


Optimizing Nitrogen Fertilization and Variety for Millet Grain Yield and Biomass Accumulation in Dry Regions



Abstract
1. Introduction
2. Materials and Methods
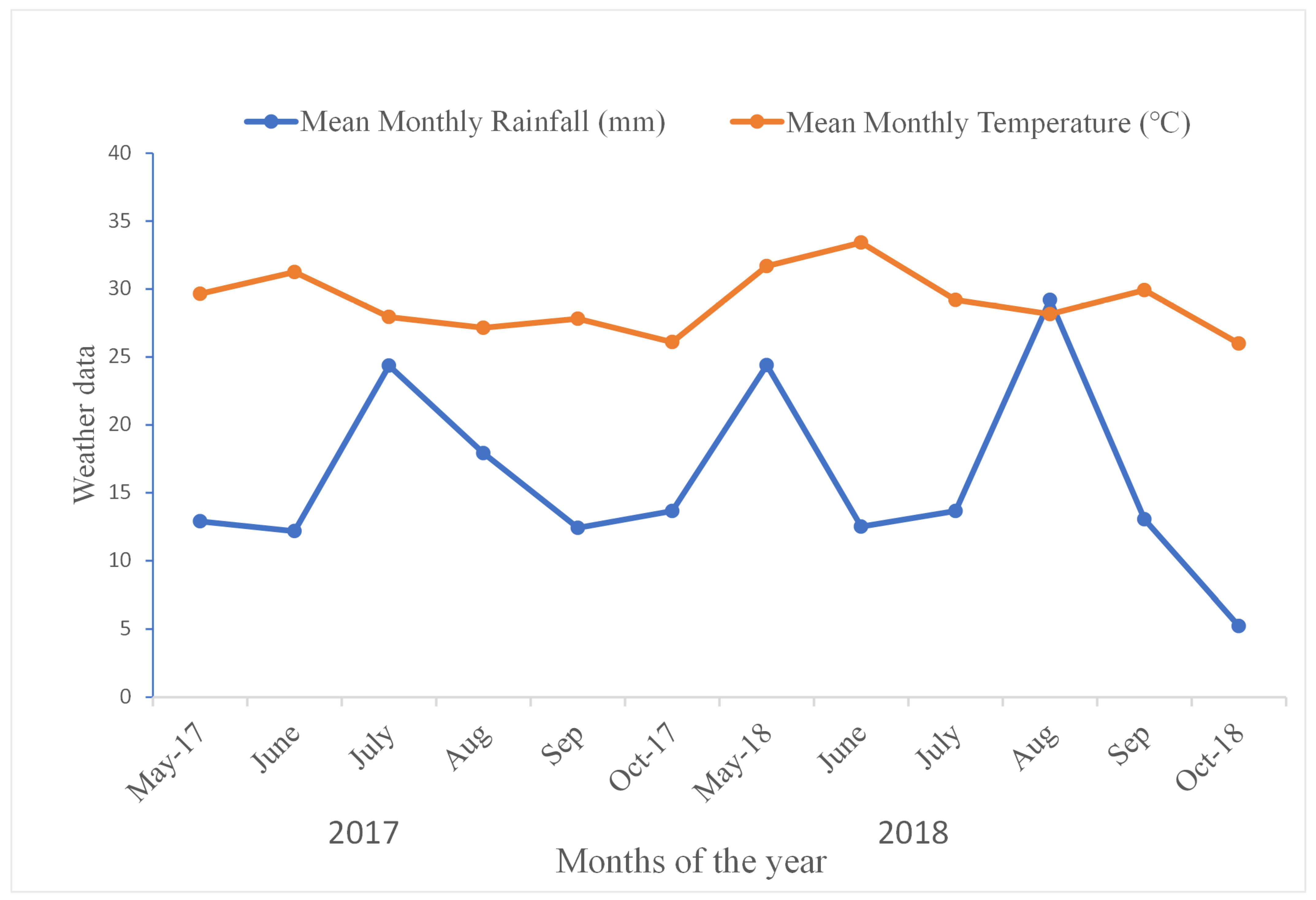
2.1. Experimental Site Details
2.2. Experimental Design and Cultivar Details
2.3. Field Preparation and Crop Management
2.4. Vegetative and Reproductive Organ Biomass Accumulation
2.5. Foxtail Millet (FM) Grain
2.6. Biomass Simulation
2.7. Statistical Analysis
3. Results
3.1. Foxtail Millet (FM) Grain
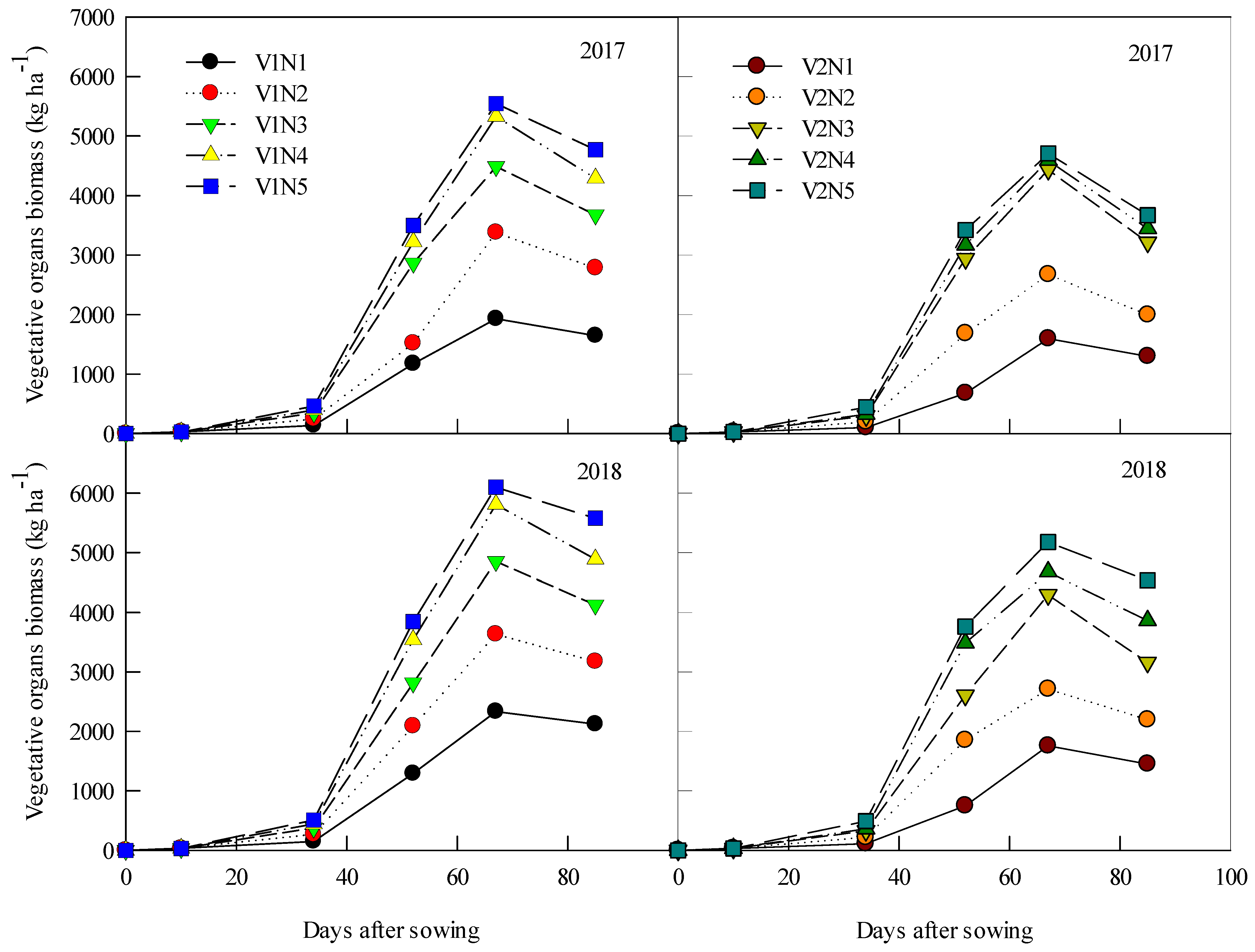
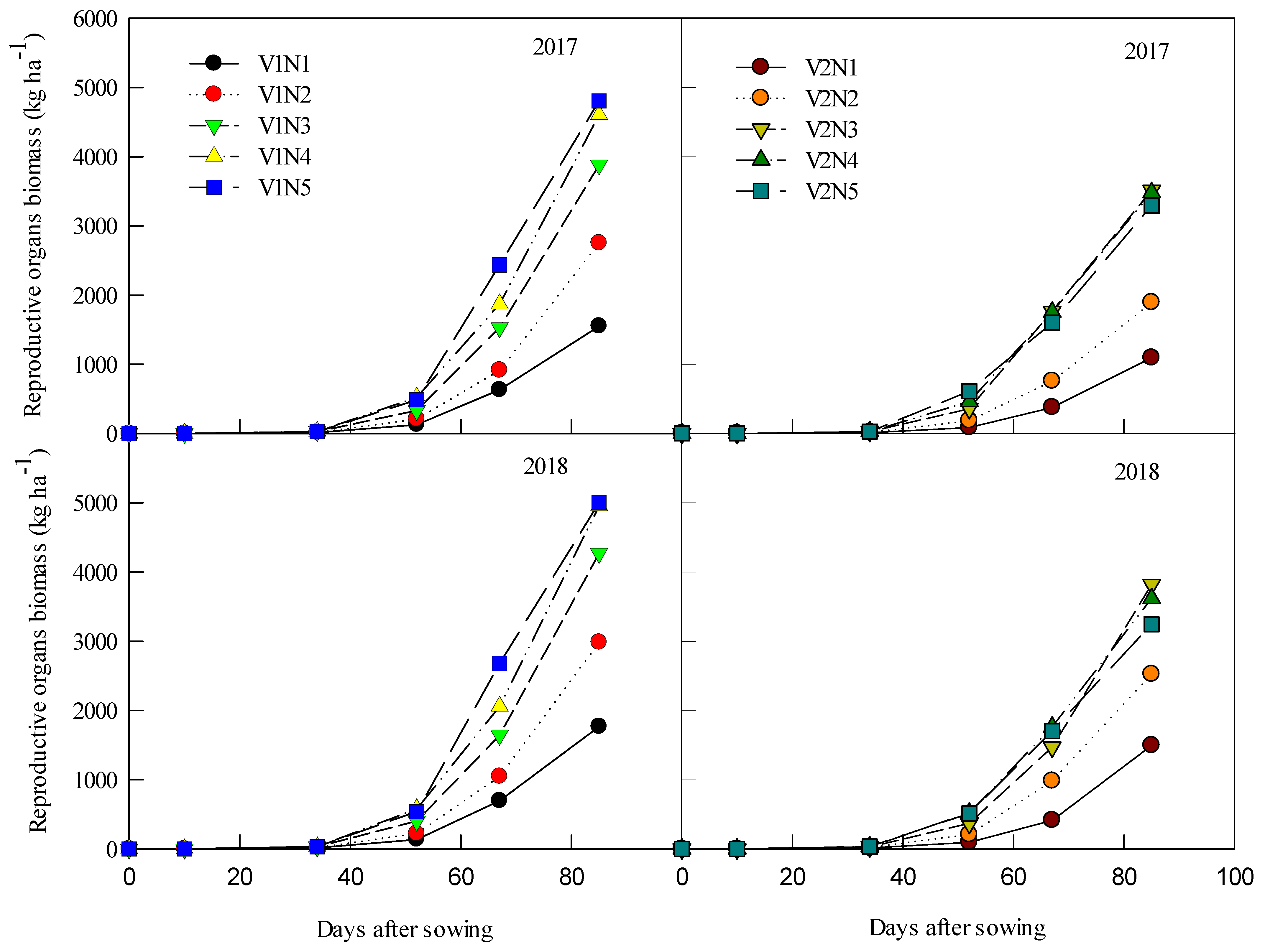
3.2. Millet Plant Biomass Accumulation
3.3. Average Accumulation Rate of Vegetative Organs Biomass
3.4. Average Accumulation Rate of Reproductive Organ Biomass (ROB)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.; Jia, G.; Zhi, H.; Niu, Z.; Chai, Y.; Li, W. Genetic diversity and population structure of Chinese foxtail millet landraces. Genes Genome Genet. 2012, 2, 769–777. [Google Scholar] [CrossRef]
- Liu, F.; Liu, M.; Zhao, Y.; Nan, C.; Xia, X.; Li, S.; Xu, L. Development trends of China’s foxtail millet and broomcom millet industries. Agric. Outlook 2017, 13, 40–43. [Google Scholar]
- Doust, A.N.; Kellogg, E.A.; Devos, K.M.; Bennetzen, J.L. Foxtail millet: A sequence-driven grass model system. Plant Physiol. 2009, 149, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Diao, X.M.; Schnable, J.; Bennetzen, J.L.; Li, J.Y. Initiation of Setaria as a model plant. Front. Agric. Sci. Eng. 2014, 1, 16–20. [Google Scholar]
