organic carbon fluxes in rice paddies under different water management practices. Plant Soil 2016, 401, 273–290. [Google Scholar] [CrossRef]
- Said-Pullicino, D.; Giannetta, B.; Demeglio, B.; Missong, A.; Gottselig, N.; Romani, M.; Bol, R.; Klumpp, E.; Celi, L. Redox-driven changes in water-dispersible colloids and their role in carbon cycling in hydromorphic soils. Geoderma 2021, 385, 114894. [Google Scholar] [CrossRef]
- Heiberg, L.; Pedersen, T.V.; Jensen, H.S.; Kjaergaard, C.; Hansen, H.C. A comparative study of phosphate sorption in lowland soils under oxic and anoxic conditions. J. Environ. Qual. 2010, 39, 734–743. [Google Scholar] [CrossRef]
- Hinkle, M.A.; Wang, Z.; Giammar, D.E.; Catalano, J.G. Interaction of Fe (II) with phosphate and sulfate on iron oxide surfaces. Geochim. Cosmochim. Acta 2015, 158, 130–146. [Google Scholar] [CrossRef]
- Celi, L.; Barberis, E. Abiotic stabilization of organic phosphorus in the environment. In Organic Phosphorus in the Environment; Turner, B.L., Frossard, E., Baldwin, D., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 113–132. [Google Scholar]
- Kour, D.; Rana, K.L.; Kaur, L.; Yadav, N.; Yadav, A.N.; Kumar, M.; Kumar, V.; Dhaliwal, H.S.; Saxena, A.K. Biodiversity, current developments and potential biotechnological applications of phosphorus-solubilizing and -mobilizing microbes: A review. Pedosphere 2021, 31, 43–75. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N Fertilization Level (kg N ha−1) | Cover Crop Management | Grain Yield (t ha−1) | Total Biomass (t ha−1) | Straw Biomass (t ha−1) | Harvest Index (%) |
|---|---|---|---|---|---|
| Average | +Vetch | 8.5 a | 17.2 a | 8.7 a | 50 a |
| −Vetch | 7.6 b | 15.2 b | 7.6 b | 50 a | |
| 0 | 6.5 c | 12.8 c | 6.3 c | 51a | |
| 80 | 8.3 b | 16.4 b | 8.2 b | 50 ab | |
| 120 | 8.8 a | 17.7 a | 9.0 a | 49 ab | |
| 160 | 8.7 a | 17.8 a | 9.1 a | 49 b | |
| 0 | +Vetch | 7.6 b | 14.5 c | 6.9 e | 52 a |
| −Vetch | 5.5c | 11.0 d | 5.5 f | 50 ab | |
| 80 | +Vetch | 8.8 a | 17.4 ab | 8.7 bc | 50 ab |
| −Vetch | 7.8 b | 15.5 c | 7.7 de | 50 ab | |
| 120 | +Vetch | 8.9 a | 18.4 a | 9.5 ab | 48 bc |
| −Vetch | 8.6 a | 17.1 b | 8.5 cd | 50 ab | |
| 160 | +Vetch | 8.9 a | 18.6 a | 9.7 a | 48 c |
| −Vetch | 8.5 a | 17.0 b | 8.5 cd | 50 abc | |
| Sources | P(F) values | ||||
| Cover crop management | 0.050 | 0.010 | 0.050 | ns | |
| Fertilization | 0.010 | 0.050 | 0.010 | ns | |
| Cover crop management × fertilization | 0.010 | ns | ns | 0.050 | |
| N Fertilization (kg N ha−1) | Cover Crop Management | Spikelets (panicle−1) | 103 Seeds Weight (g) | Panicle Density (m−2) | Sterility (%) | Tillering Rate | Milled Rice Yield (%) | Damaged Kernels (%) | Chalkiness (%) |
|---|---|---|---|---|---|---|---|---|---|
| Average | +Vetch | 142 a | 25.1 a | 509 a | 16.0 a | 2.3 a | 71.4 a | 0.48 a | 1.3 a |
| −Vetch | 129 b | 24.3 b | 490 a | 14.2 a | 1.9 a | 71.5 a | 0.54 a | 1.0 a | |
| 0 | 134 a | 25.5 a | 426 c | 10.1 c | 1.7 c | 71.6 a | 0.61 ab | 0.9 b | |
| 80 | 132 a | 24.8 b | 483 b | 14.2 b | 2.0 bc | 71.5 ab | 0.63 a | 1.2 ab | |
