
What Factors Control the Crude Protein Content Variation of a Basaltic “Campos” Native Grassland of South America?


 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
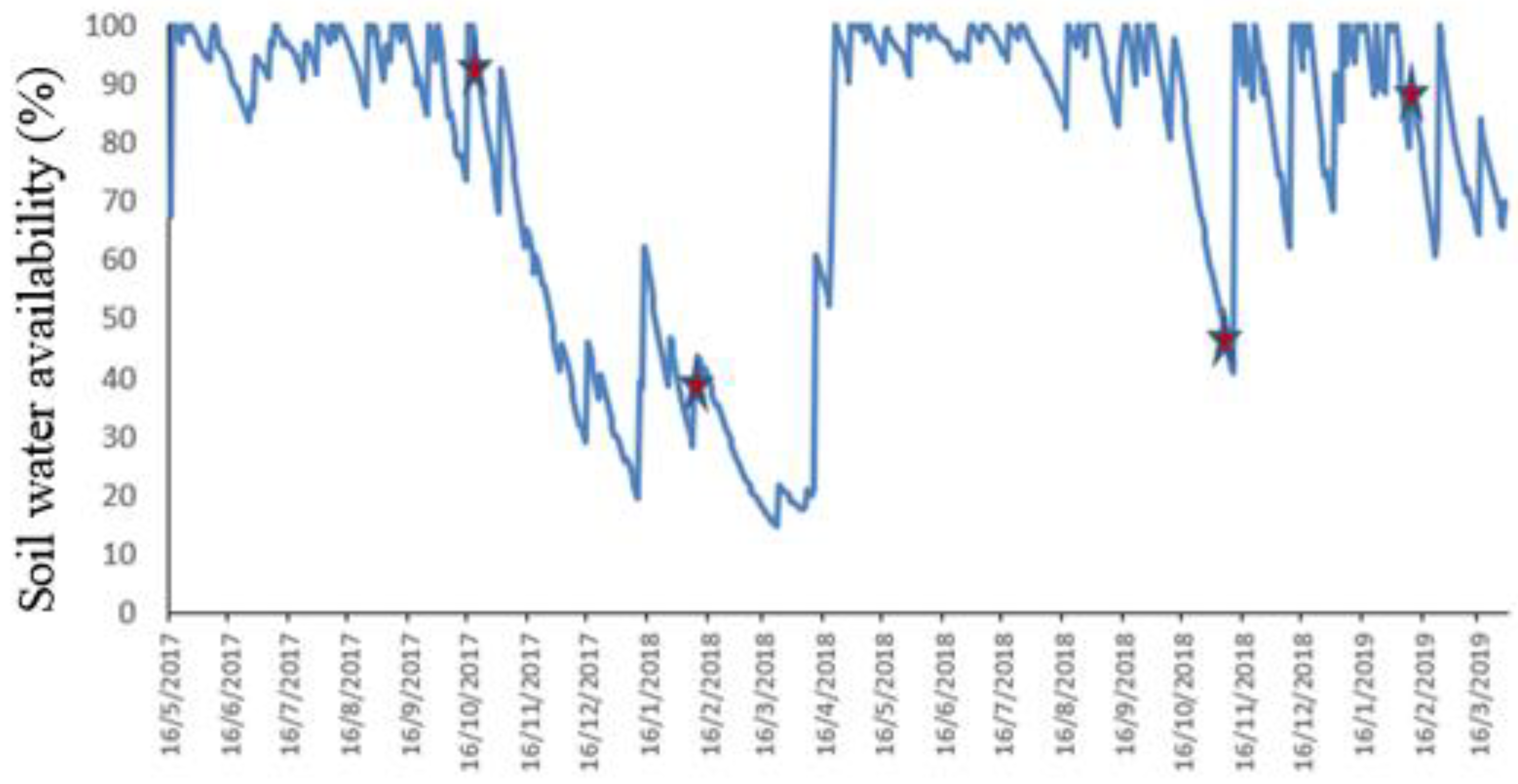
2.1. Site Description and Experimental Design
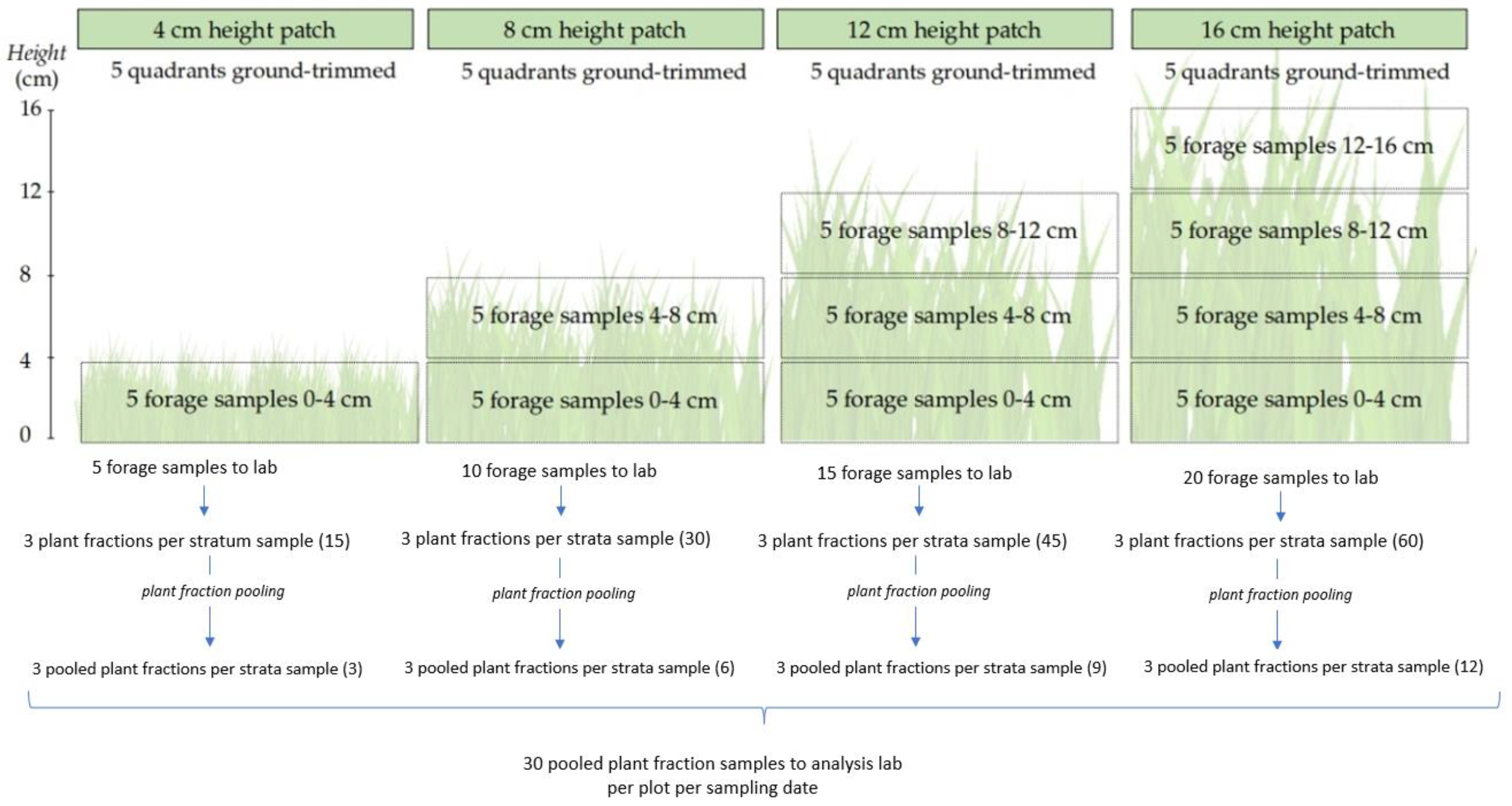
2.2. Grasslands Measurements and Forage Sampling
2.3. Forage Crude Protein Analysis
2.4. Data Analysis
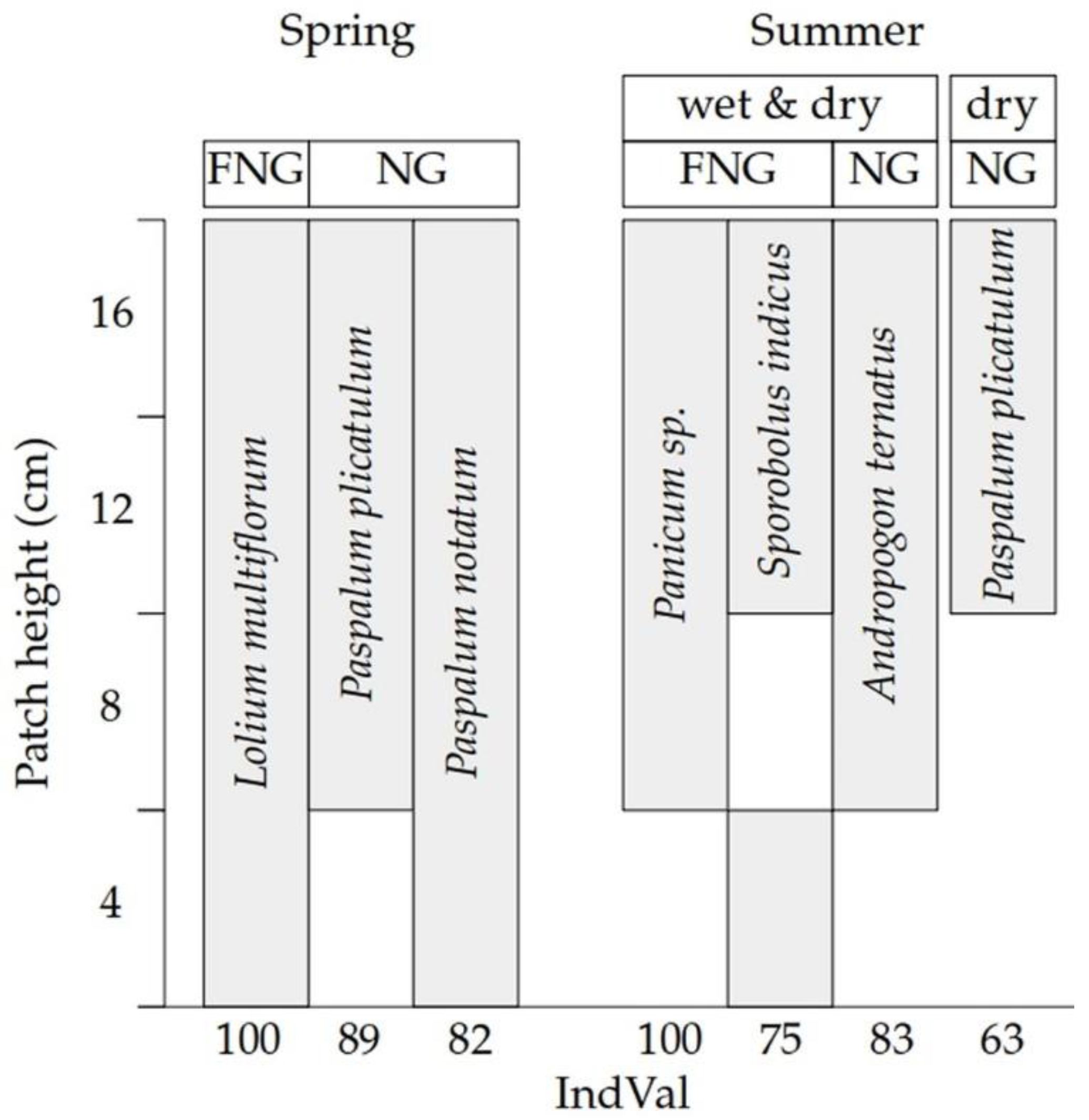
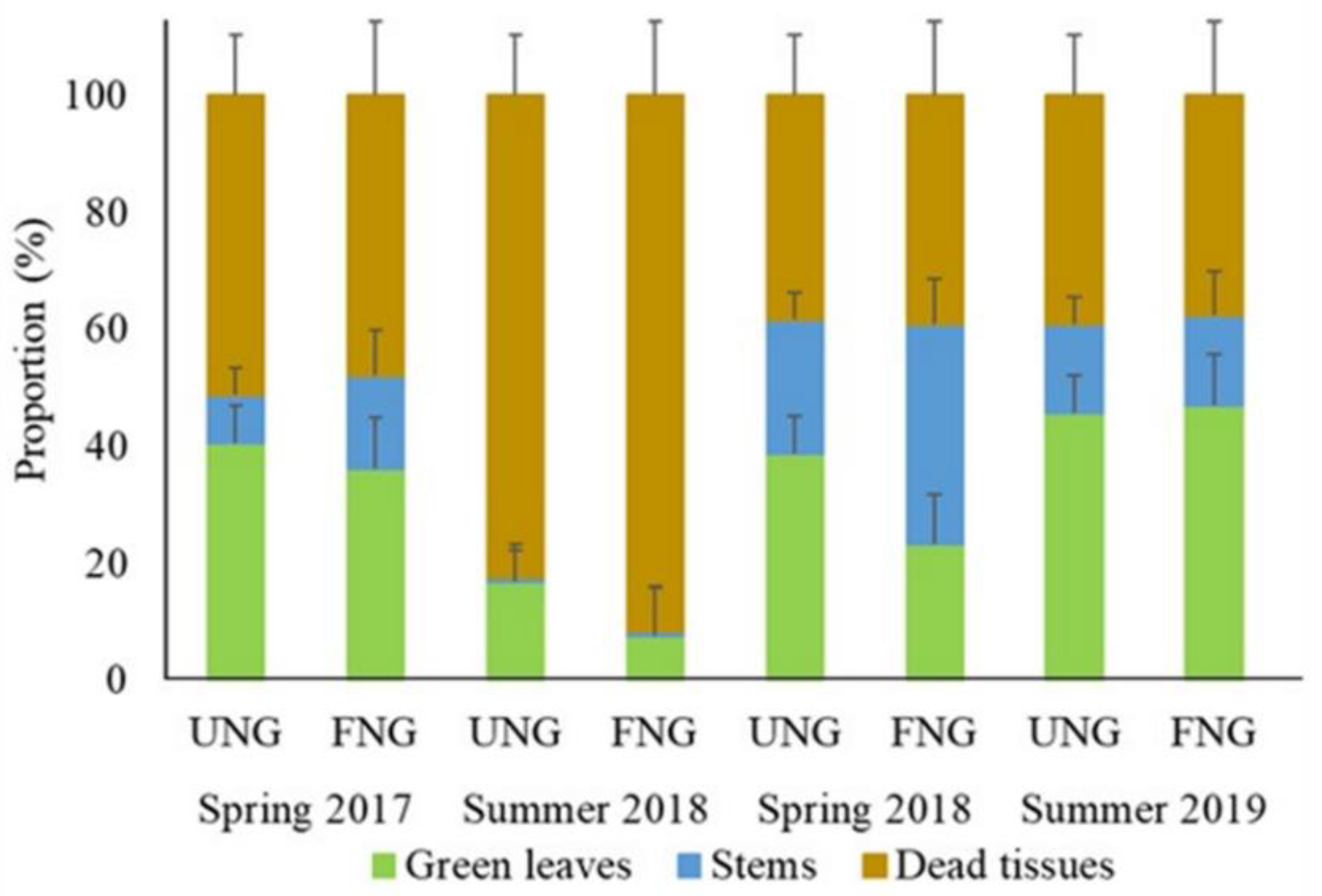
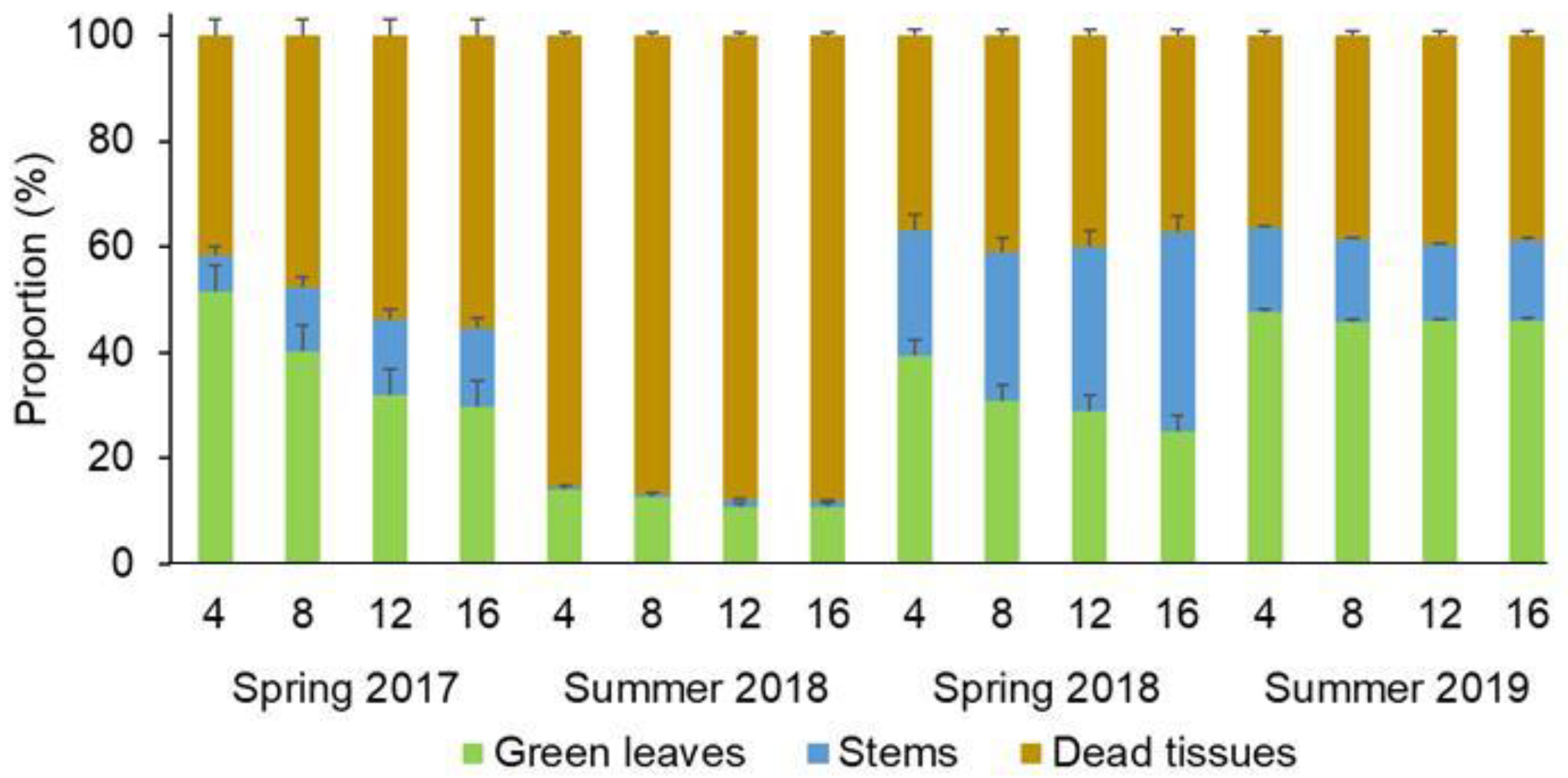
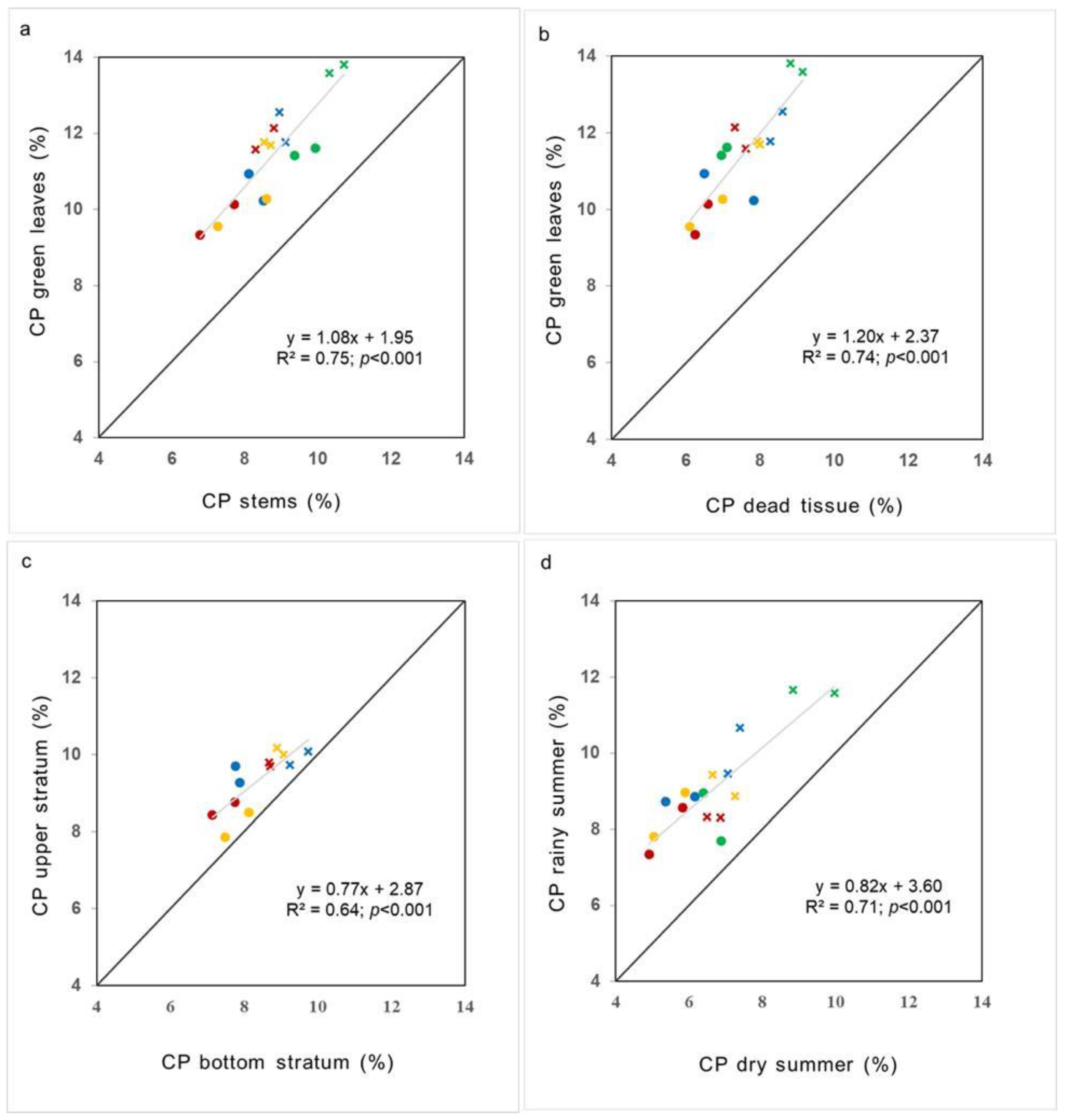
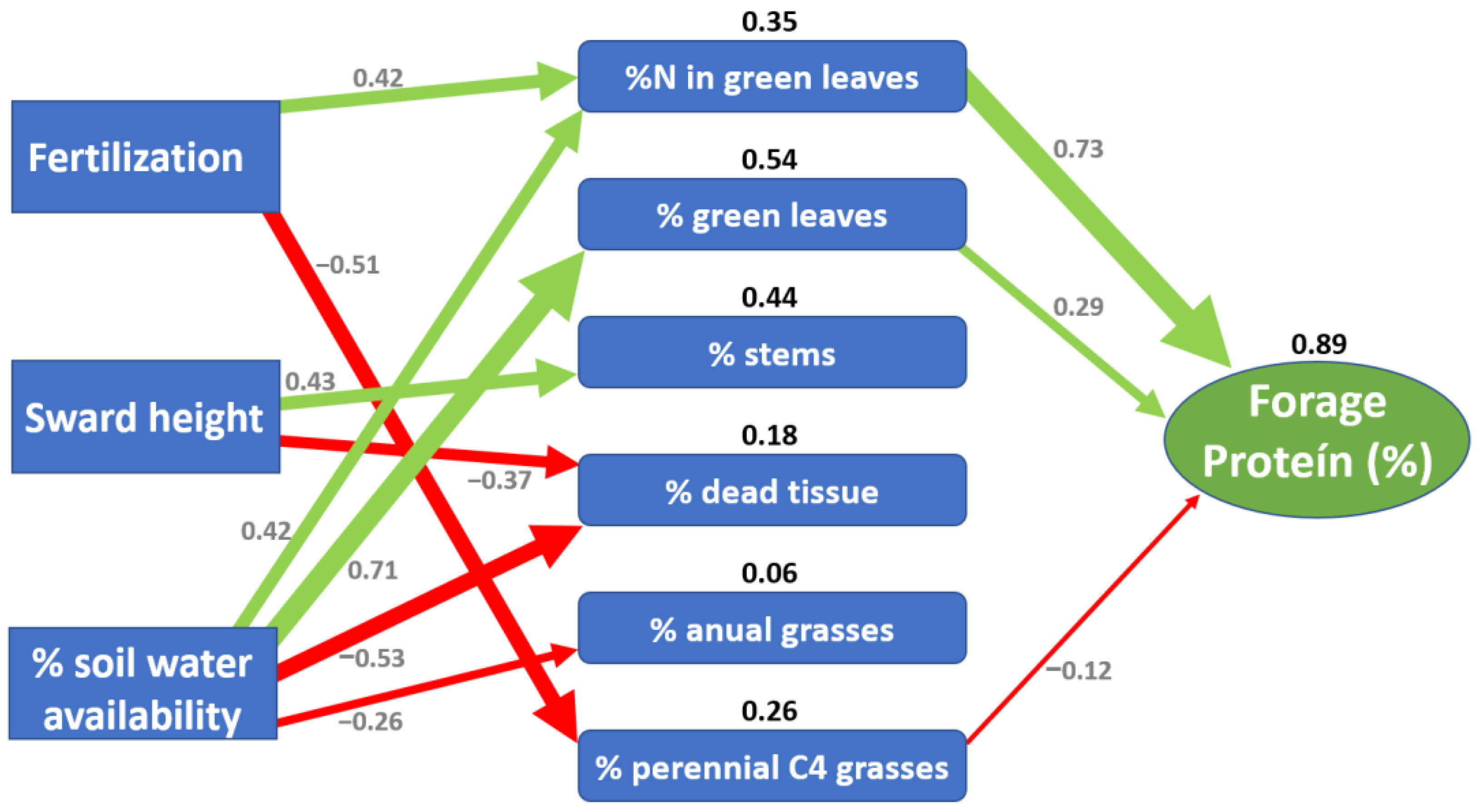
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

Appendix B
References
- Jaurena, M.; Durante, M.; Devincenzi, T.; Savian, J.V.; Bendersky, D.; Moojen, F.G.; Pereira, M.; Soca, P.; Quadros, F.L.F.; Pizzio, R.; et al. Native Grasslands at the Core: A New Paradigm of Intensification for the Campos of Southern South America to Increase Economic and Environmental Sustainability. Front. Sustain. Food. Syst. 2021, 1–11. [Google Scholar] [CrossRef]