- Zhang, G.; Liu, X.; Quan, Z.; Cheng, S.; Xu, X.; Pan, S. Genome sequence of foxtail millet (Setaria italica) provides insights into grass evolution and biofuel potential. Nat. Biotechnol. 2012, 30, 549–554. [Google Scholar] [CrossRef]
- Joshi, N.L.; Kalla, J.C. Economizing nitrogen use of pearl millet in the Indian arid zone. Ann. Arid. Zone 1986, 25, 321–325. [Google Scholar] [CrossRef]
- Khan, A.; Tan, D.K.Y.; Munsif, F.; Afridi, M.Z.; Shah, F.; Wei, F.; Zhou, R. Nitrogen nutrition in cotton and control strategies for greenhouse gas emissions: A review. Environ. Sci. Pollut. Res. 2017, 24, 23471–23487. [Google Scholar] [CrossRef]
- Khan, A.; Tan, D.K.Y.; Afridi, M.Z.; Luo, H.; Tung, S.A.; Ajab, M.; Fahad, S. Nitrogen fertility and abiotic stresses management in cotton crop: A review. Environ. Sci. Pollut. Res. 2017, 24, 14551–14566. [Google Scholar] [CrossRef]
- Chen, Z.; Khan, A.; Shi, X.; Hao, X.; Tan, D.K.Y.; Luo, H. Water-nutrient management enhances root morpho-physiological functioning, phosphorus absorption, transportation and utilization of cotton in arid region. Ind. Crops Prod. 2020, 143, 1–8. [Google Scholar] [CrossRef]
- Nadeem, F.; Ahmad, Z.; Wang, R.; Hassam, M.; Diao, X.; Li, X. Adaptation of Foxtail Millet (Setaria italica L.) to Abiotic Stresses: A Special Perspective of Responses to Nitrogen and Phosphate Limitations. Front. Plant Sci. 2018, 11, 11–187. [Google Scholar] [CrossRef]
- Uppal, R.K.; Wani, S.P.; Garg, K.K.; Alagarswamy, G. Balanced nutrition increases yield of pearl millet under drought. Field Crops Res. 2015, 177, 86–97. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, X.; Yang, J.; Cheng, B.; Luo, S. Effects of different using methods of nitrogen fertilizer on the growth and yield of millet. J. Agric. Sci. 2015, 36, 25–28. [Google Scholar] [CrossRef]