| 120 | 133 a | 24.4 c | 528 ab | 16.7 b | 2.3 ab | 71.3 b | 0.42 bc | 1.2 ab | |
| 160 | 145 a | 24.1 d | 562 a | 19.5 a | 2.4 a | 71.3 b | 0.39 c | 1.3 a | |
| 0 | +Vetch | 143 ab | 25.0 bc | 432 de | 11.0 d | 1.8 bc | 71.6 a | 0.69 ab | 1.1 ab |
| −Vetch | 124 b | 26.0 a | 420 e | 9.1 d | 1.5 c | 71.6 a | 0.53 abc | 0.7 b | |
| 80 | +Vetch | 133 ab | 24.6 cd | 497 bcd | 15.7 bc | 2.1 ab | 71.5 a | 0.52 abc | 1.2 ab |
| −Vetch | 131 b | 25.0 b | 469 cde | 12.7 cd | 1.9 bc | 71.5 a | 0.74 a | 1.2 ab | |
| 120 | +Vetch | 141 ab | 24.0 e | 530 abc | 18.6 ab | 2.4 a | 71.1 b | 0.40 bc | 1.3 a |
| −Vetch | 124 b | 24.9 bc | 527 abc | 14.7 c | 2.2 ab | 71.5 a | 0.41 bc | 1.1 ab | |
| 160 | +Vetch | 151 a | 23.8 e | 579 a | 19.0 ab | 2.6 a | 71.3 ab | 0.31 c | 1.5 a |
| −Vetch | 138 ab | 24.4 d | 545 ab | 20.0 a | 2.2 ab | 71.3 ab | 0.47 abc | 1.1 ab | |
| Sources | P(F) values | ||||||||
| Cover crop management | 0.050 | 0.050 | ns | ns | ns | ns | ns | ns | |
| Fertilization | ns | 0.010 | 0.010 | 0.010 | 0.010 | 0.050 | 0.050 | ns | |
| Cover crop management × fertilization | ns | ns | ns | ns | ns | ns | ns | ns | |
| N Fertilization (kg N ha−1) | Cover Crop Management | Grain N (%) | Straw N (%) | Total N Uptake (kg ha−1) | Apparent N Recovery (%) | Grain P (%) | Straw P (%) | Total P Uptake (kg ha−1) | Grain K (%) | Straw K (%) | Total K Uptake (kg ha−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Average | +Vetch | 1.40 a | 0.84 a | 195 a | 67.3 a | 0.26 a | 0.16 a | 38.5 a | 0.28 a | 2.21 a | 218 a |
| −Vetch | 1.21 b | 0.74 b | 149 b | 62.0 a | 0.28 a | 0.16 a | 32.0 b | 0.27 a | 1.69 b | 150 b | |
| 0 | 1.11 c | 0.65 c | 114 c | - | 0.24 c | 0.14 b | 24.4 b | 0.23 c | 1.77 d | 127 b | |
| 80 | 1.26 b | 0.75 bc | 166 b | 63.3 a | 0.28 b | 0.15 b | 34.7 a | 0.29 b | 1.87 c | 179 ab | |
| 120 | 1.40 a | 0.85 ab | 199 a | 71.2 a | 0.29 a | 0.17 a | 41.3 a | 0.30 a | 2.04 b | 211 a | |
| 160 | 1.45 a | 0.92 a | 209 a | 59.5 a | 0.28 b | 0.18 a | 40.6 a | 0.28 b | 2.11 a | 218 a | |
| 0 | +Vetch | 1.17 cd | 0.66 c | 134 c | - | 0.27 ab | 0.13 bc | 29.7 bc | 0.28 ab | 1.97 b | 158 bc |
| −Vetch | 1.05 d | 0.65 c | 93.9 d | - | 0.21 b | 0.14 bc | 19.1 c | 0.18 b | 1.58 c | 97.3 c | |
| 80 | +Vetch | 1.38 b | 0.80 abc | 190 b | 65.7 a | 0.28 a | 0.15 b | 37.4 ab | 0.28 ab | 2.12 b | 208 ab |
| −Vetch | 1.13 d | 0.70 c | 143 c | 60.9 a | 0.28 a | 0.14 bc | 32.1 b | 0.30 a | 1.63 c | 150 bc | |
| 120 | +Vetch | 1.52 a | 0.96 a | 227 a | 77.1 a | 0.29 a | 0.19 a | 43.5 a | 0.30 a | 2.33 a | 247 a |
| −Vetch | 1.27 bc | 0.74 bc | 172 b | 65.3 a | 0.30 a | 0.16 b | 39.0 ab | 0.31 a | 1.76 b | 175 abc | |
| 160 | +Vetch | 1.53 a | 0.96 a | 229 a | 59.1 a | 0.29 a | 0.18 ab | 43.2 a | 0.28 ab | 2.42 a | 259 a |
| −Vetch | 1.36 b | 0.88 ab | 189 b | 59.9 a | 0.26 bc | 0.18 ab | 38.0 ab | 0.27 ab | 1.81 b | 178 abc | |
| Sources | P(F) values | ||||||||||
| Cover crop management | 0.010 | 0.050 | 0.010 | ns | ns | ns | 0.050 | ns | 0.010 | 0.001 | |
