
Remote Sensing-Based Evaluation of Heat Stress Damage on Paddy Rice Using NDVI and PRI Measured at Leaf and Canopy Scales

 ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
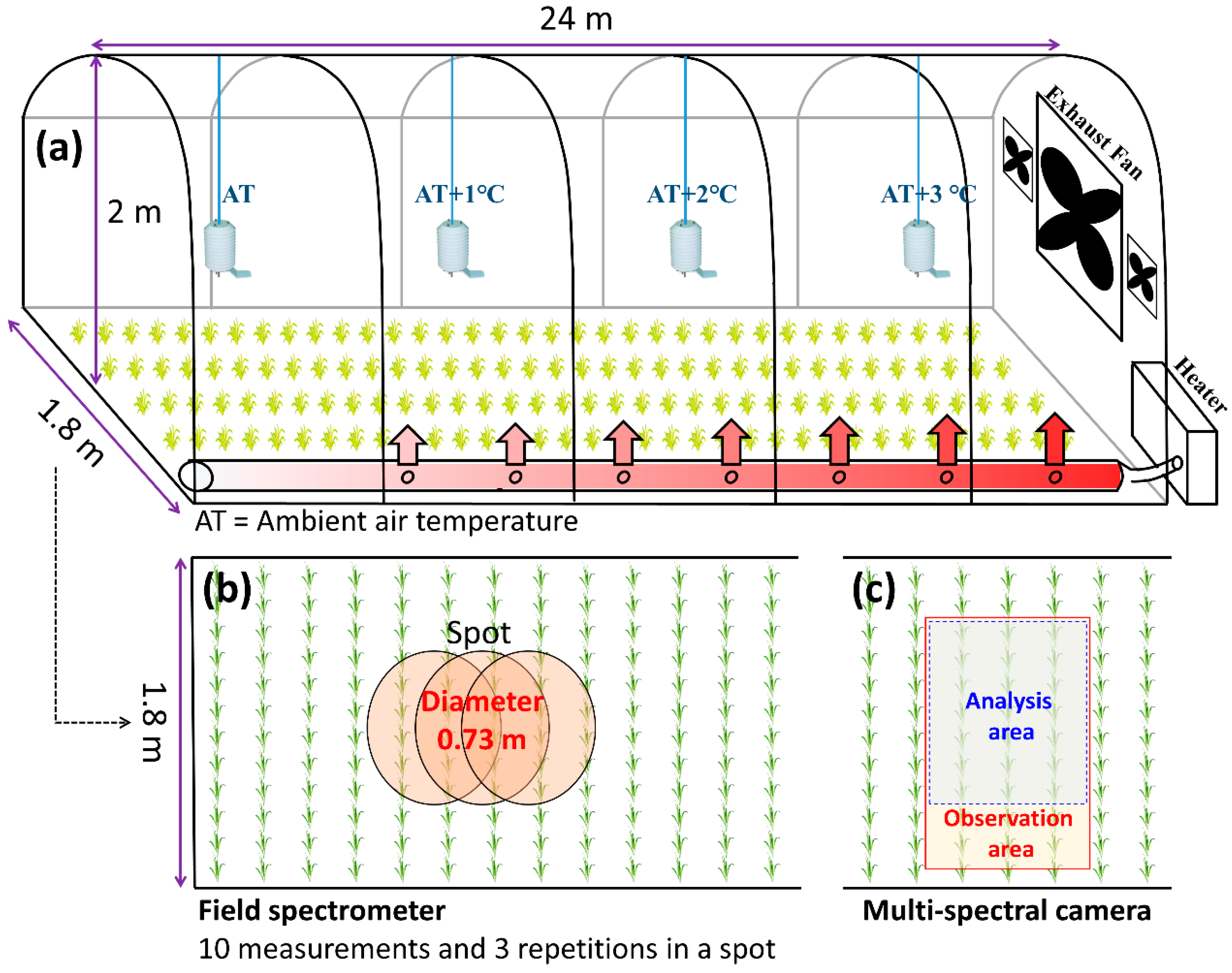
2.1. Experimental Facility and Cultivation of Paddy Rice
2.2. Remote Sensing Data
2.2.1. Leaf-Spectrometer
2.2.2. Field-Spectrometer
2.2.3. Multi-Spectral Camera
2.3. Relative Change in Vegetation Index after Heading