- Carvalho, P.C.; Batello, C. Access to Land, Livestock Production and Ecosystem Conservation in the Brazilian Campos Biome: The Natural Grasslands Dilemma. Livest. Sci. 2009, 120, 158–162. [Google Scholar] [CrossRef]
- Viglizzo, E.F.; Frank, F.C. Land-Use Options for Del Plata Basin in South America: Tradeoffs Analysis Based on Ecosystem Service Provision. Ecol. Econ. 2006, 57, 140–151. [Google Scholar] [CrossRef]
- Weyland, F.; Barral, M.P.; Laterra, P. Assessing the Relationship between Ecosystem Functions and Services: Importance of Local Ecological Conditions. Ecol. Indic. 2017, 81, 201–213. [Google Scholar] [CrossRef]
- Da Trindade, J.K.; Neves, F.P.; Pinto, C.E.; Bremm, C.; Mezzalira, J.C.; Nadin, L.B.; Genro, T.C.; Gonda, H.L.; Carvalho, P.C. Daily Forage Intake by Cattle on Natural Grassland: Response to Forage Allowance and Sward Structure. Rangel. Ecol. Manag. 2016, 69, 59–67. [Google Scholar] [CrossRef]
- Moore, J.E.; Brant, M.H.; Kunkle, W.E.; Hopkins, D.I. Forage Supplementation and Grazing-Effects of Supplementation on Voluntary Forage Intake. J. Anim. Sci. 1999, 77 (Suppl. 2), 122–135. [Google Scholar] [CrossRef]