- Yan, Z.; Guo, P.; Yuan, X.; Xiao, X.; Wang, D. Effect of foliar application of nitrogen fertilizer on photosynthetic characteristics and yield of foxtail millet cultivar ‘zhang zagu 10′ Setaria italic (L.) Beauv. J. Shanxi Agric. Univ. 2018, 38, 37–41. [Google Scholar]
- Wani, S.P.; Sahrawat, K.L.; Sreedevi, T.K.; Bhattacharyya, T.; Srinivasa, R.C. Carbon sequestration in the semi-arid tropics for improving livelihoods. Int. J. Environ. Stud. 2007, 64, 719–727. [Google Scholar] [CrossRef]
- Ciećko, Z.; Żołnowski, A.; Mierzejewska, A. Effect of foliar nitrogen and magnesium fertilization on the total, protein nitrogen and nitrates(V) content in potato tubers. Ecol. Chem. Eng. 2010, 17, 593–600. [Google Scholar]
- Sumner, M.E.; Miller, W.P. Cation exchange capacity and exchange coefficients. In Methods of Soil Analysis, Part 3, Chemical Methods; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 1201–1229. [Google Scholar] [CrossRef]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis, Part 3. Chemical Methods; Black, C.A., Ed.; Soil Science of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 961–1010. [Google Scholar]
- Dorich, R.A.; Nelson, D.W. Evaluation of manual cadmium reduction methods for determination of nitrate in potassium chloride extracts of soils. J. Soil Sci. Soc. Am. 1984, 48, 72–75. [Google Scholar] [CrossRef]
- Kosteckas, R.; Marcinkevicienee, A. The integrated evaluation of the influence of catch crops and manure on spring barley agrocenosis in organic farming. Agron. Res. 2009, 1, 355–362. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; Chinese Agriculture Press: Beijing, China, 2012; pp. 8–9. [Google Scholar]