2.4. Meteorological Characteristics
3. Results
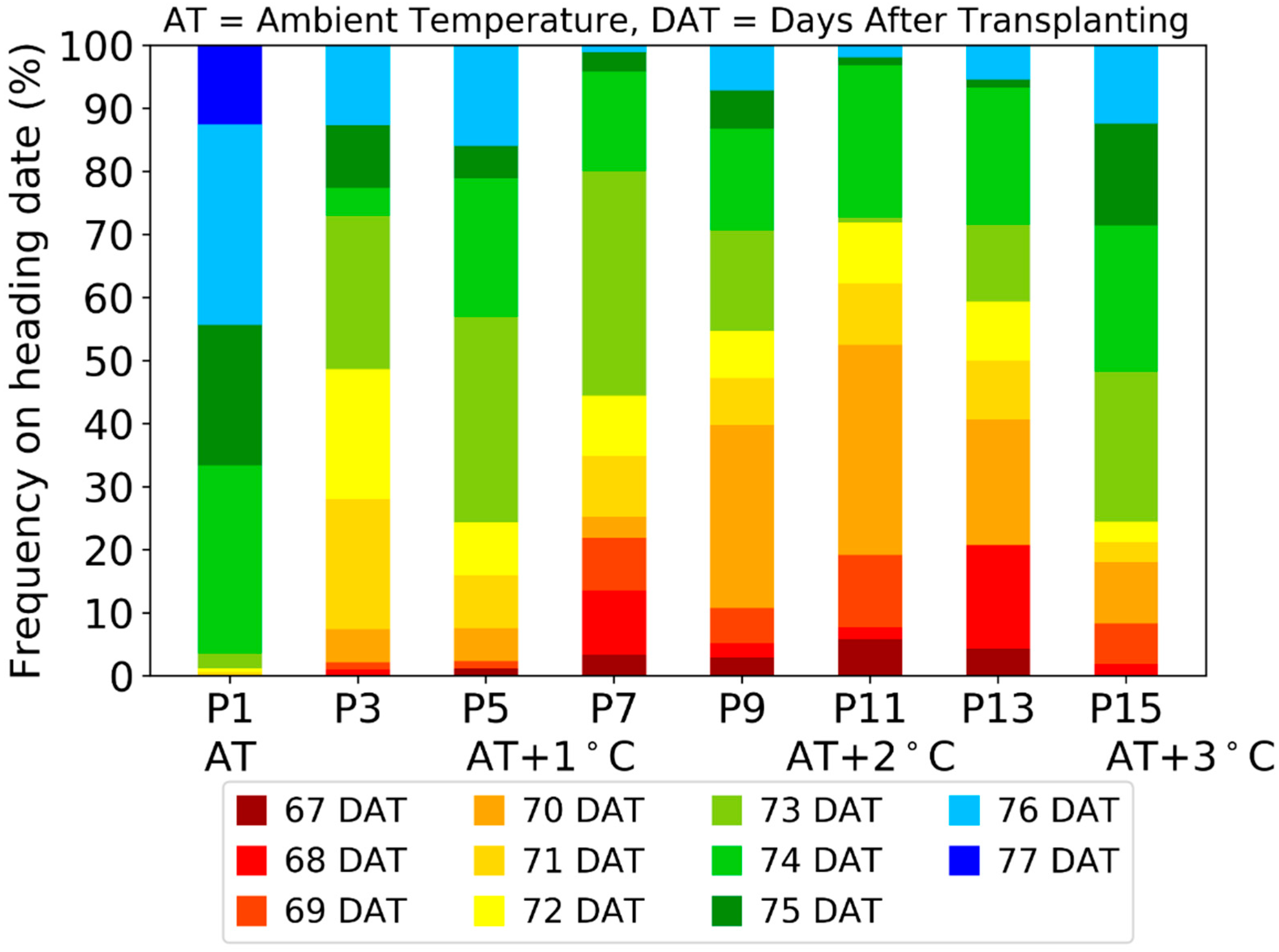
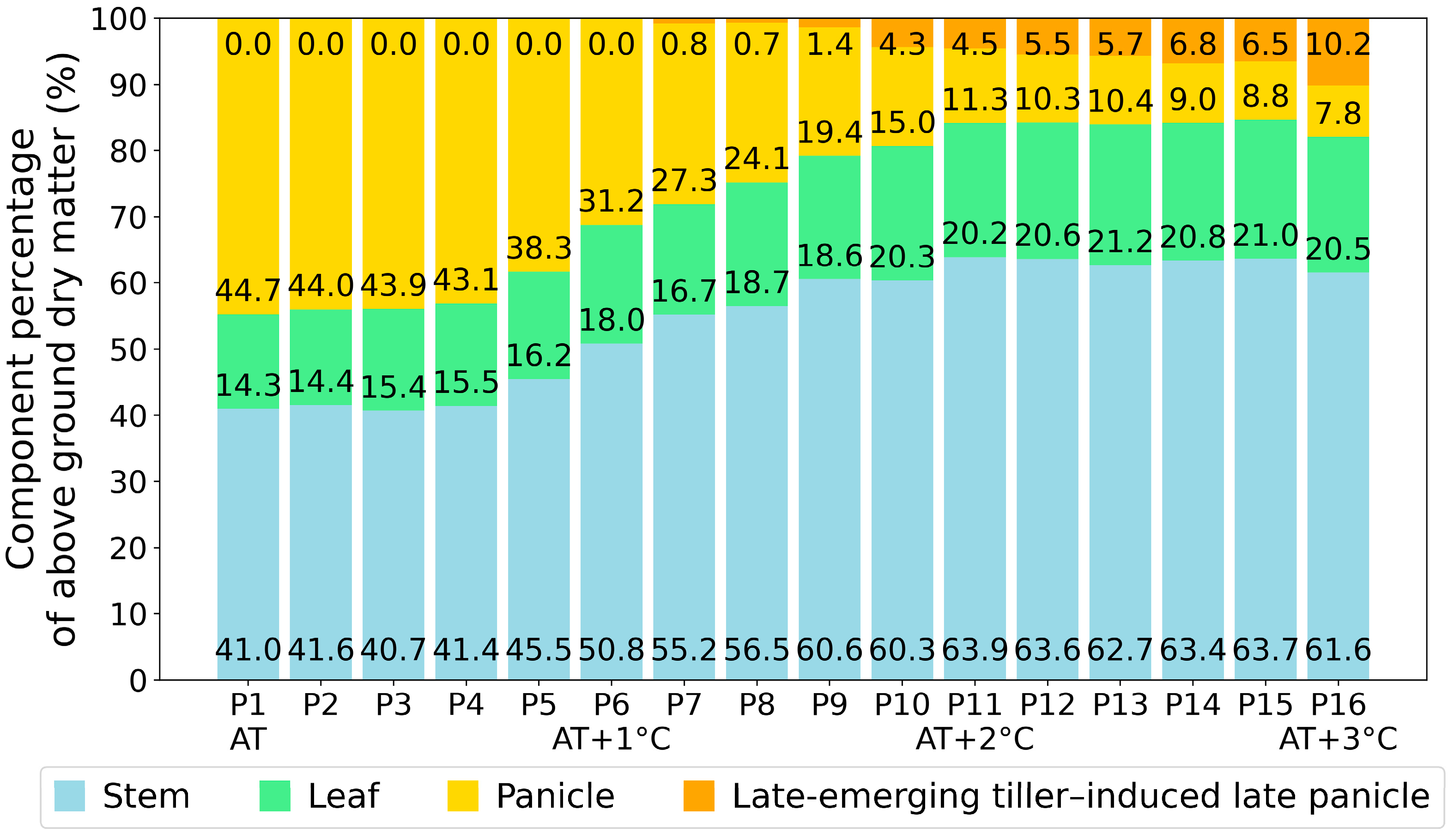
3.1. Response of Paddy Rice under Elevated Air Temperature
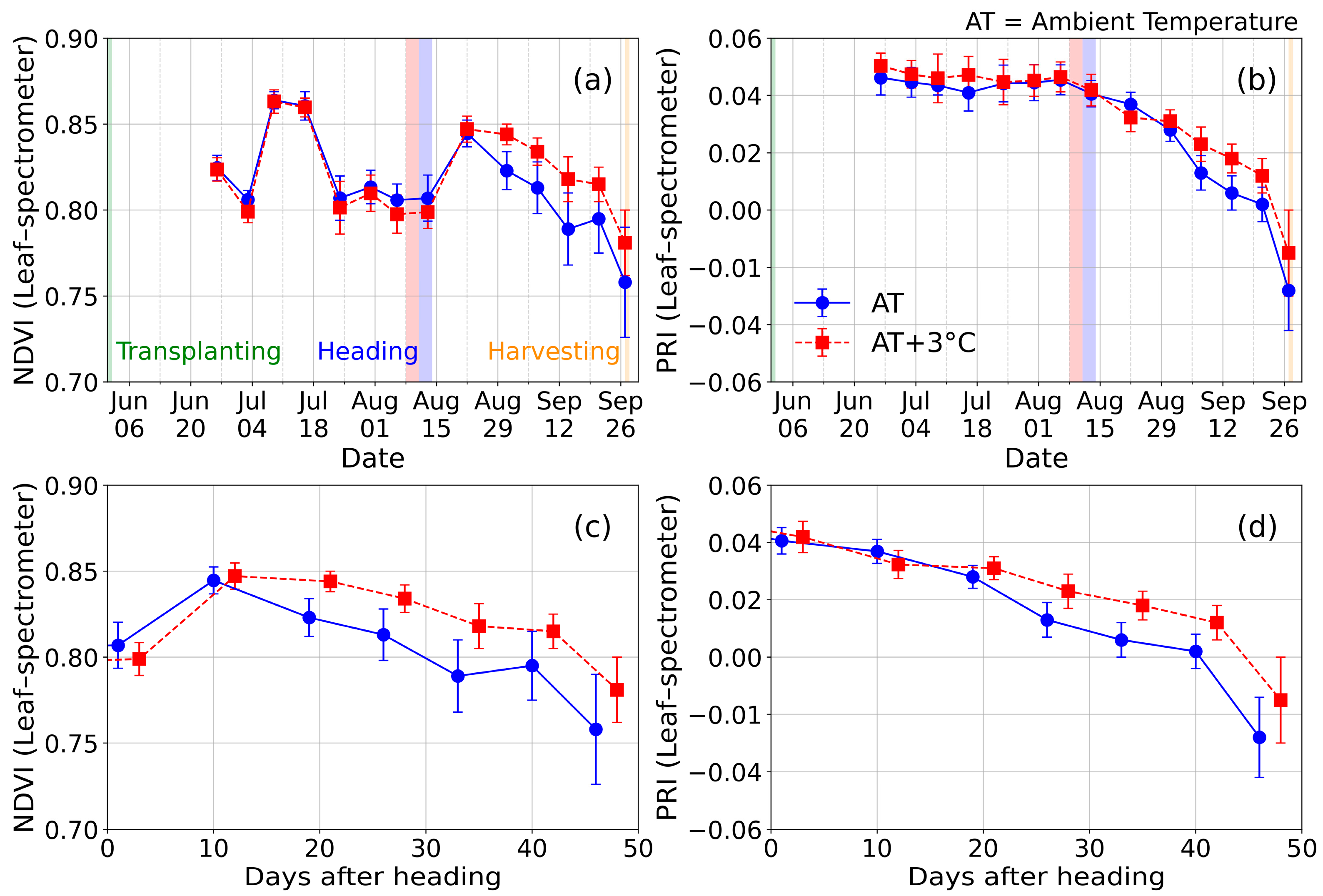
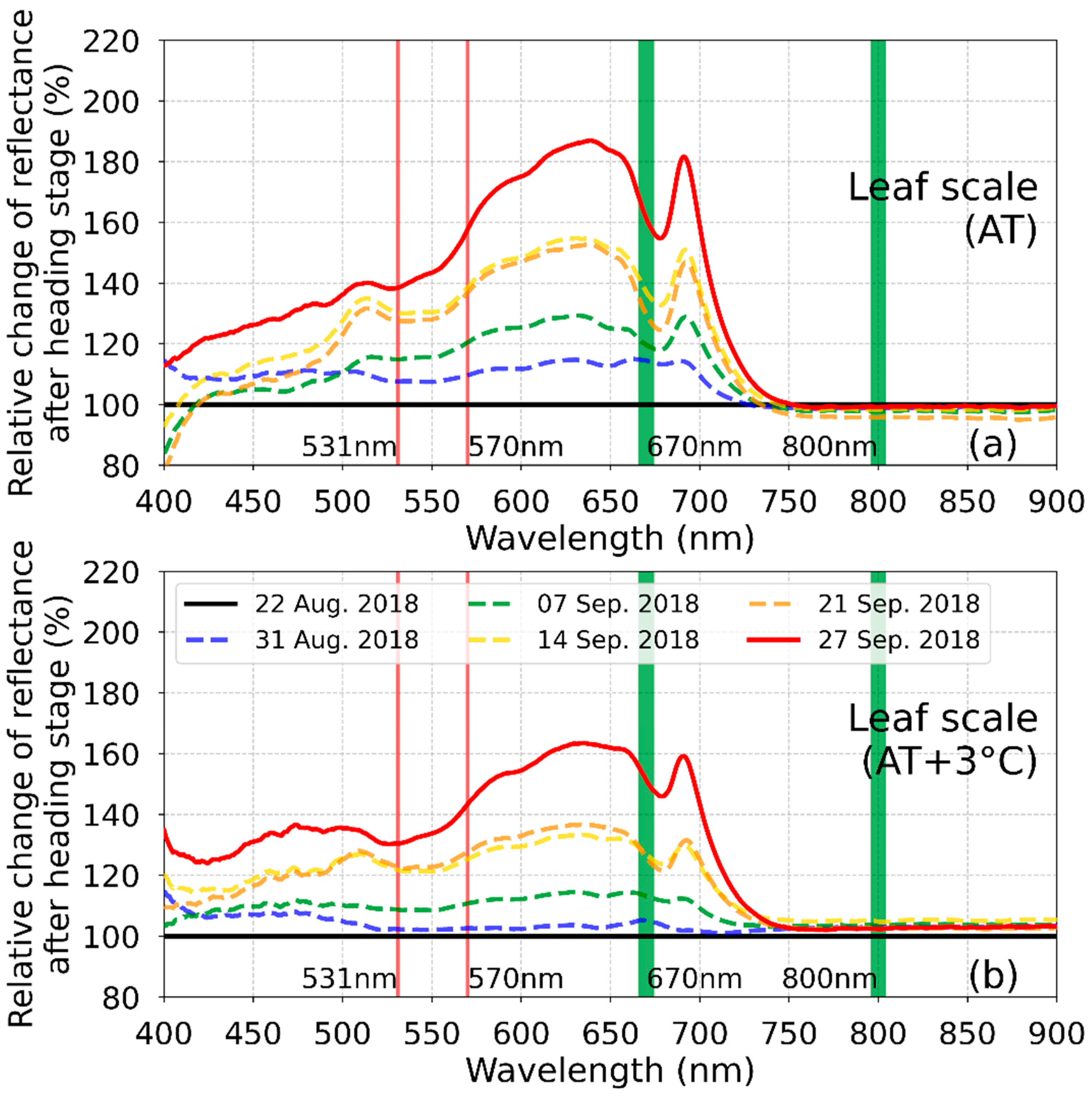
3.2. Time Series of NDVI and PRI at Leaf Scale
3.3. Time Series of NDVI and PRI at Canopy Scale
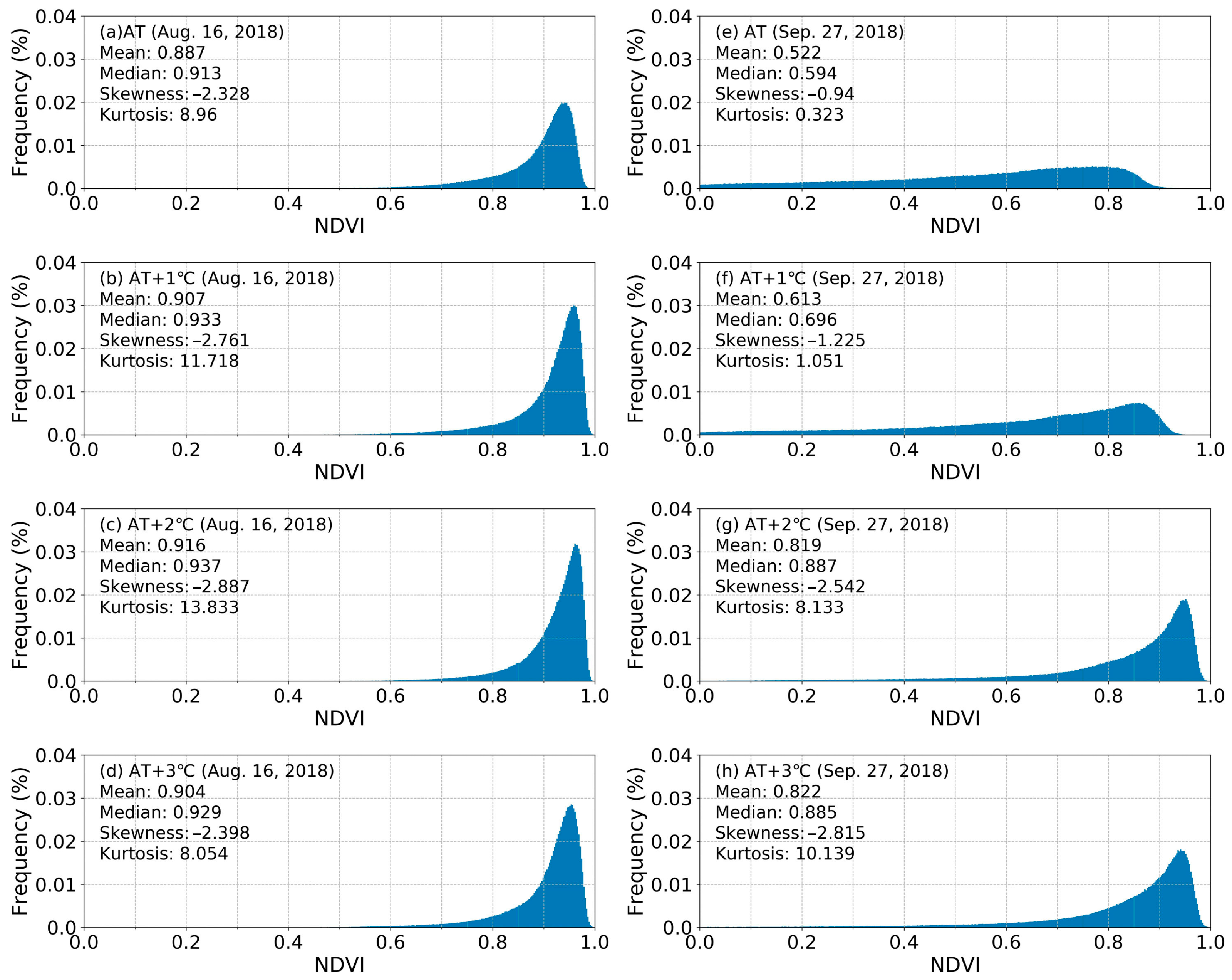
3.4. NDVI Captured by a Multi-Spectral Camera