- Poppi, D.P.; Quigley, S.P.; da Silva, T.A.C.C.; McLennan, S.R. Challenges of Beef Cattle Production from Tropical Pastures. Rev. Bras. Zootec. 2018, 47, 1–9. [Google Scholar] [CrossRef]
- Ramos, Z.; De Barbieri, I.; van Lier, E.; Montossi, F. Body and Wool Growth of Lambs Grazing on Native Pastures Can Be Improved with Energy and Protein Supplementation. Small Rumin. Res. 2019, 171, 92–98. [Google Scholar] [CrossRef]
- Soussana, J.-F.; Lüscher, A. Temperate Grasslands and Global Atmospheric Change: A Review. Grass Forage Sci. 2007, 62, 127–134. [Google Scholar] [CrossRef]
- Chaneton, E.J. Factores que determinan la heterogeneidad de la comunidad vegetal en diferentes escalas espaciales. In La Heterogeneidad de la Vegetación de los Agroecosistemas; Oesterheld, M., Aguiar, M.R., Ghersa, C.M., Paruelo, J.M., Eds.; Facultad de Agronomía, Universidad de Buenos Aires: Buenos Aires, Argentina, 2005; pp. 19–42. [Google Scholar]
- Tonn, B.; Densing, E.M.; Gabler, J.; Isselstein, J. Grazing-Induced Patchiness, Not Grazing Intensity, Drives Plant Diversity in European Low-Input Pastures. J. Appl. Ecol. 2019, 56, 1624–1636. [Google Scholar] [CrossRef]
- Cruz, P.; Lezana, L.; Durante, M.; Jaurena, M.; Figari, M.; Bittencourt, L.; Theau, J.-P.; Massa, E.; Viegas, J.; Ferreira de Quadros, F.L. Una Clasificación Funcional de 63 Poáceas Comunes de Los Pastizales Naturales de Sudamérica. Ecol. Austral. 2019, 29, 239–248. [Google Scholar] [CrossRef]
- Jaurena, M.; Lezama, F.; Cruz, P. Perennial Grasses Traits as Functional Markers of Grazing Intensity in Basaltic Grasslands of Uruguay. Chil. J. Agric. Res. 2012, 72, 541–549. [Google Scholar] [CrossRef]
- Lezama, F.; Altesor, A.; León, R.J.; Paruelo, J.M. Heterogeneidad de La Vegetación En Pastizales Naturales de La Región Basáltica de Uruguay. Ecol. Austral. 2006, 16, 167–182. [Google Scholar]
- Wilson, J.R.; Minson, D.J. Prospects for Improving the Digestibility and Intake of Tropical Grasses. Trop. Grassl. 1980, 14, 253–259. [Google Scholar]