- Knudsen, D.; Peterson, G.A.; Pratt, P.F. Lithium, sodium, and potassium. In Methods of Soil Analysis, Part 2. Chemical and Microbiological Properties; Page, A.L., Ed.; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1982; pp. 225–246. [Google Scholar] [CrossRef]
- Yang, G.; Tang, H.; Nie, Y.; Zhang, X. Response of cotton growth, yield, and biomass to nitrogen split application ratio. Eur. J. Agron. 2011, 35, 164–170. [Google Scholar] [CrossRef]
- Khan, A.; Kong, X.; Najeeb, U.; Zheng, J.; Tan, D.K.Y.; Akhtar, K.; Zhou, R. Planting density induced changes in cotton biomass yield, fiber quality, and phosphorus distribution under beta growth model. Agronomy 2019, 9, 500. [Google Scholar] [CrossRef]
- Song, M.; Li, Z.; Feng, H. Effects of irrigation and nitrogen regimes on dry matter dynamic accumulation and yield of winter wheat. Trans. Chin. Soc. Agric. Eng. 2016, 32, 119–126. [Google Scholar] [CrossRef]
- Khan, A.; Najeeb, U.; Wang, L. Planting density and sowing date strongly influence growth and lint yield of cotton crops. Field Crops Res. 2017, 209, 129–135. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, J.; Wang, Y. Effect of drought stress on dry matter accumulation and yield of summer millet. J. Northeast Agric. Univ. 2016, 47, 15–22. [Google Scholar]
- Zhang, Y.; Wang, Z.; Fu, Y.; Meng, A.; Feng, G.; Hu, X. Effect of nitrogen application amount on the growth and yield of foxtail millet by drip irrigation under mulch. J. Xinjiang Agric. Sci. 2017, 54, 1061–1065. [Google Scholar]