3.5. Characteristics of NDVI and PRI after the Heading Stage
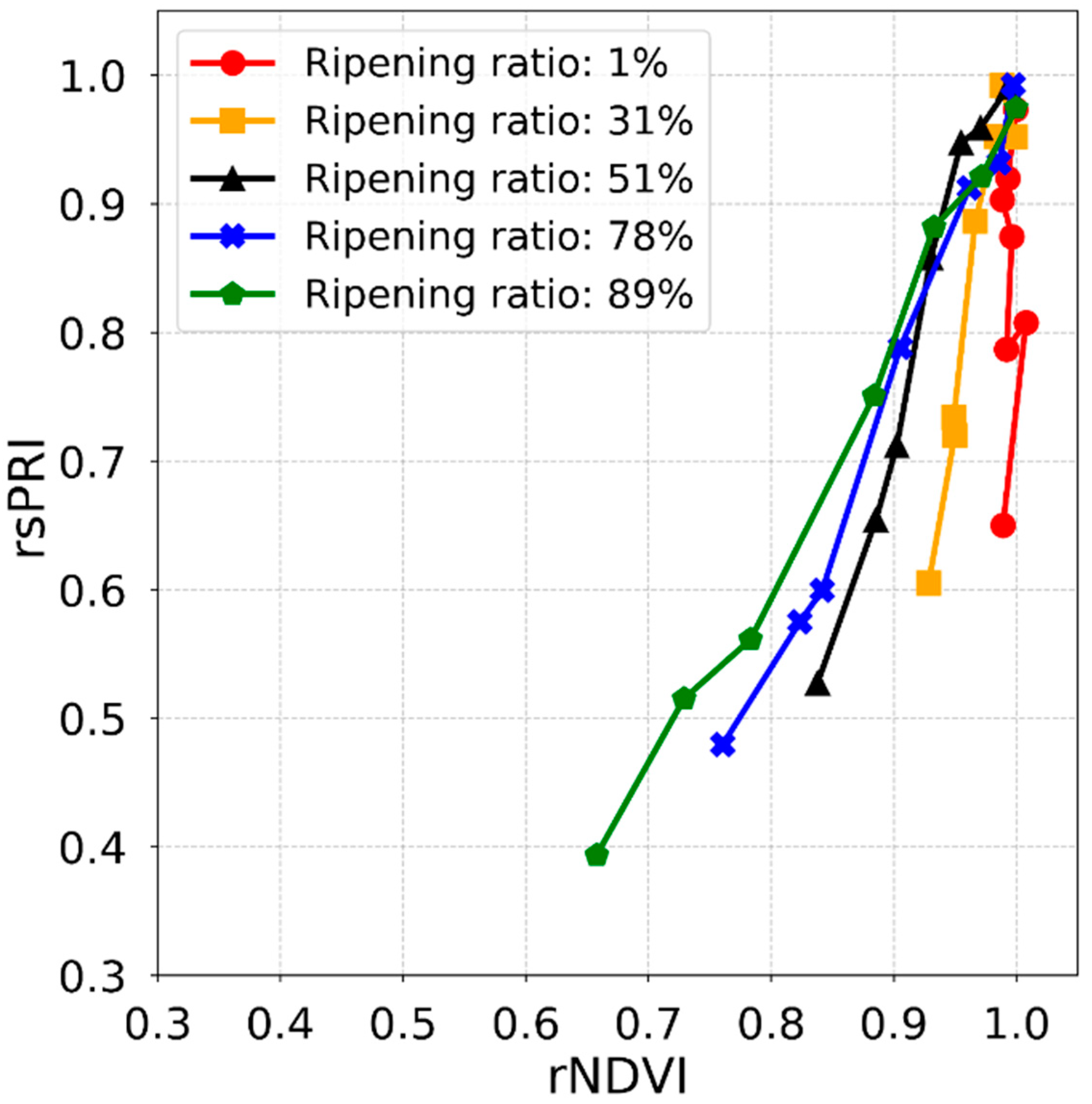
3.6. Estimation of Ripening Ratio of Paddy Rice
4. Discussion
5. Conclusions
- (1)
- The changing trend of the vegetation indices are dependent on the degree of heat stress after the heading stage at the leaf and canopy scales. Vegetation indices of NDVI and PRI decrease under normal conditions after the heading stage due to leaf senescence; however, the decreased degree of vegetation indices is minimal under extremely high air temperatures.
- (2)
- The spectral reflectance at the red wavelength causes different NDVI trends after the heading stage. The changing degree of the spectral reflectance at the red wavelength at the positions exposed to the heatwave is lower than that under normal conditions.
- (3)
- The PRI, which uses visible wavelengths, decreases after the heading stage regardless of the damage of heat stress. However, the decreasing degree at AT+3 °C is lower than that at AT, similar to the NDVI.
- (4)
- The low spectral reflectance value at the red wavelength indicates that paddy rice maintains a high chlorophyll content, as the translocation of assimilates from the stems (leaf sheath and culms) to the panicle does not occur due to spikelet sterility.