- Herrera, L.P.; Laterra, P. Relative Influence of Size, Connectivity and Disturbance History on Plant Species Richness and Assemblages in Fragmented Grasslands. Appl. Veg. Sci. 2011, 14, 181–188. [Google Scholar] [CrossRef]
- Azambuja Filho, J.C.R.; de Faccio Carvalho, P.C.; Bonnet, O.J.F.; Bastianelli, D.; Jouven, M. Functional Classification of Feed Items in Pampa Grassland, Based on Their Near-Infrared Spectrum. Rangel. Ecol. Manag. 2020, 73, 358–367. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought Effect on Plant Nitrogen and Phosphorus: A Meta-Analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef]
- Dumont, B.; Andueza, D.; Niderkorn, V.; Lüscher, A.; Porqueddu, C.; Picon-Cochard, C. A Meta-Analysis of Climate Change Effects on Forage Quality in Grasslands: Specificities of Mountain and Mediterranean Areas. Grass. Forage Sci. 2015, 70, 239–254. [Google Scholar] [CrossRef]
- Marriott, C.A.; Carrère, P. Structure and Dynamics of Grazed Vegetation. Ann. Zootech. 1998, 47, 359–369. [Google Scholar] [CrossRef]
- Dos Santos, A.B.; de Quadros, F.L.F.; Rossi, G.E.; de Pereira, L.P.; Kuinchtner, B.C.; de Carvalho, R.M.R. Valor Nutritivo de Gramíneas Nativas Do Rio Grande Do Sul/Brasil, Classificadas segundo uma tipologia funcional, sob queima e pastejo. Cienc. Rural. 2013, 43, 342–347. [Google Scholar] [CrossRef][Green Version]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Colin Prentice, I.; et al. The Global Spectrum of Plant Form and Function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, G.; Belanger, G. Allometries in Plants as Drivers of Forage Nutritive Value: A Review. Agriculture 2019, 10, 5. [Google Scholar] [CrossRef]
- Rodríguez Palma, R.; Rodríguez, T. Campo Natural de Basalto: Cuánto Responde En Producción de Forraje? In Proceedings of the XXIV Reunión del Grupo Técnico en Forrajeras del Cono Sur, Tacuarembó, Uruguay, 13–14 July 2017; Ayala, W., Boggiano, P., Alvarez, O., Eds.; Instituto Nacional de Investigación Agropecuaria: Tacuarembó, Uruguay, 2017; pp. 57–59. [Google Scholar]