- Ali, H.; Afzal, M.N.; Ahmad, S.; Muhammad, D. Effect of cultivars and sowing dates on yield and quality of (Gossypium hirsutum). J. Food Agric. Environ. 2009, 7, 244–247. [Google Scholar]
- Khan, A.; Wang, L.; Ali, S.; Tung, S.A.; Hafeez, A.; Yang, G. Optimal planting density and sowing date can improve cotton yield by maintaining reproductive organ biomass and enhancing potassium uptake. Field Crops Res. 2017, 214, 164–174. [Google Scholar] [CrossRef]
- Khan, A.; Munsif, F.; Akhtar, K.; Afridi, M.Z.; Ahmad, Z.; Fahad, S.; Ullah, R.; Khan, F.A.; Din, M. Response of fodder maize to various levels of nitrogen and phosphorus. Am. J. Plant Sci. 2014, 5, 2323–2329. [Google Scholar] [CrossRef][Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Grain Yield (kg ha−1) | |
|---|---|---|
| 2017 | 2018 | |
| Variety (V) | ||
| V1 | 2813.5a | 3048.3a |
| V2 | 2087.0b | 2370.9b |
| Nitrogen (N) | ||
| N1 | 1058.2d | 1304.1d |
| N2 | 1841.0c | 2203.4c |
| N3 | 2946.1b | 3252.2b |
| N4 | 3225.9a | 3436.6a |
| N5 | 3181.1a | 3351.8ab |
| Interaction (V × N) | ||
| V1N1 | 1227.7f | 1401.3g |
| V1N2 | 2179.6d | 2386.9e |
| V1N3 | 3156.4b | 3444.8b |
| V1N4 | 3730.2a | 3977.0a |
| V1N5 | 3773.5a | 4022.2a |
| V2N1 | 888.7g | 1197.8g |
| V2N2 | 1502.3e | 2019.9f |
| V2N3 | 2734.0c | 3059.5c |
| V2N4 | 2721.6c | 2896.1cd |
| V2N5 | 2588.6c | 2681.4de |
| Source of variance (SOV) | ||
| V | <0.0001 | <0.0001 |
| N | <0.0001 | <0.0001 |
| V × N | <0.0001 | <0.0068 |
| Year | Treatment | DAS | DAS | DAS | DAS | DAS |
|---|---|---|---|---|---|---|
| 0–10 | 10–34 | 34–52 | 52–67 | 67–85 | ||
| Variety | ||||||
| 2017 | V1 | 2.9 | 12.1a | 118.7a | 112.0a | −39.0a |