- (5)
- The vegetation index reflects the conditions of paddy rice caused by heat stress, and the relative change in the vegetation index after the heading stage exhibits a significant sigmoid relation with the ripening ratio affected by the spikelet sterility of paddy rice.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Horie, T. Global warming and rice production in Asia: Modeling, impact prediction and adaptation. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 211–245. [Google Scholar] [CrossRef] [Green Version]
- Horie, T.; Nakagawa, H.; Nakano, J.; Hamotani, K.; Kim, H.Y. Temperature gradient chambers for research on global environment change. III. A system designed for rice in Kyoto, Japan. Plant Cell Environ. 1995, 18, 1064–1069. [Google Scholar] [CrossRef]
- Kim, H.Y.; Horie, T.; Nakagawa, H.; Wada, K. Effects of elevated CO2 concentration and high temperature on growth and yield of rice: II. The effect on yield and its components of Akihikari rice. Jpn. J. Crop. Sci. 1996, 65, 644–651. [Google Scholar] [CrossRef] [Green Version]
- Oh-e, I.; Saitoh, K.; Kuroda, T. Effects of high temperature on growth, yield and dry-matter production of rice grown in the paddy field. Plant Prod. Sci. 2007, 10, 412–422. [Google Scholar] [CrossRef]
- Prasad, P.V.V.; Boote, K.J.; Alle, L.H., Jr.; Sheehy, J.E.; Thomas, J.M.G. Species, ecotype and cultivar differences in spikelet fertility and harvest index of rice in response to high temperature stress. Field Crops Res. 2006, 95, 398–411. [Google Scholar] [CrossRef]
- You, L.; Rosegrant, M.W.; Wood, S.; Sun, D. Impact of growing season temperature on wheat productivity in China. Agric. For. Meteorol. 2009, 149, 1009–1014. [Google Scholar] [CrossRef]
- Tian, Y.; Chen, J.; Chen, C.; Deng, A.; Song, Z.; Zheng, C.; Hoodgmoed, W.; Zhang, W. Warming impacts on winter wheat phenophase and grain yield under field conditions in Yangtze Delta Plain, China. Field Crops Res. 2012, 134, 193–199. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Long, S.P. 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Glob. Change Biol. 2020, 27, 27–49. [Google Scholar] [CrossRef]
- Kobayasi, K.; Sakai, H.; Tokida, T.; Nakamura, H.; Usui, Y.; Yoshimoto, M.; Hasegawa, T. Effects of free-air CO2 enrichment on flower opening time in rice. Plant Prod. Sci. 2019, 22, 367–373. [Google Scholar] [CrossRef] [Green Version]
- McLeod, A.R.; Long, S.P. Free-air carbon dioxide enrichment (FACE) in global change research: A review. Adv. Ecol. Res. 1999, 28, 1–56. [Google Scholar] [CrossRef]
- Usui, Y.; Sakai, H.; Tokida, T.; Nakamura, H.; Nakagawa, H.; Hasegawa, T. Rice grain yield and quality responses to free-air CO2 enrichment combined with soil and water warming. Glob. Change Biol. 2016, 22, 1256–1270. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Chen, F.; Lin, X.; Liu, Z.; Zhang, H.; Zhao, J.; Li, K.; Ye, Q.; Li, Y.; Lv, S.; et al. Potential benefits of climate change for crop productivity in China. Agric. For. Meteorol. 2015, 208, 76–84. [Google Scholar] [CrossRef]
- Long, S.P.; Ort, D.R. More than taking the heat: Crops and global change. Curr. Opin Plant. Biol. 2010, 13, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Yamori, W.; Hikosaka, K.; Way, D.A. Temperature response of photosynthesis in C 3, C 4, and CAM plants: Temperature acclimation and temperature adaptation. Photosynth. Res. 2014, 119, 101–117. [Google Scholar] [CrossRef]
- Wu, C.; Tang, S.; Li, G.; Wang, S.; Fahad, S.; Ding, Y. Response of phytohormone homeostasis to heat stress and the roles of phytohormones in rice grain yield: A review. PeerJ Prepr. 2019, 7, e27843v1. [Google Scholar] [CrossRef]
- Ryu, J.H.; Jeong, H.; Cho, J. Performances of Vegetation Indices on Paddy Rice at Elevated Air Temperature, Heat Stress, and Herbicide Damage. Remote Sens. 2020, 12, 2654. [Google Scholar] [CrossRef]
- Matsui, T.; Namuco, O.S.; Ziska, L.H.; Horie, T. Effects of high temperature and CO2 concentration on spikelet sterility in indica rice. Field Crops Res. 1997, 51, 213–219. [Google Scholar] [CrossRef]
- Matsui, T.; Omasa, K.; Horie, T. The Difference in sterility due to high temperatures during the flowering period among japonica-rice varieties. Plant Prod. Sci. 2001, 4, 90–93. [Google Scholar] [CrossRef]
- Maruyama, A.; Weerakoon, W.M.W.; Wakiyama, Y.; Ohba, K. Effects of increasing temperatures on spikelet fertility in different rice cultivars based on temperature gradient chamber experiments. J. Agron. Crop. Sci. 2013, 199, 416–423. [Google Scholar] [CrossRef]
- Lai, Y.J.; Li, C.F.; Lin, P.H.; Wey, T.H.; Chang, C.S. Comparison of MODIS land surface temperature and ground-based observed air temperature in complex topography. Int. J. Remote Sens. 2012, 33, 7685–7702. [Google Scholar] [CrossRef]
- Yan, H.; Zhang, J.; Hou, Y.; He, Y. Estimation of air temperature from MODIS data in east China. Int. J. Remote Sen. 2009, 30, 6261–6275. [Google Scholar] [CrossRef]
- Ryu, J.H.; Han, K.S.; Cho, J.; Lee, C.S.; Yoon, H.J.; Yeom, J.M.; Ou, M.L. Estimating midday near-surface air temperature by weighted consideration of surface and atmospheric moisture conditions using COMS and SPOT satellite data. Int. J. Remote Sens. 2015, 36, 3503–3518. [Google Scholar] [CrossRef]
- Lu, J.; Miao, Y.; Huang, Y.; Shi, W.; Hu, X.; Wang, X.; Wan, J. Evaluating an unmanned aerial vehicle-based remote sensing system for estimation of rice nitrogen status. In Proceedings of the 2015 Fourth International Conference on Agro-Geoinformatics (Agro-Geoinformatics), Istanbul, Turkey, 20–24 July 2015; IEEE: Piscataway, NJ, USA, 2015; pp. 198–203. [Google Scholar] [CrossRef]