- Ávila, M.R.D.; Nabinger, C.; Brambilla, D.M.; Carassai, I.J.; Kunrath, T.R. The Effects of Nitrogen Enrichment on Tiller Population Density and Demographics of Annual Ryegrass Overseeded on Natural Pastures South of Brazil. Afr. J. Agric. Res. 2013, 8, 3013–3018. [Google Scholar] [CrossRef]
- Mott, G.O.; Lucas, H.L. The Design, Conduct and Interpretation of Grazing Trials on Cultivated and Improved Pastures. In Proceedings of the International Grassland Congress, State College, PA, USA, 17–23 August 1952; pp. 1380–1395. [Google Scholar]
- Ellenberg, D.; Mueller-Dombois, D. Aims and Methods of Vegetation Ecology; Wiley: New York, NY, USA, 1974. [Google Scholar]
- Hodgson, J. Grazing Management: Science into Practice; Longman Handbooks in Agriculture; Longman Scientific and Technical and John Wiley: New York, NY, USA, 1990; 203p. [Google Scholar]
- AOAC International. AOAC Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2000. [Google Scholar]
- Di Rienzo, J.; Casanoves, F.; Balzarini, M.; Gonzalez, L.; Tablada, M.; Robledo, C. InfoStat Statistical software2011. Available online: https://www.infostat.com.ar/index.php (accessed on 30 May 2022).