| V2 | 3.0 | 10.2b | 116.8a | 81.6b | −48.7b | |
| 2018 | V1 | 3.2 | 13.2a | 131.6a | 121.9a | −31.5a |
| V2 | 3.2 | 11.2b | 121.6b | 82.8b | −38.0b | |
| Nitrogen | ||||||
| 2017 2018 | N1 | 3.0 | 3.8e | 44.8d | 55.8c | −16.1a |
| N2 | 3.0 | 8.1d | 76.6c | 87.3b | −35.5b | |
| N3 | 2.9 | 12.2c | 143.2b | 112.0a | −56.5d | |
| N4 | 2.9 | 14.0b | 157.3a | 117.8a | −60.7d | |
| N5 | 2.8 | 17.8a | 166.9a | 111.2a | −50.4c | |
| N1 | 3.3 | 4.1e | 49.4d | 68.3b | −14.4a | |
| N2 | 3.2 | 8.8d | 95.9c | 80.4b | −27.2b | |
| N3 | 3.1 | 13.3c | 131.2b | 124.2a | −51.7c | |
| N4 | 3.2 | 15.5b | 172.9a | 115.3a | −48.2c | |
| N5 | 3.1 | 19.6a | 183.6a | 122.3a | −32.3b | |
| Source of variance (SOV) | ||||||
| 2017 | V | 0.307 | <0.0003 | 0.5983 | 0.0004 | 0.0002 |
| N | 0.5348 | <0.0001 | <0.0001 | 0.0002 | <0.0001 | |
| V × N | 0.6455 | 0.5298 | 0.0294 | 0.0241 | 0.0252 | |
| 2018 | V | 0.4052 | <0.0003 | 0.015 | <0.0001 | 0.028 |
| N | 0.5559 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| V × N | 0.5452 | 0.4492 | 0.0663 | 0.0546 | 0.0756 | |
| Year | Treatment | DAS | DAS | DAS | DAS | DAS |
|---|---|---|---|---|---|---|
| 0–10 | 10–34 | 34–52 | 52–67 | 67–85 | ||
| Variety | ||||||
| 2017 | V1 | 0 | 1.0 | 17.4 | 70.6a | 113.5a |
| V2 | 0 | 1.0 | 17.7 | 60.6b | 78.0b | |
| 2018 | V1 | 0 | 1.1 | 19.5 | 83.3a | 120.6a |
| V2 | 0 | 1.1 | 17.5 | 61.8b | 93.1b | |
| Nitrogen | ||||||
| 2017 | N1 | 0 | 0.5c | 5.3d | 26.4d | 45.5d |
| N2 | 0 | 0.8b | 10.0c | 42.6c | 82.7c | |
| N3 | 0 | 1.3a | 17.6b | 78.3b | 113.8ab | |
| N4 | 0 | 1.3a | 25.8a | 91.3ab | 124.0a | |
| N5 | 0 | 1.3a | 28.9a | 102.9a | 112.9b | |