- Satir, O.; Berberoglu, S. Crop yield prediction under soil salinity using satellite derived vegetation indices. Field Crops Res. 2016, 192, 134–143. [Google Scholar] [CrossRef]
- Gao, B.C. NDWI—A normalized difference water index for remote sensing of vegetation liquid water from space. Remote Sens. Environ. 1996, 58, 257–266. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Zur, Y.; Chivkunova, O.B.; Merzlyak, M.N. Assessing carotenoid content in plant leaves with reflectance spectroscopy. Photochem. Photobiol. 2002, 75, 272–281. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Pattey, E.; Zarco-Tejada, P.J.; Strachan, I.B. Hyperspectral vegetation indices and novel algorithms for predicting green LAI of crop canopies: Modeling and validation in the context of precision agriculture. Remote Sens. Environ. 2004, 90, 337–352. [Google Scholar] [CrossRef]
- Kim, M.S.; Daughtry, C.S.T.; Chappelle, E.W.; McMurtrey, J.E.; Walthall, C.L. The use of high spectral resolution bands for estimating absorbed photosynthetically active radiation (Apar). In Proceedings of the 6th Symposium on Physical Measurements and Signatures in Remote Sensing, Val D’Isere, France, 17–21 January 1994. [Google Scholar]
- Verhulst, N.; Govaerts, B.; Nelissen, V.; Sayre, K.D.; Crossa, J.; Raes, D.; Deckers, J. The effect of tillage, crop rotation and residue management on maize and wheat growth and development evaluated with an optical sensor. Field Crops Res. 2011, 120, 58–67. [Google Scholar] [CrossRef]
- Hmimina, G.; Dufrêne, E.; Pontailler, J.Y.; Delpierre, N.; Aubinet, M.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; Granier, A.; et al. Evaluation of the potential of MODIS satellite data to predict vegetation phenology in different biomes: An investigation using ground-based NDVI measurements. Remote Sens. Environ. 2013, 132, 145–158. [Google Scholar] [CrossRef]
- Wu, C.; Peng, D.; Soudani, K.; Siebicke, L.; Gough, C.M.; Arain, M.A.; Bohrer, G.; Lafleur, P.M.; Peichl, M.; Gonsamo, A.; et al. Land surface phenology derived from normalized difference vegetation index (NDVI) at global FLUXNET sites. Agric. For. Meteorol. 2017, 233, 171–182. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Ho, C.-H.; Gim, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Change Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- He, M.; Kimball, J.S.; Running, S.; Ballantyne, A.; Guan, K.; Huemmrich, F. Satellite detection of soil moisture related water stress impacts on ecosystem productivity using the MODIS-based photochemical reflectance index. Remote Sens. Environ. 2016, 186, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Suárez, L.; Zarco-Tejada, P.J.; Sepulcre-Cantó, G.; Pérez-Priego, O.; Miller, J.R.; Jiménez-Muñoz, J.C.; Sobrino, J. Assessing canopy PRI for water stress detection with diurnal airborne imagery. Remote Sens. Environ. 2008, 112, 560–575. [Google Scholar] [CrossRef]
- Cao, Z.; Yao, X.; Liu, H.; Liu, B.; Cheng, T.; Tian, Y.; Cao, W.; Zhu, Y. Comparison of the abilities of vegetation indices and photosynthetic parameters to detect heat stress in wheat. Agric. For. Meteorol. 2019, 265, 121–136. [Google Scholar] [CrossRef]
- Park, E.; Kim, Y.-S.; Omari, M.K.; Suh, H.-K.; Faqeerzada, M.A.; Kim, M.S.; Baek, I.; Cho, B.-K. High-Throughput Phenotyping Approach for the Evaluation of Heat Stress in Korean Ginseng (Panax ginseng Meyer) Using a Hyperspectral Reflectance Image. Sensors 2021, 21, 5634. [Google Scholar] [CrossRef]
- Xie, X.J.; Zhang, Y.H.; Li, R.Y.; Shen, S.H.; Bao, Y.X. Prediction model of rice crude protein content, amylose content and actual yield under high temperature stress based on hyper-spectral remote sensing. Qual. Assur. Saf. Crop. 2019, 11, 517–527. [Google Scholar] [CrossRef]
- Ryu, J.-H.; Na, S.-I.; Cho, J. Inter-Comparison of normalized difference vegetation index measured from different footprint sizes in cropland. Remote Sens. 2020, 12, 2980. [Google Scholar] [CrossRef]
- Liu, L.; Hoogenboom, G. Ingram, K.T. Controlled-environment sunlit plant growth chambers. Crit. Rev. Plant. Sci. 2000, 19, 347–375. [Google Scholar] [CrossRef]
- Okada, M.; Hamasaki, T.; Hayashi, T. Temperature gradient chambers for research on global environment change. I. Thermal environment in a large chamber. Biotronics 1995, 24, 85–97. [Google Scholar] [CrossRef]
- Hadley, P.; Batts, G.R.; Ellis, R.H.; Morison, J.I.L.; Pearson, S.; Wheeler, T.R. Temperature gradient chambers for research on global environment change. II. A twin-wall tunnel system for low-stature, field-grown crops using a split heat pump. Plant Cell Environ. 1995, 18, 1055–1063. [Google Scholar] [CrossRef]
- Tang, R.S.; Zheng, J.C.; Jin, Z.Q.; Zhang, D.D.; Huang, Y.H.; Chen, L.G. Possible correlation between high temperature-induced floret sterility and endogenous levels of IAA, GAs and ABA in rice (Oryza sativa L.). Plant Growth Regul. 2008, 54, 37–43. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.; Schell, J.; Deering, D. Monitoring vegetation systems in the Great Plains with ERTS-1. In Proceedings of the Third Earth Resources Technology Satellite Symposium, Washington, DC, USA, 10–14 December 1974. [Google Scholar]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Yeom, J.M.; Ko, J.; Hwang, J.; Lee, C.S.; Choi, C.U.; Jeong, S. Updating absolute radiometric characteristics for KOMPSAT-3 and KOMPSAT-3A multispectral imaging sensors using well-characterized pseudo-invariant tarps and microtops II. Remote Sens. 2018, 10, 697. [Google Scholar] [CrossRef] [Green Version]