- Dufrêne, M.; Legendre, P. Species Assemblages and Indicator Species: The Need for a Flexible Asymmetrical Approach. Ecol. Monogr. 1997, 67, 345–366. [Google Scholar] [CrossRef]
- Hair, J.F.; Ringle, C.M.; Sarstedt, M. PLS-SEM: Indeed a Silver Bullet. J. Mark. Theory Pract. 2011, 19, 139–152. [Google Scholar] [CrossRef]
- Zanoniani, R.; Boggiano, P.; Cadenazzi, M. Respuesta Invernal de Un Campo Natural a Fertilización Nitrogenada y Ofertas de Forraje. Agrocienc. Urug. 2011, 15, 115–124. [Google Scholar] [CrossRef]
- Jaurena, M.; Lezama, F.; Salvo, L.; Cardozo, G.; Ayala, W.; Terra, J.; Nabinger, C. The dilemma of improving native grasslands by overseeding legumes: Production intensification or diversity conservation. Rangel. Ecol. Manag. 2016, 69, 35–42. [Google Scholar] [CrossRef]
- Berendse, F.; Elberse, W.T.; Geerts, R.H.M.E. Competition and Nitrogen Loss from Plants in Grassland Ecosystems. Ecology 1992, 73, 46–53. [Google Scholar] [CrossRef]
- Gastal, F.; Lemaire, G. N Uptake and Distribution in Crops: An Agronomical and Ecophysiological Perspective. J. Exp. Bot. 2002, 53, 789–799. [Google Scholar] [CrossRef]
- Madeira, W. Efectos de La Fertilización Primavero-Estival Nitrogeno Fosfatada y Del Riego Suplementario En La Productividad y Eficiencia de Uso de Nutrientes Del Campo Natural. Master’s Thesis, UdelaR, Montevideo, Uruguay, 2019. [Google Scholar]
- Duru, M.; Lemaire, G.; Cruz, P. Grasslands. In Diagnosis of the Nitrogen Status in Crops; Lemaire, G., Ed.; Crops; Lemaire; Springer: Berlin/Heidelberg, Germany, 1997; p. 239. ISBN 978-3-642-64506-8. [Google Scholar]
- Hejcman, M.; Szaková, J.; Schellberg, J.; Tlustoš, P. The Rengen Grassland Experiment: Relationship between Soil and Biomass Chemical Properties, Amount of Elements Applied, and Their Uptake. Plant Soil 2010, 333, 163–179. [Google Scholar] [CrossRef]
- Lemaire, G.; Ciampitti, I. Nitrogen Use Efficiency across Genotype-by-Environment-by-Management Scenarios: A Review. Plants 2020, 9, 1309. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Briggs, J.M.; Koelliker, J.K. Frequency and Extent of Water Limitation to Primary Production in a Mesic Temperate Grassland. Ecosystems 2001, 4, 19–28. [Google Scholar] [CrossRef]
- Deng, L.; Peng, C.; Kim, D.-G.; Li, J.; Liu, Y.; Hai, X.; Liu, Q.; Huang, C.; Shangguan, Z.; Kuzyakov, Y. Drought Effects on Soil Carbon and Nitrogen Dynamics in Global Natural Ecosystems. Earth-Sci. Rev. 2021, 214, 103501. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Núñez, L.; Hirigoyen, A.; Durante, M.; Arroyo, J.M.; Cazzuli, F.; Bremm, C.; Jaurena, M. What Factors Control the Crude Protein Content Variation of a Basaltic “Campos” Native Grassland of South America? Agronomy 2022, 12, 1756. https://doi.org/10.3390/agronomy12081756
Núñez L, Hirigoyen A, Durante M, Arroyo JM, Cazzuli F, Bremm C, Jaurena M. What Factors Control the Crude Protein Content Variation of a Basaltic “Campos” Native Grassland of South America? Agronomy. 2022; 12(8):1756. https://doi.org/10.3390/agronomy12081756
Chicago/Turabian StyleNúñez, Laura, Andrés Hirigoyen, Martín Durante, José María Arroyo, Fiorella Cazzuli, Carolina Bremm, and Martín Jaurena. 2022. "What Factors Control the Crude Protein Content Variation of a Basaltic “Campos” Native Grassland of South America?" Agronomy 12, no. 8: 1756. https://doi.org/10.3390/agronomy12081756
APA StyleNúñez, L., Hirigoyen, A., Durante, M., Arroyo, J. M., Cazzuli, F., Bremm, C., & Jaurena, M. (2022). What Factors Control the Crude Protein Content Variation of a Basaltic “Campos” Native Grassland of South America? Agronomy, 12(8), 1756. https://doi.org/10.3390/agronomy12081756