| 2018 | N1 | 0 | 0.6b | 5.7d | 29.2d | 59.8c |
| N2 | 0 | 0.8b | 10.7c | 53.3c | 96.7b | |
| N3 | 0 | 1.2a | 19.8b | 78.1b | 138.3a | |
| N4 | 0 | 1.3a | 28.9a | 90.8b | 132.1a | |
| N5 | 0 | 1.4a | 27.3a | 111.3a | 107.3b | |
| Source of variance (SOV) | ||||||
| 2017 | V | 0 | 0.4152 | 0.8086 | 0.0035 | <0.0001 |
| N | 0 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| V × N | 0 | 0.0062 | <0.0001 | 0.0078 | 0.0064 | |
| 2018 | V | 0 | 0.7564 | 0.0907 | 0.0004 | <0.0001 |
| N | 0 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | |
| V × N | 0 | 0.6529 | 0.9722 | 0.0697 | 0.0595 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, S.; Wang, L.; Akhtar, K.; Ahmad, I.; Khan, A. Optimizing Nitrogen Fertilization and Variety for Millet Grain Yield and Biomass Accumulation in Dry Regions. Agronomy 2022, 12, 2116. https://doi.org/10.3390/agronomy12092116
Yang S, Wang L, Akhtar K, Ahmad I, Khan A. Optimizing Nitrogen Fertilization and Variety for Millet Grain Yield and Biomass Accumulation in Dry Regions. Agronomy. 2022; 12(9):2116. https://doi.org/10.3390/agronomy12092116
Chicago/Turabian StyleYang, Shuang, Leishan Wang, Kashif Akhtar, Ijaz Ahmad, and Aziz Khan. 2022. "Optimizing Nitrogen Fertilization and Variety for Millet Grain Yield and Biomass Accumulation in Dry Regions" Agronomy 12, no. 9: 2116. https://doi.org/10.3390/agronomy12092116
APA StyleYang, S., Wang, L., Akhtar, K., Ahmad, I., & Khan, A. (2022). Optimizing Nitrogen Fertilization and Variety for Millet Grain Yield and Biomass Accumulation in Dry Regions. Agronomy, 12(9), 2116. https://doi.org/10.3390/agronomy12092116