- Mosleh, M.; Hassan, Q.; Chowdhury, E. Application of remote sensors in mapping rice area and forecasting its production: A review. Sensors 2015, 15, 769–791. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Tao, F.; Zhang, Z. Changes in extreme temperatures and their impacts on rice yields in southern China from 1981 to 2009. Field Crops Res. 2016, 189, 43–50. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.G.; Cho, H.S.; Choi, S.H.; Lee, J.M.; Pyon, J.Y. Varietal Difference in Heading Date of Rice by Night Illumination. Korean J. Crop. Sci. 2003, 38, 209–215, (In Korean with English Abstract). [Google Scholar]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V.R. High-temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, J.; Song, G.; Long, Z.; Chen, C. Impacts of recent temperatures rise on double-rice phenology across Southern China. Int. J. Plant Prod. 2019, 13, 1–10. [Google Scholar] [CrossRef]
- Tanaka, A.; Toriyama, K.; Kobayashi, K. Less yield reduction induced by high temperature in a paddy field under organic fertilizer management in Tochigi prefecture. J. Agric. Meteorol. 2011, 67, 249–258. [Google Scholar] [CrossRef]
- Ekanayake, I.J.; Datta, S.D.; Steponkus, P.L. Spikelet sterility and flowering response of rice to water stress at anthesis. Ann. Bot. 1989, 63, 257–264. [Google Scholar] [CrossRef]
- Rang, Z.W.; Jagadish, S.V.K.; Zhou, Q.M.; Craufurd, P.Q.; Heuer, S. Effect of high temperature and water stress on pollen germination and spikelet fertility in rice. Environ. Exp. Bot. 2011, 70, 58–65. [Google Scholar] [CrossRef] [Green Version]
- Gerhards, M.; Rock, G.; Schlerf, M.; Udelhoven, T. Water stress detection in potato plants using leaf temperature, emissivity, and reflectance. Int. J. Appl. Earth Obs. Geoinf. 2016, 53, 27–39. [Google Scholar] [CrossRef]
- Sims, D.A.; Gamon, J.A. Relationships between leaf pigment content and spectral reflectance across a wide range of species, leaf structures and developmental stages. Remote Sens. Environ. 2002, 81, 337–354. [Google Scholar] [CrossRef]
- Shon, J.; Kim, J.; Lee, C.; Yang, W. Effect of high temperature on leaf physiological changes as chlorophyll composition and photosynthesis rate of rice. Korean J. Crop. Sci. 2015, 60, 266–272. [Google Scholar] [CrossRef]
- He, J.; Zhang, N.; Su, X.; Lu, J.; Yao, X.; Cheng, T.; Zhu, Y.; Cao, W.; Tian, Y. Estimating leaf area index with a new vegetation index considering the influence of rice panicles. Remote Sens. 2019, 11, 1809. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.; Merzlyak, M.N. Spectral reflectance changes associated with autumn senescence of Aesculus hippocastanum L. and Acer platanoides L. leaves. Spectral features and relation to chlorophyll estimation. J. Plant. Physiol. 1994, 143, 286–292. [Google Scholar] [CrossRef]
- Li, Z.W.; Xiong, J.; Li, Z.F.; Qi, X.H.; Chen, H.F.; Shao, C.H.; Wang, J.Y.; Liang, Y.Y.; Lin, W.X. Analysis of differential expression of proteins in rice leaf sheath during grain filling. Acta Agron. Sin. 2008, 34, 619–626. [Google Scholar] [CrossRef]
- Chen, H.-J.; Wang, S.-J. Molecular regulation of sink–source transition in rice leaf sheaths during the heading period. Acta Physiol. Plant. 2008, 30, 639–649. [Google Scholar] [CrossRef]
- Rahman, M.A.; Haque, M.E.; Sikdar, B.; Islam, M.A.; Matin, M.N. Correlation analysis of flag leaf with yield in several rice cultivars. J. Life Earth Sci. 2013, 8, 49–54. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Optical Devices | Years | Number of Observation Positions |
|---|---|---|
| Leaf-spectrometer | 2018 | 2 |
| Field-spectrometer | 2016, 2017, 2018, 2019 | 10, 12, 16, 6 |
| Multi-spectral camera | 2018 | 4 |
| Input Variables | R-Square | RMSE (%) | Equation |
|---|---|---|---|
| Air temperature | 0.961 | 7.180 | Y = 88.933/(1 + exp(−(X − 27.244)/−0.330)) − 1.225 if (Y > 0, Y, 0) |
| (1−rsPRI)/(1−rNDVI) | 0.922 | 10.162 | Y = 83.414/(1 + exp(−(X − 3.809)/−0.636)) + 8.066, if (rNDVI < 0.99, rNDVI, 0.99) |
| rNDVI | 0.954 | 7.820 | Y = 97.317/(1 + exp(−(X − 0.900)/−0.044)) − 7.462 if (Y > 0, Y, 0) |
| rsPRI | 0.943 | 8.640 | Y = 91.386/(1 + exp(−(X − 0.608)/−0.043)) − 1.515, if (Y > 0, Y, 0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, J.-H.; Oh, D.; Ko, J.; Kim, H.-Y.; Yeom, J.-M.; Cho, J. Remote Sensing-Based Evaluation of Heat Stress Damage on Paddy Rice Using NDVI and PRI Measured at Leaf and Canopy Scales. Agronomy 2022, 12, 1972. https://doi.org/10.3390/agronomy12081972
Ryu J-H, Oh D, Ko J, Kim H-Y, Yeom J-M, Cho J. Remote Sensing-Based Evaluation of Heat Stress Damage on Paddy Rice Using NDVI and PRI Measured at Leaf and Canopy Scales. Agronomy. 2022; 12(8):1972. https://doi.org/10.3390/agronomy12081972
Chicago/Turabian StyleRyu, Jae-Hyun, Dohyeok Oh, Jonghan Ko, Han-Yong Kim, Jong-Min Yeom, and Jaeil Cho. 2022. "Remote Sensing-Based Evaluation of Heat Stress Damage on Paddy Rice Using NDVI and PRI Measured at Leaf and Canopy Scales" Agronomy 12, no. 8: 1972. https://doi.org/10.3390/agronomy12081972
APA StyleRyu, J.-H., Oh, D., Ko, J., Kim, H.-Y., Yeom, J.-M., & Cho, J. (2022). Remote Sensing-Based Evaluation of Heat Stress Damage on Paddy Rice Using NDVI and PRI Measured at Leaf and Canopy Scales. Agronomy, 12(8), 1972. https://doi.org/10.3390/agronomy12081972