| Fertilization | 0.010 | 0.010 | 0.010 | ns | 0.050 | 0.050 | 0.001 | 0.050 | 0.050 | 0.010 | |
| Cover crop management × fertilization | ns | ns | ns | ns | ns | ns | 0.001 | ns | ns | 0.001 | |
| N Fertilization (kg N ha−1) | Cover Crop Management | Days after Seeding | ||||||
|---|---|---|---|---|---|---|---|---|
| 31 | 42 | 55 | 67 | 81 | 94 | 109 | ||
| Average | +Vetch | 38.6 a | 42.9 a | 37.1 a | 34.5 a | 38.0 a | 40.2 a | 39.2 a |
| −Vetch | 35.0 b | 41.9 a | 36.0 a | 31.8 b | 35.3 b | 37.5 b | 36.1 b | |
| 0 | 36.4 a | 36.0 c | 32.0 d | 30.9 c | 32.1 d | 34.7 c | 33.3 d | |
| 80 | 37.1 a | 43.6 b | 35.7 c | 32.8 b | 35.7 c | 38.4 b | 36.6 c | |
| 120 | 36.9 a | 45.0 a | 38.4 b | 33.9 b | 38.1 b | 40.6 a | 39.2 b | |
| 160 | 36.9 a | 45.3 a | 40.1 a | 35.1 a | 40.6 a | 41.6 a | 41.5 a | |
| 0 | +Vetch | 38.2 a | 37.4 a | 32.9 a | 32.2 a | 33.7 a | 36.1 a | 34.3 a |
| −Vetch | 34.7 b | 34.6 b | 31.2 a | 29.6 b | 30.6 b | 33.3 b | 32.2 b | |
| 80 | +Vetch | 39.1 a | 44.4 a | 36.5 a | 34.6 a | 36.8 a | 39.7 a | 38.1 a |
| −Vetch | 35.1 b | 42.8 b | 34.9 b | 31.0 b | 34.7 b | 37.1 b | 35.1 b | |
| 120 | +Vetch | 38.7 a | 45.0 a | 38.7 a | 35.4 a | 39.5 a | 42.1 a | 40.9 a |
| −Vetch | 35.2 b | 45.0 a | 38.1 a | 32.5 b | 36.7 b | 39.1 b | 37.5 b | |
| 160 | +Vetch | 38.6 a | 45.1 a | 40.4 a | 36.0 a | 42.1 a | 42.8 a | 43.5 a |
| −Vetch | 35.2 b | 45.4 a | 39.9 a | 34.3 b | 39.2 b | 40.4 b | 39.5 b | |
| Sources | P(F) values | |||||||
| Cover crop management | 0.050 | ns | ns | 0.010 | 0.010 | 0.010 | 0.010 | |
| Fertilization | ns | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | 0.010 | |
| Cover crop management × fertilization | ns | 0.050 | ns | ns | ns | ns | ns | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lizcano Toledo, R.; Lerda, C.; Moretti, B.; Miniotti, E.; Santoro, V.; Fernandez-Ondoño, E.; Martin, M.; Said-Pullicino, D.; Romani, M.; Celi, L. Cover Crops Increase N and P Cycling and Rice Productivity in Temperate Cropping Systems. Agronomy 2022, 12, 2193. https://doi.org/10.3390/agronomy12092193
Lizcano Toledo R, Lerda C, Moretti B, Miniotti E, Santoro V, Fernandez-Ondoño E, Martin M, Said-Pullicino D, Romani M, Celi L. Cover Crops Increase N and P Cycling and Rice Productivity in Temperate Cropping Systems. Agronomy. 2022; 12(9):2193. https://doi.org/10.3390/agronomy12092193
Chicago/Turabian StyleLizcano Toledo, Rodolfo, Cristina Lerda, Barbara Moretti, Eleonora Miniotti, Veronica Santoro, Emilia Fernandez-Ondoño, Maria Martin, Daniel Said-Pullicino, Marco Romani, and Luisella Celi. 2022. "Cover Crops Increase N and P Cycling and Rice Productivity in Temperate Cropping Systems" Agronomy 12, no. 9: 2193. https://doi.org/10.3390/agronomy12092193
APA StyleLizcano Toledo, R., Lerda, C., Moretti, B., Miniotti, E., Santoro, V., Fernandez-Ondoño, E., Martin, M., Said-Pullicino, D., Romani, M., & Celi, L. (2022). Cover Crops Increase N and P Cycling and Rice Productivity in Temperate Cropping Systems. Agronomy, 12(9), 2193. https://doi.org/10.3390/agronomy12092193